
Anorexigenic Effects of Intermittent Hypoxia on the Gut—Brain Axis in Sleep Apnea Syndrome

 , and
, and 
{kind=link}
{kind=link}
Abstract
1. Introduction
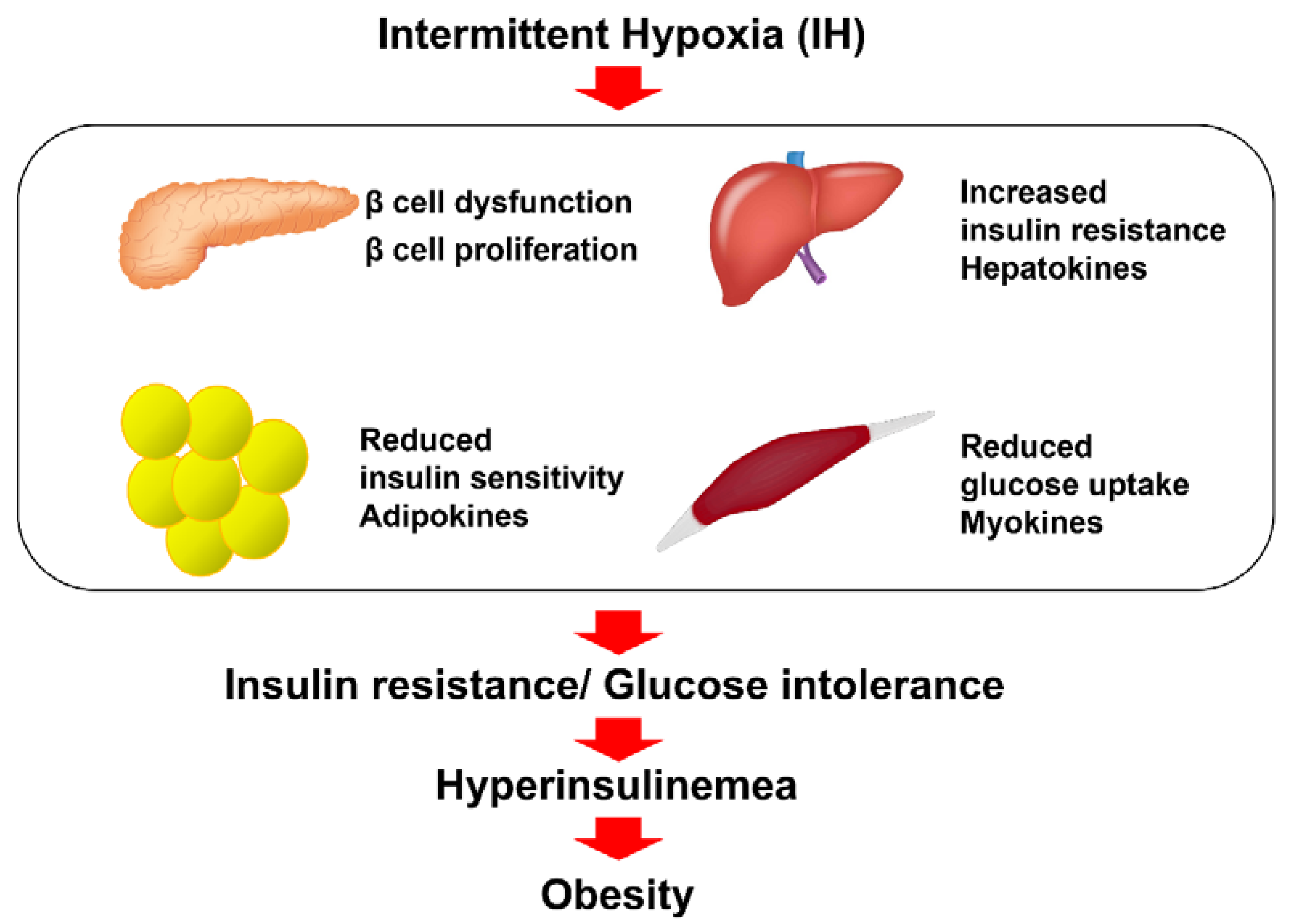
2. The Reciprocal Relationship between IH and Obesity
3. Molecular Mechanisms of the IH-Induced Gene Expression
4. IH and Impaired Glucose Tolerance
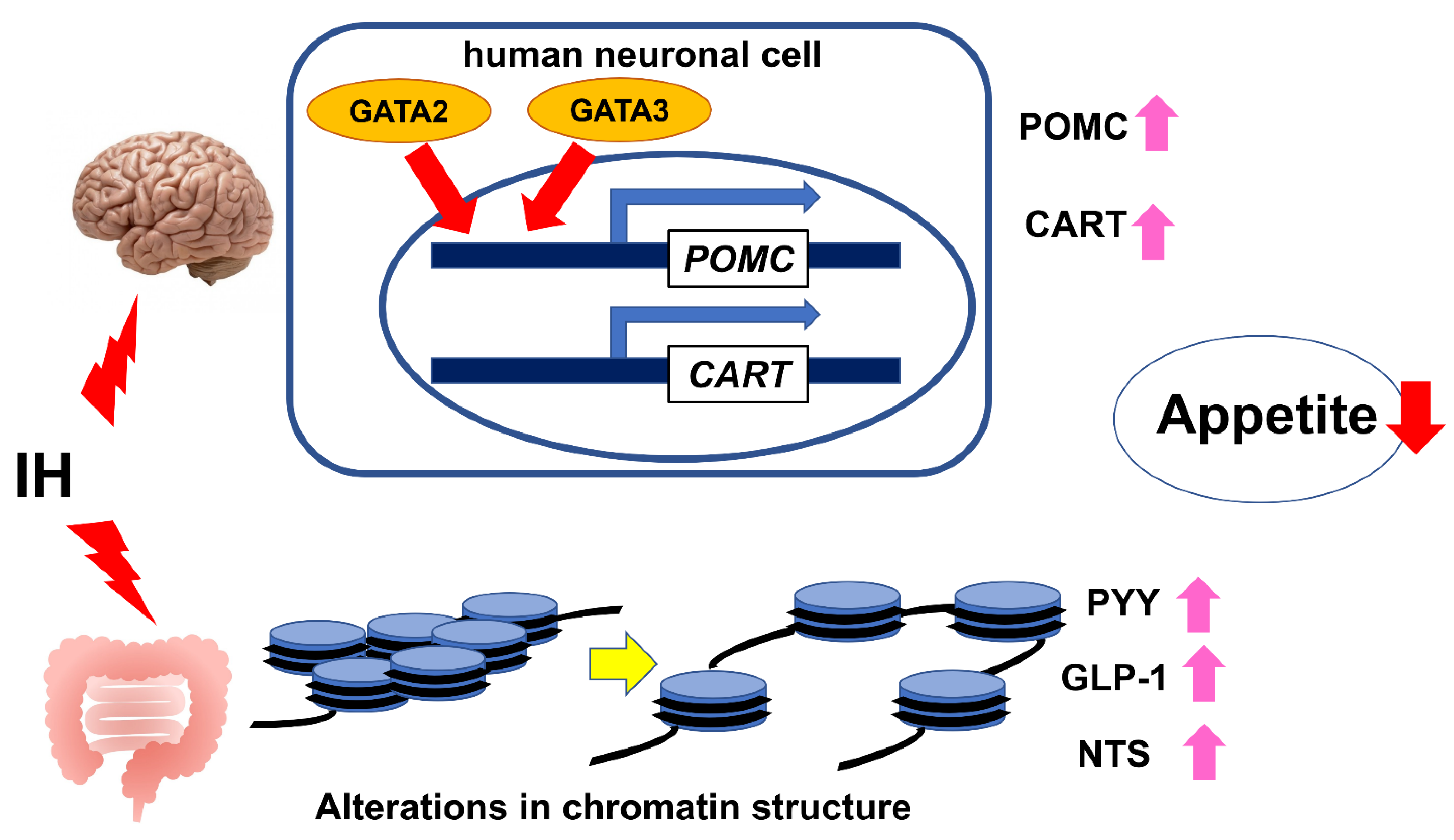
5. Appetite Regulation in SAS Patients
6. IH in Neuronal Cells
7. IH in Enteroendocrine Cells
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AGRP | Agouti-related peptide |
| AHI | Apnea-hypopnea index |
| ARC | Arcuate nucleus |
| ARNT | Aryl hydrocarbon receptor nuclear translocator |
| 5AZC | 5-Azacytidine |
| Bcl-xL | B cell lymphoma-extra large |
| Bcl-2 | B cell lymphoma 2 |
| BMI | Body Mass Index |
| CART | Cocaine- and amphetamine-regulated transcript |
| cADPR | Cyclic ADP-ribose |
| CCL2 | C-C motif chemokine ligand 2 |
| CD38 | Cluster of differentiation 38 |
| CNS | Central nervous system |
| CSA | Central sleep apnea |
| CXCL2 | Chemokine (C-X-C motif) ligand 2 |
| DM | Diabetes mellitus |
| DNA | Deoxyribonucleic acid |
| ENS | Enteric nervous system |
| GAL | Galanin |
| GALP | Galanin-like peptide |
| GCG | Glucagon |
| GLP-1 | Glucagon-like peptide-1 |
| GHRL | Ghrelin |
| GI | Gastrointestinal |
| HIF | Hypoxia-inducible factor |
| HIP/PAP | Hepatocarcinoma-intestine-pancreas/pancreatitis-associated protein |
| H3K27 | Lysine 27 of histone H3 |
| HRE | Hypoxia-response element |
| IH | Intermittent hypoxia |
| IL | Interleukin |
| KDMs | Lysine demethylases |
| MC4R | Melanocortin 4 receptor |
| miR | MicroRNA |
| NADPH | Nicotinamide adenine dinucleotide phosphate reduced form |
| Nox | NADPH oxidase(s) |
| NPY | Neuropeptide Y |
| NTS | Neurotensin |
| OSA | Obstructive sleep apnea |
| POMC | Proopiomelanocortin |
| PYY | Peptide YY |
| QRFP | Pyroglutamylated RFamide peptide |
| Reg | Regenerating gene |
| RETN | Resistin |
| ROS | Reactive oxygen species |
| SAS | Sleep apnea syndrome |
| siRNA | Small interfering RNA |
| TNF-α | Tumor necrosis factor-α |
| TSA | Trichostatin A |
References
- Morgenthaler, T.I.; Kagramanov, V.; Hanak, V.; Decker, P.A. Complex sleep apnea syndrome: Is it a unique clinical syndrome? Sleep 2006, 29, 1203–1209. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, J.A.; Veasey, S.C.; Morgan, B.J.; O’Donnell, C.P. Pathophysiology of sleep apnea. Physiol. Rev. 2010, 90, 47–112. [Google Scholar] [CrossRef] [PubMed]
- Nannapaneni, S.; Ramar, K.; Surani, S. Effect of obstructive sleep apnea on type 2 diabetes mellitus: A comprehensive literature review. World J. Diabetes 2013, 15, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Rajan, P.; Greenberg, H. Obstructive sleep apnea as a risk factor for type 2 diabetes mellitus. Nat. Sci. Sleep 2015, 5, 113–125. [Google Scholar] [CrossRef]
- Uchiyama, T.; Ota, H.; Itaya-Hironaka, A.; Shobatake, R.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Makino, M.; Kimura, H.; Takeda, M.; Ohbayashi, C.; et al. Up-regulation of selenoprotein P and HIP/PAP mRNAs in hepatocytes by intermittent hypoxia via down-regulation of miR-203. Biochem. Biophys. Rep. 2017, 11, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Ota, H.; Tamaki, S.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Morioka, T.; Takasawa, S.; Kimura, H. Attenuation of glucose-induced insulin secretion by intermittent hypoxia via down-regulation of CD38. Life Sci. 2012, 90, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, R.; Singh, M.; Nida, M.; Waheed, I.; Khan, A.; Ahmed, S.; Naseem, J.; Champeau, D. Effect of obstructive sleep apnea hypopnea syndrome on lipid profile: A meta-regression analysis. J. Clin. Sleep Med. 2014, 15, 475–489. [Google Scholar] [CrossRef] [PubMed]
- Bradley, T.D.; Floras, J.S. Obstructive sleep apnoea and its cardiovascular consequences. Lancet 2009, 373, 82–93. [Google Scholar] [CrossRef]
- Javaheri, S.; Javaheri, S.; Javaheri, A. Sleep apnea, heart failure, and pulmonary hypertension. Curr. Heart Fail. Rep. 2013, 10, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Vaessen, T.J.A.; Overeem, S.; Sitskoorn, M.M. Cognitive complaints in obstructive sleep apnea. Sleep Med. Rev. 2015, 19, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Bucks, R.S.; Olaithe, M.; Eastwood, P. Neurocognitive function in obstructive sleep apnoea: A meta-review. Respirology 2013, 18, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Wallace, A.; Bucks, R.S. Memory and obstructive sleep apnea: A meta-analysis. Sleep 2013, 36, 203–220. [Google Scholar] [CrossRef] [PubMed]
- Carotenuto, M.; Esposito, M.; Parisi, L.; Gallai, B.; Marotta, R.; Pascotto, A.; Roccella, M. Depressive symptoms and childhood sleep apnea syndrome. Neuropsychiatr. Dis. Treat. 2012, 8, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Franklin, K.A.; Lindberg, E. Obstructive sleep apnea is a common disorder in the population-a review on the epidemiology of sleep apnea. J. Thorac. Dis. 2015, 8, 1311–1322. [Google Scholar] [CrossRef]
- Arnaud, C.; Poulain, L.; Lévy, P.; Dematteis, M. Inflammation contributes to the atherogenic role of intermittent hypoxia in apolipoprotein-E knock out mice. Atherosclerosis 2011, 219, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Ota, H.; Fujita, Y.; Yamauchi, M.; Muro, S.; Kimura, H.; Takasawa, S. Relationship between intermittent hypoxia and Type 2 diabetes in sleep apnea syndrome. Int. J. Mol. Sci. 2019, 20, 4756. [Google Scholar] [CrossRef] [PubMed]
- Drager, L.F.; Yao, Q.; Hernandez, K.L.; Shin, M.-K.; Bevans-Fonti, S.; Gay, J.; Sussan, T.E.; Jun, J.C.; Myers, A.C.; Olivercrona, G.; et al. Chronic intermittent hypoxia induces atherosclerosis via activation of adipose angiopoietin-like 4. Am. J. Respir. Crit. Care. Med. 2013, 188, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Lavie, L. Intermittent hypoxia: The culprit of oxidative stress, vascular inflammation and dyslipidemia in obstructive sleep apnea. Expert Rev. Respir. Med. 2008, 2, 75–84. [Google Scholar] [CrossRef]
- Savransky, V.; Nanayakkara, A.; Li, J.; Bevans, S.; Smith, P.L.; Rodriguez, A.; Polotsky, V.Y. Chronic intermittent hypoxia induces atherosclerosis. Am. J. Respir. Crit. Care Med. 2007, 175, 1290–1297. [Google Scholar] [CrossRef]
- Htoo, A.K.; Greenberg, H.; Tongia, S.; Chen, G.; Henderson, T.; Wilson, D.; Liu, S.F. Activation of nuclear factor κB in obstructive sleep apnea: A pathway leading to systemic inflammation. Sleep Breath 2006, 10, 43–50. [Google Scholar] [CrossRef]
- Oliver, K.M.; Garvey, J.F.; Ng, C.T.; Veale, D.J.; Fearon, U.; Cummins, E.P.; Taylor, C.T. Hypoxia activates NF-κB-dependent gene expression through the canonical signaling pathway. Antioxid. Redox Signal. 2009, 11, 2057–2064. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, M.; Nakano, H.; Maekawa, J.; Okamoto, Y.; Ohnishi, Y.; Suzuki, T.; Kimura, H. Oxidative stress in obstructive sleep apnea. Chest 2005, 127, 1674–1679. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, E.C. Sympathetic over activity in the etiology of hypertension of obstructive sleep apnea. Sleep 2003, 26, 15–19. [Google Scholar] [CrossRef]
- Young, T.; Skatrud, J.; Peppard, P.E. Risk factors for obstructive sleep apnea in adults. JAMA 2004, 291, 2013–2016. [Google Scholar] [CrossRef] [PubMed]
- Eckert, D.J.; White, D.P.; Jordan, A.S.; Malhotra, A.; Wellman, A. Defining phenotypic causes of obstructive sleep apnea. Idetifcation of novel therapeutic targets. Am. J. Respir. Crit. Care Med. 2013, 188, 996–1004. [Google Scholar] [CrossRef]
- Osman, A.M.; Carter, S.G.; Carberry, J.C.; Eckert, D.J. Obstructive sleep apnea: Current perspectives. Nat. Sci. Sleep 2018, 10, 21–34. [Google Scholar] [CrossRef]
- Tasali, E.; Ip, M.S.M. Obstructive sleep apnea and metabolic syndrome: Alterations in glucose metabolism and inflammation. Proc. Am. Thorac. Soc. 2008, 5, 207–217. [Google Scholar] [CrossRef]
- Young, T.; Peppard, P.E.; Taheri, S. Excess weight and sleep-disordered breathing. J. Appl. Physiol. 2005, 99, 1592–1599. [Google Scholar] [CrossRef]
- Strohl, K.P.; Redline, S. Recognition of obstructive sleep apnea. Am. J. Respir. Crit. Care Med. 1996, 154, 279–289. [Google Scholar] [CrossRef]
- Levinson, P.D.; McGarvey, S.T.; Carlisle, C.C.; Eveloff, S.E.; Herbert, P.N.; Millman, R.P. Adiposity and cardiovascular risk factors in men with obstructive sleep apnea. Chest 1993, 103, 1336–1342. [Google Scholar] [CrossRef] [PubMed]
- Phillips, B.; Cook, Y.; Schmitt, F.; Berry, D. Sleep apnea: Prevalence of risk factors in a general population. South Med. J. 1989, 82, 1090–1092. [Google Scholar] [CrossRef] [PubMed]
- Vgontzas, A.N.; Tan, T.L.; Bixler, E.O.; Martin, L.F.; Shubert, D.; Kales, A. Sleep apnea and sleep disruption in obese patients. Arch. Intern. Med. 1994, 154, 1705–1711. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, O.R.; Carmo, I.; Paiva, T.; Figueira, M.L. Body mass index and neuropsychological and emotional variables: Joint contribution for the screening of sleep apnoea syndrome in obese. Sleep Sci. 2021, 14, 19–26. [Google Scholar] [CrossRef]
- Peppard, P.E.; Young, T.; Palta, M.; Dempsey, J.; Skatrud, J. Longitudinal study of moderate weight change and sleep-disordered breathing. JAMA 2000, 284, 3015–3021. [Google Scholar] [CrossRef] [PubMed]
- Heinzer, R.; Vat, S.; Marques-Vidal, P.; Marti-Soler, H.; Andries, D.; Tobback, N.; Mooser, V.; Preisig, M.; Malhotra, A.; Waeber, G.; et al. Prevalence of sleep-disordered breathing in the general population: The HypnoLaus study. Lancet Respir. Med. 2015, 3, 310–318. [Google Scholar] [CrossRef]
- Li, K.K.; Kushida, C.; Powell, N.B.; Riley, R.W.; Guilleminault, C. Obstructive sleep apnea syndrome: A comparison between Far-East Asian and white men. Laryngoscope 2000, 110, 1689–1693. [Google Scholar] [CrossRef]
- Zhang, C.; Cheng, Y.; Liu, F.; Ma, J.; Wang, G. A community study of the risk for obstructive sleep apnea and respiratory inflammation in an adult Chinese population. Postgrad. Med. 2021, 133, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Shechter, A. Obstructive sleep apnea and energy balance regulation: A systematic review. Sleep Med. Rev. 2017, 34, 59–69. [Google Scholar] [CrossRef]
- Romero-Corral, A.; Caples, S.M.; Lopez-Jimenez, F.; Somers, V.K. Interactions between obesity and obstructive sleep apnea: Implications for treatment. Chest 2010, 137, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.S.; Waight, C.; Doyle, G.; Rossa, K.R.; Sullivan, K.A. Liking for high fat foods in patients with obstructive sleep apnoea. Appetite 2014, 78, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Kechribari, I.; Kontogianni, M.D.; Georgoulis, M.; Lamprou, K.; Mourati, I.; Vagiakis, E.; Yiannakouris, N. Associations between red meat intake and sleep parameters in patients with obstructive sleep apnea. J. Acad. Nutr. Diet 2020, 120, 1042–1053. [Google Scholar] [CrossRef]
- Kechribari, I.; Kontogianni, M.D.; Georgoulis, M.; Lamprou, K.; Vagiakis, E.; Yiannakouris, N. Higher refined Cereal Grain intake is Positively Associated with Apnoea-Hypopnoea Index in Patients with Obstructive Sleep Apnoea. 2021. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/jhn.12933 (accessed on 25 November 2021).
- Gaona-Pineda, E.B.; Martinez-Tapia, B.; Rodríguez-Ramírez, S.; Guerrero-Zúñiga, S.; Perez-Padilla, R.; Shamah-Levy, T. Dietary patterns and sleep disorders in Mexican adults from a National Health and Nutrition Survey. J. Nutr. Sci. 2021, 10, e34. [Google Scholar] [CrossRef] [PubMed]
- de Melo, C.M.; Re, M.P.D.; Quaresma, M.V.L.D.S.; Antunes, H.K.M.; Togeiro, S.M.; Ribeiro, S.M.L.; Tufik, S.; de Mello, M.T. Relationship of evening meal with sleep quality in obese individuals with obstructive sleep apnea. Clin. Nutr. ESPE 2019, 29, 231–236. [Google Scholar] [CrossRef] [PubMed]
- Nanduri, J.; Peng, Y.J.; Yuan, G.; Kumar, G.K.; Prabhakar, N.R. Hypoxia-inducible factors and hypertension: Lessons from sleep apnea syndrome. J. Mol. Med. 2015, 93, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.J.; Yuan, G.; Ramakrishnan, D.; Sharma, S.D.; Bosch-Marce, M.; Kumar, G.K.; Semenza, G.L.; Prabhakar, N.R. Heterozygous HIF-1α deficiency impairs carotid body-mediated systemic responses and reactive oxygen species generation in mice exposed to intermittent hypoxia. J. Physiol. 2006, 577, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L.; Prabhakar, N.R. HIF-1-dependent respiratory, cardiovascular, and redox responses to chronic intermittent hypoxia. Antioxid. Redox Signal. 2007, 9, 1391–1396. [Google Scholar] [CrossRef]
- Yuan, G.; Nanduri, J.; Bhasker, C.R.; Semenza, G.L.; Prabhakar, N.R. Ca2+/calmodulin kinase-dependent activation of hypoxia inducible factor 1 transcriptional activity in cells subjected to intermittent hypoxia. J. Biol. Chem. 2005, 280, 4321–4328. [Google Scholar] [CrossRef]
- Mole, D.R.; Blancher, C.; Copley, R.R.; Pollard, P.J.; Gleadle, J.M.; Ragoussis, J.; Ratcliffe, P.J. Genome-wide association of hypoxia-inducible factor (HIF)-1α and HIF-2α DNA binding with expression profiling of hypoxia-inducible transcripts. J. Biol. Chem. 2009, 284, 16767–16775. [Google Scholar] [CrossRef]
- Semenza, G.L. Regulation of oxygen homeostasis by hypoxia-inducible factor 1. Physiology 2009, 24, 97–106. [Google Scholar] [CrossRef]
- Prabhakar, N.R.; Semenza, G.L. Adaptive and maladaptive cardiorespiratory responses to continuous and intermittent hypoxia mediated by hypoxia-inducible factors 1 and 2. Physiol. Rev. 2012, 92, 967–1003. [Google Scholar] [CrossRef]
- Wood, S.M.; Gleadle, J.M.; Pugh, C.W.; Hankinson, O.; Ratcliffe, P.J. The role of the aryl hydrocarbon receptor nuclear translocator (ARNT) in hypoxic induction of gene expression. Studies in ARNT-deficient cells. J. Biol. Chem. 1996, 271, 15117–15123. [Google Scholar] [CrossRef] [PubMed]
- Gabryelska, A.; Szmyd, B.; Szemraj, J.; Stawski, R.; Sochal, M.; Białasiewicz, P. Patients with obstructive sleep apnea present with chronic upregulation of serum HIF-1α protein. J. Clin. Sleep Med. 2020, 16, 1761–1768. [Google Scholar] [CrossRef]
- Peng, Y.J.; Nanduri, J.; Yuan, G.; Wang, N.; Deneris, E.; Pendyala, S.; Natarajan, V.; Kumar, G.K.; Prabhakar, N.R. NADPH oxidase is required for the sensory plasticity of the carotid body by chronic intermittent hypoxia. J. Neurosci. 2009, 29, 4903–4910. [Google Scholar] [CrossRef] [PubMed]
- Souvannakitti, D.; Nanduri, J.; Yuan, G.; Kumar, G.K.; Fox, A.P.; Prabhakar, N.R. NADPH oxidase-dependent regulation of T-type Ca2+ channels and ryanodine receptors mediate the augmented exocytosis of catecholamines from intermittent hypoxia-treated neonatal rat chromaffin cells. J. Neurosci. 2010, 30, 10763–10772. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Khan, S.A.; Luo, W.; Nanduri, J.; Semenza, G.L.; Prabhakar, N.R. Hypoxia-inducible factor 1 mediates increased expression of NADPH oxidase-2 in response to intermittent hypoxia. J. Cell Physiol. 2011, 226, 2925–2933. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.L.; Dunne, K.; Walport, L.J.; Flashman, E.; Kawamura, A. Epigenetic regulation by histone demethylases in hypoxia. Epigenomics 2015, 7, 791–811. [Google Scholar] [CrossRef] [PubMed]
- Nanduri, J.; Wang, N.; Wang, B.L.; Prabhakar, N.R. Lysine demethylase KDM6B regulates HIF-1α-mediated systemic and cellular responses to intermittent hypoxia. Physiol. Genom. 2021, 53, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Reichmuth, K.J.; Austin, D.; Skatrud, J.B.; Young, T. Association of sleep apnea and type II diabetes: A population-based study. Am. J. Respir. Crit. Care Med. 2005, 172, 1590–1595. [Google Scholar] [CrossRef]
- Tasali, E.; Mokhlesi, B.; Cauter, E.V. Obstructive sleep apnea and type 2 diabetes: Interacting epidemics. Chest 2008, 133, 496–506. [Google Scholar] [CrossRef]
- Gabryelska, A.; Chrzanowski, J.; Sochal, M.; Kaczmarski, P.; Turkiewicz, S.; Ditmer, M.; Karuga, F.F.; Czupryniak, L.; Białasiewicz, P. Nocturnal oxygen saturation parameters as independent risk factors for Type 2 diabetes mellitus among obstructive sleep apnea patients. J. Clin. Med. 2021, 10, 3770. [Google Scholar] [CrossRef]
- Shao, Y.; Lv, C.; Yuan, Q.; Wang, Q. Levels of serum 25(OH)VD3, HIF-1α, VEGF, vWf, and IGF-1 and their correlation in type 2 diabetes patients with different urine albumin creatinine ratio. J. Diabetes Res. 2016, 2016, 1925424. [Google Scholar] [CrossRef]
- Nagao, A.; Kobayashi, M.; Koyasu, S.; Chow, C.C.T.; Harada, H. HIF-1–dependent reprogramming of glucose metabolic pathway of cancer cells and its therapeutic significance. Int. J. Mol. Sci. 2019, 20, 238. [Google Scholar] [CrossRef]
- Xu, J.; Long, Y.; Gozal, D.; Epstein, P.N. β-cell death and proliferation after intermittent hypoxia: Role of oxidative stress. Free Radic. Biol. Med. 2009, 46, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Takasawa, S.; Nata, K.; Yonekura, H.; Okamoto, H. Cyclic ADP-ribose in insulin secretion from pancreatic β cells. Science 1993, 259, 370–373. [Google Scholar] [CrossRef] [PubMed]
- Takasawa, S.; Tohgo, A.; Noguchi, N.; Koguma, T.; Nata, K.; Sugimoto, T.; Yonekura, H.; Okamoto, H. Synthesis and hydrolysis of cyclic ADP-ribose by human leukocyte antigen CD38 and inhibition of the hydrolysis by ATP. J. Biol. Chem. 1993, 268, 26052–26054. [Google Scholar] [CrossRef]
- Takasawa, S.; Akiyama, T.; Nata, K.; Kuroki, M.; Tohgo, A.; Noguchi, N.; Kobayashi, S.; Kato, I.; Katada, T.; Okamoto, H. Cyclic ADP-ribose and inositol 1,4,5-trisphospate as alternate second messengers for intracellular Ca2+ mobilization in normal and diabetic β-cells. J. Biol. Chem. 1998, 273, 2497–2500. [Google Scholar] [CrossRef]
- Kato, I.; Yamamoto, Y.; Fujimura, M.; Noguchi, N.; Takasawa, S.; Okamoto, H. CD38 disruption impairs glucose-induced increases in cyclic ADP-ribose, [Ca2+]i, and insulin secretion. J. Biol. Chem. 1999, 274, 1869–1872. [Google Scholar] [CrossRef]
- Okamoto, H.; Takasawa, S. Recent advances in the Okamoto model: The CD38-cyclic ADP-ribose signal system and the regenerating gene protein (Reg)-Reg receptor system in β-cells. Diabetes 2002, 51, S462–S473. [Google Scholar] [CrossRef]
- Takasawa, S.; Kuroki, M.; Nata, K.; Noguchi, N.; Ikeda, T.; Yamauchi, A.; Ota, H.; Itaya-Hironaka, A.; Sakuramoto-Tsuchida, S.; Takahashi, I.; et al. A novel ryanodine receptor expressed in pancreatic islets by alternative splicing from type 2 ryanodine receptor gene. Biochem. Biophys. Res. Commun. 2010, 397, 140–145. [Google Scholar] [CrossRef]
- Okamoto, H.; Takasawa, S. Okamoto model for necrosis and its expansions, CD38-cyclic ADP-ribose signal system for intracelllular Ca2+ mobilization and Reg ([Regenerating gene] protein)-Reg receptor system for cell regeneration. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2021, 79, 423–461. [Google Scholar] [CrossRef]
- Terazono, K.; Yamamoto, H.; Takasawa, S.; Shiga, K.; Yonemura, Y.; Tochino, Y.; Okamoto, H. A novel gene activated in regenerating islets. J. Biol. Chem. 1988, 263, 2111–2114. [Google Scholar] [CrossRef]
- Watanabe, T.; Yonemura, Y.; Yonekura, H.; Suzuki, Y.; Miyashita, H.; Sugiyama, K.; Moriizumi, S.; Unno, M.; Tanaka, O.; Kondo, H.; et al. Pancreatic beta-cell replication and amelioration of surgical diabetes by Reg protein. Proc. Natl. Acad. Sci. USA 1994, 91, 3589–3592. [Google Scholar] [CrossRef]
- Takasawa, S. Regenerating gene (REG) product and its potential clinical usage. Expert Opin. Ther. Targets 2016, 20, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Shervani, N.J.; Takasawa, S.; Uchigata, Y.; Akiyama, T.; Nakagawa, K.; Noguchi, N.; Takada, H.; Takahashi, I.; Yamauchi, A.; Ikeda, T.; et al. Autoantibodies to REG, a beta-cell regeneration factor, in diabetic patients. Eur. J. Clin. Investig. 2004, 34, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Ota, H.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Miyaoka, T.; Fujimura, T.; Tsujinaka, H.; Yoshimoto, K.; Nakagawa, K.; Tamaki, S.; et al. Pancreatic β-cell proliferation by intermittent hypoxia via up-regulation of Reg family genes and HGF gene. Life Sci. 2013, 93, 664–672. [Google Scholar] [CrossRef] [PubMed]
- Takeda, Y.; Itaya-Hironaka, A.; Yamauchi, A.; Makino, M.; Sakuramoto-Tsuchida, S.; Ota, H.; Kawaguchi, R.; Takasawa, S. Intermittent hypoxia upregulates the renin and Cd38 mRNAs in renin-producing cells via the downregulation of miR-203. Int. J. Mol. Sci. 2021, 22, 10127. [Google Scholar] [CrossRef]
- Uchiyama, T.; Itaya-Hironaka, A.; Yamauchi, A.; Makino, M.; Sakuramoto-Tsuchida, S.; Shobatake, R.; Ota, H.; Takeda, M.; Ohbayashi, C.; Takasawa, S. Intermittent hypoxia up-regulates CCL2, RETN, and TNFα mRNAs in adipocytes via down-regulation of miR-452. Int. J. Mol. Sci. 2019, 20, 1960. [Google Scholar] [CrossRef] [PubMed]
- Takasawa, S.; Shobatake, R.; Itaya-Hironaka, A.; Yamauchi, A.; Makino, M.; Sakuramoto-Tsuchida, S.; Uchiyama, T.; Tsuchida, C.; Ota, H. Up-regulation of diabetes-associated myokines by intermittent hypoxia in mammalian skeletal muscle cells. In Proceedings of the IDF World Diabetes Congress, Busan, Korea, 2–6 December 2019. [Google Scholar]
- Takasawa, S.; Shobatake, R.; Itaya-Hironaka, A.; Makino, M.; Sakuramoto-Tsuchida, S.; Uchiyama, T.; Ota, H.; Yamauchi, A. Intermittent hypoxia increased the expressions of interleukin (IL)-8, osteonectin, and myonectin via OCT1 and NRF2 binding to the promoters in muscle cells. Diabetes 2020, 69 (Suppl. 1), 1887-P. [Google Scholar] [CrossRef]
- Takasawa, S.; Shobatake, R.; Itaya-Hironaka, A.; Makino, M.; Sakuramoto-Tsuchida, S.; Uchiyama, T.; Ota, H.; Yamauchi, A. Up-regulation of IL-8, osteonectin, and myonectin mRNAs by intermittent hypoxia via OCT1- and NRF2-mediated mechanisms in skeletal muscle cells. Diabetologia 2020, 63 (Suppl. 1), 453. [Google Scholar]
- Uchiyama, T.; Ota, H.; Ohbayashi, C.; Takasawa, S. Effect of intermittent hypoxia on cytokine expression involved in insulin resistance. Int. J. Mol. Sci. 2021, 22, 12898. [Google Scholar] [CrossRef]
- Clarke, I.J. Hypothalamus as an endocrine organ. Compr. Physiol. 2015, 5, 217–253. [Google Scholar] [CrossRef] [PubMed]
- Schneeberger, M.; Gomis, R.; Claret, M. Hypothalamic and brainstem neuronal circuits controlling homeostatic energy balance. J. Endocrinol. 2014, 220, T25–T46. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Simpson, K.A.; Minnion, J.S.; Shillito, J.C.; Bloom, S.R. The role of gut hormones and the hypothalamus in appetite regulation. Endocr. J. 2010, 57, 359–372. [Google Scholar] [CrossRef] [PubMed]
- Gautron, L.; Elmquist, J.K.; Williams, K.W. Neural control of energy balance: Translating circuits to therapies. Cell 2015, 161, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Flier, J.S. Obesity wars: Molecular progress confronts an expanding epidemic. Cell 2004, 116, 337–350. [Google Scholar] [CrossRef]
- Robinson, J.K.; Brewer, A. Galanin: A potential role in mesolimbic dopamine-mediated instrumental behavior. Neurosci. Biobehav. Rev. 2008, 32, 1485–1493. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bruzzone, F.; Lectez, B.; Tollemer, H.; Leprince, J.; Dujardin, C.; Rachidi, W.; Chatenet, D.; Baroncini, M.; Beauvillain, J.; Vallarino, M.; et al. Anatomical distribution and biochemical characterization of the novel RFamide peptide 26RFa in the human hypothalamus and spinal cord. J. Neurochem. 2006, 99, 616–627. [Google Scholar] [CrossRef]
- Lawrence, C.; Fraley, G.S. Galanin-like peptide (GALP) is a hypothalamic regulator of energy homeostasis and reproduction. Front. Neuroendocrinol. 2011, 32, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Andermann, M.L.; Lowell, B.B. Toward a wiring diagram understanding of appetite control. Neuron 2017, 95, 757–778. [Google Scholar] [CrossRef]
- Filpa, V.; Moro, E.; Protasoni, M.; Crema, F.; Frigo, G.; Giaroni, C. Role of glutamatergic neurotransmission in the enteric nervous system and brain-gut axis in health and disease. Neuropharmacology 2016, 111, 14–33. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.K.; Chang, R.B.; Strochlic, D.E.; Umans, B.D.; Lowell, B.B.; Liberles, S.D. Sensory neurons that detect stretch and nutrients in the digestive system. Cell 2016, 166, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Tellez, L.A.; Perkins, M.H.; Perez, I.O.; Qu, T.; Ferreira, J.; Ferreira, T.L.; Quinn, D.; Liu, Z.; Gao, X.; et al. A neural circuit for gut-induced reward. Cell 2018, 175, 665–678. [Google Scholar] [CrossRef] [PubMed]
- Kaelberer, M.M.; Buchanan, K.L.; Klein, M.E.; Barth, B.B.; Montoya, M.M.; Shen, X.; Bohórquez, D.V. A gut-brain neural circuit for nutrient sensory transduction. Science 2018, 361, eaat5236. [Google Scholar] [CrossRef]
- Bauer, P.V.; Hamr, S.C.; Duca, F.A. Regulation of energy balance by a gut-brain axis and involvement of the gut microbiota. Cell. Mol. Life Sci. 2016, 73, 737–755. [Google Scholar] [CrossRef]
- Basoglu, O.K.; Sarac, F.; Sarac, S.; Uluer, H.; Yilmaz, C. Metabolic syndrome, insulin resistance, fibrinogen, homocysteine, leptin, and C-reactive protein in obese patients with obstructive sleep apnea syndrome. Ann. Thorac. Med. 2011, 6, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Kapsimalis, F.; Varouchakis, G.; Manousaki, A.; Daskas, S.; Nikita, D.; Kryger, M.; Gourgoulianis, K. Association of sleep apnea severity and obesity with insulin resistance, C-reactive protein, and leptin levels in male patients with obstructive sleep apnea. Lung 2008, 186, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Papaioannou, I.; Patterson, M.; Twigg, G.L.; Vazir, A.; Ghatei, M.; Morrell, M.J.; Polkey, M.I. Lack of association between impaired glucose tolerance and appetite regulating hormones in patients with obstructive sleep apnea. J. Clin. Sleep Med. 2011, 7, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Chihara, Y.; Akamizu, T.; Azuma, M.; Murase, K.; Harada, Y.; Tanizawa, K.; Handa, T.; Oga, T.; Mishima, M.; Chin, K. Among metabolic factors, significance of fasting and postprandial increases in acyl and desacyl ghrelin and the acyl/desacyl ratio in obstructive sleep apnea before and after treatment. J. Clin. Sleep Med. 2015, 11, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Ursavas, A.; Ilcol, Y.O.; Nalci, N.; Karadag, M.; Ege, E. Ghrelin, leptin, adiponectin, and resistin levels in sleep apnea syndrome: Role of obesity. Ann. Thorac. Med. 2010, 5, 161–165. [Google Scholar] [CrossRef]
- Sun, M.; Niu, X.; Xiao, X.; Chen, X. The differences in plasma/serum ghrelin levels between obstructive sleep apnea-hypopnea patients and controls: A protocol for systematic review and meta-analysis. Medicine 2021, 100, e24368. [Google Scholar] [CrossRef]
- Matsumoto, T.; Harada, N.; Azuma, M.; Chihara, Y.; Murase, K.; Tachikawa, R.; Minami, T.; Hamada, S.; Tanizawa, K.; Inouchi, M.; et al. Plasma incretin levels and dipeptidyl peptidase-4 activity in patients with obstructive sleep apnea. Ann. Am. Thorac. Soc. 2016, 13, 1378–1387. [Google Scholar] [CrossRef] [PubMed]
- Shobatake, R.; Takasawa, K.; Ota, H.; Itaya-Hironaka, A.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Uchiyama, T.; Makino, M.; Sugie, K.; Takasawa, S.; et al. Up-regulation of POMC and CART mRNAs by intermittent hypoxia via GATA transcription factors in human neuronal cells. Int. J. Biochem. Cell Biol. 2018, 95, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Park, A.M.; Nagase, H.; Kumar, S.V.; Suzuki, Y.J. Acute intermittent hypoxia activates myocardial cell survival signaling. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H751–H757. [Google Scholar] [CrossRef] [PubMed]
- Shobatake, R.; Itaya-Hironaka, A.; Yamauchi, A.; Makino, M.; Sakuramoto-Tsuchida, S.; Uchiyama, T.; Ota, H.; Takahashi, N.; Ueno, S.; Sugie, K.; et al. Intermittent hypoxia up-regulates gene expressions of peptide YY (PYY), glucagon-like peptide-1 (GLP-1), and neurotensin (NTS) in enteroendocrine cells. Int. J. Mol. Sci. 2019, 20, 1849. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shobatake, R.; Ota, H.; Takahashi, N.; Ueno, S.; Sugie, K.; Takasawa, S. Anorexigenic Effects of Intermittent Hypoxia on the Gut—Brain Axis in Sleep Apnea Syndrome. Int. J. Mol. Sci. 2022, 23, 364. https://doi.org/10.3390/ijms23010364
Shobatake R, Ota H, Takahashi N, Ueno S, Sugie K, Takasawa S. Anorexigenic Effects of Intermittent Hypoxia on the Gut—Brain Axis in Sleep Apnea Syndrome. International Journal of Molecular Sciences. 2022; 23(1):364. https://doi.org/10.3390/ijms23010364
Chicago/Turabian StyleShobatake, Ryogo, Hiroyo Ota, Nobuyuki Takahashi, Satoshi Ueno, Kazuma Sugie, and Shin Takasawa. 2022. "Anorexigenic Effects of Intermittent Hypoxia on the Gut—Brain Axis in Sleep Apnea Syndrome" International Journal of Molecular Sciences 23, no. 1: 364. https://doi.org/10.3390/ijms23010364
APA StyleShobatake, R., Ota, H., Takahashi, N., Ueno, S., Sugie, K., & Takasawa, S. (2022). Anorexigenic Effects of Intermittent Hypoxia on the Gut—Brain Axis in Sleep Apnea Syndrome. International Journal of Molecular Sciences, 23(1), 364. https://doi.org/10.3390/ijms23010364