
Knockdown of RRM1 with Adenoviral shRNA Vectors to Inhibit Tumor Cell Viability and Increase Chemotherapeutic Sensitivity to Gemcitabine in Bladder Cancer Cells

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
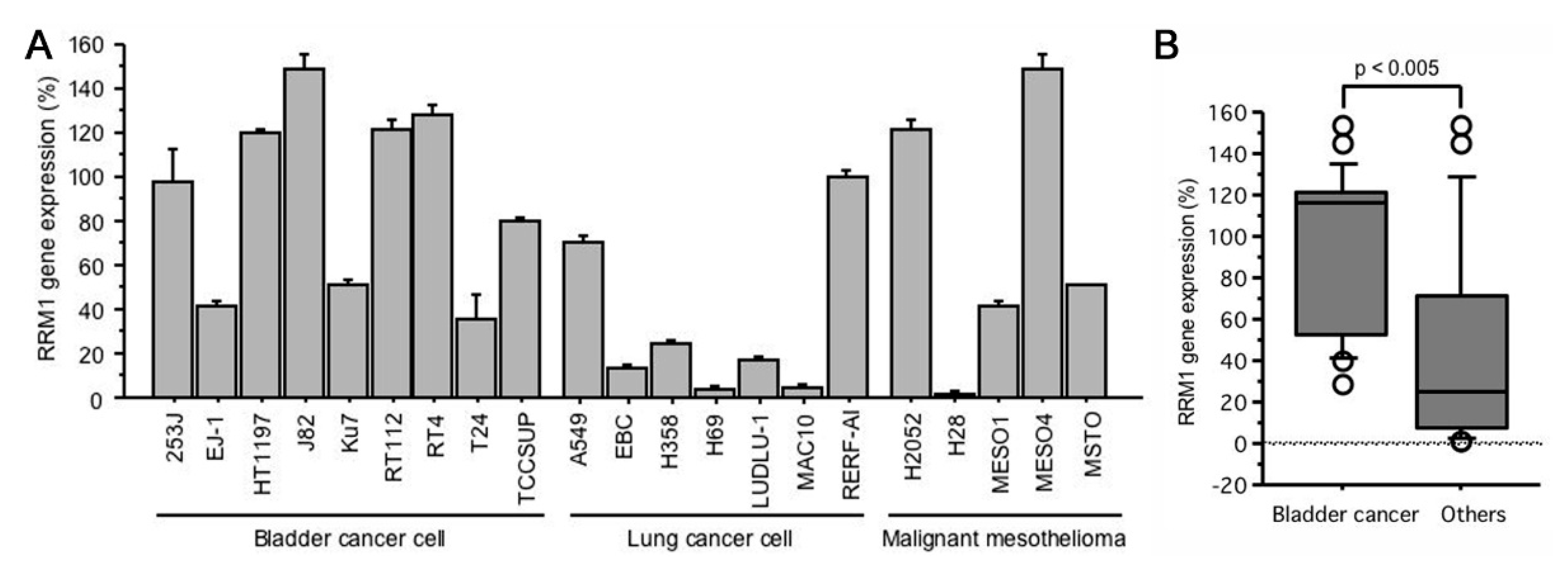
2.1. RRM1 Gene Expression Is Higher in Bladder Cancer Cell Lines
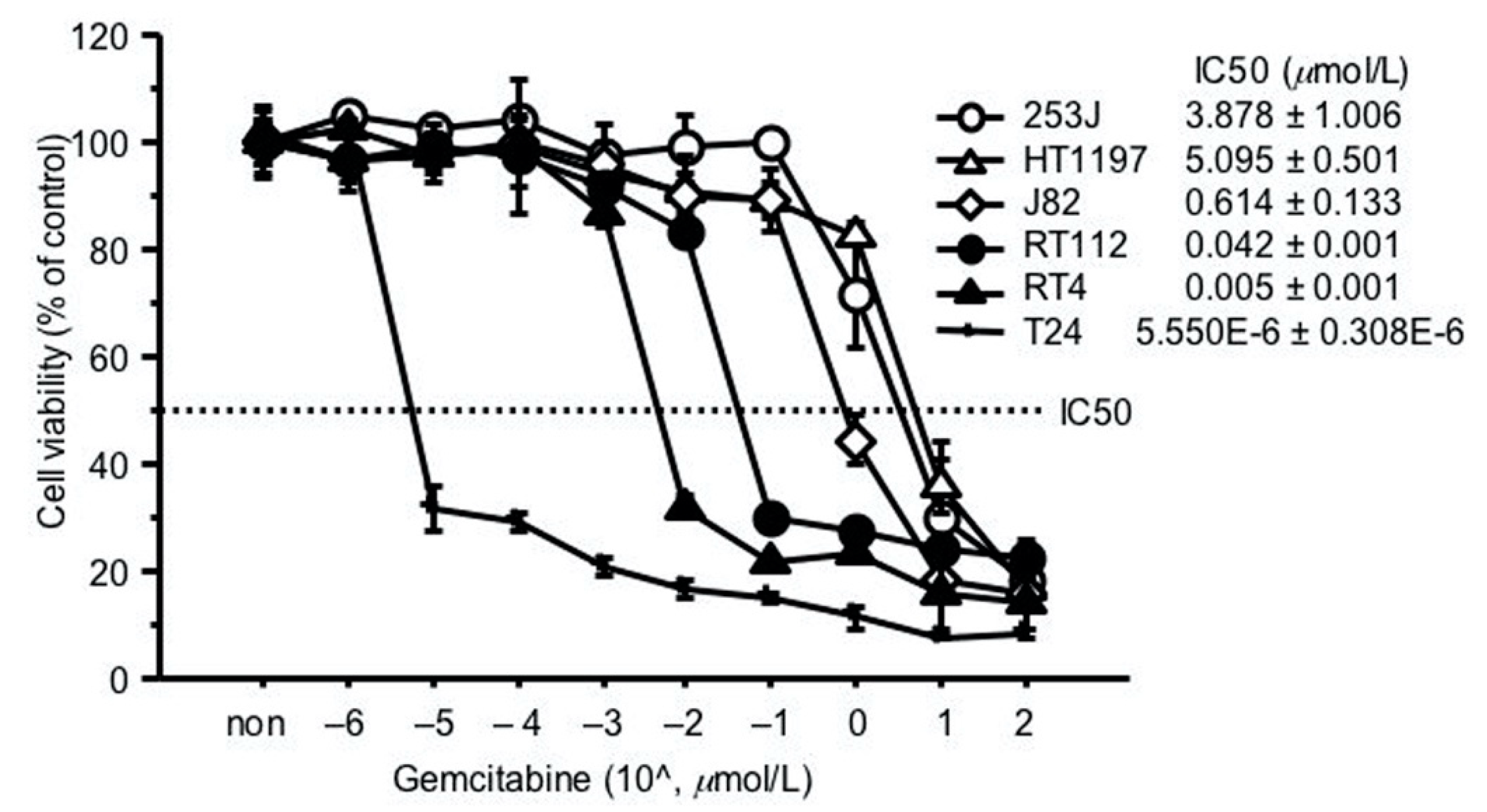
2.2. GEM Resistance in Bladder Cancer Cell Lines
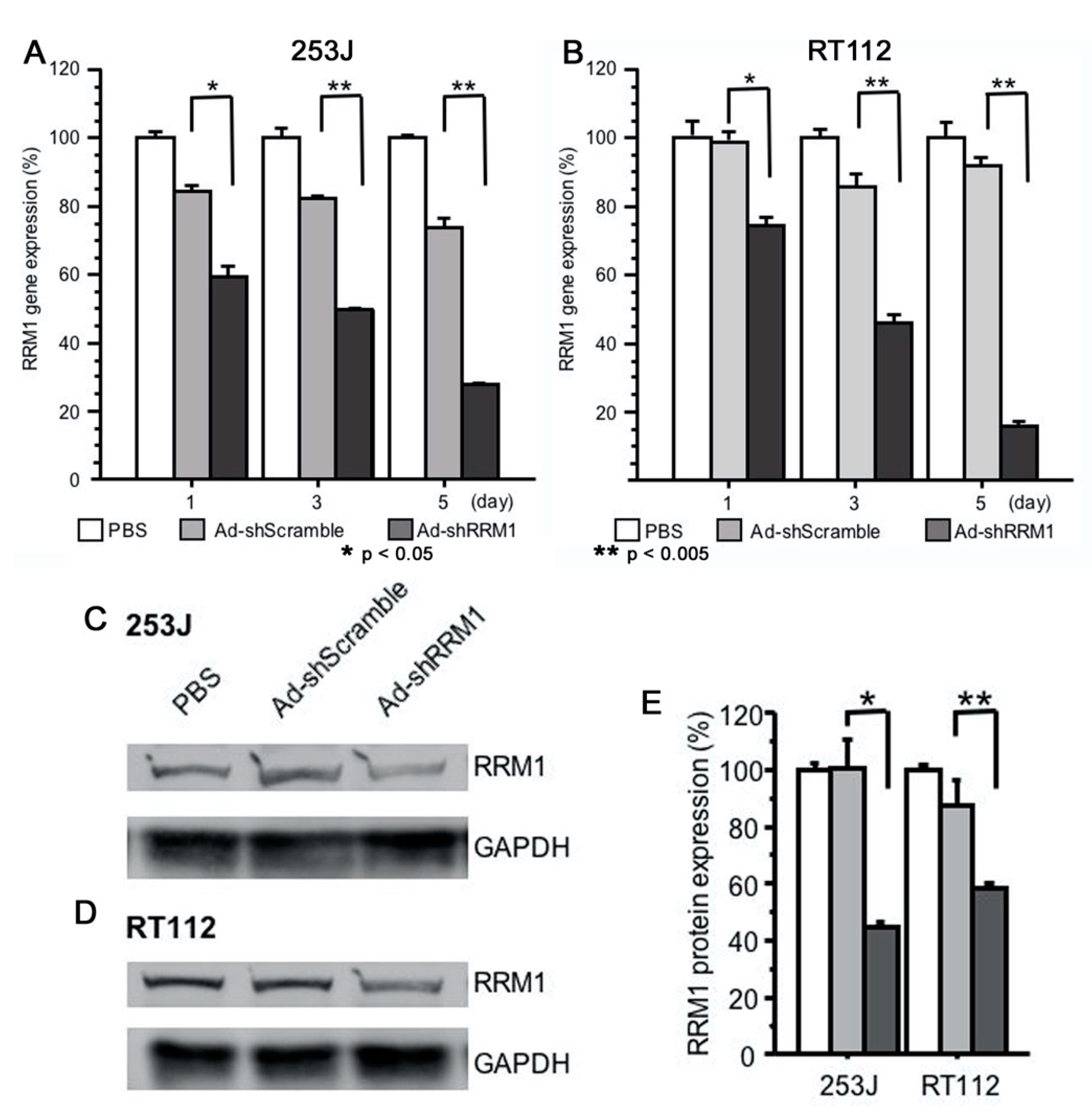
2.3. Ad-shRRM1 Effectively Downregulates RRM1 Expression in Bladder Cancer Cells
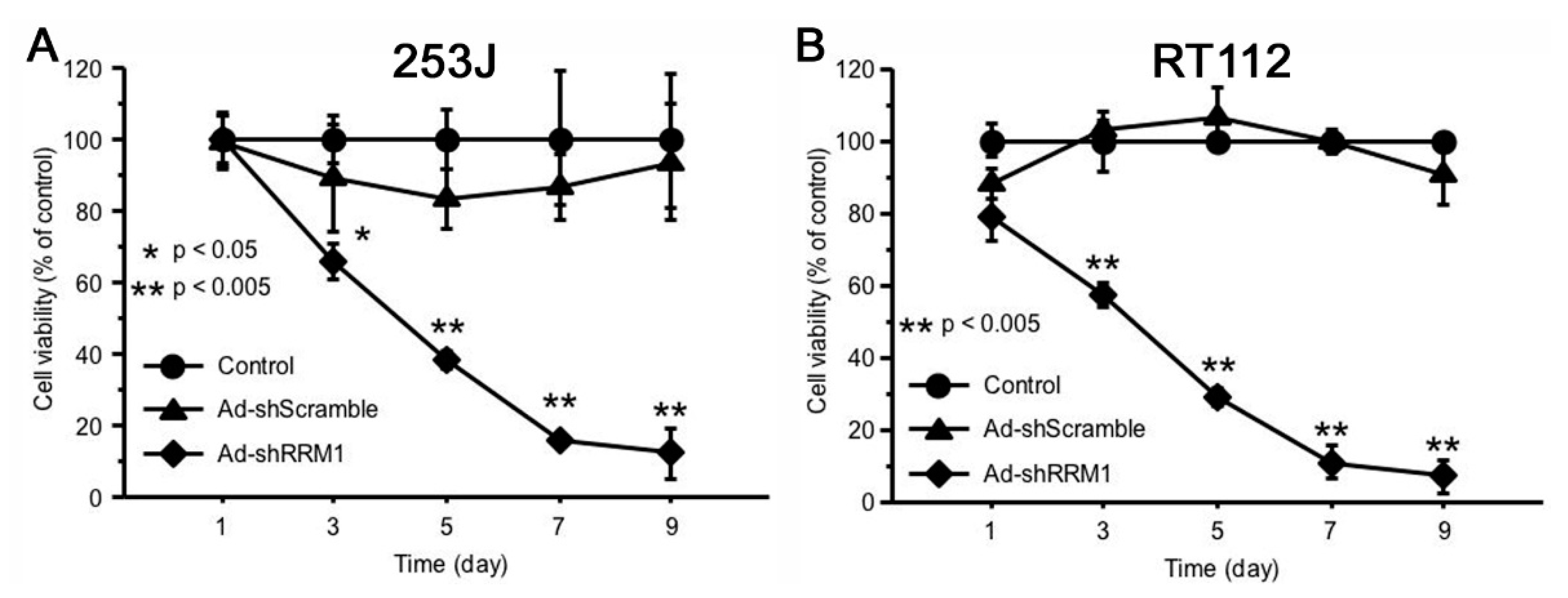
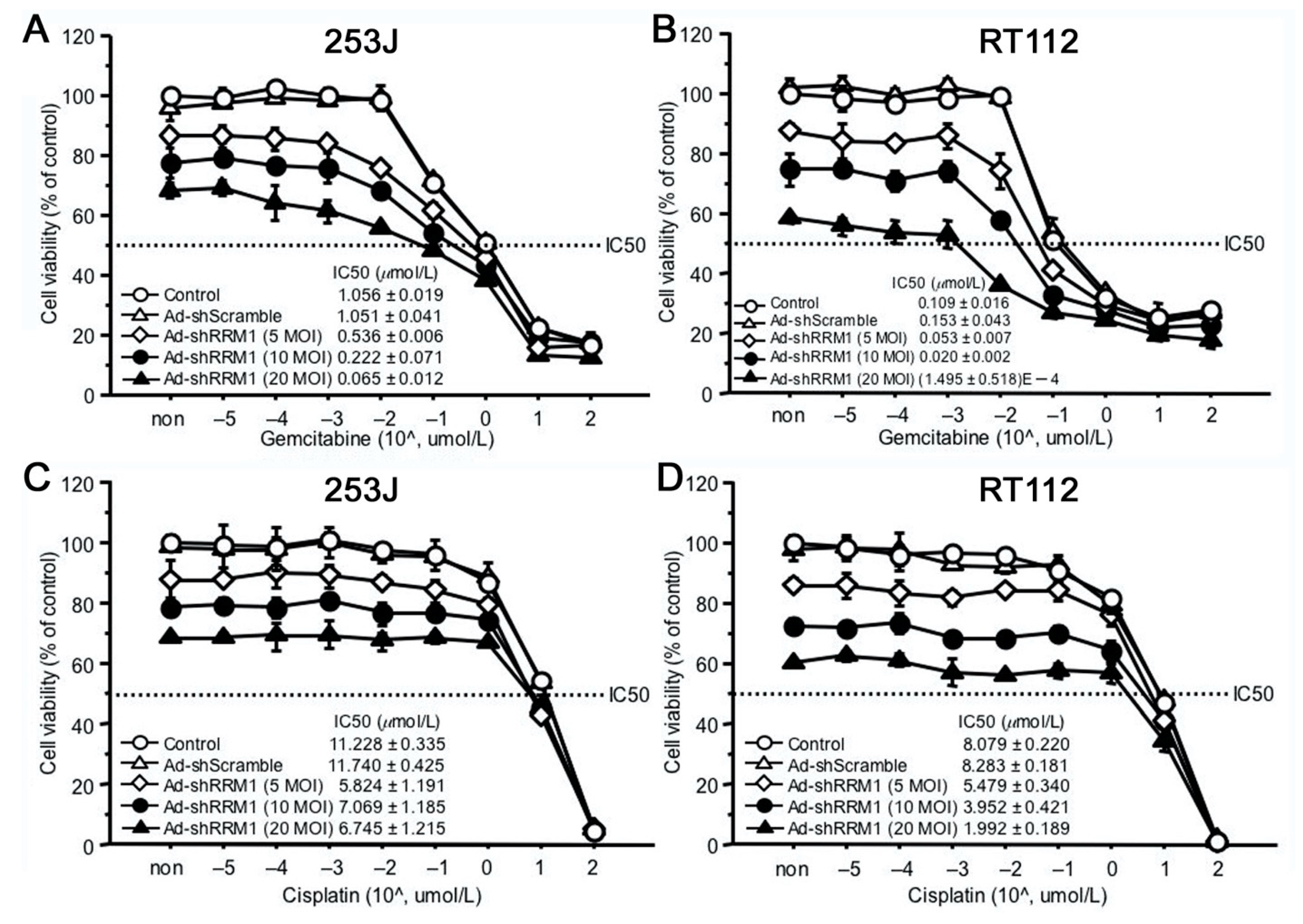
2.4. Ad-shRRM1 Inhibits Cell Viability in RRM1 Highly Expressed Bladder Cancer Cells
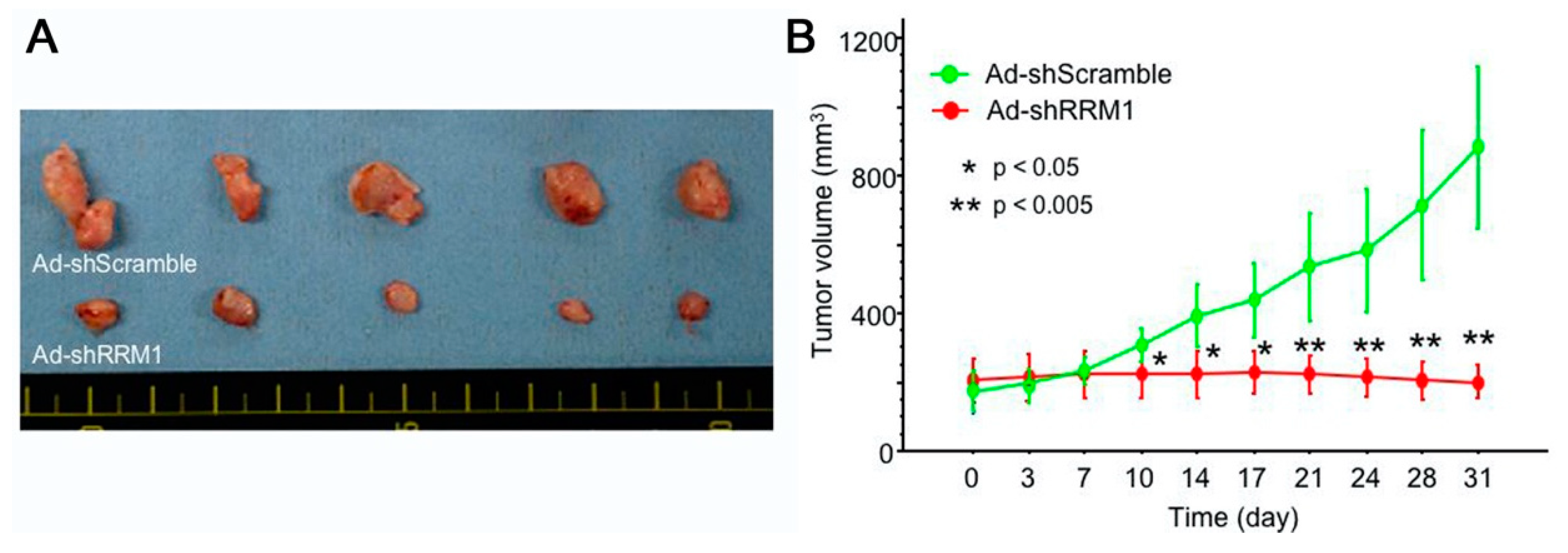
2.5. Ad-shRRM1 Inhibits Tumor Growth in Bladder Cancer Cells Xenografts
2.6. Ad-shRRM1 Specifically Increases GEM Sensitivity in Bladder Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Cell Lines
4.2. Real-Time RT-PCR for mRNA Expression
4.3. Western Blot Analysis for Protein Expression
4.4. Construction of Adenoviral Vectors
4.5. Cells Viability Assay
4.6. Cell Sensitivity to GEM
4.7. Xenograft Tumor Model in Nude Mice
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Abraham, J.; Gulley, J.L. The Bethesda Handbook of Clinical Oncology, 5th ed.; Wolters Kluwer Health: Philadelphia, PA, USA, 2019; ISBN 978-1-4963-4418-2. [Google Scholar]
- Alfred Witjes, J.; Lebret, T.; Compérat, E.; Cowan, N.; De Santis, M.; Bruins, H.; Hernández, V.; Espinós, E.; Dunn, J.; Rouanne, M.; et al. EAU guidelines on muscle-invasive and metastatic bladder cancer. Eur. Urol. 2017, 71, 462–475. [Google Scholar] [CrossRef] [PubMed]
- Stein, J.P.; Lieskovsky, G.; Cote, R.; Groshen, S.; Feng, A.C.; Boyd, S.; Skinner, E.; Bochner, B.; Thangathurai, D.; Mikhail, M.; et al. Radical cystectomy in the treatment of invasive bladder cancer: Long-term results in 1,054 patients. J. Clin. Oncol. 2001, 19, 666–675. [Google Scholar] [CrossRef] [PubMed]
- Gschwend, J.E.; Dahm, P.; Fair, W.R. Disease specific survival as endpoint of outcome for bladder cancer patients following radical cystectomy. Eur. Urol. 2002, 41, 440–448. [Google Scholar] [CrossRef]
- Wong, A.; Soo, R.A.; Yong, W.P.; Innocenti, F. Clinical pharmacology and pharmacogenetics of gemcitabine. Drug Metab. Rev. 2009, 41, 77–88. [Google Scholar] [CrossRef]
- Von der Maase, H.; Hansen, S.W.; Roberts, J.T.; Dogliotti, L.; Oliver, T.; Moore, M.J.; Bodrogi, I.; Albers, P.; Knuth, A.; Lippert, C.M.; et al. Gemcitabine and cisplatin versus methotrexate, vinblastine, doxorubicin, and cisplatin in advanced or metastatic bladder cancer: Results of a large, randomized, multinational, multicenter, phase III study. J. Clin. Oncol. 2000, 18, 3068–3077. [Google Scholar] [CrossRef] [PubMed]
- Pectasides, D.; Pectasides, M.; Economopoulos, T. Systemic chemotherapy in locally advanced and/or metastatic bladder cancer. Cancer Treat. Rev. 2006, 32, 456–470. [Google Scholar] [CrossRef] [PubMed]
- Elledge, S.J.; Zhou, Z.; Allen, J.B. Ribonucleotide reductase: Regulation, regulation, regulation. Trends Biochem. Sci. 1992, 17, 119–123. [Google Scholar] [CrossRef]
- Ohtaka, K.; Kohya, N.; Sato, K.; Kitajima, Y.; Ide, T.; Mitsuno, M.; Miyazaki, K. Ribonucleotide reductase subunit M1 is a possible chemoresistance marker to gemcitabine in biliary tract carcinoma. Oncol. Rep. 2008, 20, 279–286. [Google Scholar]
- Mini, E.; Nobili, S.; Caciagli, B.; Landini, I.; Mazzei, T. Cellular pharmacology of gemcitabine. Ann. Oncol. 2006, 17, 7–12. [Google Scholar] [CrossRef]
- Davidson, J.D.; Ma, L.; Flagella, M.; Geeganage, S.; Gelbert, L.M.; Slapak, C.A. An increase in the expression of ribonucleotide reductase large subunit 1 is associated with gemcitabine resistance in non-small cell lung cancer cell lines. Cancer Res. 2004, 64, 3761–3766. [Google Scholar] [CrossRef]
- Nakano, Y.; Tanno, S.; Koizumi, K.; Nishikawa, T.; Nakamura, K.; Minoguchi, M.; Izawa, T.; Mizukami, Y.; Okumura, T.; Kohgo, Y. Gemcitabine chemoresistance and molecular markers associated with gemcitabine transport and metabolism in human pancreatic cancer cells. Br. J. Cancer 2007, 96, 457–463. [Google Scholar] [CrossRef]
- Shilkrut, M.; Wu, A.; Thomas, D.G.; Hamstra, D.A. Expression of ribonucleoside reductase subunit M1, but not excision repair cross-complementation group 1, is predictive in muscle-invasive bladder cancer treated with chemotherapy and radiation. Mol. Clin. Oncol. 2014, 2, 479–487. [Google Scholar] [CrossRef]
- Kim, M.; Ku, J.H.; Kwak, C.; Kim, H.H.; Lee, E.; Keam, B.; Kim, T.M.; Heo, D.S.; Lee, S.H.; Moon, K.C. Predictive and prognostic value of ribonucleotide reductase regulatory subunit M1 and excision repair cross-complementation group 1 in advanced urothelial carcinoma (UC) treated with first-line gemcitabine plus platinum combination chemotherapy. PLoS ONE 2015, 10, e0133371. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, N.; Nakamura, Y.; Kohjimoto, Y.; Nishizawa, S.; Kikkawa, K.; Iba, A.; Kodama, Y.; Hara, I. Overexpression of ribonucleotide reductase subunit M1 protein predicts shorter survival in metastatic bladder cancer patients treated with gemcitabine-containing combination chemotherapy. Int. J. Urol. 2017, 24, 230–235. [Google Scholar] [CrossRef][Green Version]
- Tokunaga, Y.; Liu, D.; Nakano, J.; Zhang, X.; Nii, K.; Go, T.; Huang, C.L.; Yokomise, H. Potent effect of adenoviral vector expressing short hairpin RNA targeting ribonucleotide reductase large subunit M1 on cell viability and chemotherapeutic sensitivity to gemcitabine in non-small cell lung cancer cells. Eur. J. Cancer 2015, 51, 2480–2489. [Google Scholar] [CrossRef]
- Beck, B.; Chen, Y.-F.; Dere, W.; Devanarayan, V.; Eastwood, B.J.; Farmen, M.W.; Iturria, S.J.; Iversen, P.W.; Kahl, S.D.; Moore, R.A.; et al. Assay Operations for SAR Support; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bthesda, MA, USA, 2004. [Google Scholar]
- Parker, N.J.; Begley, C.G.; Fox, R.M. Human gene for the large subunit of ribonucleotide reductase (RRM1): Functional analysis of the promoter. Genomics 1995, 27, 280–285. [Google Scholar] [CrossRef] [PubMed]
- Nordlund, P.; Reichard, P. Ribonucleotide reductases. Annu. Rev. Biochem. 2006, 75, 681–706. [Google Scholar] [CrossRef]
- Jordheim, L.P.; Sève, P.; Trédan, O.; Dumontet, C. The ribonucleotide reductase large subunit (RRM1) as a predictive factor in patients with cancer. Lancet Oncol. 2011, 12, 693–702. [Google Scholar] [CrossRef]
- Reid, G.; ’t Wallant, N.C.; Patel, R.; Antonic, A.; Saxon-Aliifaalogo, F.; Cao, H.; Webster, G.; Watson, J.D. Potent subunit-specific effects on cell growth and drug sensitivity from optimised siRNA-mediated silencing of ribonucleotide reductase. J. RNAi Gene Silenc. Int. J. RNA Gene Target. Res. 2009, 5, 321–330. [Google Scholar]
- Sagawa, M.; Ohguchi, H.; Harada, T.; Samur, M.K.; Tai, Y.T.; Munshi, N.C.; Kizaki, M.; Hideshima, T.; Anderson, K.C. Ribonucleotide reductase catalytic Subunit M1 (RRM1) as a novel therapeutic target in multiple myeloma. Clin. Cancer Res. 2017, 23, 5225–5237. [Google Scholar] [CrossRef] [PubMed]
- Schiller, J.H.; Harrington, D.; Belani, C.P.; Langer, C.; Sandler, A.; Krook, J.; Zhu, J.; Johnson, D.H.; Eastern Cooperative Oncology Group. Comparison of four chemotherapy regimens for advanced non-small-cell lung cancer. N. Engl. J. Med. 2002, 346, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Spielmann, M.; Llombart-Cussac, A.; Kalla, S.; Espié, M.; Namer, M.; Ferrero, J.M.; Diéras, V.; Fumoleau, P.; Cuvier, C.; Perrocheau, G.; et al. Single-agent gemcitabine is active in previously treated metastatic breast cancer. Oncology 2001, 60, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Oettle, H.; Post, S.; Neuhaus, P.; Gellert, K.; Langrehr, J.; Ridwelski, K.; Schramm, H.; Fahlke, J.; Zuelke, C.; Burkart, C.; et al. Adjuvant chemotherapy with gemcitabine vs observation in patients undergoing curative-intent resection of pancreatic cancer: A randomized controlled trial. J. Am. Med. Assoc. 2007, 297, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Witjes, J.A.; van der Heijden, A.G.; Vriesema, J.L.J.; Peters, G.J.; Laan, A.; Schalken, J.A. Intravesical gemcitabine: A phase 1 and pharmacokinetic study. Eur. Urol. 2004, 45, 182–186. [Google Scholar] [CrossRef]
- Yafi, F.A.; North, S.; Kassouf, W. First- and second-line therapy for metastatic urothelial carcinoma of the bladder. Curr. Oncol. 2011, 18, e25–e34. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.I.; Choo, S.H. Intravesical Chemotherapy. In Bladder Cancer; Ku, J.H., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 263–276. ISBN 9780128099407. [Google Scholar]
- Muramaki, M.; So, A.; Hayashi, N.; Sowery, R.; Miyake, H.; Fujisawa, M.; Gleave, M.E. Chemosensitization of gemcitabine-resistant human bladder cancer cell line both in vitro and in vivo using antisense oligonucleotide targeting the anti-apoptotic gene, clusterin. BJU Int. 2009, 103, 384–390. [Google Scholar] [CrossRef]
- Crystal, R.G. Adenovirus: The first effective in vivo gene delivery vector. Hum. Gene Ther. 2014, 25, 3–11. [Google Scholar] [CrossRef]
- Lee, C.S.; Bishop, E.S.; Zhang, R.; Yu, X.; Farina, E.M.; Yan, S.; Zhao, C.; Zheng, Z.; Shu, Y.; Wu, X.; et al. Adenovirus-mediated gene delivery: Potential applications for gene and cell-based therapies in the New Era of personalized medicine. Genes Dis. 2017, 4, 43–63. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, D.; Hayashida, Y.; Okazoe, H.; Hashimoto, T.; Ueda, N.; Sugimoto, M.; Kakehi, Y. G Protein-coupled Receptor 87 (GPR87) promotes cell proliferation in human bladder cancer cells. Int. J. Mol. Sci. 2015, 16, 24319–24331. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Taoka, R.; Liu, D.; Matsuoka, Y.; Tohi, Y.; Kakehi, Y.; Sugimoto, M. Knockdown of RRM1 with Adenoviral shRNA Vectors to Inhibit Tumor Cell Viability and Increase Chemotherapeutic Sensitivity to Gemcitabine in Bladder Cancer Cells. Int. J. Mol. Sci. 2021, 22, 4102. https://doi.org/10.3390/ijms22084102
Zhang X, Taoka R, Liu D, Matsuoka Y, Tohi Y, Kakehi Y, Sugimoto M. Knockdown of RRM1 with Adenoviral shRNA Vectors to Inhibit Tumor Cell Viability and Increase Chemotherapeutic Sensitivity to Gemcitabine in Bladder Cancer Cells. International Journal of Molecular Sciences. 2021; 22(8):4102. https://doi.org/10.3390/ijms22084102
Chicago/Turabian StyleZhang, Xia, Rikiya Taoka, Dage Liu, Yuki Matsuoka, Yoichiro Tohi, Yoshiyuki Kakehi, and Mikio Sugimoto. 2021. "Knockdown of RRM1 with Adenoviral shRNA Vectors to Inhibit Tumor Cell Viability and Increase Chemotherapeutic Sensitivity to Gemcitabine in Bladder Cancer Cells" International Journal of Molecular Sciences 22, no. 8: 4102. https://doi.org/10.3390/ijms22084102
APA StyleZhang, X., Taoka, R., Liu, D., Matsuoka, Y., Tohi, Y., Kakehi, Y., & Sugimoto, M. (2021). Knockdown of RRM1 with Adenoviral shRNA Vectors to Inhibit Tumor Cell Viability and Increase Chemotherapeutic Sensitivity to Gemcitabine in Bladder Cancer Cells. International Journal of Molecular Sciences, 22(8), 4102. https://doi.org/10.3390/ijms22084102