
Diosmin Mitigates Cyclophosphamide Induced Premature Ovarian Insufficiency in Rat Model

 , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Effect of Cyclophosphamide on Ovarian Morphology
2.2. Chemotherapy-Induced Serum Hormonal Level Changes in the POI Model
2.3. Oxidative Stress Markers in the POI Model
2.4. Effect of Cyclophosphamide and Diosmin on Follicular Growth
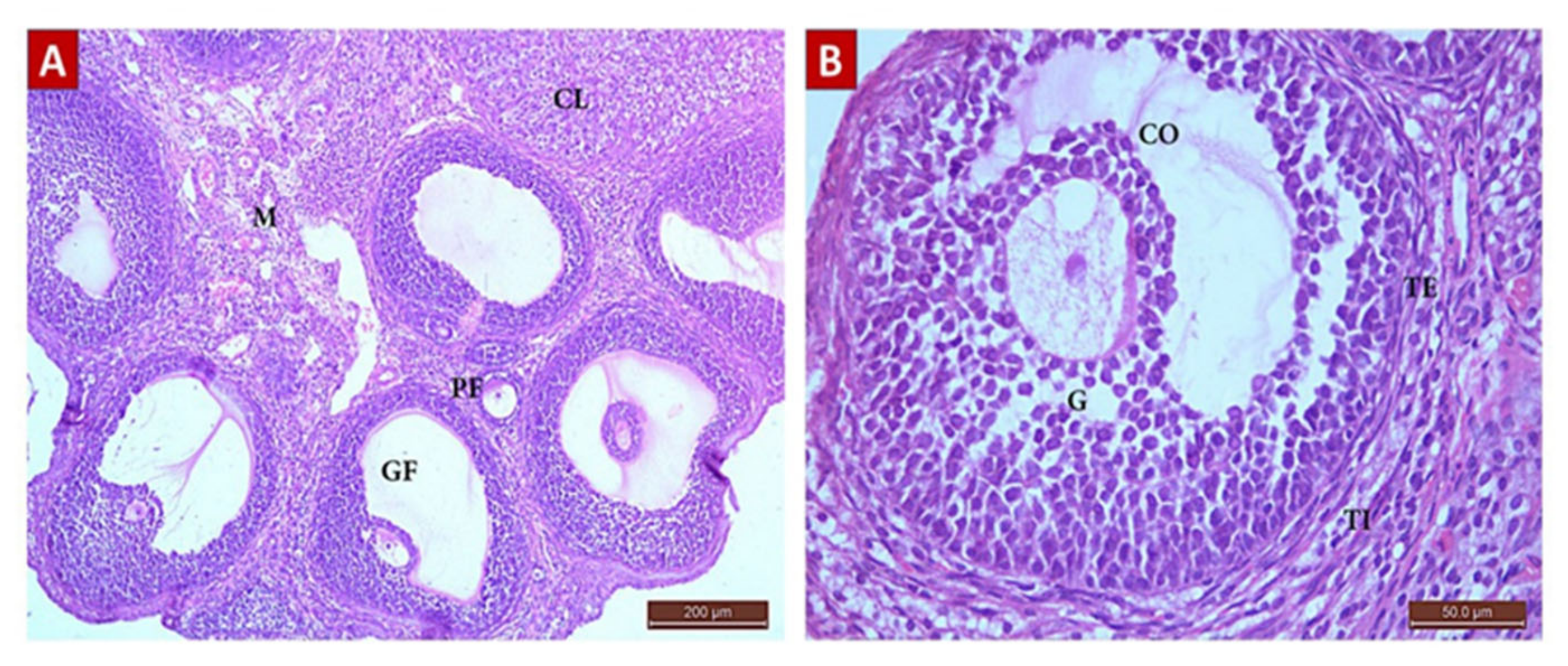
2.4.1. Hematoxylin and Eosin staining results
The Control Group (Group I, Figure 1, Figure 2, Figure 3 and Figure 4)
POI Group (Group II, Figure 5)
Diosmin 50 (Group III, Figure 6)
Diosmin 100 (Group IV, Figure 7)
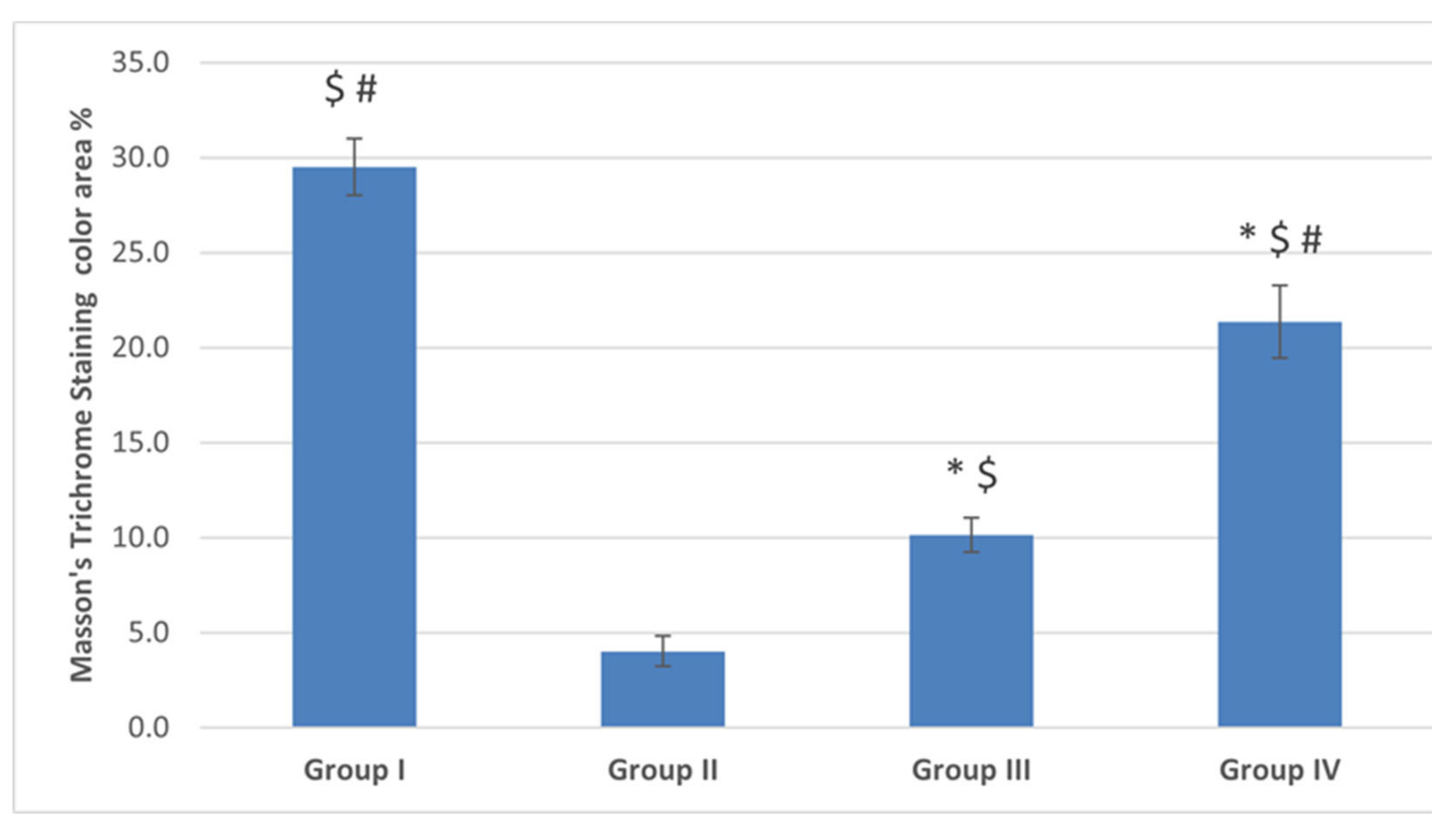
2.4.2. Masson’s Trichrome Staining Results
The control (G I) Group
The POI (G II) Group
Diosmin 50 (Group III) and Diosmin 100 (Group IV) Groups
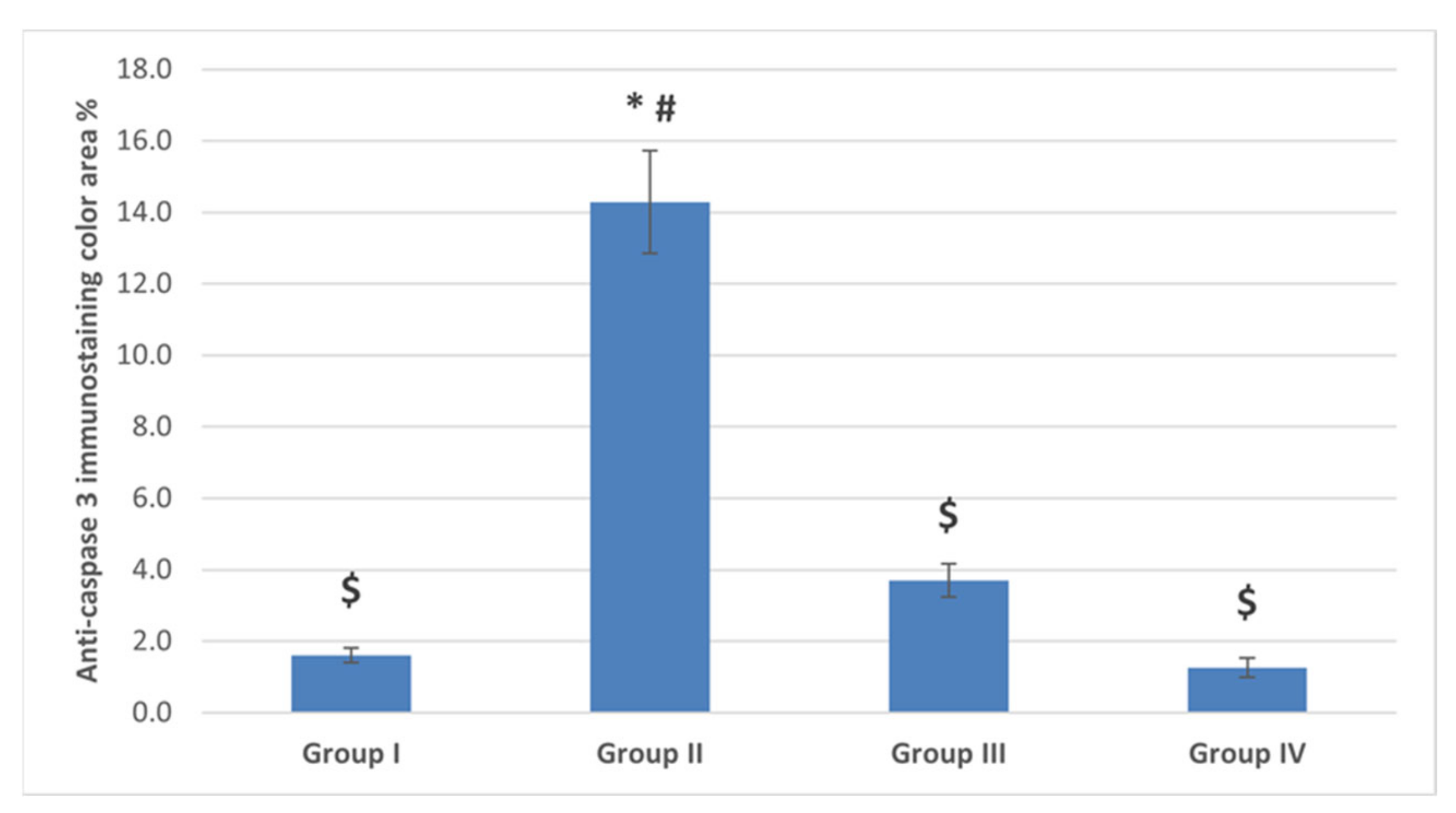
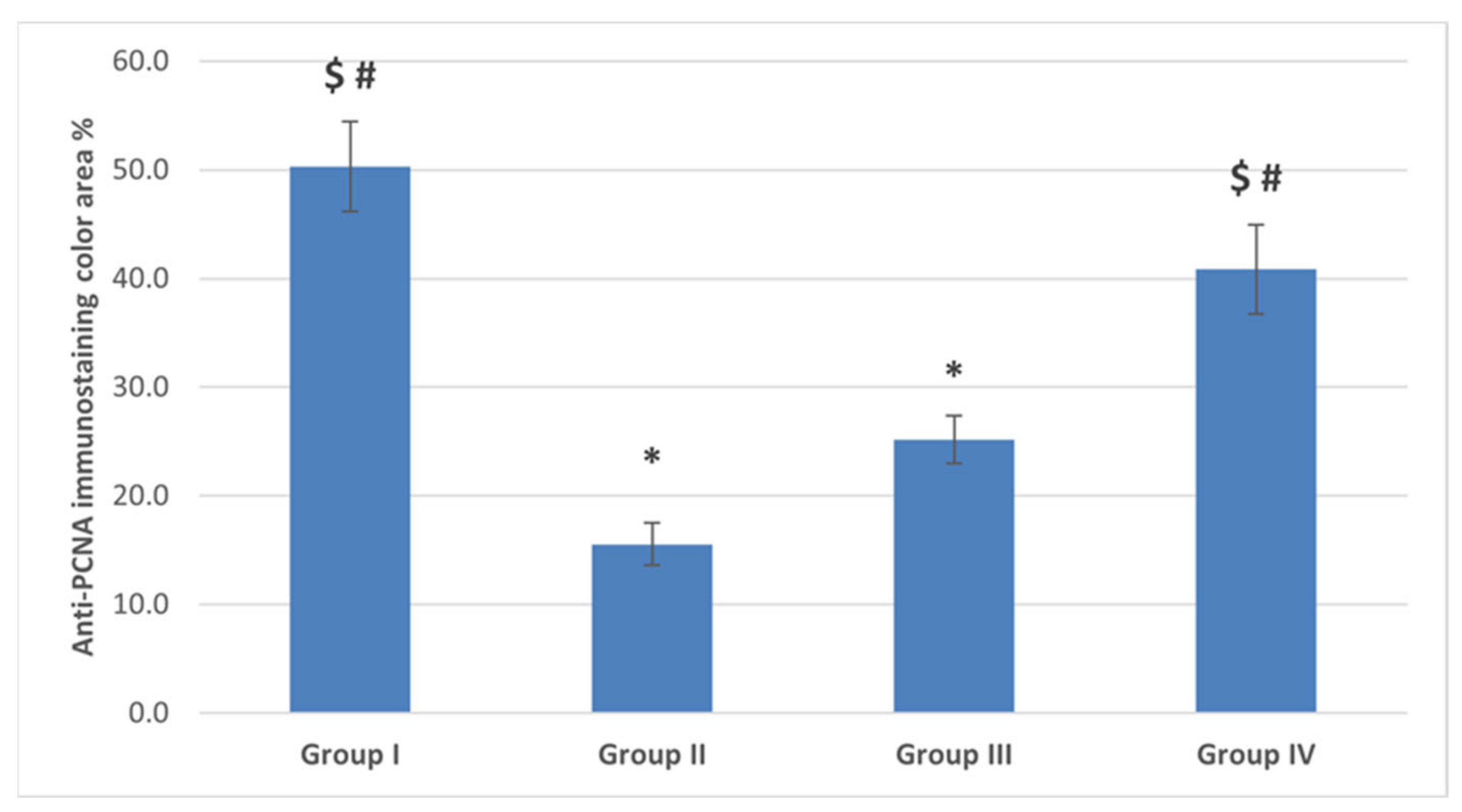
2.4.3. Immunostaining Results
The Anti-Caspase 3
The Anti-PCNA
2.5. Cyclophosphamide or Diosmin Effect on MiRNA-145 and Target Genes
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Chemicals
4.3. Experimental Model of Premature Ovarian Insufficiency
4.4. Study Design
4.5. Histopathological Examination
4.6. Immunohistochemistry Staining
4.7. Serum Biochemical Analysis
4.7.1. Oxidative Stress Markers
4.7.2. Hormonal Assays for FSH, E2, and AMH
4.8. RNA Isolation, Reverse Transcription, and Target Gene Expression
4.9. microRNA-145 Expression Analysis
4.10. In Silico Data Analysis
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Weghofer, A.; Gleicher, N. Ovarian function: A theory of relativity. Hum. Reprod. 2009, 24, 17–19. [Google Scholar] [CrossRef]
- Hsueh, A.J.W.; Kawamura, K.; Cheng, Y.; Fauser, B.C.J.M. Intraovarian control of early folliculogenesis. Endocr. Rev. 2015, 36, 1–24. [Google Scholar] [CrossRef]
- Liu, Y.; Fang, Y.; Yang, X. MicroRNAs in ovarian function and disorders. J. Ovarian Res. 2015, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Asselin, E.; Xiao, C.W.; Wang, Y.F.; Tsang, B.K. Mammalian follicular development and atresia: Role of apoptosis. Neurosignals 2000, 9, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Tilly, J.L.; Kowalski, K.I.; Johnson, A.L.; Hsueh, A.J. Involvement of apoptosis in ovarian follicular atresia and postovulatory regression. Endocrinology 1991, 129, 2799–2801. [Google Scholar] [CrossRef] [PubMed]
- Valdez, K.E.; Cuneo, S.P.; Turzillo, A.M. Regulation of apoptosis in the atresia of dominant bovine follicles of the first follicular wave following ovulation. Reproduction 2005, 130, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Saleh, R.A.; Bedaiwy, M.A. Role of reactive oxygen species in the pathophysiology of human reproduction. Fertil. Steril. 2003, 79, 829–843. [Google Scholar] [CrossRef]
- Green, D.M.; Nolan, V.G.; Goodman, P.J.; Whitton, J.A.; Srivastava, D.; Leisenring, W.M.; Neglia, J.P.; Sklar, C.A.; Kaste, S.C.; Hudson, M.M.; et al. The cyclophosphamide equivalent dose as an approach for quantifying alkylating agent exposure: A report from the Childhood Cancer Survivor Study. Pediatric Blood Cancer 2014, 61, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Sheweita, S.A.; El-Hosseiny, L.S.; Nashashibi, M.A. Protective effects of essential oils as natural antioxidants against hepatotoxicity induced by cyclophosphamide in mice. PLoS ONE 2016, 11, e0165667. [Google Scholar] [CrossRef] [PubMed]
- Murali, V.P.; Kuttan, G. Curculigo orchioides gaertn effectively ameliorates the uro-and nephrotoxicities induced by cyclophosphamide administration in experimental animals. Integr. Cancer Ther. 2016, 15, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Kumar, A. Protective Effect of Edaravone on Cyclophosphamide Induced Oxidative Stress and Neurotoxicity in Rats. Curr. Drug Saf. 2019, 14, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Aly, F.M.; Othman, A.; Haridy, M.A. Protective effects of fullerene C60 nanoparticles and virgin olive oil against genotoxicity induced by cyclophosphamide in rats. Oxidative Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef] [PubMed]
- Tsai-Turton, M.; Luong, B.T.; Tan, Y.; Luderer, U. Cyclophosphamide-induced apoptosis in COV434 human granulosa cells involves oxidative stress and glutathione depletion. Toxicol. Sci. 2007, 98, 216–230. [Google Scholar] [CrossRef] [PubMed]
- Nogata, Y.; Sakamoto, K.; Shiratsuchi, H.; Ishii, T.; Yano, M.; Ohta, H. Flavonoid composition of fruit tissues of citrus species. Biosci. Biotechnol. Biochem. 2006, 70, 178–192. [Google Scholar] [CrossRef]
- Pari, L.; Srinivasan, S. Antihyperglycemic effect of diosmin on hepatic key enzymes of carbohydrate metabolism in streptozotocin-nicotinamide-induced diabetic rats. Biomed. Pharmacother. 2010, 64, 477–481. [Google Scholar] [CrossRef]
- Maksimovic, Z.V.; Maksimovic, M.; Jadranin, D.; Kuzmanovic, I.; Andonovic, O. Medicamentous treatment of chronic venous insufficiency using semisynthetic diosmin: A prospective study. Acta Chir. Iugosl. 2008, 55, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Firuzi, O.; Miri, R.; Tavakkoli, M.; Saso, L. Antioxidant therapy: Current status and future prospects. Curr. Med. Chem. 2011, 18, 3871–3888. [Google Scholar] [CrossRef] [PubMed]
- Delbarre, B.; Delbarre, G.; Calinon, F. Effect of Daflon 500 mg, a flavonoid drug, on neurological signs, levels of free radicals and electroretinogram in the gerbil after ischemia-reperfusion injury. Int. J. Microcirc. 1995, 15, 27–33. [Google Scholar] [CrossRef]
- Ramelet, A.-A. Pharmacologic aspects of a phlebotropic drug in CVI-associated edema. Angiology 2000, 51, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Manthey, J.A. Biological properties of flavonoids pertaining to inflammation. Microcirculation 2000, 7, S29–S34. [Google Scholar] [CrossRef] [PubMed]
- Rapavi, E.; Kocsis, I.; Fehér, E.; Szentmihályi, K.; Lugasi, A.; Székely, E.; Blázovics, A. The effect of citrus flavonoids on the redox state of alimentary-induced fatty liver in rats. Nat. Prod. Res. 2007, 21, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Labrid, C. Pharmacologic properties of Daflon 500 mg. Angiology 1994, 45, 45. [Google Scholar]
- Bartel, D.P. MicroRNAs: Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef]
- Shukla, G.C.; Singh, J.; Barik, S. MicroRNAs: Processing, maturation, target recognition and regulatory functions. Mol. Cell. Pharmacol. 2011, 3, 83. [Google Scholar] [PubMed]
- Li, R.; Yan, G.; Li, Q.; Sun, H.; Hu, Y.; Sun, J.; Xu, B. MicroRNA-145 protects cardiomyocytes against hydrogen peroxide (H 2 O 2)-induced apoptosis through targeting the mitochondria apoptotic pathway. PLoS ONE 2012, 7, e44907. [Google Scholar] [CrossRef]
- Lin, Y.; Liu, X.; Cheng, Y.; Yang, J.; Huo, Y.; Zhang, C. Involvement of MicroRNAs in hydrogen peroxide-mediated gene regulation and cellular injury response in vascular smooth muscle cells. J. Biol. Chem. 2009, 284, 7903–7913. [Google Scholar] [CrossRef]
- Hou, Y.; Wang, Y.; Xu, S.; Qi, G.; Wu, X. Bioinformatics identification of microRNAs involved in polycystic ovary syndrome based on microarray data. Mol. Med. Rep. 2019, 20, 281–291. [Google Scholar] [CrossRef]
- Rehman, M.U.; Tahir, M.; Khan, A.Q.; Khan, R.; Lateef, A.; Hamiza, O.O.; Ali, F.; Sultana, S. Diosmin protects against trichloroethylene-induced renal injury in Wistar rats: Plausible role of p53, Bax and caspases. Br. J. Nutr. 2013, 110, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Tahir, M.; Rehman, M.U.; Lateef, A.; Khan, R.; Khan, A.Q.; Qamar, W.; Ali, F.; O’Hamiza, O.; Sultana, S. Diosmin protects against ethanol-induced hepatic injury via alleviation of inflammation and regulation of TNF-α and NF-κB activation. Alcohol 2013, 47, 131–139. [Google Scholar] [CrossRef]
- Germoush, M. Diosmin Protects Against Cyclophosphamide-induced Liver Injury Through Attenuation of Oxidative Stress, Inflammation and Apoptosis. Int. J. Pharmacol. 2016, 12, 644–654. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Khalifa, H.A.; Abushouk, A.I.; Dkhil, M.A.; Al-Quraishy, S.A. Diosmin Attenuates Methotrexate-Induced Hepatic, Renal, and Cardiac Injury: A Biochemical and Histopathological Study in Mice. Oxidative Med. Cell. Longev. 2017, 2017, 1–10. [Google Scholar] [CrossRef]
- Pascuali, N.; Scotti, L.; Di Pietro, M.; Oubiña, G.; Bas, D.; May, M.; Muñoz, A.G.; Cuasnicú, P.S.; Cohen, D.J.; Tesone, M.; et al. Ceramide-1-phosphate has protective properties against cyclophosphamide-induced ovarian damage in a mice model of premature ovarian failure. Hum. Reprod. 2018, 33, 844–859. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Zhong, Y.; Qian, C.; Zou, Q.; Ou, J.; Shi, Y.; Gao, L.; Wang, G.; Liu, Z.; Li, H.; et al. Human umbilical cord mesenchymal stem cells therapy in cyclophosphamide-induced premature ovarian failure rat model. BioMed Res. Int. 2016, 2016. [Google Scholar] [CrossRef]
- Hamzeh, M.; Hosseinimehr, S.J.; Mohammadi, H.R.; Beklar, S.Y.; Dashti, A.; Amiri, F.T. Atorvastatin attenuates the ovarian damage induced by cyclophosphamide in rat: An experimental study. Int. J. Reprod. Biomed. 2018, 16, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Yener, N.A.; Sinanoğlu, O.; Ilter, E.; Celik, A.; Sezgin, G.; Midi, A.; Deveci, U.; Aksungar, F. Effects of Spirulina on Cyclophosphamide-Induced Ovarian Toxicity in Rats: Biochemical and Histomorphometric Evaluation of the Ovary. Biochem. Res. Int. 2013, 2013, 764262. [Google Scholar] [CrossRef]
- Melekoglu, R.; Ciftci, O.; Eraslan, S.; Cetin, A.; Basak, N. Beneficial effects of curcumin and capsaicin on cyclophosphamide-induced premature ovarian failure in a rat model. J. Ovarian Res. 2018, 11, 1–8. [Google Scholar] [CrossRef]
- Liu, T.; Wang, S.; Li, Q.; Huang, Y.; Chen, C.; Zheng, J. Telocytes as potential targets in a cyclophosphamide-induced animal model of premature ovarian failure. Mol. Med. Rep. 2016, 14, 2415–2422. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Liu, T.; Wang, S.; Chi, H.; Chen, C.; Zheng, J. Cyclophosphamide promotes the proliferation inhibition of mouse ovarian granulosa cells and premature ovarian failure by activating the lncRNA-Meg3-p53-p66Shc pathway. Gene 2017, 596, 120138. [Google Scholar] [CrossRef]
- Liu, W.Y.; Liou, S.-S.; Hong, T.-Y.; Liu, I.M. The Benefits of the Citrus Flavonoid Diosmin on Human Retinal Pigment Epithelial Cells under High-Glucose Conditions. Molecules 2017, 22, 2251. [Google Scholar] [CrossRef]
- Queenthy, S.S.; Prince, P.S.M.; John, B. Diosmin Prevents Isoproterenol-Induced Heart Mitochondrial Oxidative Stress in Rats. Cardiovasc. Toxicol. 2018, 18, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Shalkami, A.S.; Hassan, M.I.A.; Bakr, A.G. Anti-inflammatory, antioxidant and anti-apoptotic activity of diosmin in acetic acid-induced ulcerative colitis. Hum. Exp. Toxicol. 2017, 37, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Yacobi, K.; Wojtowicz, A.K.; Tsafriri, A.; Gross, A. Gonadotropins Enhance Caspase-3 and -7 Activity and Apoptosis in the Theca-Interstitial Cells of Rat Preovulatory Follicles in Culture. Endocrinology 2004, 145, 1943–1951. [Google Scholar] [CrossRef]
- Weenen, C.; Laven, J.S.; Von Bergh, A.R.; Cranfield, M.; Groome, N.P.; Visser, J.A.; Kramer, P.; Fauser, B.C.; Themmen, A.P. Anti-Mullerian hormone expression pattern in the human ovary: Potential implications for initial and cyclic follicle recruitment. Mol. Hum. Reprod. 2004, 10, 77–83. [Google Scholar] [CrossRef]
- Visser, J.; Schipper, I.; Laven, J.; Themmen, A. Anti-Müllerian hormone: An ovarian reserve marker in primary ovarian insufficiency. Nat. Rev. Endocrinol. 2012, 8, 331–341. [Google Scholar] [CrossRef]
- Kwee, J.; Schats, R.; McDonnell, J.; Themmen, A.; Jong, F.; Lambalk, C.B. Evaluation of AMH as test for the prediction of ovarian reserve. Fertil. Steril. 2007, 90, 737–743. [Google Scholar] [CrossRef]
- Sukur, Y.E.; Kivancli, I.B.; Ozmen, B. Ovarian aging and premature ovarian failure. J. Turk. Gynecol. Assoc. 2014, 15, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Cai, G.; Ma, X.; Chen, B.; Huang, Y.; Liu, S.; Yang, H.; Zou, W. MicroRNA-145 Negatively Regulates Cell Proliferation Through Targeting IRS1 in Isolated Ovarian Granulosa Cells From Patients With Polycystic Ovary Syndrome. Reprod. Sci. 2016, 24, 902–910. [Google Scholar] [CrossRef]
- Ding, W.; Tan, H.; Zhao, C.; Li, X.; Li, Z.; Jiang, C.; Zhang, Y.; Wang, L. MiR-145 suppresses cell proliferation and motility by inhibiting ROCK1 in hepatocellular carcinoma. Tumor Biol. 2015, 37, 6255–6260. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Zhao, L.; Yang, F.; Li, Z.; Li, G. MicroRNA-145 inhibits the malignant phenotypes of gastric carcinoma cells via downregulation of fascin 1 expression. Mol. Med. Rep. 2015, 13, 1033–1039. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zheng, M.; Sun, X.; Li, Y.; Zuo, W. MicroRNA-145 inhibits growth and migration of breast cancer cells through targeting oncoprotein ROCK1. Tumor Biol. 2016, 37, 8189–8196. [Google Scholar] [CrossRef] [PubMed]
- Kenific, C.M.; Debnath, J. Cellular and metabolic functions for autophagy in cancer cells. Trends Cell Biol. 2015, 25, 37–45. [Google Scholar] [CrossRef]
- Lu, Y.; Chopp, M.; Zheng, X.; Katakowski, M.; Buller, B.; Jiang, F. MiR-145 reduces ADAM17 expression and inhibits in vitro migration and invasion of glioma cells. Oncol. Rep. 2013, 29, 67–72. [Google Scholar] [CrossRef]
- Lin, Y.; Ge, X.; Wen, Y.; Shi, Z.-M.; Chen, Q.-D.; Wang, M.; Liu, L.-Z.; Jiang, B.-H.; Lu, Y. MiRNA-145 increases therapeutic sensibility to gemcitabine treatment of pancreatic adenocarcinoma cells. Oncotarget 2016, 7, 70857–70868. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.M.; Sohel, M.M.H.; Schellander, K.; Tesfaye, D. Characterization and importance of microRNAs in mammalian gonadal functions. Cell Tissue Res. 2012, 349, 679–690. [Google Scholar] [CrossRef]
- Hossain, M.; Ghanem, N.; Hoelker, M.; Rings, F.; Phatsara, C.; Tholen, E.; Schellander, K.; Tesfaye, D. Identification and characterization of miRNAs expressed in the bovine ovary. BMC Genom. 2009, 10, 443. [Google Scholar] [CrossRef]
- Feldo, M.; Wójciak-Kosior, M.; Sowa, I.; Kocki, J.; Bogucki, J.; Zubilewicz, T.; Kęsik, J.; Bogucka-Kocka, A. Effect of Diosmin Administration in Patients with Chronic Venous Disorders on Selected Factors Affecting Angiogenesis. Moleculers 2019, 24, 3316. [Google Scholar] [CrossRef] [PubMed]
- Imam, F.; Al-Harbi, N.O.; Al-Harbi, M.M.; Ansari, M.A.; Zoheir, K.M.A.; Iqbal, M.; AnweR, K.; Hoshani, A.R.A.; Attia, S.M.; Ahmad, S.F. Diosmin downregulates the expression of T cell receptors, pro-inflammatory cytokines and NF-κB activation against LPS-induced acute lung injury in mice. Pharmacol. Res. 2015, 102, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Yagi, K. Assay for serum lipid peroxide level and its clinical significance. Lipid Peroxides Biol. Med. 1982, 223, 242. [Google Scholar]
- Aebi, H. Catalase in vitro. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1984; pp. 121–126. [Google Scholar]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Sticht, C.; De La Torre, C.; Parveen, A.; Gretz, N. miRWalk: An online resource for prediction of microRNA binding sites. PLOS ONE 2018, 13, e0206239. [Google Scholar] [CrossRef] [PubMed]
- Coordinators, N.R. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2016, 44, D7–D19. [Google Scholar] [CrossRef]
- The UniProt, C. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2018, 47, D506–D515. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [PubMed]
- Millán, R.M.S.; Martínez-Ballesteros, I.; Rementeria, A.; Garaizar, J.; Bikandi, J. Online exercise for the design and simulation of PCR and PCR-RFLP experiments. BMC Res. Notes 2013, 6, 513. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Buchner, A.; Lang, A.-G. Statistical power analyses using G*Power 3.1: Tests for correlation and regression analyses. Behav. Res. Methods 2009, 41, 1149–1160. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Body Weight (gm) | Ovarian Weight (gm) |
|---|---|---|
| G I | 139 ± 15 | 18 ± 5 |
| G II | 130 ± 10 | 8 ± 2 * |
| G III | 132 ± 22 | 10 ± 5 *,$ |
| G IV | 134 ± 15 | 15 ± 3 $ |
| Groups | FSH Level (mIU/mL) | AMH (mIU/mL) | E2 Level (pg/mL) |
|---|---|---|---|
| G I | 0.015 ±0.007 | 0.330 ± 0.148 | 18.35 ± 1.77 |
| G II | 0.052 ± 0.031 * | 0.036 ± 0.017 * | 8.53 ± 1.02* |
| G III | 0.003 ± 0.003 *,$ | 0.237 ± 0.124 *,$ | 12.15 ± 2.51 *,$ |
| G IV | 0.011 ± 0.007 $,# | 0.284 ± 0.120 *,$ | 14.68 ± 3.87 *,$ |
| Groups | MDA Level (nmol/mL) | Catalase Activity (U/L) |
|---|---|---|
| G I | 9.08 ± 0.16 $ | 78.78 ± 1.98 |
| G II | 20.17 ± 7.91 * | 59.85 ± 20.08 |
| G III | 4.68 ± 1.35 $ | 83.44 ± 5.37 |
| G IV | 6.18 ± 1. 35 $ | 86.35 ± 2.07 |
| Groups | Primordial | Primary | Antral | Graafian | Atretic | Total |
|---|---|---|---|---|---|---|
| G I | 2.6 ± 1.34 | 3.8 ± 1.47 | 2.5 ± 0.52 | 1.5 ± 0.52 | 0.6 ± 0.51 | 11 |
| G II | 0.5 ± 0.52 * | 0.5 ± 0.52 * | 0.4 ± 0.41 * | 0.2 ± 0.22 * | 2.8 ± 0.63 * | 4.4 |
| G III | 2 ± 0.86 | 2.5 ± 0.84 | 1.5 ± 0.7 | 0.7 ± 0.48 | 1 ± 0.8 | 7.7 |
| G IV | 3 ± 1.41 | 3.2 ± 1.31 | 1.7 ± 0.81 | 1.4 ± 0.51 $ | 0.7 ± 0.67 | 10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abogresha, N.M.; Mohammed, S.S.; Hosny, M.M.; Abdallah, H.Y.; Gadallah, A.M.; Greish, S.M. Diosmin Mitigates Cyclophosphamide Induced Premature Ovarian Insufficiency in Rat Model. Int. J. Mol. Sci. 2021, 22, 3044. https://doi.org/10.3390/ijms22063044
Abogresha NM, Mohammed SS, Hosny MM, Abdallah HY, Gadallah AM, Greish SM. Diosmin Mitigates Cyclophosphamide Induced Premature Ovarian Insufficiency in Rat Model. International Journal of Molecular Sciences. 2021; 22(6):3044. https://doi.org/10.3390/ijms22063044
Chicago/Turabian StyleAbogresha, Noha M., Sally S. Mohammed, Marwa M. Hosny, Hoda Y. Abdallah, Ahmed M. Gadallah, and Sahar M. Greish. 2021. "Diosmin Mitigates Cyclophosphamide Induced Premature Ovarian Insufficiency in Rat Model" International Journal of Molecular Sciences 22, no. 6: 3044. https://doi.org/10.3390/ijms22063044
APA StyleAbogresha, N. M., Mohammed, S. S., Hosny, M. M., Abdallah, H. Y., Gadallah, A. M., & Greish, S. M. (2021). Diosmin Mitigates Cyclophosphamide Induced Premature Ovarian Insufficiency in Rat Model. International Journal of Molecular Sciences, 22(6), 3044. https://doi.org/10.3390/ijms22063044