
3D Structural Insights into β-Glucans and Their Binding Proteins


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
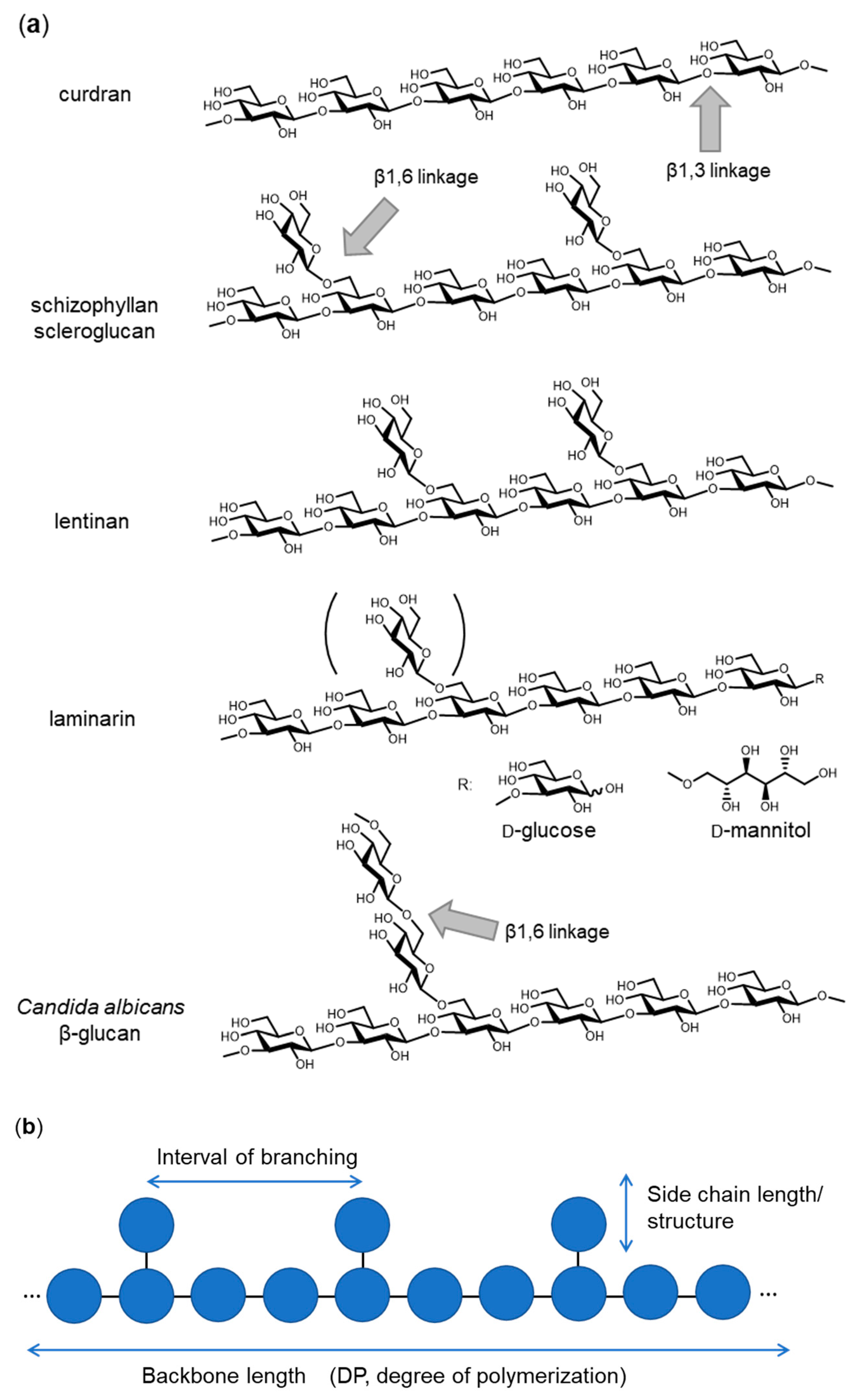
1. Introduction—Variation of β-Glucan Primary Structures
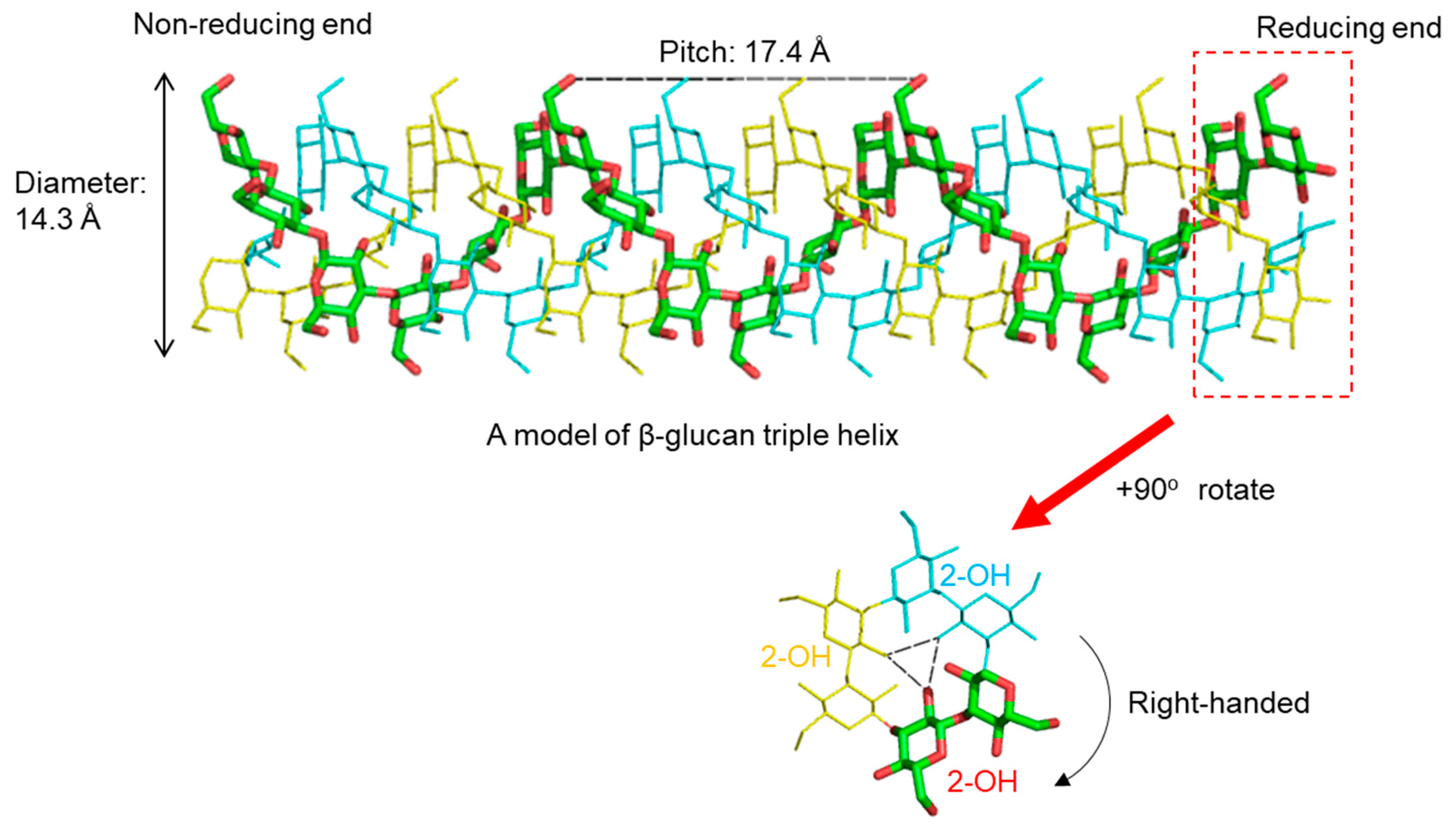
2. 3D Structure of β-Glucan—What Does β-Glucan Look Like?
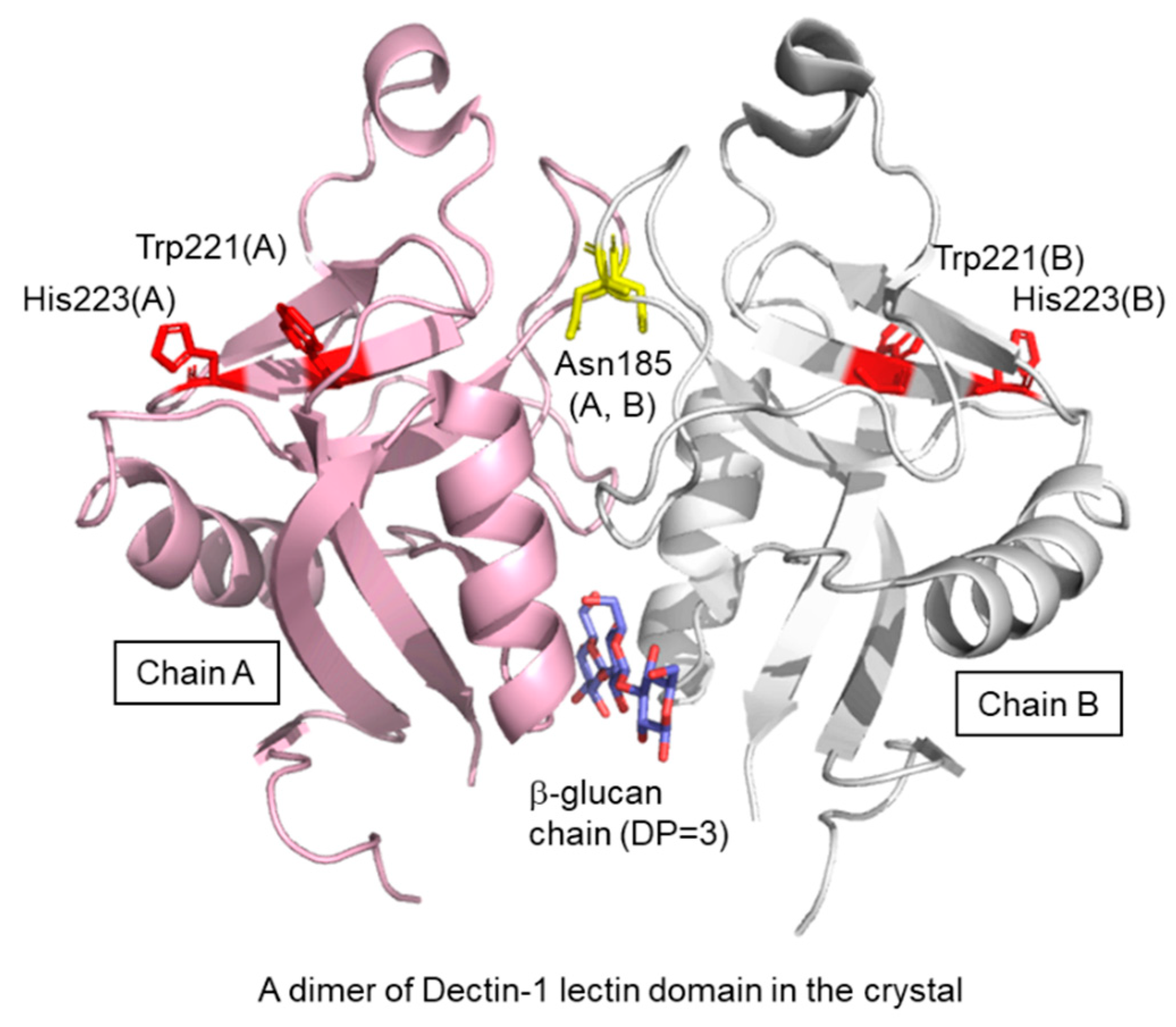
3. Dectin-1–β Glucan Interaction
4. Complement Receptor 3(CR3)–β-Glucan Interaction
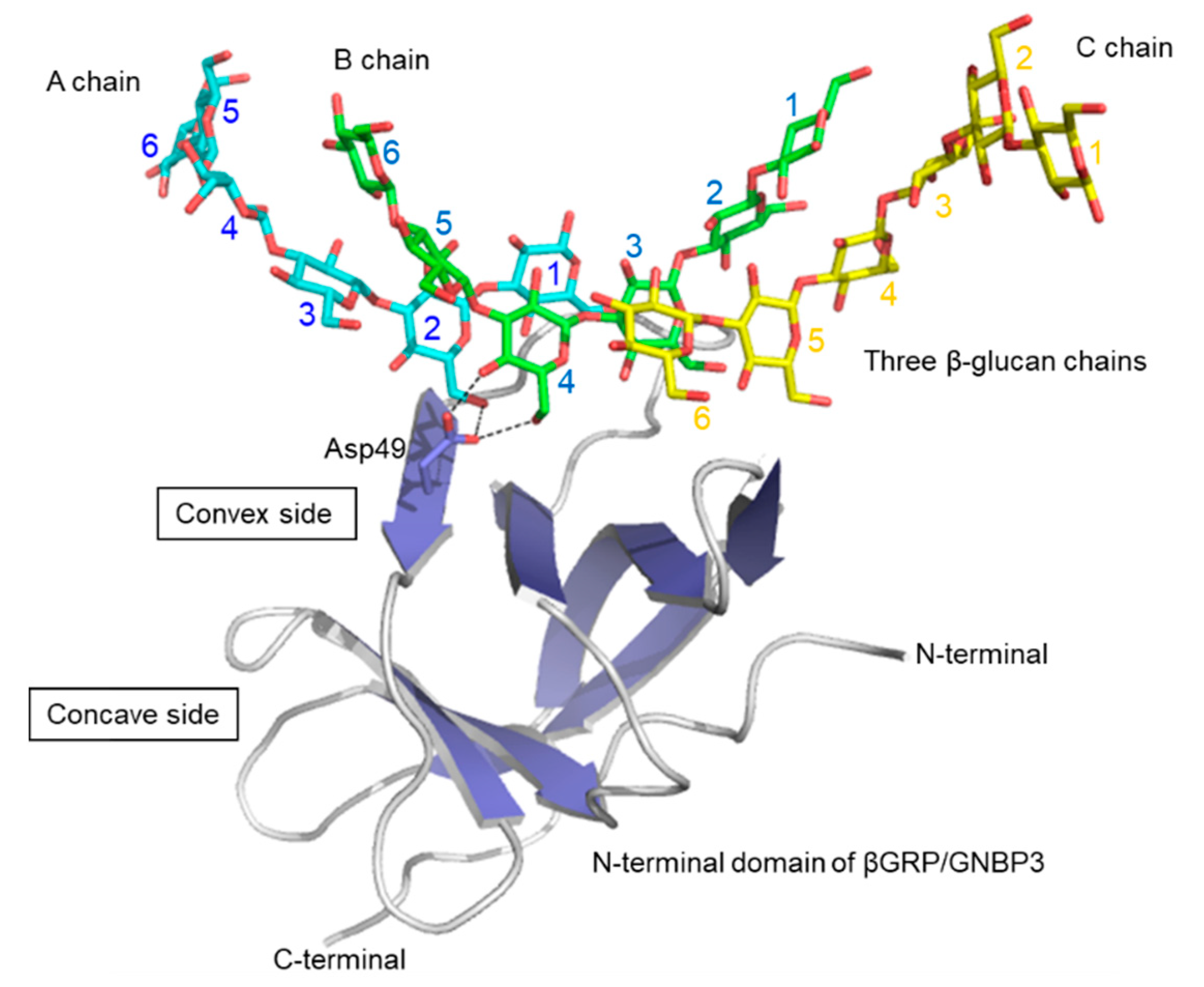
5. βGRP/GNBP3–β-Glucan Interaction
6. Factor G–β-Glucan Interaction
7. Other β-Glucan–Protein Interactions
8. Effect of β(1,6)-Branching on β-Glucan Conformation
9. Summary and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Harada, T.; Masuda, M.; Fujimori, K.; Maeda, I. Production of a firm, resilient gel-forming polysaccharide by a mutant of Alcaligenes faecalis var. myxogenes 10C3. Agric. Biol. Chem. 1966, 30, 196–198. [Google Scholar]
- Saito, H.; Misaki, A.; Harada, T. A comparison of strucure of curdlan and pachyman. Agric. Biol. Chem. 1968, 32, 1261–1269. [Google Scholar] [CrossRef]
- Harada, T.; Misaki, A.; Saito, H. Curdlan: A bacterial gel-forming beta-1,3-glucan. Arch. Biochem. Biophys. 1968, 124, 292–298. [Google Scholar] [CrossRef]
- Tabata, K.; Ito, W.; Kojima, T.; Kawabata, S.; Misaki, A. Ultrasonic degradation of schizophyllan, an antitumor polysaccharide produced by Schizophyllum commune Fries. Carbohydr. Res. 1981, 89, 121–135. [Google Scholar] [CrossRef]
- Rinaudo, M.; Vincendon, M. 13C NMR structural investigation of scleroglucan. Carbohydr. Polym. 1982, 2, 135–144. [Google Scholar] [CrossRef]
- Sasaki, T.; Takasuka, N. Further study of the structure of lentinan, an anti-tumor polysaccharide from Lentinus edodes. Carbohydr. Res. 1976, 47, 99–104. [Google Scholar] [CrossRef]
- Read, S.M.; Currie, G.; Bacic, A. Analysis of the structural heterogeneity of laminarin by electrospray-ionisation-mass spectrometry. Carbohydr. Res. 1996, 281, 187–201. [Google Scholar] [CrossRef]
- Ohno, N.; Uchiyama, M.; Tsuzuki, A.; Tokunaka, K.; Miura, N.N.; Adachi, Y.; Aizawa, M.W.; Tamura, H.; Tanaka, S.; Yadomae, T. Solubilization of yeast cell-wall beta-(1-->3)-D-glucan by sodium hypochlorite oxidation and dimethyl sulfoxide extraction. Carbohydr. Res. 1999, 316, 161–172. [Google Scholar] [CrossRef]
- Sletmoen, M.; Stokke, B.T. Higher order structure of (1,3)-β-D-glucans and its influence on their biological activities and complexation abilities. Biopolymers 2008, 89, 310–321. [Google Scholar] [CrossRef]
- Bluhm, T.L.; Sarko, A. The triple helical structure of lentinan, a linear β-(1→3)-D-glucan. Can. J. Chem. 1977, 55, 293–299. [Google Scholar] [CrossRef]
- Chuah, C.T.; Sarko, A.; Deslandes, Y.; Marchessault, R.H. Packing analysis of carbohydrates and polysaccharides. Part 14. Triple-helical crystalline-structure of curdlan and paramylon hydrates. Macromolecules 1983, 16, 1375–1382. [Google Scholar] [CrossRef]
- Bluhm, T.L.; Deslandes, Y.; Marchessault, R.H.; Perez, S.; Rinaudo, M. Solid-state and solution conformation of scleroglucan. Carbohydr. Res. 1982, 100, 117–130. [Google Scholar] [CrossRef]
- Kashiwagi, Y.; Norisuye, T.; Fujita, H. Triple helix of Schizophyllum commune polysaccharide in dilute solution. 4. Light scattering and viscosity in dilute aqueous sodium hydroxide. Macromolecules 1981, 14, 1220–1225. [Google Scholar] [CrossRef]
- Ogawa, K.; Tsurugi, J.; Watanabe, T. The dependence of the conformation of a (1→3)-β-D-glucan on chain-length in alkaline solution. Carbohydr. Res. 1973, 29, 397–403. [Google Scholar] [CrossRef]
- Kojima, T.; Tabata, K.; Itoh, W.; Yanaki, T. Molecular-weight dependence of the antitumor-activity of schizophyllan. Agric. Biol. Chem. 1986, 50, 231–232. [Google Scholar]
- Hanashima, S.; Ikeda, A.; Tanaka, H.; Adachi, Y.; Ohno, N.; Takahashi, T.; Yamaguchi, Y. NMR study of short β(1-3)-glucans provides insights into the structure and interaction with Dectin-1. Glycoconj. J. 2014, 31, 199–207. [Google Scholar] [CrossRef]
- Oda, M.; Tanabe, Y.; Noda, M.; Inaba, S.; Krayukhina, E.; Fukada, H.; Uchiyama, S. Structural and binding properties of laminarin revealed by analytical ultracentrifugation and calorimetric analyses. Carbohydr. Res. 2016, 431, 33–38. [Google Scholar] [CrossRef]
- Norisuye, T.; Yanaki, T.; Fujita, H. Triple helix of a schizophyllum-commune polysaccharide in aqueous-solution. J. Polym. Sci. Part B Polym. Phys. 1980, 18, 547–558. [Google Scholar] [CrossRef]
- Yanaki, T.; Tabata, K.; Kojima, T. Melting behavior of a triple helical polysaccharide schizophyllan in aqueous-solution. Carbohyd. Polym. 1985, 5, 275–283. [Google Scholar] [CrossRef]
- Stokke, B.T.; Elgsaeter, A.; Brant, D.A.; Kuge, T.; Kitamura, S. Macromolecular cyclization of (1-->6)-branched-(1-->3)-beta-D-glucans observed after denaturation-renaturation of the triple-helical structure. Biopolymers 1993, 33, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, L.; Xu, X. Morphologies and conformation transition of Lentinan in aqueous NaOH solution. Biopolymers 2004, 75, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D. Dectin-1: A signalling non-TLR pattern-recognition receptor. Nat. Rev. Immunol. 2006, 6, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Palma, A.S.; Feizi, T.; Zhang, Y.; Stoll, M.S.; Lawson, A.M.; Díaz-Rodríguez, E.; Campanero-Rhodes, M.A.; Costa, J.; Gordon, S.; Brown, G.D.; et al. Ligands for the β-glucan receptor, Dectin-1, assigned using "designer" microarrays of oligosaccharide probes (neoglycolipids) generated from glucan polysaccharides. J. Biol. Chem. 2006, 281, 5771–5779. [Google Scholar] [CrossRef]
- Tanaka, H.; Kawai, T.; Adachi, Y.; Hanashima, S.; Yamaguchi, Y.; Ohno, N.; Takahashi, T. Synthesis of β(1,3) oligoglucans exhibiting a Dectin-1 binding affinity and their biological evaluation. Bioorg. Med. Chem. 2012, 20, 3898–3914. [Google Scholar] [CrossRef]
- Adams, E.L.; Rice, P.J.; Graves, B.; Ensley, H.E.; Yu, H.; Brown, G.D.; Gordon, S.; Monteiro, M.A.; Papp-Szabo, E.; Lowman, D.W.; et al. Differential high-affinity interaction of dectin-1 with natural or synthetic glucans is dependent upon primary structure and is influenced by polymer chain length and side-chain branching. J. Pharmacol. Exp. Ther. 2008, 325, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.; O’Callaghan, C.A.; Marshall, A.S.; Gilbert, R.J.; Siebold, C.; Gordon, S.; Brown, G.D.; Jones, E.Y. Structure of the fungal β-glucan-binding immune receptor dectin-1: Implications for function. Protein Sci. 2007, 16, 1042–1052. [Google Scholar] [CrossRef]
- Adachi, Y.; Ishii, T.; Ikeda, Y.; Hoshino, A.; Tamura, H.; Aketagawa, J.; Tanaka, S.; Ohno, N. Characterization of β-glucan recognition site on C-type lectin, dectin 1. Infect. Immun. 2004, 72, 4159–4171. [Google Scholar] [CrossRef]
- Kato, Y.; Adachi, Y.; Ohno, N. Contribution of N-linked oligosaccharides to the expression and functions of β-glucan receptor, Dectin-1. Biol. Pharm. Bull. 2006, 29, 1580–1586. [Google Scholar] [CrossRef]
- Dulal, H.P.; Adachi, Y.; Ohno, N.; Yamaguchi, Y. β-Glucan-induced cooperative oligomerization of Dectin-1 C-type lectin like domain. Glycobiology 2018, in press. [Google Scholar] [CrossRef]
- Anaya, E.U.; Amin, A.E.; Danielson, M.; Michel, K.; Neumann, A.K. Innate antifungal immune receptor, Dectin-1, undergoes ligand-induced oligomerization with highly structured β-glucans and at fungal cell contact sites. bioRxiv 2019. [Google Scholar] [CrossRef]
- Bode, K.; Bujupi, F.; Link, C.; Hein, T.; Zimmermann, S.; Peiris, D.; Jaquet, V.; Lepenies, B.; Weyd, H.; Krammer, P.H. Dectin-1 binding to annexins on apoptotic cells induces peripheral immune tolerance via NADPH oxidase-2. Cell Rep. 2019, 29, 4435–4446.e4439. [Google Scholar] [CrossRef]
- Manabe, Y.; Marchetti, R.; Takakura, Y.; Nagasaki, M.; Nihei, W.; Takebe, T.; Tanaka, K.; Kabayama, K.; Chiodo, F.; Hanashima, S.; et al. The core fucose on an IgG antibody is an endogenous ligand of Dectin-1. Angew. Chem. Int. Ed. Engl. 2019, 58, 18697–18702. [Google Scholar] [CrossRef]
- Thornton, B.P.; Vetvicka, V.; Pitman, M.; Goldman, R.C.; Ross, G.D. Analysis of the sugar specificity and molecular location of the beta-glucan-binding lectin site of complement receptor type 3 (CD11b/CD18). J. Immunol. 1996, 156, 1235–1246. [Google Scholar]
- Xia, Y.; Ross, G.D. Generation of recombinant fragments of CD11b expressing the functional beta-glucan-binding lectin site of CR3 (CD11b/CD18). J. Immunol. 1999, 162, 7285–7293. [Google Scholar]
- Brown, G.D.; Gordon, S. Immune recognition of fungal β-glucans. Cell Microbiol. 2005, 7, 471–479. [Google Scholar] [CrossRef]
- Takahasi, K.; Ochiai, M.; Horiuchi, M.; Kumeta, H.; Ogura, K.; Ashida, M.; Inagaki, F. Solution structure of the silkworm betaGRP/GNBP3 N-terminal domain reveals the mechanism for beta-1,3-glucan-specific recognition. Proc. Natl. Acad. Sci. USA 2009, 106, 11679–11684. [Google Scholar] [CrossRef]
- Mishima, Y.; Quintin, J.; Aimanianda, V.; Kellenberger, C.; Coste, F.; Clavaud, C.; Hetru, C.; Hoffmann, J.A.; Latge, J.P.; Ferrandon, D.; et al. The N-terminal domain of Drosophila Gram-negative binding protein 3 (GNBP3) defines a novel family of fungal pattern recognition receptors. J. Biol. Chem. 2009, 284, 28687–28697. [Google Scholar] [CrossRef]
- Kanagawa, M.; Satoh, T.; Ikeda, A.; Adachi, Y.; Ohno, N.; Yamaguchi, Y. Structural insights into recognition of triple-helical β-glucans by an insect fungal receptor. J. Biol. Chem. 2011, 286, 29158–29165. [Google Scholar] [CrossRef]
- Rao, X.J.; Zhan, M.Y.; Pan, Y.M.; Liu, S.; Yang, P.J.; Yang, L.L.; Yu, X.Q. Immune functions of insect betaGRPs and their potential application. Dev. Comp. Immunol. 2018, 83, 80–88. [Google Scholar] [CrossRef]
- Adachi, Y.; Ishii, M.; Kanno, T.; Tetsui, J.; Ishibashi, K.I.; Yamanaka, D.; Miura, N.; Ohno, N. N-Terminal (1-->3)-beta-d-glucan recognition proteins from insects recognize the difference in ultra-structures of (1-->3)-beta-d-glucan. Int. J. Mol. Sci. 2019, 20, 3498. [Google Scholar] [CrossRef]
- Ochiai, M.; Ashida, M. A pattern-recognition protein for β-1,3-glucan. The binding domain and the cDNA cloning of β-1,3-glucan recognition protein from the silkworm, Bombyx mori. J. Biol. Chem. 2000, 275, 4995–5002. [Google Scholar] [CrossRef]
- Ma, C.; Kanost, M.R. A β1,3-glucan recognition protein from an insect, Manduca sexta, agglutinates microorganisms and activates the phenoloxidase cascade. J. Biol. Chem. 2000, 275, 7505–7514. [Google Scholar] [CrossRef]
- Gottar, M.; Gobert, V.; Matskevich, A.A.; Reichhart, J.M.; Wang, C.; Butt, T.M.; Belvin, M.; Hoffmann, J.A.; Ferrandon, D. Dual detection of fungal infections in Drosophila via recognition of glucans and sensing of virulence factors. Cell 2006, 127, 1425–1437. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Hiromasa, Y.; Takahashi, D.; VanderVelde, D.; Fabrick, J.A.; Kanost, M.R.; Krishnamoorthi, R. An initial event in the insect innate immune response: Structural and biological studies of interactions between beta-1,3-glucan and the N-terminal domain of beta-1,3-glucan recognition protein. Biochemistry 2013, 52, 161–170. [Google Scholar] [CrossRef]
- Takaki, Y.; Seki, N.; Kawabata Si, S.; Iwanaga, S.; Muta, T. Duplicated binding sites for (1-->3)-beta-D-glucan in the horseshoe crab coagulation factor G: Implications for a molecular basis of the pattern recognition in innate immunity. J. Biol. Chem. 2002, 277, 14281–14287. [Google Scholar] [CrossRef]
- Ueda, Y.; Ohwada, S.; Abe, Y.; Shibata, T.; Iijima, M.; Yoshimitsu, Y.; Koshiba, T.; Nakata, M.; Ueda, T.; Kawabata, S. Factor G utilizes a carbohydrate-binding cleft that is conserved between horseshoe crab and bacteria for the recognition of beta-1,3-D-glucans. J. Immunol. 2009, 183, 3810–3818. [Google Scholar] [CrossRef] [PubMed]
- Saito, H.; Yoshioka, Y.; Uehara, N.; Aketagawa, J.; Tanaka, S.; Shibata, Y. Relationship between conformation and biological response for (1----3)-beta-D-glucans in the activation of coagulation factor G from limulus amebocyte lysate and host-mediated antitumor activity. Demonstration of single-helix conformation as a stimulant. Carbohydr. Res. 1991, 217, 181–190. [Google Scholar] [CrossRef]
- Theel, E.S.; Doern, C.D. beta-D-glucan testing is important for diagnosis of invasive fungal infections. J. Clin. Microbiol. 2013, 51, 3478–3483. [Google Scholar] [CrossRef] [PubMed]
- Boraston, A.B.; Bolam, D.N.; Gilbert, H.J.; Davies, G.J. Carbohydrate-binding modules: Fine-tuning polysaccharide recognition. Biochem. J. 2004, 382, 769–781. [Google Scholar] [CrossRef]
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef]
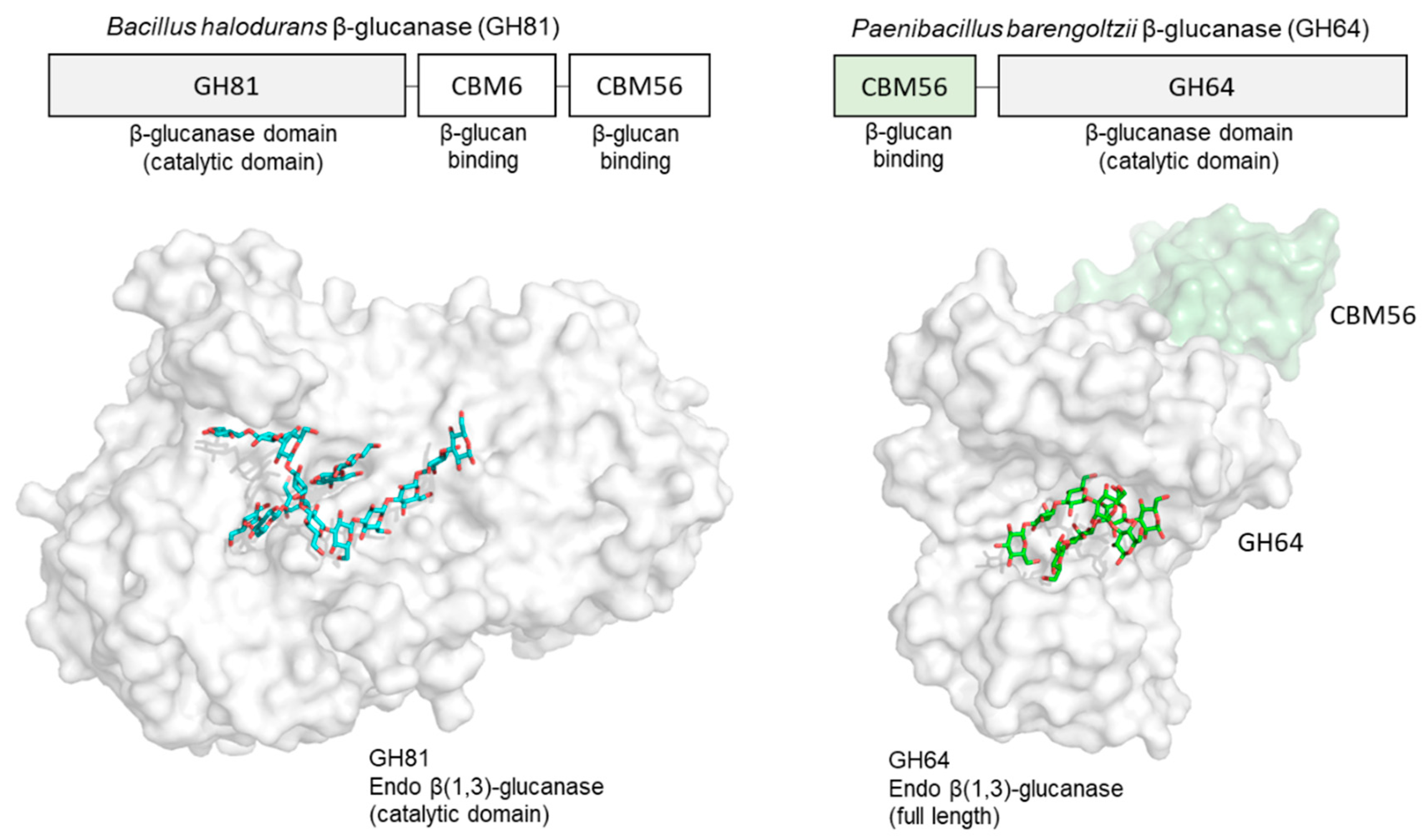
- Pluvinage, B.; Fillo, A.; Massel, P.; Boraston, A.B. Structural analysis of a family 81 glycoside hydrolase implicates its recognition of β-1,3-glucan quaternary structure. Structure 2017, 25, 1348–1359.e3. [Google Scholar] [CrossRef]
- van Bueren, A.L.; Morland, C.; Gilbert, H.J.; Boraston, A.B. Family 6 carbohydrate binding modules recognize the non-reducing end of beta-1,3-linked glucans by presenting a unique ligand binding surface. J. Biol. Chem. 2005, 280, 530–537. [Google Scholar] [CrossRef]
- Hettle, A.; Fillo, A.; Abe, K.; Massel, P.; Pluvinage, B.; Langelaan, D.N.; Smith, S.P.; Boraston, A.B. Properties of a family 56 carbohydrate-binding module and its role in the recognition and hydrolysis of beta-1,3-glucan. J. Biol. Chem. 2017, 292, 16955–16968. [Google Scholar] [CrossRef]
- Qin, Z.; Yang, D.; You, X.; Liu, Y.; Hu, S.; Yan, Q.; Yang, S.; Jiang, Z. The recognition mechanism of triple-helical β-1,3-glucan by a β-1,3-glucanase. Chem. Commun. 2017, 53, 9368–9371. [Google Scholar] [CrossRef] [PubMed]
- Sung, K.H.; Josewski, J.; Dubel, S.; Blankenfeldt, W.; Rau, U. Structural insights into antigen recognition of an anti-β-(1,6)-β-(1,3)-D-glucan antibody. Sci. Rep. 2018, 8, 13652. [Google Scholar] [CrossRef]
- Okobira, T.; Miyoshi, K.; Uezu, K.; Sakurai, K.; Shinkai, S. Molecular dynamics studies of side chain effect on the β-1,3-D-glucan triple helix in aqueous solution. Biomacromolecules 2008, 9, 783–788. [Google Scholar] [CrossRef] [PubMed]
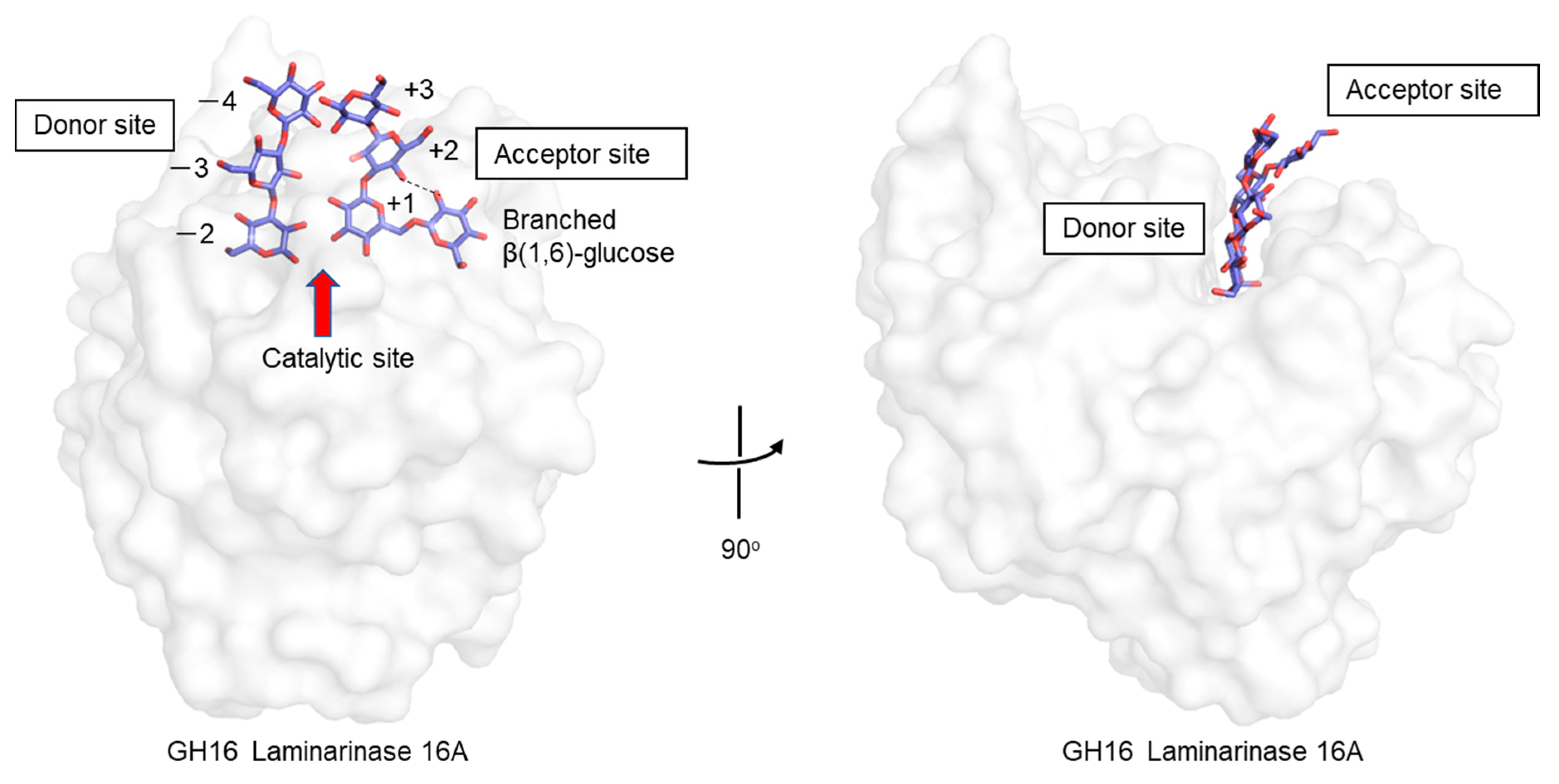
- Vasur, J.; Kawai, R.; Andersson, E.; Igarashi, K.; Sandgren, M.; Samejima, M.; Stahlberg, J. X-ray crystal structures of Phanerochaete chrysosporium Laminarinase 16A in complex with products from lichenin and laminarin hydrolysis. FEBS J. 2009, 276, 3858–3869. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, Y.; Uehara, N.; Saito, H. Conformation-dependent change in antitumor activity of linear and branched (1-->3)-β-D-glucans on the basis of conformational elucidation by carbon-13 nuclear magnetic resonance spectroscopy. Chem. Pharm. Bull. 1992, 40, 1221–1226. [Google Scholar] [CrossRef] [PubMed]
- Kulicke, W.M.; Lettau, A.I.; Thielking, H. Correlation between immunological activity, molar mass, and molecular structure of different (1-->3)-beta-D-glucans. Carbohydr. Res. 1997, 297, 135–143. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manabe, N.; Yamaguchi, Y. 3D Structural Insights into β-Glucans and Their Binding Proteins. Int. J. Mol. Sci. 2021, 22, 1578. https://doi.org/10.3390/ijms22041578
Manabe N, Yamaguchi Y. 3D Structural Insights into β-Glucans and Their Binding Proteins. International Journal of Molecular Sciences. 2021; 22(4):1578. https://doi.org/10.3390/ijms22041578
Chicago/Turabian StyleManabe, Noriyoshi, and Yoshiki Yamaguchi. 2021. "3D Structural Insights into β-Glucans and Their Binding Proteins" International Journal of Molecular Sciences 22, no. 4: 1578. https://doi.org/10.3390/ijms22041578
APA StyleManabe, N., & Yamaguchi, Y. (2021). 3D Structural Insights into β-Glucans and Their Binding Proteins. International Journal of Molecular Sciences, 22(4), 1578. https://doi.org/10.3390/ijms22041578