
MFN1 and MFN2 Are Dispensable for Sperm Development and Functions in Mice

Abstract
:1. Introduction
2. Results
2.1. Mfn1 and Mfn2 mRNA Are Highly Expressed in Testes
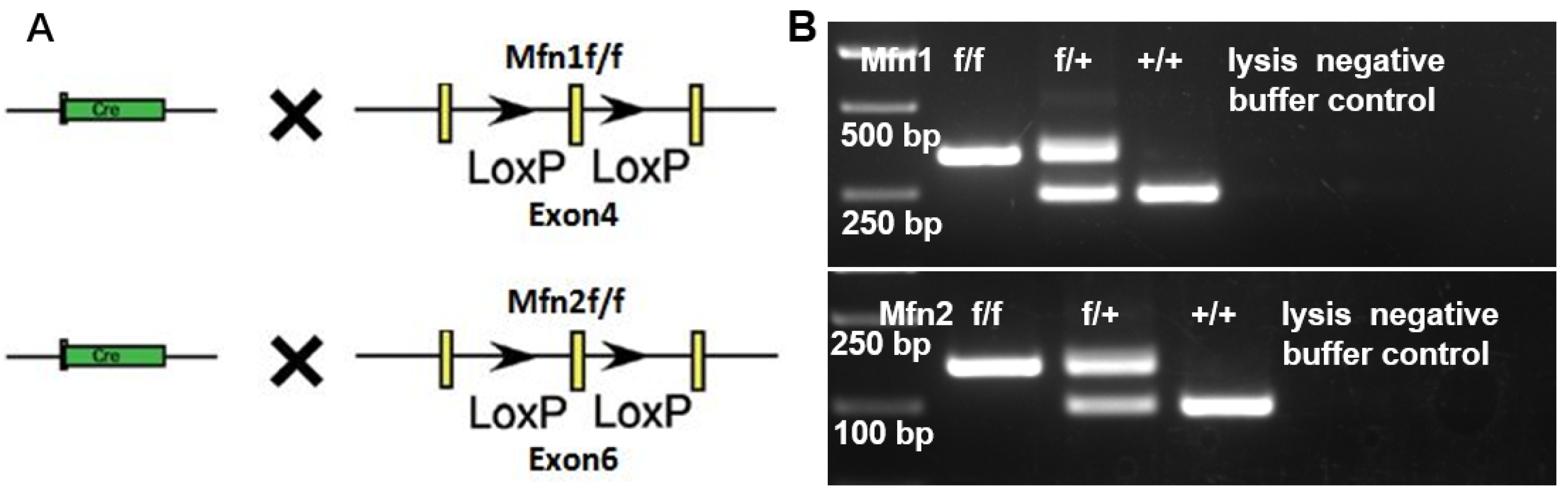
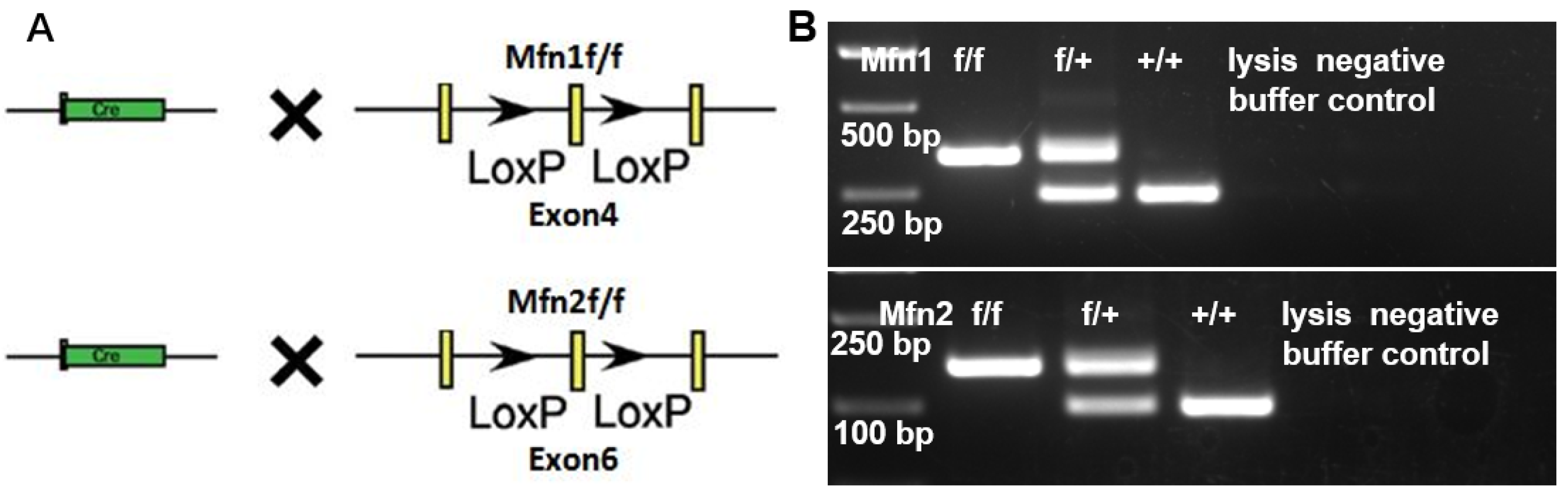
2.2. Generation of Mfn1 and Mfn2 Conditional Knockout (cKO) Mice
2.3. Either Mfn1 or Mfn2 Deletion in Sperm Does Not Affect Sperm Development and Functions
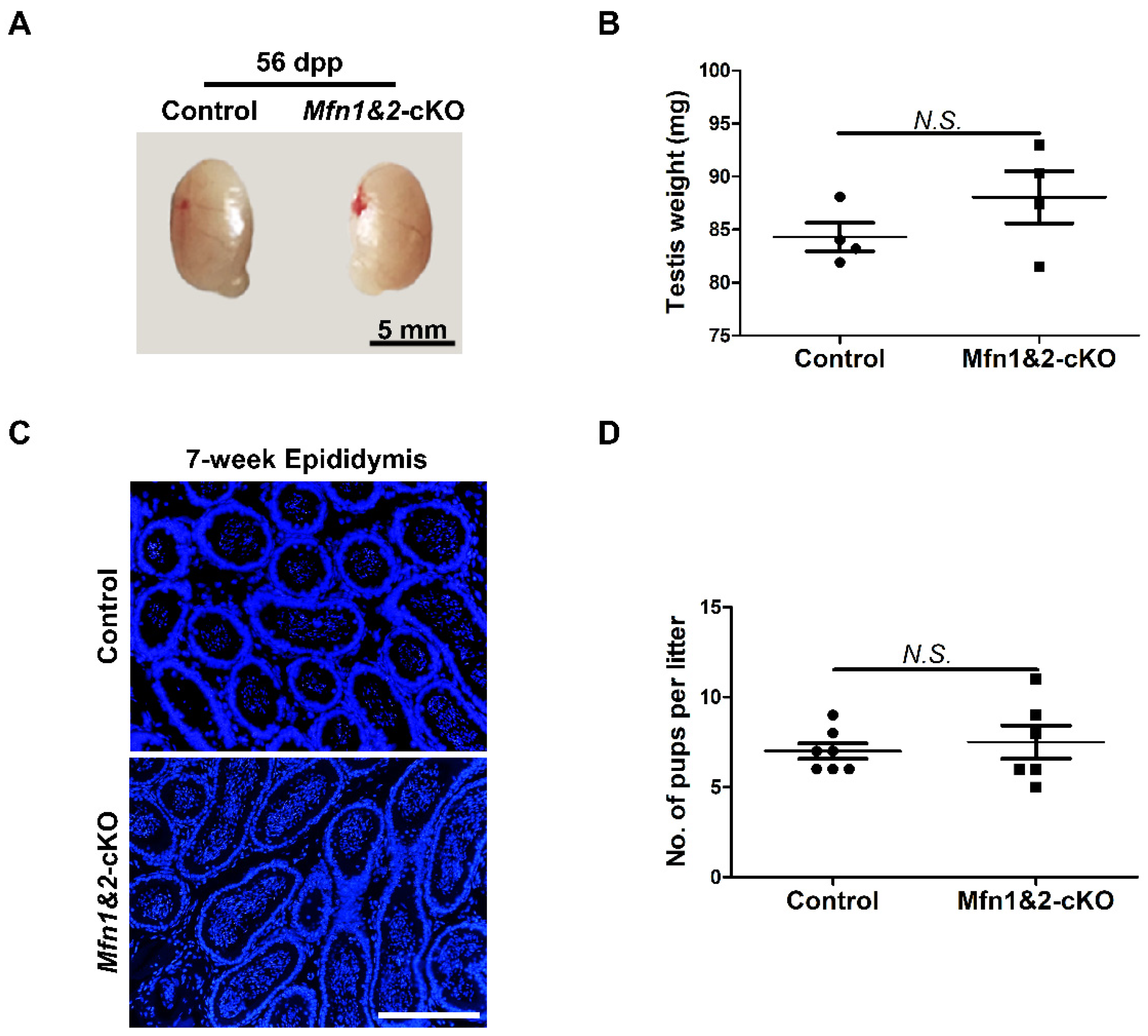
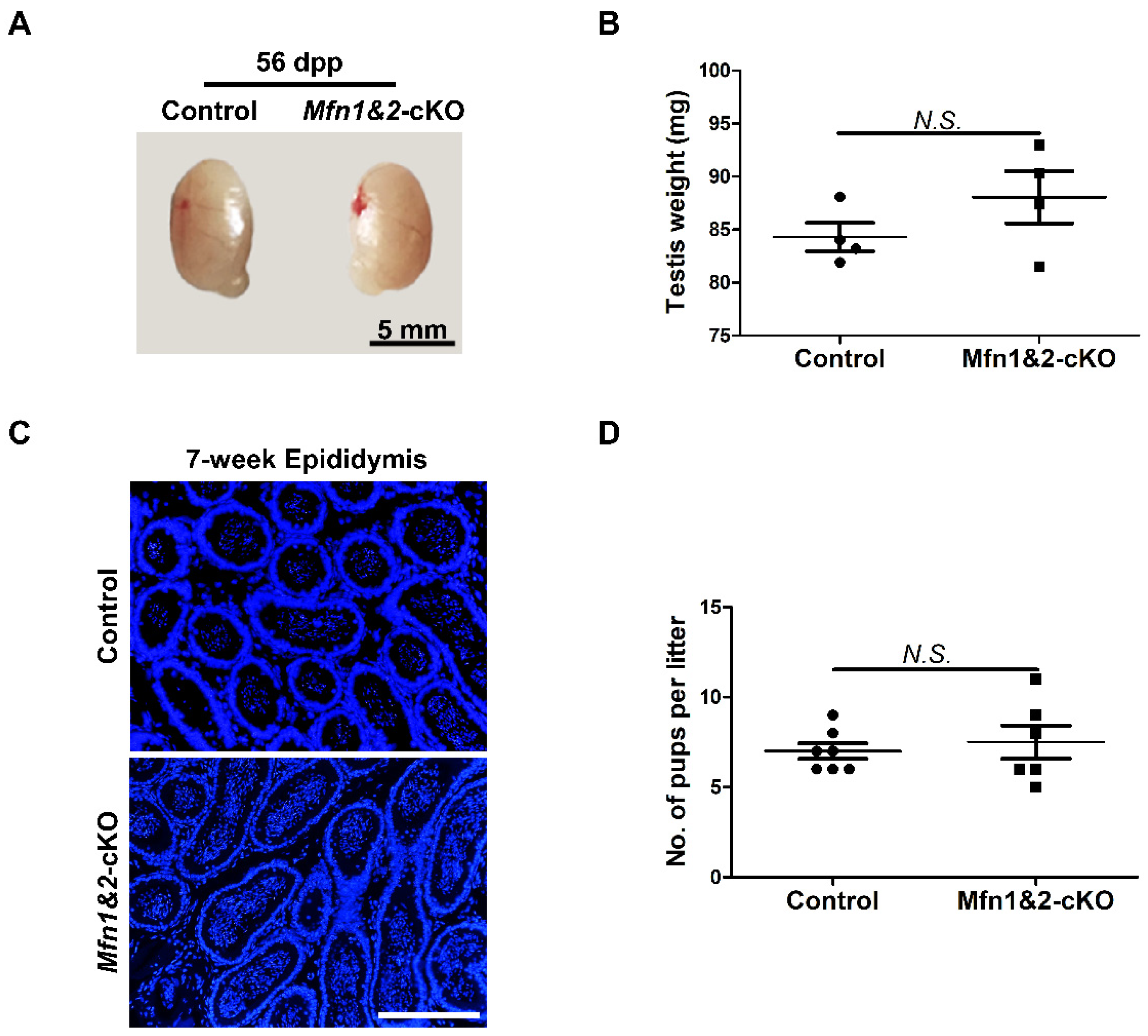
2.4. Combined Deletion of Mfn1 and Mfn2 in Sperm Does Not Affect Sperm Development and Functions
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Mouse Lines, Animal Care, and Fertility Test
4.3. Genomic DNA Extraction and Genotyping
4.4. Total RNA Extraction, Reverse Transcription, and Real-Time PCR
4.5. Histology and Immunohistofluorensce (IHF)
4.6. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Mfn1 | Mitofusin 1 |
| Mfn2 | Mitofusin 2 |
| SYCP3 | Synaptonemal complex protein 3 |
| TRA98 | Germ cell specific antigen 98 |
| mtDNAs | Mitochondrial DNAs |
| OPA1 | Optic atrophy 1 |
| DRP1 | Dynamin-related protein 1 |
| Prm1 | Protamine 1 |
| cKO | Conditional knockout |
| IHF | Immunohistofluorescence |
| dpp | Days postpartum |
| MEFs | Mouse embryonic fibroblasts |
| PCR | Polymerase chain reaction |
| PFA | Paraformaldehyde |
References
- Chen, H.; Chan, D.C. Mitochondrial Dynamics in Regulating the Unique Phenotypes of Cancer and Stem Cells. Cell Metab. 2017, 26, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Mishra, P.; Chan, D.C. Mitochondrial dynamics and inheritance during cell division, development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 634–646. [Google Scholar] [CrossRef] [Green Version]
- Samiec, M.; Skrzyszowska, M. Extranuclear Inheritance of Mitochondrial Genome and Epigenetic Reprogrammability of Chromosomal Telomeres in Somatic Cell Cloning of Mammals. Int. J. Mol. Sci. 2021, 22, 3099. [Google Scholar] [CrossRef]
- Shaw, J.M.; Nunnari, J. Mitochondrial dynamics and division in budding yeast. Trends Cell Biol. 2002, 12, 178–184. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Chan, D.C. Mitochondrial Dynamics in Mammals. Curr. Top. Dev. Biol. 2004, 59, 119–144. [Google Scholar] [CrossRef]
- Chan, D.C. Fusion and Fission: Interlinked Processes Critical for Mitochondrial Health. Annu. Rev. Genet. 2012, 46, 265–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labbé, K.; Murley, A.; Nunnari, J. Determinants and Functions of Mitochondrial Behavior. Annu. Rev. Cell Dev. Biol. 2014, 30, 357–391. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.C. Mitochondrial Fusion and Fission in Mammals. Annu. Rev. Cell Dev. Biol. 2006, 22, 79–99. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Detmer, S.A.; Ewald, A.J.; Griffin, E.E.; Fraser, S.E.; Chan, D.C. Mitofusins Mfn1 and Mfn2 coordinately regulate mitochondrial fusion and are essential for embryonic development. J. Cell Biol. 2003, 160, 189–200. [Google Scholar] [CrossRef]
- Hsiuchen, C.J. Michael McCaffery, David C Chan. Mitochondrial fusion protects against neurodegeneration in the cere-bellum. Cell 2007, 130, 548–562. [Google Scholar]
- Zhang, J.; Wang, Q.; Wang, M.; Jiang, M.; Wang, Y.; Sun, Y.; Wang, J.; Xie, T.; Tang, C.; Tang, N.; et al. GASZ and mi-tofusin-mediated mitochondrial functions are crucial for spermatogenesis. EMBO Rep. 2016, 17, 220–234. [Google Scholar] [CrossRef] [Green Version]
- Varuzhanyan, G.; Rojansky, R.; Sweredoski, M.J.; Graham, R.L.; Hess, S.; Ladinsky, M.S.; Chan, D.C. Mitochondrial fusion is required for spermatogonial differentiation and meiosis. eLife 2019, 8, 51601. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Sun, Y.; Sun, Q.; Zhang, J.; Jiang, M.; Chang, C.; Huang, X.; Wang, C.; Wang, P.; Zhang, Z.; et al. MFN2 Plays a Distinct Role from MFN1 in Regulating Spermatogonial Differentiation. Stem Cell Rep. 2020, 14, 803–817. [Google Scholar] [CrossRef]
- Wang, X.; Wen, Y.; Zhang, J. Grace Swanson, Shuangshuang Guo, Congcong Cao, Stephen A Krawetz, Zhibing Zhang, Shuiqiao Yuan. Mitochondrial fusion and fission in mammals. Development 2021, 148, dev196295. [Google Scholar] [CrossRef]
- Honda, S. Shigehisa Hirose. Stage-specific enhanced expression of mitochondrial fusion and fission factors during sper-matogenesis in rat testis. Biochem. Biophys. Res. Commun. 2003, 11, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Vadnais, M.L.; Lin, A.M.; Gerton, G.L. Mitochondrial fusion protein MFN2 interacts with the mitostatin-related protein MNS1 required for mouse sperm flagellar structure and function. Cilia 2014, 3, 5. [Google Scholar] [CrossRef] [Green Version]
- Liu, R.; Chan, D.C. The mitochondrial fission receptor Mff selectively recruits oligomerized Drp1. Mol. Biol. Cell 2015, 26, 4466–4477. [Google Scholar] [CrossRef]
- Varuzhanyan, G.; Chen, H.; Rojansky, R.; Ladinsky, M.S.; Michael, J. McCaffery, David C Chan. Mitochon-drial fission factor (Mff) is required for organization of the mitochondrial sheath in spermatids. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129845. [Google Scholar] [CrossRef]
- Varuzhanyan, G.; Ladinsky, M.S.; Yamashita, S.-I.; Abe, M.; Sakimura, K.; Kanki, T.; Chan, D.C. Fis1 ablation in the male germline disrupts mitochondrial morphology and mitophagy, and arrests spermatid maturation. Development 2021, 148, 199686. [Google Scholar] [CrossRef]
- Hecht, N.B.; Bower, P.A.; Waters, S.H.; Yelick, P.C.; Distel, R.J. Evidence for haploid expression of mouse testicular genes. Exp. Cell Res. 1986, 164, 183–190. [Google Scholar] [CrossRef]
- Matsumura, H.; Hasuwa, H.; Inoue, N.; Ikawa, M.; Okabe, M. Lineage-specific cell disruption in living mice by Cre-mediated expression of diphtheria toxin A chain. Biochem. Biophys. Res. Commun. 2004, 321, 275–279. [Google Scholar] [CrossRef]
- Tanaka, H.; Pereira, L.A.V.D.; Nozaki, M.; Tsuchida, J.; Sawada, K.; Mori, H.; Nishimune, Y. A germ cell-specific nuclear antigen recognized by a monoclonal antibody raised against mouse testicular germ cells. Int. J. Androl. 1998, 20, 361–366. [Google Scholar] [CrossRef]
- Liu, J.-G.; Yuan, L.; Brundell, E.; Bjorkroth, B.; Daneholt, B.; Höög, C. Localization of the N-terminus of SCP1 to the Central Element of the Synaptonemal Complex and Evidence for Direct Interactions between the N-termini of SCP1 Molecules Organized Head-to-Head. Exp. Cell Res. 1996, 226, 11–19. [Google Scholar] [CrossRef]
- Santel, A.; Fuller, M.T. Control of mitochondrial morphology by a human mitofusin. J. Cell Sci. 2001, 114, 867–874. [Google Scholar] [CrossRef] [PubMed]
- Legros, F.; Lombès, A.; Frachon, P.; Rojo, M. Mitochondrial Fusion in Human Cells Is Efficient, Requires the Inner Membrane Potential, and Is Mediated by Mitofusins. Mol. Biol. Cell 2002, 13, 4343–4354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojo, M.; Legros, F. Danielle Chateau, Anne Lombès. Membrane topology and mitochondrial targeting of mitofusins, ubiquitous mammalian homologs of the transmembrane GTPase Fzo. J. Cell Sci. 2002, 115, 1663–1674. [Google Scholar] [CrossRef] [PubMed]
- Santel, A.; Frank, S.; Gaume, B.; Herrler, M.; Youle, R.J.; Fuller, M.T. Mitofusin-1 protein is a generally expressed mediator of mitochondrial fusion in mammalian cells. J. Cell Sci. 2003, 116, 2763–2774. [Google Scholar] [CrossRef] [Green Version]
- Cipolat, S.; De Brito, O.M.; Zilio, B.D.; Scorrano, L. OPA1 requires mitofusin 1 to promote mitochondrial fusion. Proc. Natl. Acad. Sci. USA 2004, 101, 15927–15932. [Google Scholar] [CrossRef] [Green Version]
- Ishihara, N.; Eura, Y.; Mihara, K. Mitofusin 1 and 2 play distinct roles in mitochondrial fusion reactions via GTPase activity. J. Cell Sci. 2004, 117, 6535–6546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Chomyn, A.; Chan, D.C. Disruption of Fusion Results in Mitochondrial Heterogeneity and Dysfunction. J. Biol. Chem. 2005, 280, 26185–26192. [Google Scholar] [CrossRef] [Green Version]
- O’Gorman, S.; Dagenais, N.A.; Qian, M.; Marchuk, Y. Protamine-Cre recombinase transgenes efficiently recombine target sequences in the male germ line of mice, but not in embryonic stem cells. Proc. Natl. Acad. Sci. USA 1997, 94, 14602–14607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batista, F.; Lu, L.; Williams, S.A.; Stanley, P. Complex N-Glycans Are Essential, but Core 1 and 2 Mucin O-Glycans, O-Fucose Glycans, and NOTCH1 Are Dispensable, for Mammalian Spermatogenesis1. Biol. Reprod. 2012, 86, 179. [Google Scholar] [CrossRef] [PubMed]
- Diao, F.; Jiang, C.; Wang, X.-X.; Zhu, R.-L.; Wang, Q.; Yao, B.; Li, C.-J. Alteration of protein prenylation promotes spermatogonial differentiation and exhausts spermatogonial stem cells in newborn mice. Sci. Rep. 2016, 6, 28917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Vermulst, M.; Wang, Y.E.; Chomyn, A.; Prolla, T.A.; McCaffery, M.J. David C Chan. Mito-chondrial fusion is required for mtDNA stability in skeletal muscle and tolerance of mtDNA mutations. Cell 2010, 141, 280–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Martino, C.; Floridi, A.; Marcante, M.L.; Malorni, W.; Barcellona, P.S.; Bellocci, M.; Silvestrini, B. Morphological, histochemical and biochemical studies on germ cell mitochondria of normal rats. Z. Zellforsch. Mikrosk. Anat. 1979, 196, 1–22. [Google Scholar] [CrossRef]
- Ramalho-Santos, J.; Varum, S.; Amaral, S.; Mota, P.C.; Sousa, A.P.; Amaral, A. Mitochondrial functionality in reproduction: From gonads and gametes to embryos and embryonic stem cells. Hum. Reprod. Updat. 2009, 15, 553–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaral, A.; Lourenço, B.; Marques, M.; Ramalho-Santos, J. Mitochondria functionality and sperm quality. Reproduction 2013, 146, R163–R174. [Google Scholar] [CrossRef] [Green Version]
- Varuzhanyan, G.; Chan, D.C. Mitochondrial dynamics during spermatogenesis. J. Cell Sci. 2020, 133, 23593. [Google Scholar] [CrossRef] [PubMed]
- Otani, H.; Tanaka, O.; Kasai, K.-I.; Yoshioka, T. Development of mitochondrial helical sheath in the middle piece of the mouse spermatid tail: Regular dispositions and synchronized changes. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1988, 222, 26–33. [Google Scholar] [CrossRef]
- Ho, H.-C.; Wey, S. Three-dimensional rendering of the mitochondrial sheath morphogenesis during mouse spermi-ogenesis. Microsc. Res. Tech. 2007, 70, 719–723. [Google Scholar] [CrossRef]
- Pham, A.H.; McCaffery, J.M.; Chan, D.C. Mouse lines with photo-activatable mitochondria to study mitochondrial dynamics. Genes 2012, 50, 833–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Wang, P.; Liang, D.; Wang, Y. BRG1 Is Dispensable for Sertoli Cell Development and Functions in Mice. Int. J. Mol. Sci. 2020, 21, 4358. [Google Scholar] [CrossRef] [PubMed]
- Truett, G.; Heeger, P.; Mynatt, R.; Truett, A.; Walker, J.; Warman, M. Preparation of PCR-Quality Mouse Genomic DNA with Hot Sodium Hydroxide and Tris (HotSHOT). Biotechniques 2000, 29, 52–54. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Floxed Alleles | NO. of Males | NO. of Litters | NO. of Pups | F/+ Pups | Del/+ Pups | Transmission Del/+ Allele |
|---|---|---|---|---|---|---|
| Mfn1f/f; Prm1-Cre | 3 | 6 | 43 | 0 | 43 | 100% |
| Mfn2f/f; Prm1-Cre | 3 | 7 | 52 | 0 | 52 | 100% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miao, J.; Chen, W.; Wang, P.; Zhang, X.; Wang, L.; Wang, S.; Wang, Y. MFN1 and MFN2 Are Dispensable for Sperm Development and Functions in Mice. Int. J. Mol. Sci. 2021, 22, 13507. https://doi.org/10.3390/ijms222413507
Miao J, Chen W, Wang P, Zhang X, Wang L, Wang S, Wang Y. MFN1 and MFN2 Are Dispensable for Sperm Development and Functions in Mice. International Journal of Molecular Sciences. 2021; 22(24):13507. https://doi.org/10.3390/ijms222413507
Chicago/Turabian StyleMiao, Junru, Wei Chen, Pengxiang Wang, Xin Zhang, Lei Wang, Shuai Wang, and Yuan Wang. 2021. "MFN1 and MFN2 Are Dispensable for Sperm Development and Functions in Mice" International Journal of Molecular Sciences 22, no. 24: 13507. https://doi.org/10.3390/ijms222413507
APA StyleMiao, J., Chen, W., Wang, P., Zhang, X., Wang, L., Wang, S., & Wang, Y. (2021). MFN1 and MFN2 Are Dispensable for Sperm Development and Functions in Mice. International Journal of Molecular Sciences, 22(24), 13507. https://doi.org/10.3390/ijms222413507