
Correlation of MET-Receptor Overexpression with MET Gene Amplification and Patient Outcome in Malignant Mesothelioma

 , ,
, ,
Abstract
:1. Introduction
2. Results
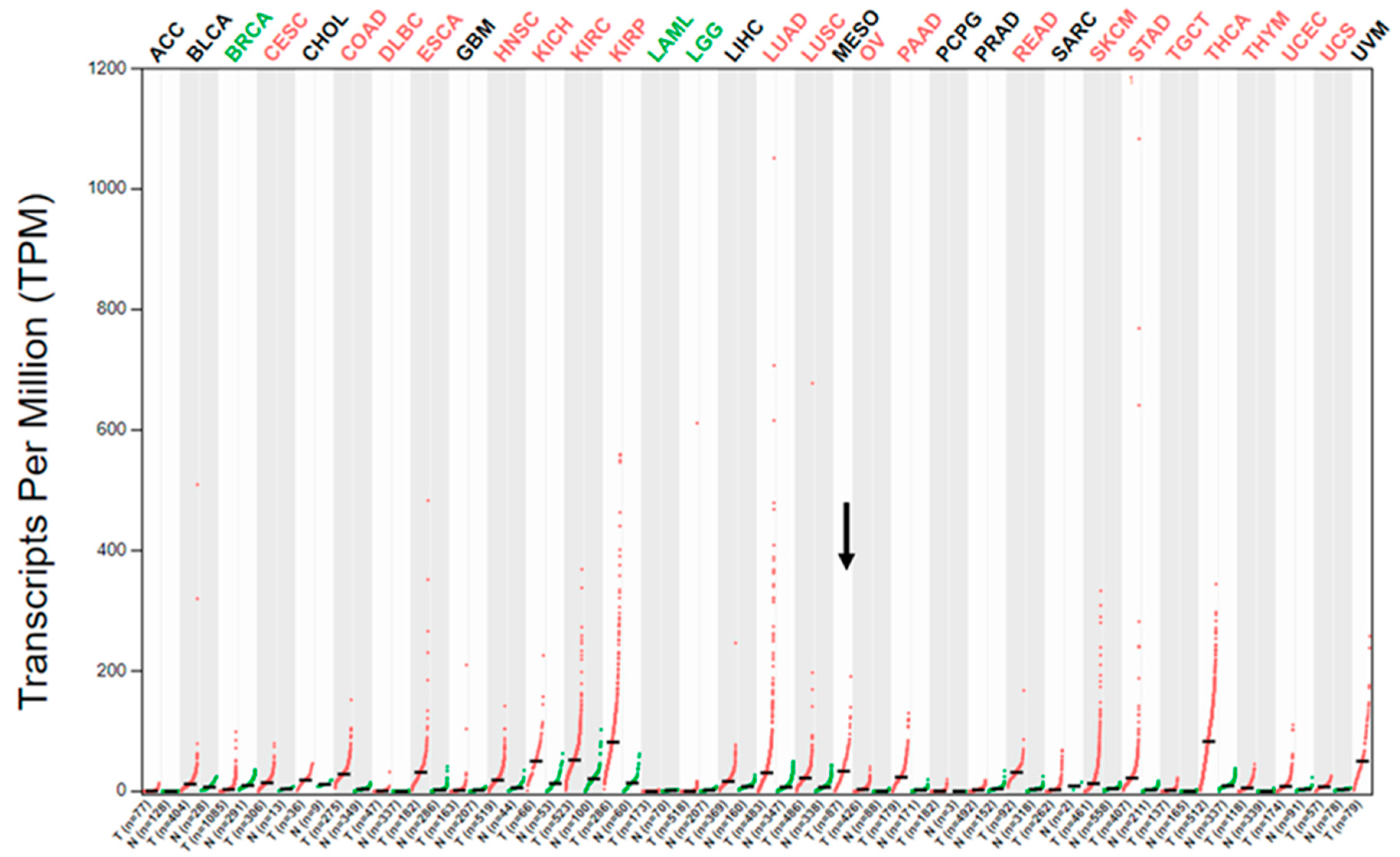
2.1. MET Gene Transcript Profile in MM
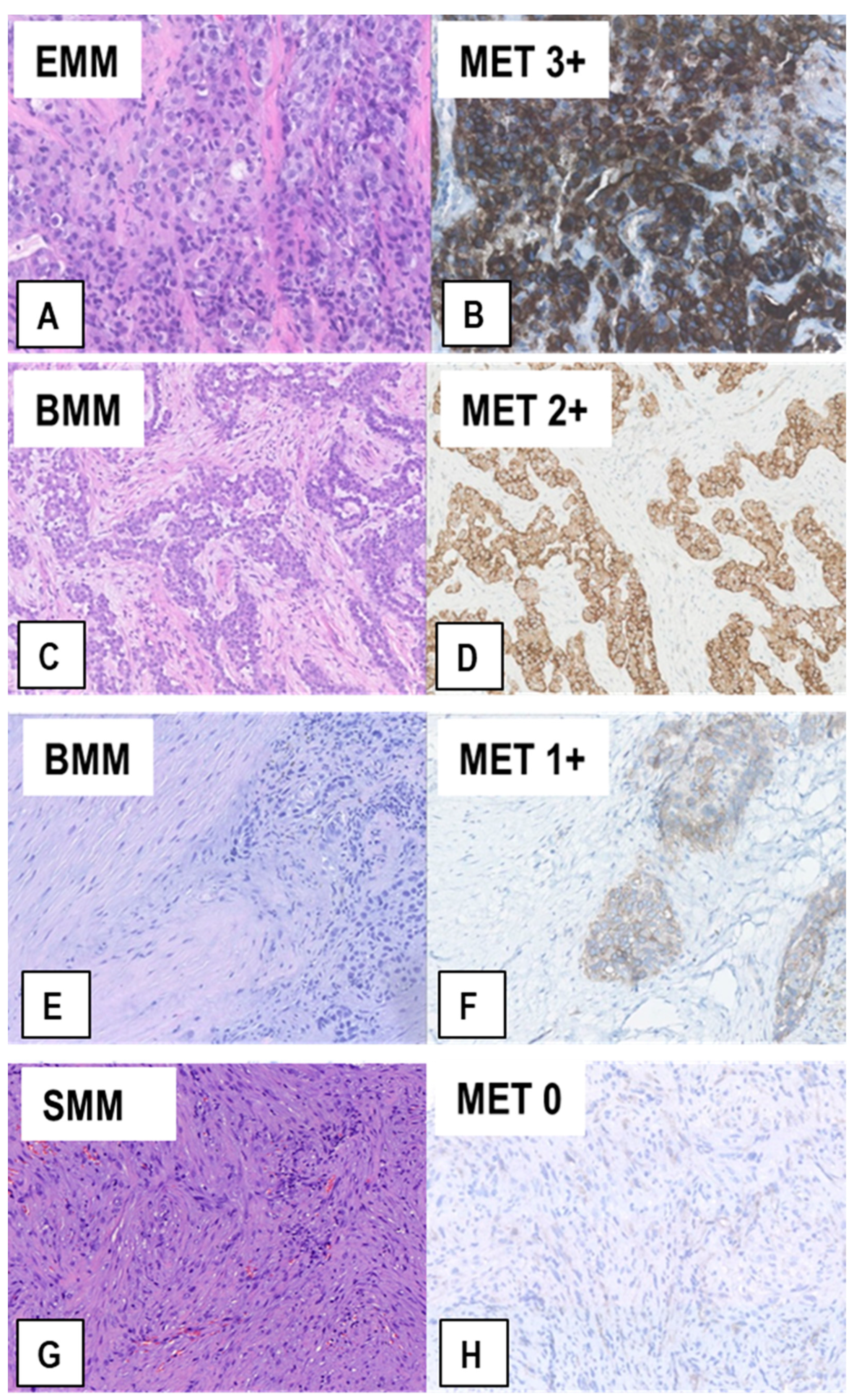
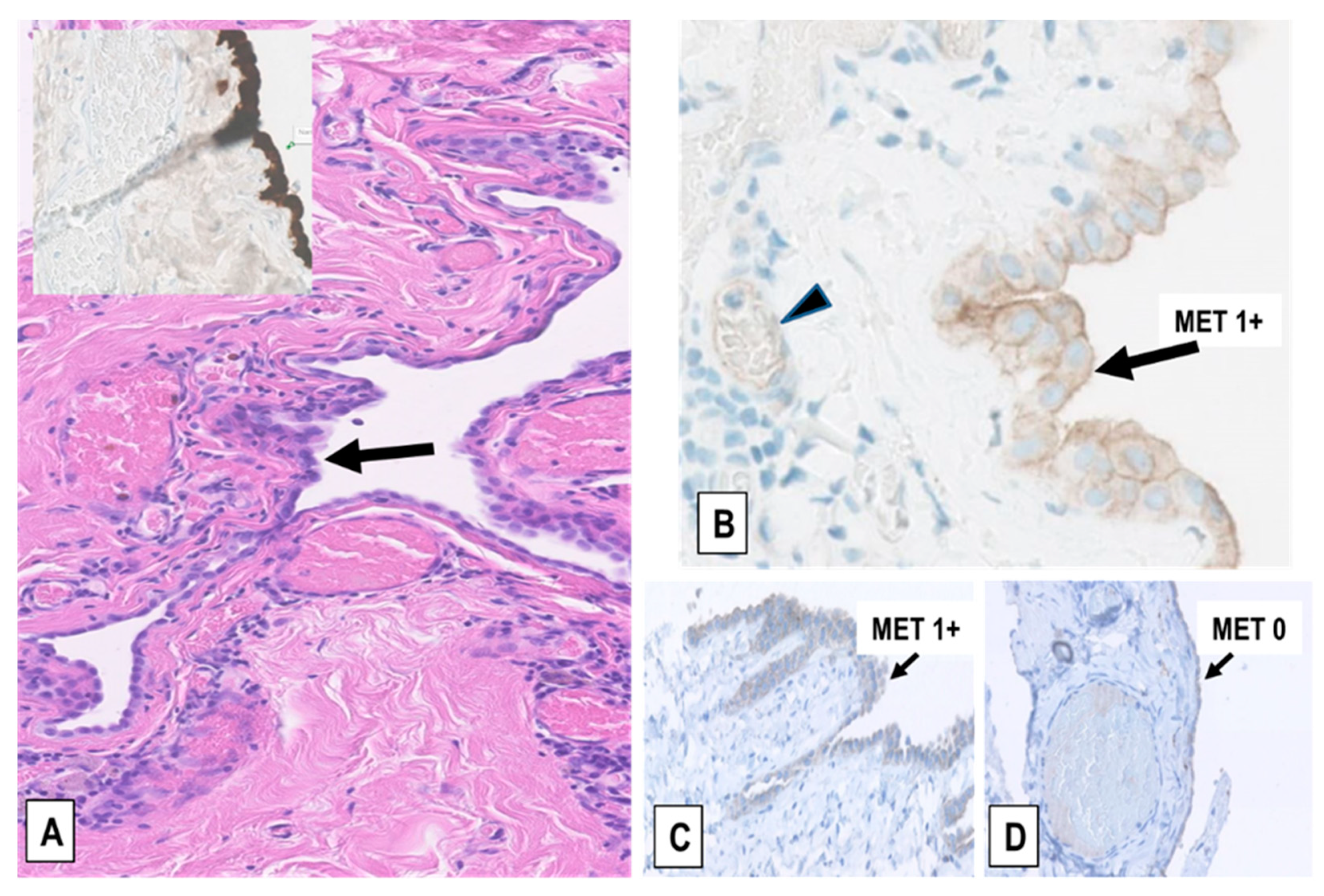
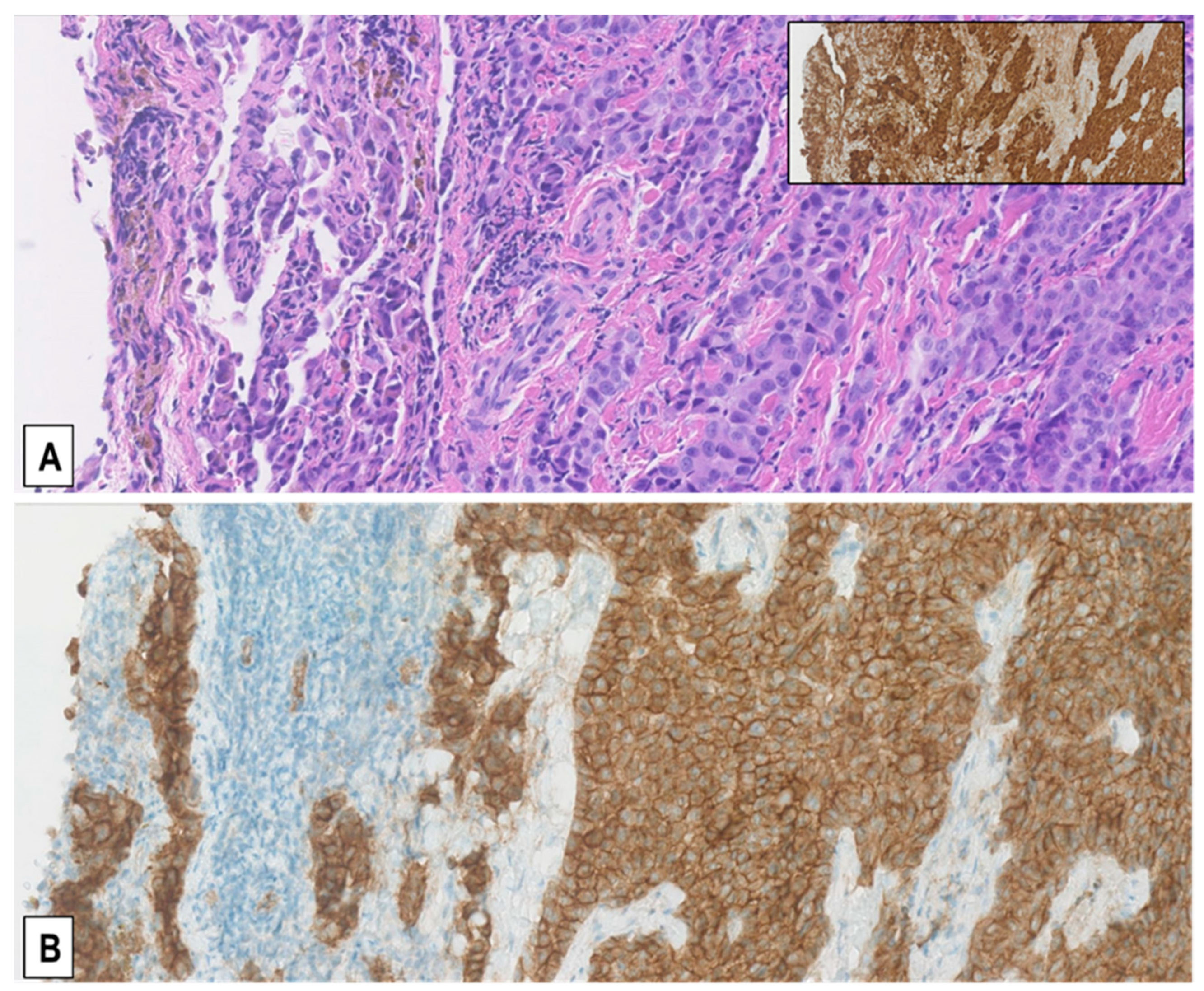
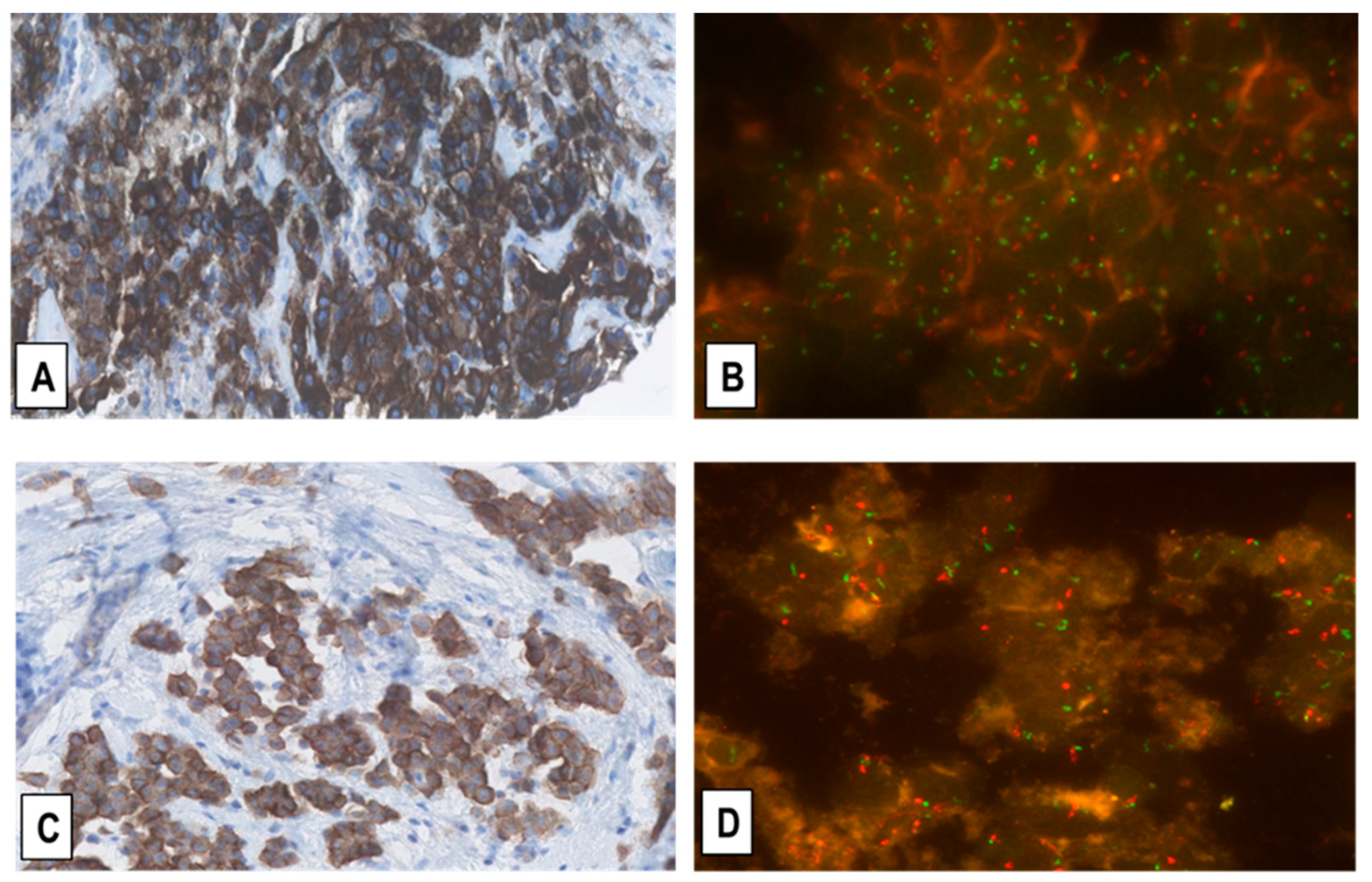
2.2. Expression of MET-Receptor in MM Assessed by IHC
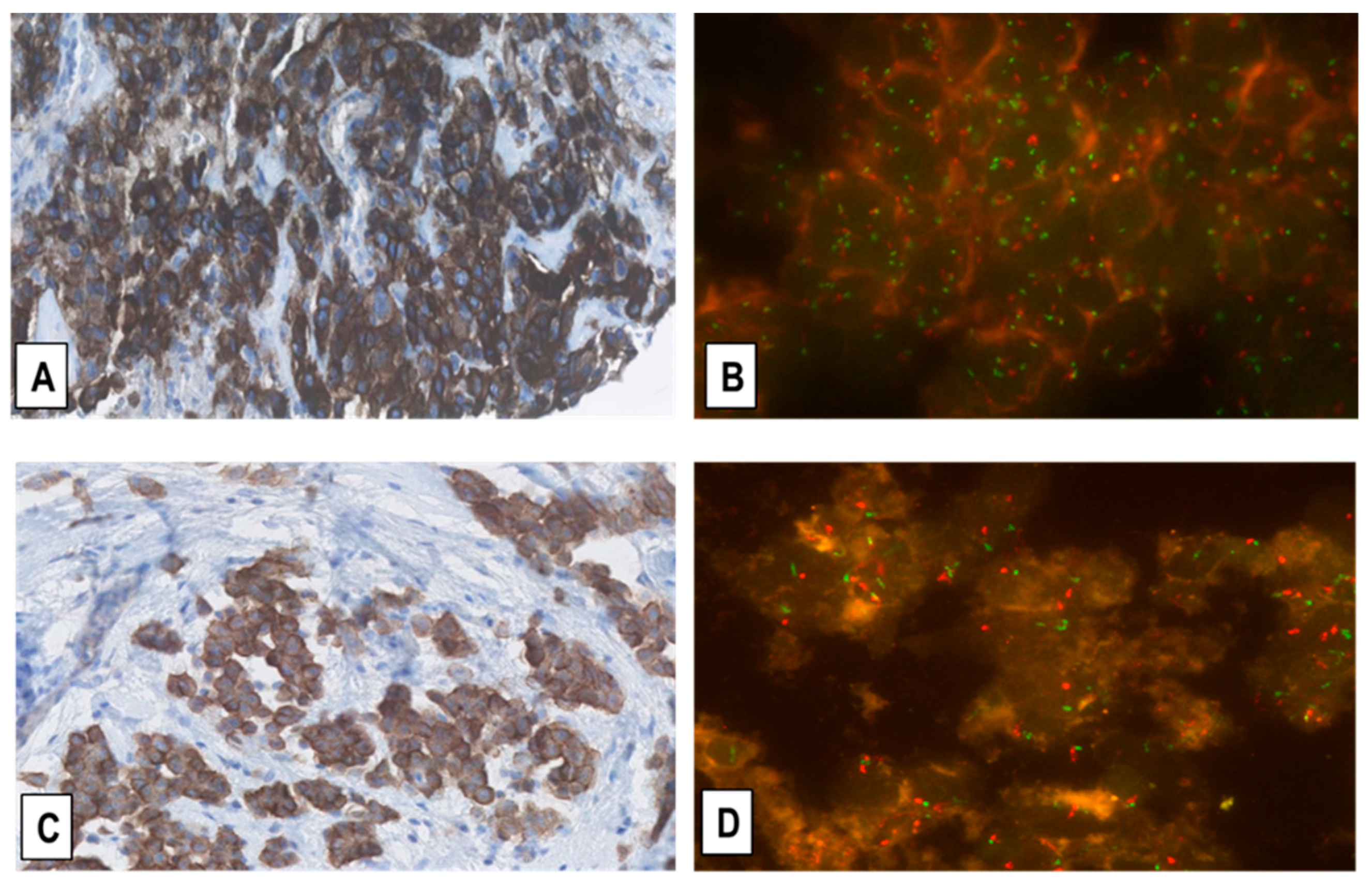
2.3. Assessment of MET-GCNG/GA in MM by FISH
2.4. MET Overexpression in MM Specimens Is Not Associated with MET Exon 14 Skipping Mutations
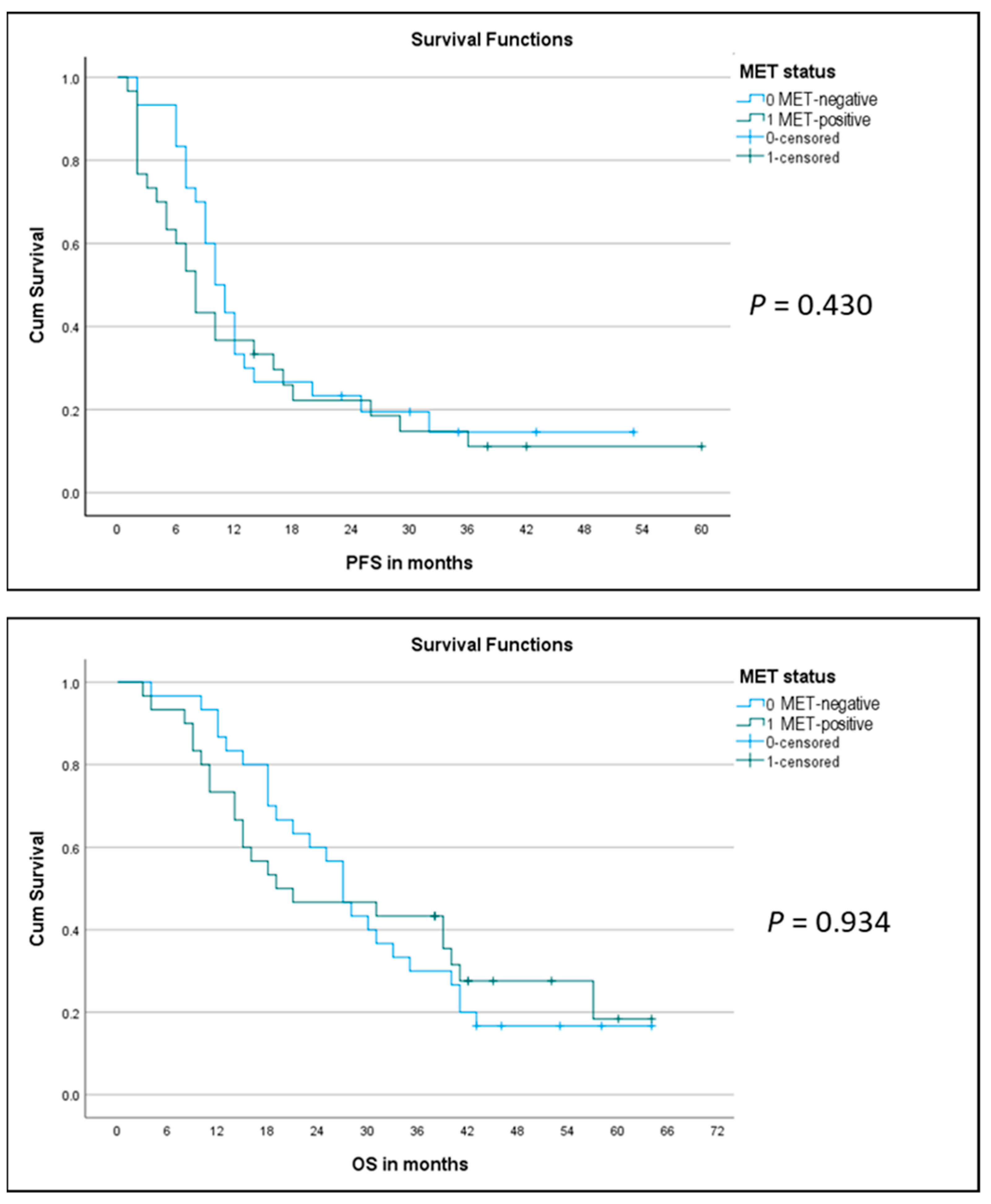
2.5. Correlation of MET Overexpression with Patient Outcome
3. Discussion
4. Materials and Methods
4.1. Tissue Samples
4.2. Immunohistochemistry
4.3. Fluorescence In Situ Hybridization
4.4. RNA Next-Generation Sequencing of MET Exon 14 Skipping Mutations
4.5. Correlation of MET-Amplification and Expression with Treatment Outcome
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Comoglio, P.M.; Trusolino, L.; Boccaccio, C. Known and novel roles of the MET oncogene in cancer: A coherent approach to targeted therapy. Nat. Rev. Cancer 2018, 18, 341–358. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Jain, P.; Wang, F.; Ma, P.C.; Borczuk, A.; Halmos, B. MET alterations and their impact on the future of non-small cell lung cancer (NSCLC) targeted therapies. Expert Opin. Ther. Targets 2021, 25, 249–268. [Google Scholar] [CrossRef] [PubMed]
- Miranda, O.; Farooqui, M.; Siegfried, J.M. Status of Agents Targeting the HGF/c-Met Axis in Lung Cancer. Cancers 2018, 10, 280. [Google Scholar] [CrossRef] [Green Version]
- Planchard, D. Have We Really MET a New Target? J. Clin. Oncol. 2018, 36, 3069–3071. [Google Scholar] [CrossRef] [PubMed]
- Garon, E.B.; Brodrick, P. Targeted Therapy Approaches for MET Abnormalities in Non-Small Cell Lung Cancer. Drugs 2021, 81, 547–554. [Google Scholar] [CrossRef]
- Carbone, M.; Adusumilli, P.S.; Alexander, H.R.; Baas, P.; Bardelli, F.; Bononi, A.; Bueno, R.; Felley-Bosco, E.; Galateau-Salle, F.; Jablons, D.; et al. Mesothelioma: Scientific Clues for Prevention, Diagnosis, and Therapy. CA Cancer J. Clin. 2019, 69, 402–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bueno, R.; Stawiski, E.W.; Goldstein, L.D.; Durinck, S.; De Rienzo, A.; Modrusan, Z.; Gnad, F.; Nguyen, T.T.; Jaiswal, B.S.; Chirieac, L.R.; et al. Comprehensive genomic analysis of malignant pleural mesothelioma identifies recurrent mutations, gene fusions and splicing alterations. Nat. Genet. 2016, 48, 407–416. [Google Scholar] [CrossRef]
- Hmeljak, J.; Sanchez-Vega, F.; Hoadley, K.A.; Shih, J.; Stewart, C.; Heiman, D.; Tarpey, P.; Danilova, L.; Drill, E.; Gibb, E.A.; et al. Integrative Molecular Characterization of Malignant Pleural Mesothelioma. Cancer Discov. 2018, 8, 1548–1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cakiroglu, E.; Senturk, S. Genomics and Functional Genomics of Malignant Pleural Mesothelioma. Int. J. Mol. Sci. 2020, 21, 6342. [Google Scholar] [CrossRef]
- Gaudino, G.; Yang, H.; Carbone, M. HGF/Met Signaling Is a Key Player in Malignant Mesothelioma Carcinogenesis. Biomedicines 2014, 2, 327–344. [Google Scholar] [CrossRef] [Green Version]
- Jean, D.; Daubriac, J.; Le Pimpec-Barthes, F.; Galateau-Salle, F.; Jaurand, M.C. Molecular changes in mesothelioma with an impact on prognosis and treatment. Arch. Pathol. Lab. Med. 2012, 136, 277–293. [Google Scholar] [CrossRef] [Green Version]
- Joseph, N.M.; Chen, Y.Y.; Nasr, A.; Yeh, I.; Talevich, E.; Onodera, C.; Bastian, B.C.; Rabban, J.T.; Garg, K.; Zaloudek, C.; et al. Genomic profiling of malignant peritoneal mesothelioma reveals recurrent alterations in epigenetic regulatory genes BAP1, SETD2, and DDX3X. Mod. Pathol. 2017, 30, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Levallet, G.; Vaisse-Lesteven, M.; Le Stang, N.; Ilg, A.G.; Brochard, P.; Astoul, P.; Pairon, J.C.; Bergot, E.; Zalcman, G.; Galateau-Sallé, F. Plasma cell membrane localization of c-MET predicts longer survival in patients with malignant mesothelioma: A series of 157 cases from the MESOPATH Group. J. Thorac. Oncol. 2012, 7, 599–606. [Google Scholar] [CrossRef] [Green Version]
- Bois, M.C.; Mansfield, A.S.; Sukov, W.R.; Jenkins, S.M.; Moser, J.C.; Sattler, C.A.; Smith, C.Y.; Molina, J.R.; Peikert, T.; Roden, A.C. c-Met expression and MET amplification in malignant pleural mesothelioma. Ann. Diagn. Pathol. 2016, 23, 1–7. [Google Scholar] [CrossRef]
- Salvi, S.; Varesano, S.; Boccardo, S.; Ravetti, J.L.; Canessa, P.A.; Pistillo, M.P.; Ferro, P.; Fedeli, F.; Roncella, S. FISH Analysis of Crizotinib Target Genes ROS1/ALK/MET in Malignant Mesothelioma. J. Thorac. Oncol. 2017, 12, e116–e118. [Google Scholar] [CrossRef]
- Schildhaus, H.U.; Schultheis, A.M.; Rüschoff, J.; Binot, E.; Merkelbach-Bruse, S.; Fassunke, J.; Schulte, W.; Ko, Y.D.; Schlesinger, A.; Bos, M.; et al. MET Amplification Status in Therapy-Naïve Adeno- and Squamous Cell Carcinomas of the Lung. Clin. Cancer Res. 2015, 21, 907–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jakobsen, J.N.; Santoni-Rugiu, E.; Grauslund, M.; Melchior, L.; Sørensen, J.B. Concomitant driver mutations in advanced EGFR-mutated non-small-cell lung cancer and their impact on erlotinib treatment. Oncotarget 2018, 9, 26195–26208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A Web Server for Cancer and Normal Gene Expression Profiling and Interactive Analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [Green Version]
- GEPIA. Available online: http://gepia.cancer-pku.cn/detail.php?gene=MET (accessed on 3 July 2021).
- Andersen, M.; Grauslund, M.; Ravn, J.; Sørensen, J.B.; Andersen, C.B.; Santoni-Rugiu, E. Diagnostic potential of miR-126, miR-143, miR-145, and miR-652 in malignant pleural mesothelioma. J. Mol. Diagn. 2014, 16, 418–430. [Google Scholar] [CrossRef]
- Andersen, M.; Grauslund, M.; Muhammad-Ali, M.; Ravn, J.; Sørensen, J.B.; Andersen, C.B.; Santoni-Rugiu, E. Are differentially expressed microRNAs useful in the diagnostics of malignant pleural mesothelioma? APMIS 2012, 120, 767–769. [Google Scholar] [CrossRef] [PubMed]
- Andersen, M.; Trapani, D.; Ravn, J.; Sørensen, J.B.; Andersen, C.B.; Grauslund, M.; Santoni-Rugiu, E. Methylation-associated Silencing of microRNA-126 and its Host Gene EGFL7 in Malignant Pleural Mesothelioma. Anticancer Res. 2015, 35, 6223–6229. [Google Scholar] [PubMed]
- Schuler, M.; Berardi, R.; Lim, W.T.; de Jonge, M.; Bauer, T.M.; Azaro, A.; Gottfried, M.; Han, J.Y.; Lee, D.H.; Wollner, M.; et al. Molecular correlates of response to capmatinib in advanced non-small-cell lung cancer: Clinical and biomarker results from a phase I trial. Ann. Oncol. 2020, 31, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.; Seto, T.; Han, J.Y.; Reguart, N.; Garon, E.B.; Groen, H.J.M.; Tan, D.S.W.; Hida, T.; de Jonge, M.; Orlov, S.V.; et al. Capmatinib in MET Exon 14-Mutated or MET-Amplified Non-Small-Cell Lung Cancer. N. Engl. J. Med. 2020, 383, 944–957. [Google Scholar] [CrossRef]
- Overbeck, T.R.; Cron, D.A.; Schmitz, K.; Rittmeyer, A.; Korber, W.; Hugo, S.; Schnalke, J.; Lukat, L.; Hugo, T.; Hinterthaner, M.; et al. Top-level MET gene copy number gain defines a subtype of poorly differentiated pulmonary adenocarcinomas with poor prognosis. Transl. Lung Cancer Res. 2020, 9, 603–616. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. Novartis Pharmaceuticals; Clinical Study of Oral cMET Inhibitor INC280 in Adult Patients with EGFR Wild-Type Advanced Non-Small Cell Lung Cancer. 2018. Available online: https://clinicaltrials.gov/ct2/show/NCT02414139 (accessed on 3 October 2021).
- Travis, W.D.; Brambilla, E.; Burke, A.P.; Marx, A.; Nicholson, A.G. WHO Classification of Tumours of the Lung, Pleura, Thymus and Heart, 4th ed.; International Agency for Research on Cancer (IARC): Lyon, France, 2015; pp. 153–181. [Google Scholar]
- Husain, A.N.; Colby, T.V.; Ordóñez, N.G.; Allen, T.C.; Attanoos, R.L.; Beasley, M.B.; Butnor, K.J.; Chirieac, L.R.; Churg, A.M.; Dacic, S.; et al. Guidelines for Pathologic Diagnosis of Malignant Mesothelioma 2017 Update of the Consensus Statement From the International Mesothelioma Interest Group. Arch. Pathol. Lab. Med. 2018, 142, 89–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapel, D.B.; Churg, A.; Santoni-Rugiu, E.; Tsujimura, T.; Hiroshima, K.; Husain, A.N. Molecular pathways and diagnosis in malignant mesothelioma: A review of the 14th International Conference of the International Mesothelioma Interest Group. Lung Cancer 2019, 127, 69–75. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | MPM P/D (n = 45) | MM Biopsy (n = 110) | RMP (n = 33) |
|---|---|---|---|
| Gender | |||
| Male (%) | 39 (87) | 88 (80) | 25 (76) |
| Female (%) | 6 (13) | 22 (20) | 8 (24) |
| Age, mean (range) | 68 (40–74) | 70 (40–86) | 39 (17–79) |
| Smoking history | |||
| Ex/current smoker (%) | 29 (64) | 83 (75) | 20 (60) |
| Non-smoker (%) | 16 (36) | 27 (25) | 13 (40) |
| Asbestos exposure | |||
| Yes (%) | 34 (76) | 77 (70) | 0 |
| No (%) | 11 (24) | 33 (30) | 33 (100) |
| Chemotherapy (cisplatin + pemetrexed) (%) | 45 (100) | 0 | 0 |
| Histological type | |||
| Epithelioid (%) | 20 (44) | 57 (52) | |
| Biphasic (%) | 25 (56) | 43 (39) | |
| Sarcomatoid (%) | 0 | 10 (9) | |
| Site | |||
| Pleura (%) | 45 (100) | 97 (88) | 33 (100) |
| Peritoneum (%) | 13 (12) |
| Hazard Ratio (HR) | 95% CI (HR) | p-Value | |
|---|---|---|---|
| MET status | 1.240 | 0.714–2.153 | 0.446 |
| Gender | 0.419 | 0.211–0.833 | * 0.013 |
| Histologic type | 1.012 | 0.552–1.854 | 0.970 |
| Asbestos exposure | 0.808 | 0.429–1.523 | 0.510 |
| Smoking status | 0.946 | 0.532–1.682 | 0.851 |
| Stage | 3.336 | 1.366–8.147 | * 0.008 |
| Performance status | 1.387 | 0.590–3.259 | 0.454 |
| Pleurectomy (P/D) | 0.508 | 0.290–0.892 | * 0.018 |
| Age | 1.005 | 0.970–1.041 | 0.774 |
| Hazard Ratio (HR) | 95% CI (HR) | p-Value | |
|---|---|---|---|
| MET status | 0.976 | 0.549–1.735 | 0.935 |
| Gender | 0.407 | 0.205–0.809 | * 0.010 |
| Histologic type | 0.931 | 0.489–1.773 | 0.828 |
| Asbestos exposure | 0.962 | 0.499–1.855 | 0.909 |
| Smoking status | 0.970 | 0.527–1.783 | 0.921 |
| Stage | 2.659 | 1.108–6.382 | * 0.029 |
| Performance status | 2.174 | 0.916–5.162 | 0.078 |
| Pleurectomy (P/D) | 0.385 | 0.213–0.694 | * 0.002 |
| Age | 1.024 | 0.987–1.061 | 0.206 |
| Hazard Ratio (HR) | 95% CI (HR) | p-Value | |
|---|---|---|---|
| MET status | 1.356 | 0.763–2.413 | 0.300 |
| Gender | 0.299 | 0.128–0.697 | * 0.005 |
| Histologic type | 1.278 | 0.645–2.531 | 0.483 |
| Asbestos exposure | 1.038 | 0.495–2.176 | 0.921 |
| Smoking status | 1.460 | 0.741–2.878 | 0.274 |
| Stage | 3.911 | 1.364–11.215 | * 0.011 |
| Performance status | 2.110 | 0.770–5.779 | 0.146 |
| Pleurectomy (P/D) | 0.576 | 0.286–1.160 | 0.123 |
| Age | 0.976 | 0.938–1.015 | 0.226 |
| Hazard Ratio (HR) | 95% CI (HR) | p-Value | |
|---|---|---|---|
| MET status | 1.051 | 0.563–1.962 | 0.877 |
| Gender | 0.223 | 0.085–0.586 | * 0.002 |
| Histologic type | 1.439 | 0.688–3.009 | 0.334 |
| Asbestos exposure | 1.923 | 0.825–4.478 | 0.130 |
| Smoking status | 1.551 | 0.723–3.325 | 0.260 |
| Stage | 3.319 | 1.045–9.425 | * 0.041 |
| Performance status | 2.110 | 0.770–5.779 | 0.146 |
| Pleurectomy (P/D) | 0.471 | 0.217–1.024 | 0.057 |
| Age | 0.980 | 0.937–1.025 | 0.375 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santoni-Rugiu, E.; Lü, M.J.S.; Jakobsen, J.N.; Melchior, L.C.; Ravn, J.; Sørensen, J.B. Correlation of MET-Receptor Overexpression with MET Gene Amplification and Patient Outcome in Malignant Mesothelioma. Int. J. Mol. Sci. 2021, 22, 12868. https://doi.org/10.3390/ijms222312868
Santoni-Rugiu E, Lü MJS, Jakobsen JN, Melchior LC, Ravn J, Sørensen JB. Correlation of MET-Receptor Overexpression with MET Gene Amplification and Patient Outcome in Malignant Mesothelioma. International Journal of Molecular Sciences. 2021; 22(23):12868. https://doi.org/10.3390/ijms222312868
Chicago/Turabian StyleSantoni-Rugiu, Eric, Maya Jeje Schuang Lü, Jan Nyrop Jakobsen, Linea Cecilie Melchior, Jesper Ravn, and Jens Benn Sørensen. 2021. "Correlation of MET-Receptor Overexpression with MET Gene Amplification and Patient Outcome in Malignant Mesothelioma" International Journal of Molecular Sciences 22, no. 23: 12868. https://doi.org/10.3390/ijms222312868
APA StyleSantoni-Rugiu, E., Lü, M. J. S., Jakobsen, J. N., Melchior, L. C., Ravn, J., & Sørensen, J. B. (2021). Correlation of MET-Receptor Overexpression with MET Gene Amplification and Patient Outcome in Malignant Mesothelioma. International Journal of Molecular Sciences, 22(23), 12868. https://doi.org/10.3390/ijms222312868