
Dermal Olfactory Receptor OR51B5 Is Essential for Survival and Collagen Synthesis in Human Dermal Fibroblast (Hs68 Cells)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
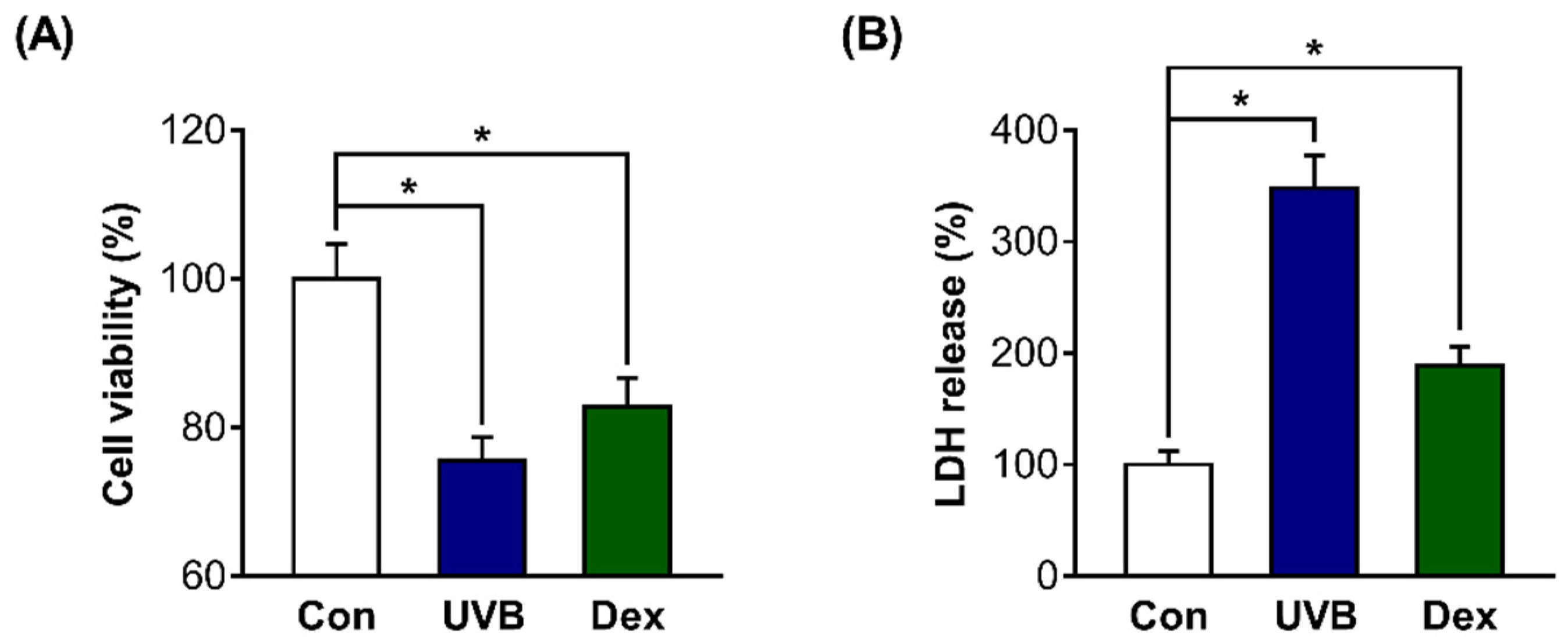
2.1. Inhibitory Effects of UVB or Dexamethasone on Hs68 Cell Viability
2.2. Inhibitory Effects of UVB or Dex on Collagen Synthesis in Hs68 Cells
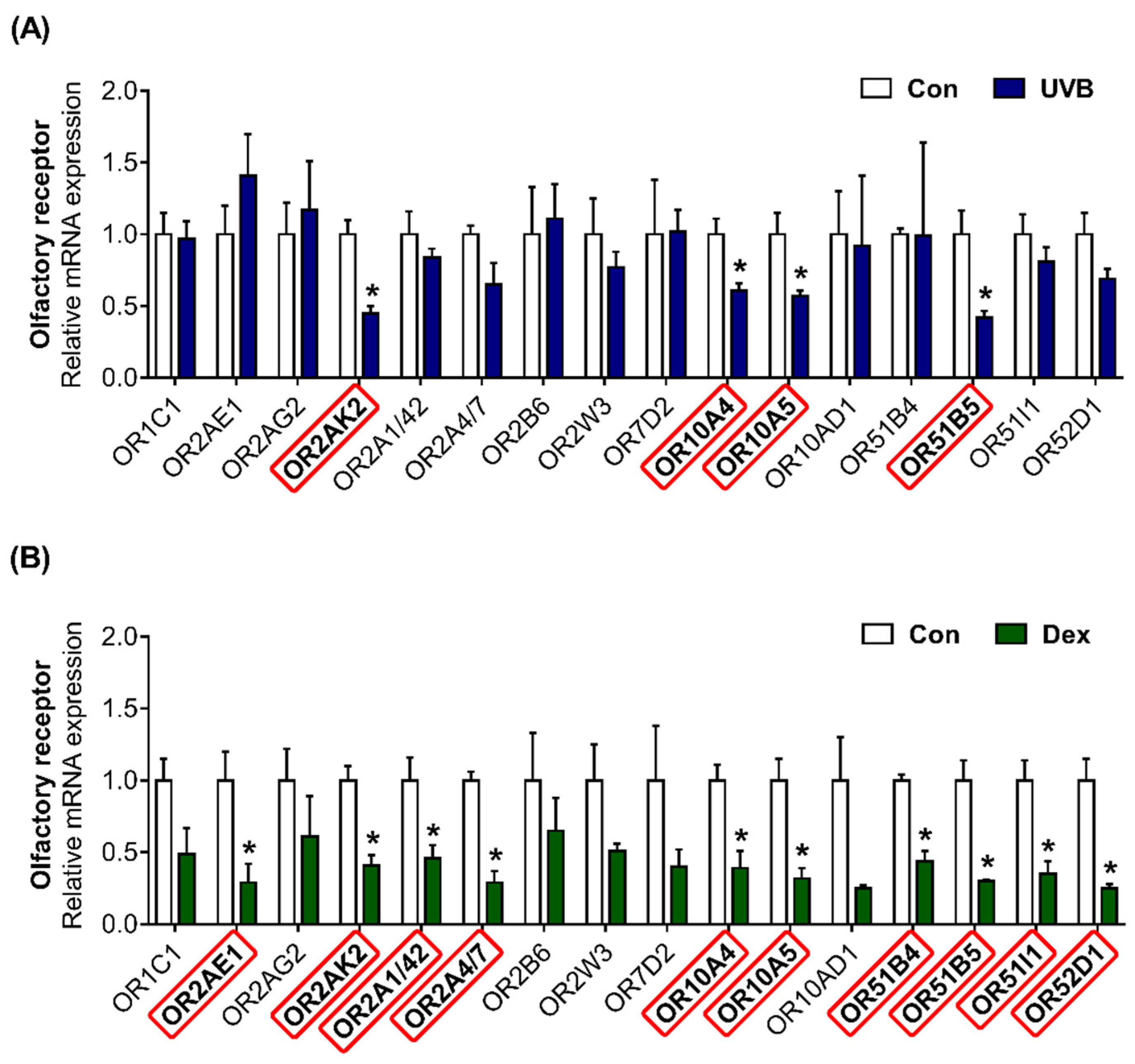
2.3. Altered Patterns of OR Gene Expression in UVB-Irradiated or Dexamethasone-Induced Hs68 Cells
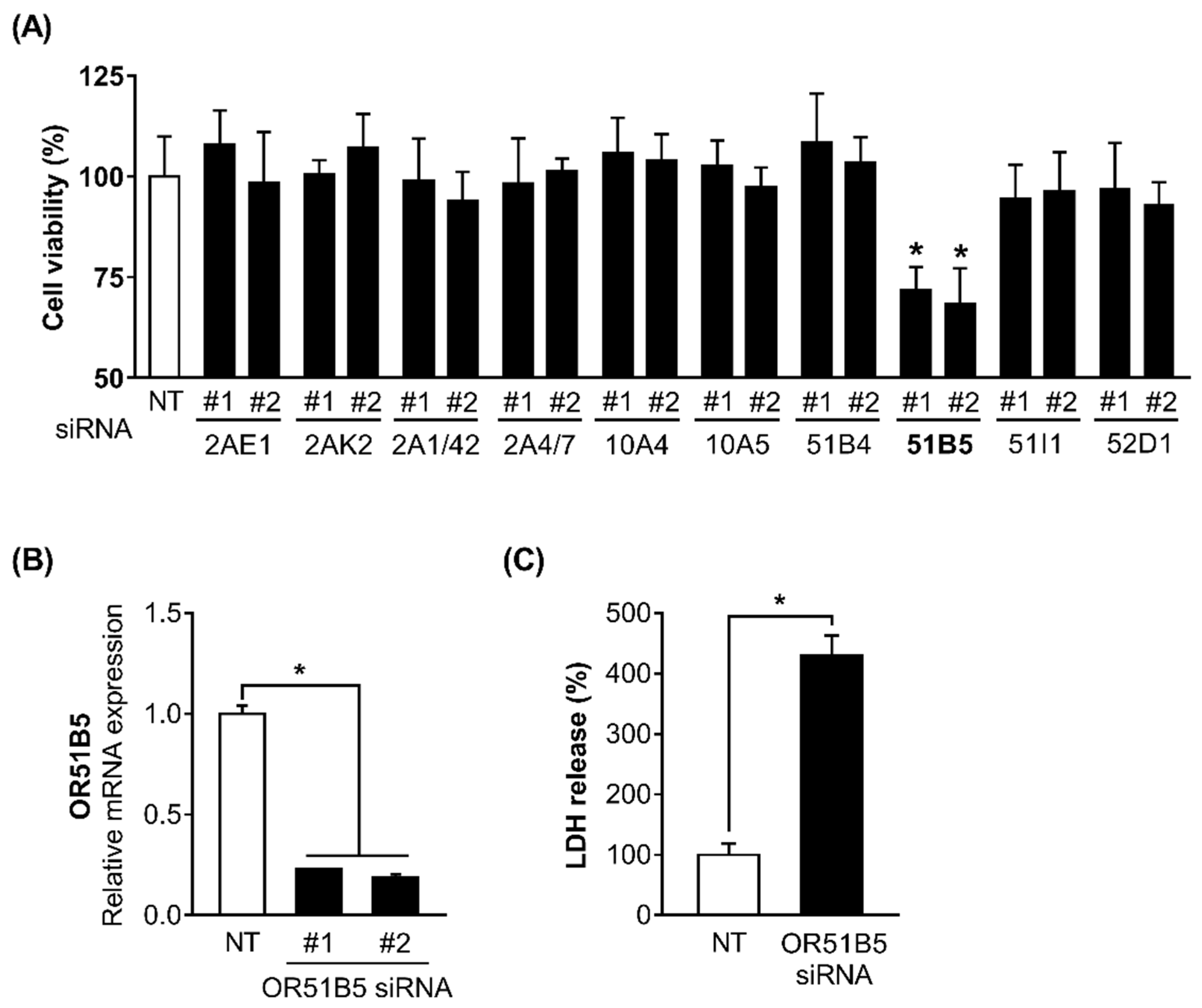
2.4. OR51B5 Knockdown Decreased Hs68 Cell Viability
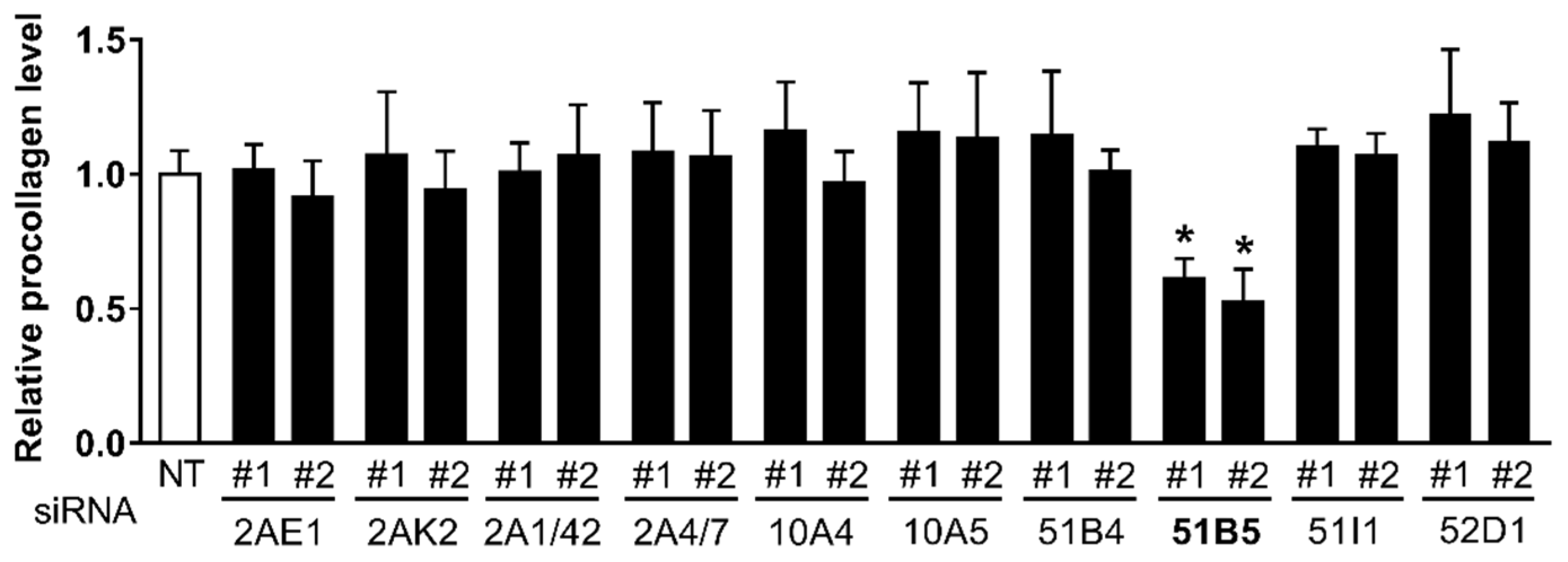
2.5. OR51B5 Knockdown Decreased Collagen Synthesis in Hs68 Cells
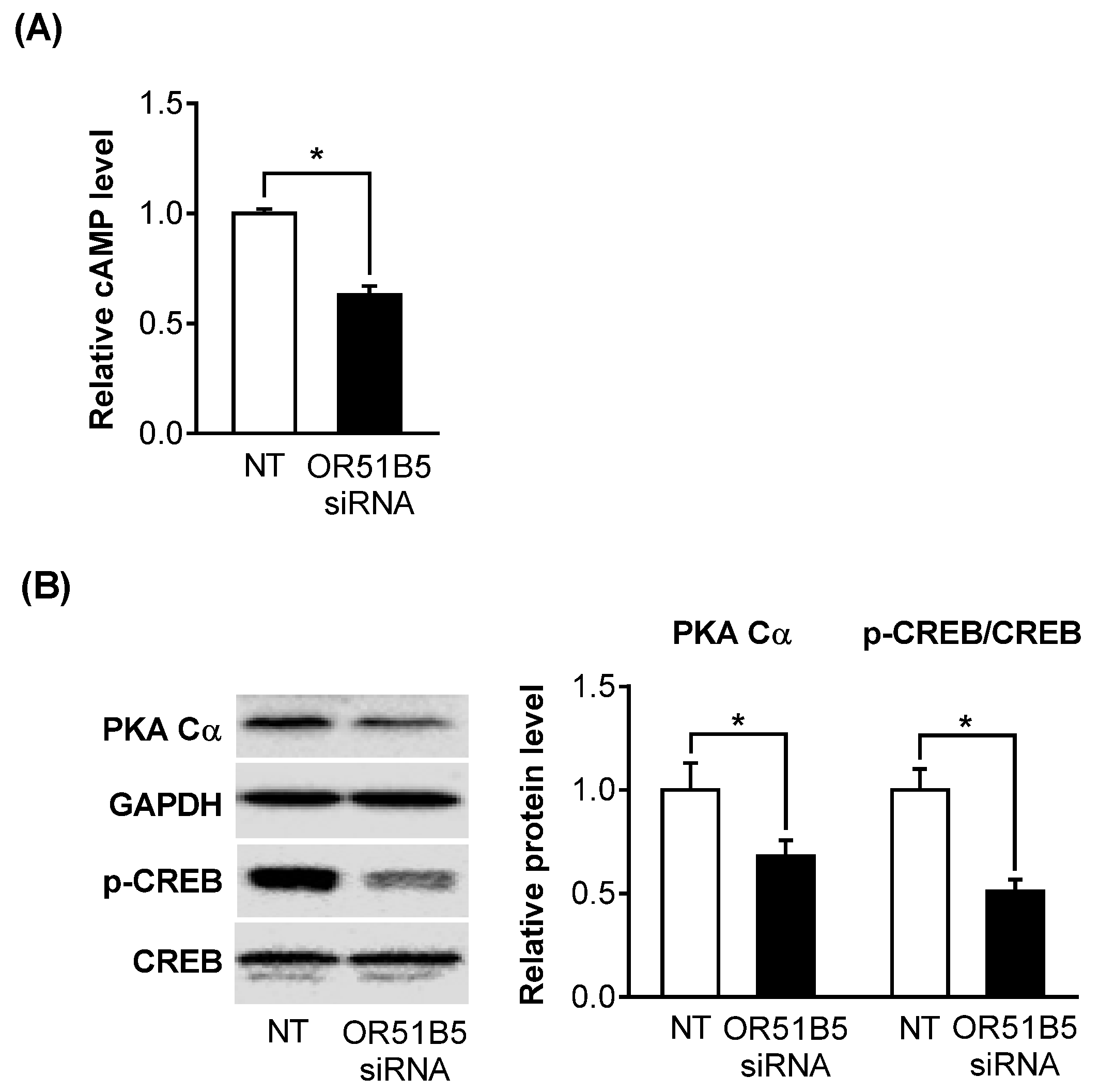
2.6. OR51B5 Knockdown Inhibited Olfactory Signaling Pathway in Hs68 Cells
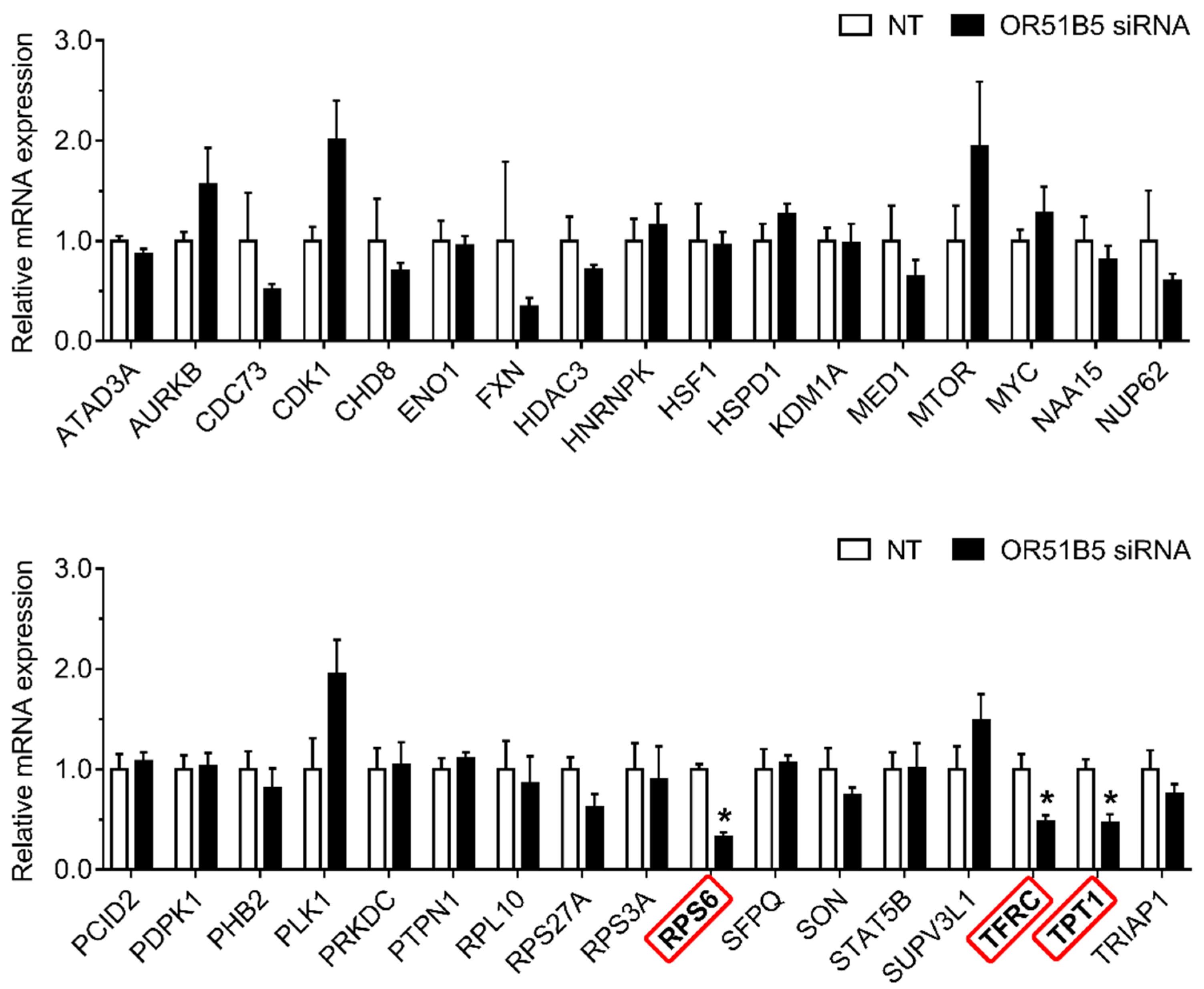
2.7. OR51B5 Knockdown Decreased Cell Survival-Related Gene Expression in Hs68 Cells
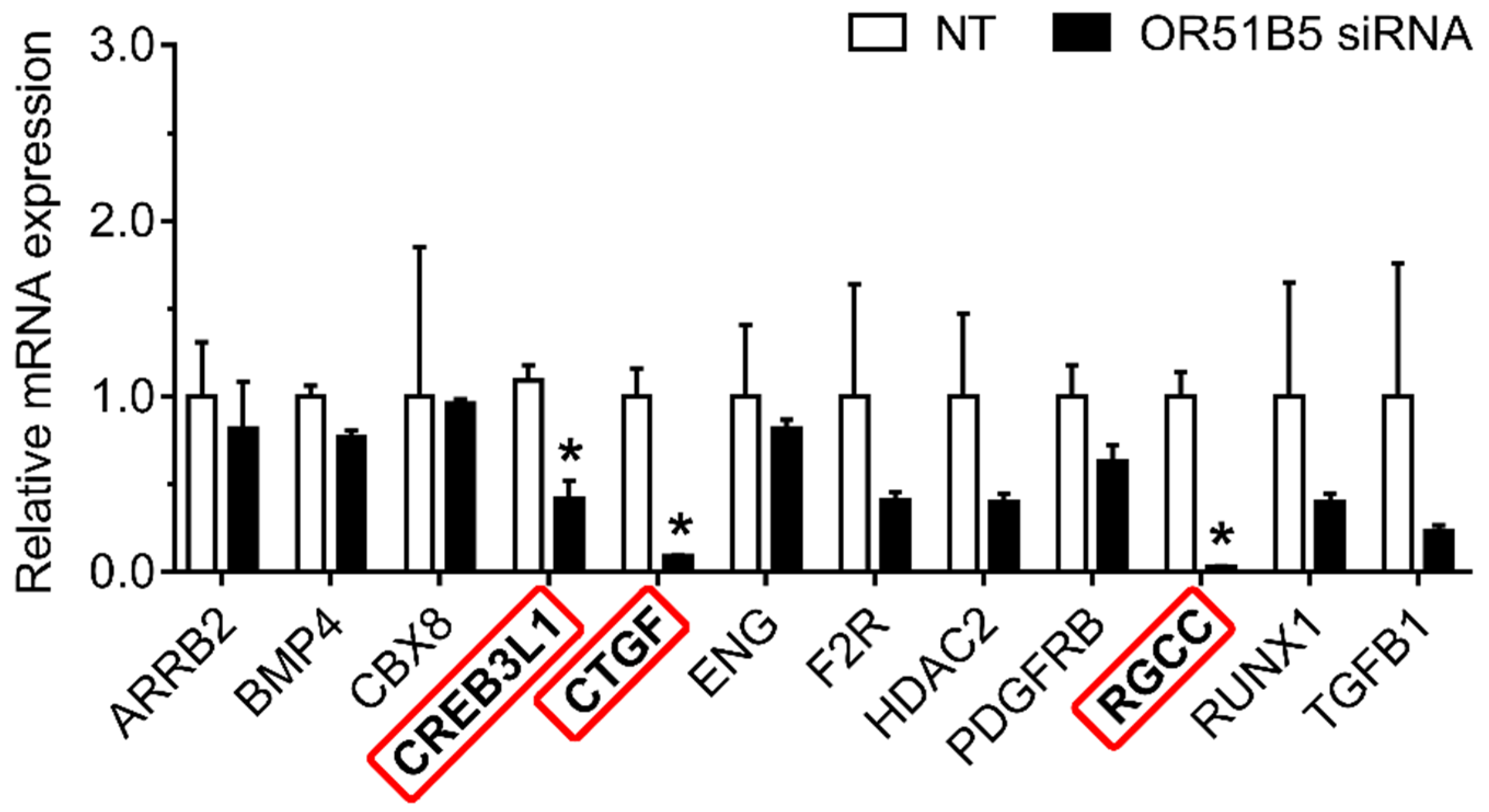
2.8. OR51B5 Knockdown Decreased Collagen Synthesis-Related Gene Expression in Hs68 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Cell Treatment
4.3. WST-1 Assay
4.4. LDH Assay
4.5. Senescence-Associated β-Galactosidase (SA-β-gal) Staining
4.6. Procollagen Type I C-Peptide Measurement
4.7. RNA Extraction, cDNA Synthesis, and Polymerase Chain Reaction (PCR)
4.8. siRNA Screening
4.9. cAMP Measurement
4.10. Western Blot
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Aust, M.C.; Fernandes, D.; Kolokythas, P.; Kaplan, H.M.; Vogt, P.M. Percutaneous collagen induction therapy: An alternative treatment for scars, wrinkles, and skin laxity. Plast. Reconstr. Surg. 2008, 121, 1421–1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quan, T.; Little, E.; Quan, H.; Voorhees, J.J.; Fisher, G.J. Elevated matrix metalloproteinases and collagen fragmentation in photodamaged human skin: Impact of altered extracellular matrix microenvironment on dermal fibroblast function. J. Investig. Dermatol. 2013, 133, 1362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cole, M.A.; Quan, T.; Voorhees, J.J.; Fisher, G.J. Extracellular matrix regulation of fibroblast function: Redefining our perspective on skin aging. J. Cell Commun. Signal. 2018, 12, 35–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, L.; Mauro, T.M.; Dang, E.; Man, G.; Zhang, J.; Lee, D.; Wang, G.; Feingold, K.R.; Elias, P.M.; Man, M.-Q. Epidermal dysfunction leads to an age-associated increase in levels of serum inflammatory cytokines. J. Investig. Dermatol. 2017, 137, 1277–1285. [Google Scholar] [CrossRef] [Green Version]
- Flament, F.; Bazin, R.; Laquieze, S.; Rubert, V.; Simonpietri, E.; Piot, B. Effect of the sun on visible clinical signs of aging in Caucasian skin. Clin. Cosmet. Investig. Dermatol. 2013, 6, 221. [Google Scholar] [CrossRef] [Green Version]
- Khmaladze, I.; Leonardi, M.; Fabre, S.; Messaraa, C.; Mavon, A. The Skin Interactome: A Holistic “Genome-Microbiome-Exposome” Approach to Understand and Modulate Skin Health and Aging. Clin. Cosmet. Investig. Dermatol. 2020, 13, 1021. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Duan, E. Fighting against skin aging: The way from bench to bedside. Cell Transplant. 2018, 27, 729–738. [Google Scholar] [CrossRef]
- Uitto, J. The role of elastin and collagen in cutaneous aging: Intrinsic aging versus photoexposure. J. Drugs Dermatol. 2008, 7, s12–s16. [Google Scholar]
- Tsatsou, F.; Trakatelli, M.; Patsatsi, A.; Kalokasidis, K.; Sotiriadis, D. Extrinsic aging: UV-mediated skin carcinogenesis. Dermato-Endocrinology 2012, 4, 285–297. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.Y.; Wang, P.W.; Huang, C.H.; Chen, M.H.; Wu, Y.R.; Pan, T.L. Skin aging caused by intrinsic or extrinsic processes characterized with functional proteomics. Proteomics 2016, 16, 2718–2731. [Google Scholar] [CrossRef]
- Weiner, F.; Czaja, M.; Jefferson, D.; Giambrone, M.; Tur-Kaspa, R.; Reid, L.; Zern, M.A. The effects of dexamethasone on in vitro collagen gene expression. J. Biol. Chem. 1987, 262, 6955–6958. [Google Scholar] [CrossRef]
- Slavin, J.; Unemori, E.; Hunt, T.; Amento, E. Transforming growth factor beta (TGF-β) and dexamethasone have direct opposing effects on collagen metabolism in low passage human dermal fibroblasts in vitro. Growth Factors 1994, 11, 205–213. [Google Scholar] [CrossRef]
- Choi, D.; Kang, W.; Park, S.; Son, B.; Park, T. β-Ionone Attenuates Dexamethasone-Induced Suppression of Collagen and Hyaluronic Acid Synthesis in Human Dermal Fibroblasts. Biomolecules 2021, 11, 619. [Google Scholar] [CrossRef]
- Jariashvili, K.; Madhan, B.; Brodsky, B.; Kuchava, A.; Namicheishvili, L.; Metreveli, N. UV damage of collagen: Insights from model collagen peptides. Biopolymers 2012, 97, 189–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fagot, D.; Asselineau, D.; Bernerd, F. Direct role of human dermal fibroblasts and indirect participation of epidermal keratinocytes in MMP-1 production after UV-B irradiation. Arch. Dermatol. Res. 2002, 293, 576–583. [Google Scholar] [CrossRef]
- Tong, T.; Ryu, S.E.; Min, Y.; Claire, A.; Bushdid, C.; Golebiowski, J.; Moon, C.; Park, T. Olfactory receptor 10J5 responding to α-cedrene regulates hepatic steatosis via the cAMP–PKA pathway. Sci. Rep. 2017, 7, 9471. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Zhao, H.; Fu, N.; Chen, L. The diversified function and potential therapy of ectopic olfactory receptors in non-olfactory tissues. J. Cell. Physiol. 2018, 233, 2104–2115. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Zhang, K.; Tong, T.; Park, T. Improved Glucose Intolerance through a Distinct Mouse Olfactory Receptor 23-Induced Signaling Pathway Mediating Glucose Uptake in Myotubes and Adipocytes. Mol. Nutr. Food Res. 2020, 64, 1901329. [Google Scholar] [CrossRef] [PubMed]
- Flegel, C.; Manteniotis, S.; Osthold, S.; Hatt, H.; Gisselmann, G. Expression profile of ectopic olfactory receptors determined by deep sequencing. PLoS ONE 2013, 8, e55368. [Google Scholar] [CrossRef] [Green Version]
- Tham, E.H.; Dyjack, N.; Kim, B.E.; Rios, C.; Seibold, M.A.; Leung, D.Y.; Goleva, E. Expression and function of the ectopic olfactory receptor OR10G7 in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2019, 143, 1838–1848.e4. [Google Scholar] [CrossRef]
- Morita, R.; Hirohashi, Y.; Torigoe, T.; Ito-Inoda, S.; Takahashi, A.; Mariya, T.; Asanuma, H.; Tamura, Y.; Tsukahara, T.; Kanaseki, T. Olfactory receptor family 7 subfamily C member 1 is a novel marker of colon cancer–initiating cells and is a potent target of immunotherapy. Clin. Cancer Res. 2016, 22, 3298–3309. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Lee, J.Y.; Song, K.C.; Kim, J.; Park, J.H.; Chun, K.-H.; Hwang, G.S. Protective effect of processed Panax ginseng, sun ginseng on UVB-irradiated human skin keratinocyte and human dermal fibroblast. J. Ginseng Res. 2012, 36, 68. [Google Scholar] [CrossRef] [Green Version]
- Fang, Y.; Hu, X.H.; Jia, Z.G.; Xu, M.H.; Guo, Z.Y.; Gao, F.H. Tiron protects against UVB-induced senescence-like characteristics in human dermal fibroblasts by the inhibition of superoxide anion production and glutathione depletion. Australas. J. Dermatol. 2012, 53, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Brenner, R.; Felger, D.; Winter, C.; Christiansen, A.; Hofmann, D.; Bartmann, P. Effects of dexamethasone on proliferation, chemotaxis, collagen I, and fibronectin-metabolism of human fetal lung fibroblasts. Pediatric Pulmonol. 2001, 32, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bose, R.; Moors, M.; Tofighi, R.; Cascante, A.; Hermanson, O.; Ceccatelli, S. Glucocorticoids induce long-lasting effects in neural stem cells resulting in senescence-related alterations. Cell Death Dis. 2010, 1, e92. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Li, Q.; Yan, H.; Jiao, G.; Wang, H.; Chi, H.; Zhou, H.; Chen, L.; Shan, Y.; Chen, Y. Resveratrol Protects Osteoblasts Against Dexamethasone-Induced Cytotoxicity Through Activation of AMP-Activated Protein Kinase. Drug Des. Dev. Ther. 2020, 14, 4451. [Google Scholar] [CrossRef]
- Morgens, D.W.; Deans, R.M.; Li, A.; Bassik, M.C. Systematic comparison of CRISPR/Cas9 and RNAi screens for essential genes. Nat. Biotechnol. 2016, 34, 634–636. [Google Scholar] [CrossRef] [Green Version]
- Consortium, G.O. The Gene Ontology (GO) database and informatics resource. Nucleic Acids Res. 2004, 32, D258–D261. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Liu, W.; Zhang, H.-M.; Xie, G.-Y.; Miao, Y.-R.; Xia, M.; Guo, A.-Y. hTFtarget: A comprehensive database for regulations of human transcription factors and their targets. Genom. Proteom. Bioinform. 2020, 18, 120–128. [Google Scholar] [CrossRef]
- Ferreira, T.; Wilson, S.R.; Choi, Y.G.; Risso, D.; Dudoit, S.; Speed, T.P.; Ngai, J. Silencing of odorant receptor genes by G protein βγ signaling ensures the expression of one odorant receptor per olfactory sensory neuron. Neuron 2014, 81, 847–859. [Google Scholar] [CrossRef] [Green Version]
- Von Dannecker, L.E.C.; Mercadante, A.F.; Malnic, B. Ric-8B, an olfactory putative GTP exchange factor, amplifies signal transduction through the olfactory-specific G-protein Gαolf. J. Neurosci. 2005, 25, 3793–3800. [Google Scholar] [CrossRef] [PubMed]
- Verbeurgt, C.; Wilkin, F.; Tarabichi, M.; Gregoire, F.; Dumont, J.E.; Chatelain, P. Profiling of olfactory receptor gene expression in whole human olfactory mucosa. PLoS ONE 2014, 9, e96333. [Google Scholar] [CrossRef] [PubMed]
- Reisert, J.; Yau, K.W.; Margolis, F.L. Olfactory marker protein modulates the cAMP kinetics of the odour-induced response in cilia of mouse olfactory receptor neurons. J. Physiol. 2007, 585, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Firestein, S.; Darrow, B.; Shepherd, G.M. Activation of the sensory current in salamander olfactory receptor neurons depends on a G protein-mediated cAMP second messenger system. Neuron 1991, 6, 825–835. [Google Scholar] [CrossRef]
- Moon, C.; Jaberi, P.; Otto-Bruc, A.; Baehr, W.; Palczewski, K.; Ronnett, G.V. Calcium-sensitive particulate guanylyl cyclase as a modulator of cAMP in olfactory receptor neurons. J. Neurosci. 1998, 18, 3195–3205. [Google Scholar] [CrossRef] [Green Version]
- Wetzel, C.H.; Spehr, M.; Hatt, H. Phosphorylation of voltage-gated ion channels in rat olfactory receptor neurons. Eur. J. Neurosci. 2001, 14, 1056–1064. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Son, B.; Park, S.; Choi, D.; Park, T. UV-Irradiation-and Inflammation-Induced Skin Barrier Dysfunction Is Associated with the Expression of Olfactory Receptor Genes in Human Keratinocytes. Int. J. Mol. Sci. 2021, 22, 2799. [Google Scholar] [CrossRef]
- Marcinek, P.; Geithe, C.; Krautwurst, D. Chemosensory G protein-coupled receptors (GPCR) in blood leukocytes. In Taste and Smell; Springer: Berlin, Germany, 2016; pp. 151–173. [Google Scholar]
- Manteniotis, S.; Wójcik, S.; Göthert, J.R.; Dürig, J.; Dührsen, U.; Gisselmann, G.; Hatt, H. Deorphanization and characterization of the ectopically expressed olfactory receptor OR51B5 in myelogenous leukemia cells. Cell Death Discov. 2016, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Tsai, T.; Veitinger, S.; Peek, I.; Busse, D.; Eckardt, J.; Vladimirova, D.; Jovancevic, N.; Wojcik, S.; Gisselmann, G.; Altmüller, J. Two olfactory receptors—OR 2A4/7 and OR 51B5—differentially affect epidermal proliferation and differentiation. Exp. Dermatol. 2017, 26, 58–65. [Google Scholar] [CrossRef]
- Uhlén, M.; Björling, E.; Agaton, C.; Szigyarto, C.A.-K.; Amini, B.; Andersen, E.; Andersson, A.-C.; Angelidou, P.; Asplund, A.; Asplund, C. A human protein atlas for normal and cancer tissues based on antibody proteomics. Mol. Cell. Proteom. 2005, 4, 1920–1932. [Google Scholar] [CrossRef] [Green Version]
- Hung, C.-F.; Fang, C.-L.; Al-Suwayeh, S.A.; Yang, S.-Y.; Fang, J.-Y. Evaluation of drug and sunscreen permeation via skin irradiated with UVA and UVB: Comparisons of normal skin and chronologically aged skin. J. Dermatol. Sci. 2012, 68, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Adil, M.D.; Kaiser, P.; Satti, N.K.; Zargar, A.M.; Vishwakarma, R.A.; Tasduq, S.A. Effect of Emblica officinalis (fruit) against UVB-induced photo-aging in human skin fibroblasts. J. Ethnopharmacol. 2010, 132, 109–114. [Google Scholar] [CrossRef]
- Debacq-Chainiaux, F.; Leduc, C.; Verbeke, A.; Toussaint, O. UV, stress and aging. Dermato-Endocrinology 2012, 4, 236–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drouin, R.; Therrien, J.P. UVB-induced cyclobutane pyrimidine dimer frequency correlates with skin cancer mutational hotspots in p53. Photochem. Photobiol. 1997, 66, 719–726. [Google Scholar] [CrossRef]
- Slominski, A.; Zbytek, B.; Semak, I.; Sweatman, T.; Wortsman, J. CRH stimulates POMC activity and corticosterone production in dermal fibroblasts. J. Neuroimmunol. 2005, 162, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Matthews, S.G.; Challis, J. Regulation of CRH and AVP mRNA in the developing ovine hypothalamus: Effects of stress and glucocorticoids. Am. J. Physiol.-Endocrinol. Metab. 1995, 268, E1096–E1107. [Google Scholar] [CrossRef] [PubMed]
- Munck, A.; Guyre, P.M. Glucocorticoid physiology, pharmacology and stress. In Steroid Hormone Resistance; Springer: Berlin, Germany, 1986; pp. 81–96. [Google Scholar]
- Siu, Y.T.; Jin, D.Y. CREB—A real culprit in oncogenesis. FEBS J. 2007, 274, 3224–3232. [Google Scholar] [CrossRef]
- Briand, L.A.; Lee, B.G.; Lelay, J.; Kaestner, K.H.; Blendy, J.A. Serine 133 phosphorylation is not required for hippocampal CREB-mediated transcription and behavior. Learn. Mem. 2015, 22, 109–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Odom, D.T.; Koo, S.-H.; Conkright, M.D.; Canettieri, G.; Best, J.; Chen, H.; Jenner, R.; Herbolsheimer, E.; Jacobsen, E. Genome-wide analysis of cAMP-response element binding protein occupancy, phosphorylation, and target gene activation in human tissues. Proc. Natl. Acad. Sci. USA 2005, 102, 4459–4464. [Google Scholar] [CrossRef] [Green Version]
- Cha-Molstad, H.; Keller, D.M.; Yochum, G.S.; Impey, S.; Goodman, R.H. Cell-type-specific binding of the transcription factor CREB to the cAMP-response element. Proc. Natl. Acad. Sci. USA 2004, 101, 13572–13577. [Google Scholar] [CrossRef] [Green Version]
- Carlezon, W.A., Jr.; Duman, R.S.; Nestler, E.J. The many faces of CREB. Trends Neurosci. 2005, 28, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Coven, E.; Ni, Y.; Widnell, K.L.; Chen, J.; Walker, W.H.; Habener, J.F.; Nestler, E.J. Cell type-specific regulation of CREB gene expression: Mutational analysis of CREB promoter activity. J. Neurochem. 1998, 71, 1865–1874. [Google Scholar] [CrossRef]
- Fang, C.-L.; Huang, L.-H.; Tsai, H.-Y.; Chang, H.-I. Dermal lipogenesis inhibits adiponectin production in human dermal fibroblasts while exogenous adiponectin administration prevents against UVA-induced dermal matrix degradation in human skin. Int. J. Mol. Sci. 2016, 17, 1129. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.; Lee, Y.S.; Kim, M.O.; Chang, M.Y.; Won, B.M.; Jin, B.S.; Park, S. Smad-induced alterations of matrix metabolism by a myristoyl tetra peptide. Cell Biochem. Funct. 2014, 32, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Choi, D.; Park, T. Decanal Protects against UVB-Induced Photoaging in Human Dermal Fibroblasts via the cAMP Pathway. Nutrients 2020, 12, 1214. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Son, B.; Kang, W.; Park, S.; Choi, D.; Park, T. Dermal Olfactory Receptor OR51B5 Is Essential for Survival and Collagen Synthesis in Human Dermal Fibroblast (Hs68 Cells). Int. J. Mol. Sci. 2021, 22, 9273. https://doi.org/10.3390/ijms22179273
Son B, Kang W, Park S, Choi D, Park T. Dermal Olfactory Receptor OR51B5 Is Essential for Survival and Collagen Synthesis in Human Dermal Fibroblast (Hs68 Cells). International Journal of Molecular Sciences. 2021; 22(17):9273. https://doi.org/10.3390/ijms22179273
Chicago/Turabian StyleSon, Bomin, Wesuk Kang, Soyoon Park, Dabin Choi, and Taesun Park. 2021. "Dermal Olfactory Receptor OR51B5 Is Essential for Survival and Collagen Synthesis in Human Dermal Fibroblast (Hs68 Cells)" International Journal of Molecular Sciences 22, no. 17: 9273. https://doi.org/10.3390/ijms22179273
APA StyleSon, B., Kang, W., Park, S., Choi, D., & Park, T. (2021). Dermal Olfactory Receptor OR51B5 Is Essential for Survival and Collagen Synthesis in Human Dermal Fibroblast (Hs68 Cells). International Journal of Molecular Sciences, 22(17), 9273. https://doi.org/10.3390/ijms22179273