
Identification of Novel RNA Binding Proteins Influencing Circular RNA Expression in Hepatocellular Carcinoma




Abstract
1. Introduction
2. Results
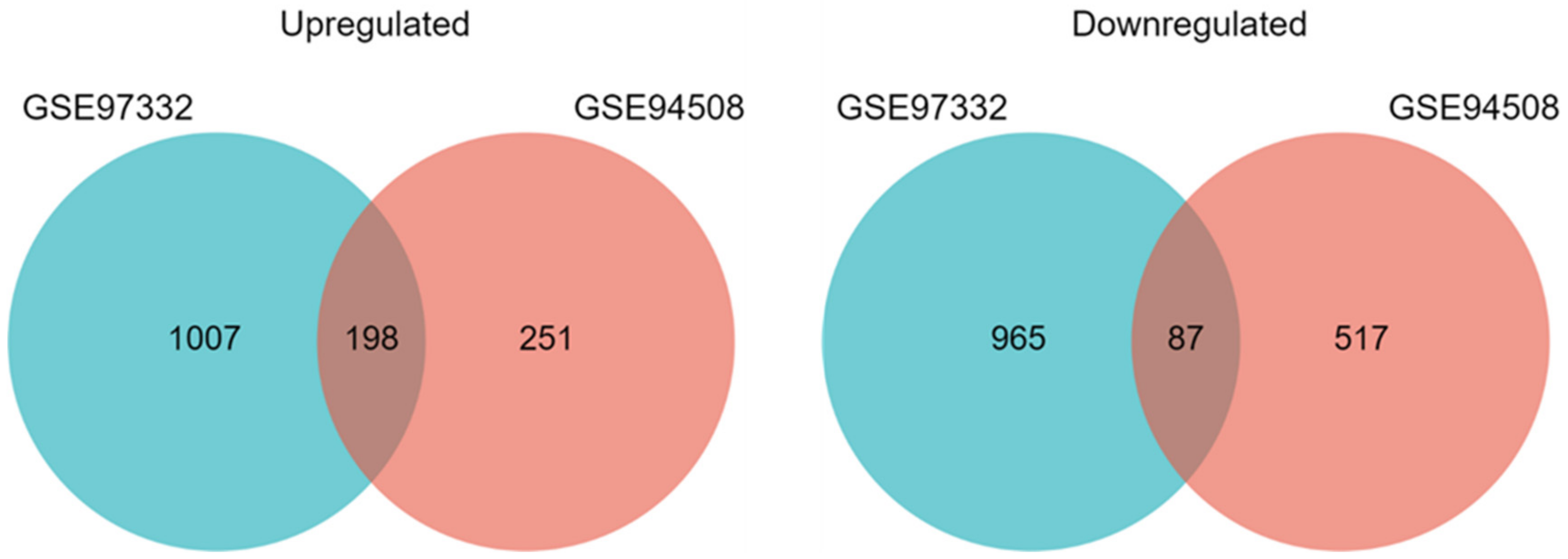
2.1. Differentially Expressed circRNAs and mRNAs in HCC
2.2. Identification of Enriched RBPs Motifs in the Groups of Differentially Expressed circRNAs
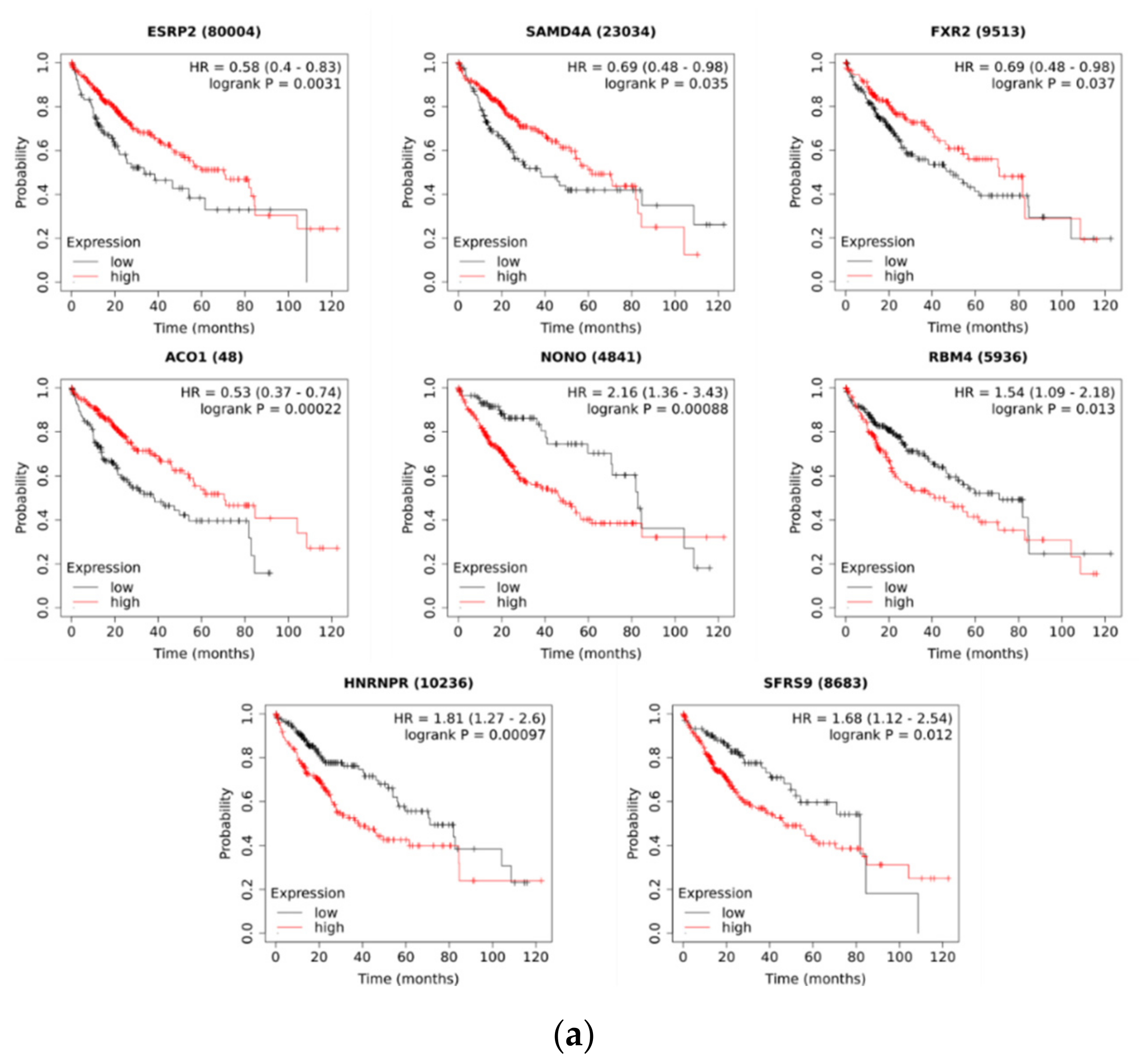
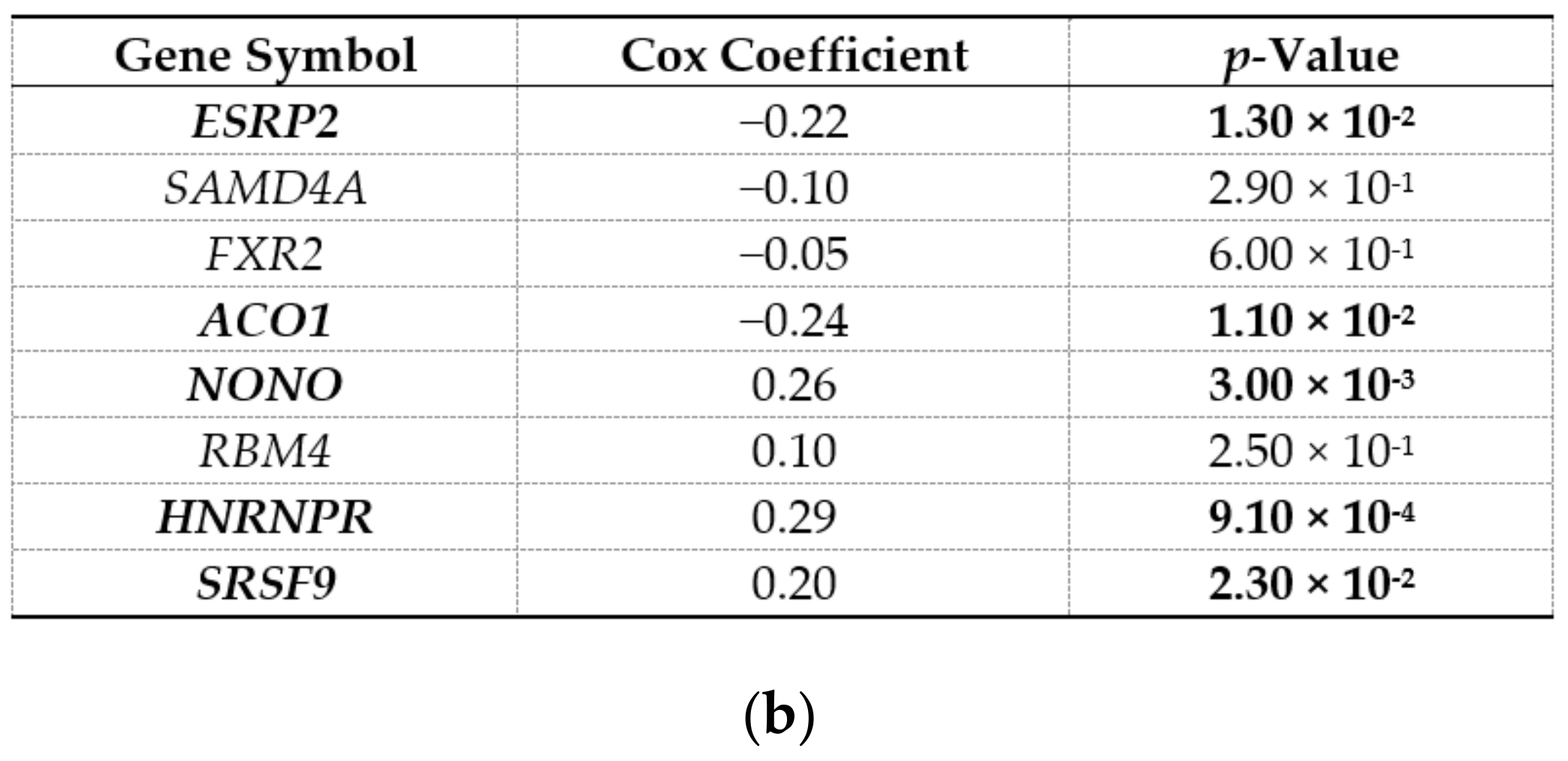
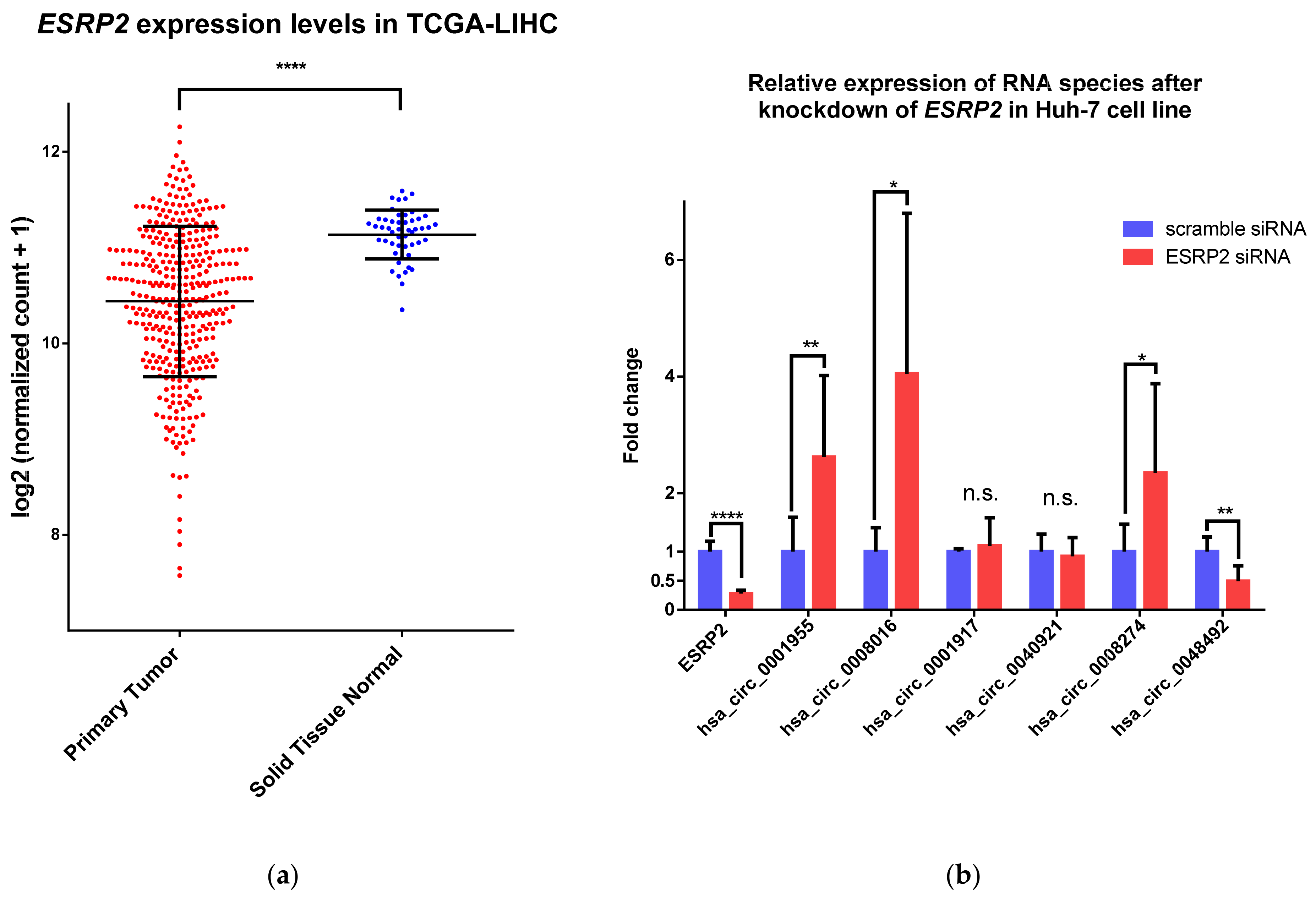
2.3. ESRP2 Is a Potential Regulator of circRNA Expression in HCC
3. Discussion
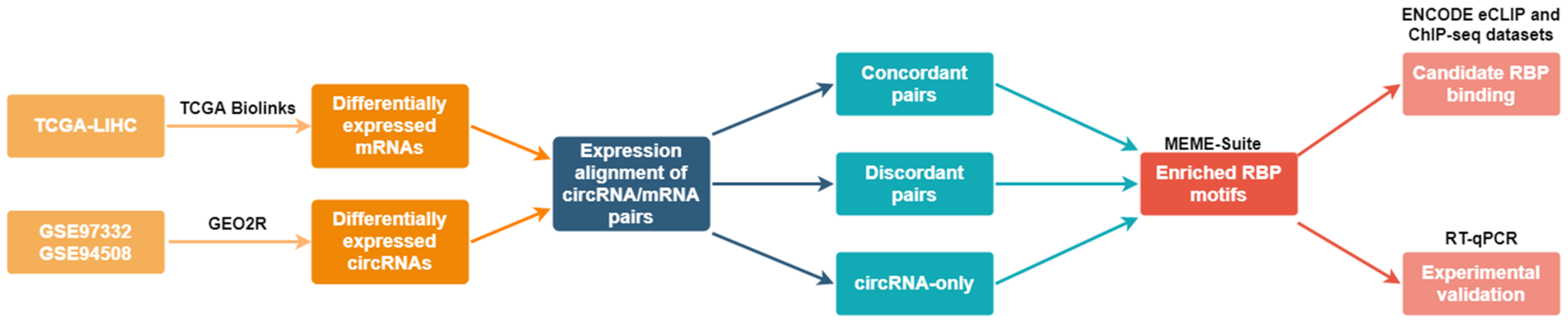
4. Materials and Methods
4.1. Identification of Differentially Expressed circRNAs and mRNAs in HCC
4.2. Expression Alignment of circRNAs and Host Gene mRNAs
4.3. Identification of Enriched Motifs of RBPs in Groups of Differentially Expressed circRNAs
4.4. Identification of Known RBP Binding Sites from Published eCLIP and ChIP-seq Data
4.5. Survival Analysis
4.6. Cell Culture Huh-7 and Knockdown of ESRP2 Expression
4.7. RNA Isolation, Reverse Transcription and RT-qPCR
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The Biogenesis, Biology and Characterization of Circular RNAs. Nat. Rev. Genet. 2019, 20, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Hansen, T.B.; Venø, M.T.; Kjems, J. Circular RNAs in Cancer: Opportunities and Challenges in the Field. Oncogene 2017, 37, 555–565. [Google Scholar] [CrossRef]
- Fu, L.; Jiang, Z.; Li, T.; Hu, Y.; Guo, J. Circular RNAs in Hepatocellular Carcinoma: Functions and Implications. Cancer Med. 2018, 7, 3101–3109. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, Q.; Bao, C.; Li, S.; Guo, W.; Zhao, J.; Chen, D.; Gu, J.; He, X.; Huang, S. Circular RNA Is Enriched and Stable in Exosomes: A Promising Biomarker for Cancer Diagnosis. Cell Res. 2015, 25, 981. [Google Scholar] [CrossRef]
- Dong, R.; Ma, X.K.; Chen, L.L.; Yang, L. Increased Complexity of CircRNA Expression during Species Evolution. RNA Biol. 2017, 14, 1064–1074. [Google Scholar] [CrossRef] [PubMed]
- Stagsted, L.V.W.; O’leary, E.T.; Ebbesen, K.K.; Hansen, T.B. The Rna-Binding Protein Sfpq Preserves Long-Intron Splicing and Regulates Circrna Biogenesis in Mammals. eLife 2021, 10, 1–26. [Google Scholar] [CrossRef]
- Salzman, J.; Chen, R.E.; Olsen, M.N.; Wang, P.L.; Brown, P.O. Cell-Type Specific Features of Circular RNA Expression. PLoS Genet. 2013, 9, e1003777. [Google Scholar] [CrossRef]
- Rybak-Wolf, A.; Stottmeister, C.; Glažar, P.; Jens, M.; Pino, N.; Hanan, M.; Behm, M.; Bartok, O.; Ashwal-Fluss, R.; Herzog, M.; et al. Circular RNAs in the Mammalian Brain Are Highly Abundant, Conserved, and Dynamically Expressed. Mol. Cell 2014, 58, 870–885. [Google Scholar] [CrossRef] [PubMed]
- Siede, D.; Rapti, K.; Gorska, A.A.; Katus, H.A.; Altmüller, J.; Boeckel, J.N.; Meder, B.; Maack, C.; Völkers, M.; Müller, O.J.; et al. Identification of Circular RNAs with Host Gene-Independent Expression in Human Model Systems for Cardiac Differentiation and Disease. J. Mol. Cell. Cardiol. 2017, 109, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, E.; Cairns, M.J. Circular RNAs Are Temporospatially Regulated throughout Development and Ageing in the Rat. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Tatomer, D.C.; Luo, Z.; Wu, H.; Yang, L.; Chen, L.L.; Cherry, S.; Wilusz, J.E. The Output of Protein-Coding Genes Shifts to Circular RNAs When the Pre-MRNA Processing Machinery Is Limiting. Mol. Cell 2017, 68, 940–954.e3. [Google Scholar] [CrossRef] [PubMed]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. CircRNA Biogenesis Competes with Pre-MRNA Splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Conn, S.J.; Pillman, K.A.; Toubia, J.; Conn, V.M.; Salmanidis, M.; Phillips, C.A.; Roslan, S.; Schreiber, A.W.; Gregory, P.A.; Goodall, G.J. The RNA Binding Protein Quaking Regulates Formation of CircRNAs. Cell 2015, 160, 1125–1134. [Google Scholar] [CrossRef]
- Ivanov, A.; Memczak, S.; Wyler, E.; Torti, F.; Porath, H.T.; Orejuela, M.R.; Piechotta, M.; Levanon, E.Y.; Landthaler, M.; Dieterich, C.; et al. Analysis of Intron Sequences Reveals Hallmarks of Circular RNA Biogenesis in Animals. Cell Rep. 2015, 10, 170–177. [Google Scholar] [CrossRef]
- Khan, M.A.F.; Reckman, Y.J.; Aufiero, S.; Van Den Hoogenhof, M.M.G.; Van Der Made, I.; Beqqali, A.; Koolbergen, D.R.; Rasmussen, T.B.; Van Der Velden, J.; Creemers, E.E.; et al. RBM20 Regulates Circular RNA Production from the Titin Gene. Circ. Res. 2016, 119, 996–1003. [Google Scholar] [CrossRef] [PubMed]
- Aktaş, T.; Avşar Ilık, İ.; Maticzka, D.; Bhardwaj, V.; Pessoa Rodrigues, C.; Mittler, G.; Manke, T.; Backofen, R.; Akhtar, A. DHX9 Suppresses RNA Processing Defects Originating from the Alu Invasion of the Human Genome. Nature 2017, 544, 115–119. [Google Scholar] [CrossRef]
- Li, X.; Liu, C.X.; Xue, W.; Zhang, Y.; Jiang, S.; Yin, Q.F.; Wei, J.; Yao, R.W.; Yang, L.; Chen, L.L. Coordinated CircRNA Biogenesis and Function with NF90/NF110 in Viral Infection. Mol. Cell 2017, 67, 214–227.e7. [Google Scholar] [CrossRef] [PubMed]
- Errichelli, L.; Dini Modigliani, S.; Laneve, P.; Colantoni, A.; Legnini, I.; Capauto, D.; Rosa, A.; De Santis, R.; Scarfò, R.; Peruzzi, G.; et al. FUS Affects Circular RNA Expression in Murine Embryonic Stem Cell-Derived Motor Neurons. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Dong, J.; Li, X.; Cheng, Z.; Zhu, Q. CPSF4 Regulates CircRNA Formation and MicroRNA Mediated Gene Silencing in Hepatocellular Carcinoma. Oncogene 2021. [Google Scholar] [CrossRef]
- Ho, J.S.Y.; Di Tullio, F.; Schwarz, M.; Low, D.; Incarnato, D.; Gay, F.; Tabaglio, T.; Zhang, J.; Wollmann, H.; Chen, L.; et al. HNRNPM Controls CircRNA Biogenesis and Splicing Fidelity to Sustain Cancer Cell Fitness. eLife 2021, 10, e59654. [Google Scholar] [CrossRef]
- Fitzmaurice, C.; Abate, D.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdel-Rahman, O.; Abdelalim, A.; Abdoli, A.; Abdollahpour, I.; Abdulle, A.S.M.; et al. Global, Regional, and National Cancer Incidence, Mortality, Years of Life Lost, Years Lived with Disability, and Disability-Adjusted Life-Years for 29 Cancer Groups, 1990 to 2017: A Systematic Analysis for the Global Burden of Disease Study. JAMA Oncol. 2019, 5, 1749–1768. [Google Scholar] [CrossRef]
- Lee, A.; Lee, F.-C. Medical Oncology Management of Advanced Hepatocellular Carcinoma 2019: A Reality Check. Front. Med. 2020, 14, 273–283. [Google Scholar] [CrossRef]
- Qiu, L.; Wang, T.; Ge, Q.; Xu, H.; Wu, Y.; Tang, Q.; Chen, K. Circular RNA Signature in Hepatocellular Carcinoma. J. Cancer 2019, 10, 3361–3372. [Google Scholar] [CrossRef] [PubMed]
- Webster, N.J.G. Alternative RNA Splicing in the Pathogenesis of Liver Disease. Front. Endocrinol. 2017, 8, 1–9. [Google Scholar] [CrossRef]
- Jimenez, M.; Arechederra, M.; Ávila, M.A.; Berasain, C. Splicing Alterations Contributing to Cancer Hallmarks in the Liver: Central Role of Dedifferentiation and Genome Instability. Transl. Gastroenterol. Hepatol. 2018, 3, 84. [Google Scholar] [CrossRef]
- Li, S.; Hu, Z.; Zhao, Y.; Huang, S.; He, X. Transcriptome-Wide Analysis Reveals the Landscape of Aberrant Alternative Splicing Events in Liver Cancer. Hepatology 2019, 69, 359–375. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Yao, T.; Chen, Q.; Mo, X.; Hu, Y.; Guo, J. Screening Differential Circular RNA Expression Profiles Reveals Hsa_circ_0004018 Is Associated with Hepatocellular Carcinoma. Oncotarget 2017, 8, 58405–58416. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Li, J.; Wang, H.; Su, X.; Hou, J.; Gu, Y.; Qian, C.; Lin, Y.; Liu, X.; Huang, M.; et al. Circular RNA CircMTO1 Acts as the Sponge of MicroRNA-9 to Suppress Hepatocellular Carcinoma Progression. Hepatology 2017, 66, 1151–1164. [Google Scholar] [CrossRef]
- Chang, K.; Creighton, C.J.; Davis, C.; Donehower, L.; Drummond, J.; Wheeler, D.; Ally, A.; Balasundaram, M.; Birol, I.; Butterfield, Y.S.N.; et al. The Cancer Genome Atlas Pan-Cancer Analysis Project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef]
- Bhate, A.; Parker, D.J.; Bebee, T.W.; Ahn, J.; Arif, W.; Rashan, E.H.; Chorghade, S.; Chau, A.; Lee, J.-H.; Anakk, S.; et al. ESRP2 Controls an Adult Splicing Programme in Hepatocytes to Support Postnatal Liver Maturation. Nat. Commun. 2015, 6, 8768. [Google Scholar] [CrossRef]
- Wu, W.; Ji, P.; Zhao, F. CircAtlas: An Integrated Resource of One Million Highly Accurate Circular RNAs from 1070 Vertebrate Transcriptomes. Genome Biol. 2020, 21, 101. [Google Scholar] [CrossRef]
- Wilusz, J.E. A 360° View of Circular RNAs: From Biogenesis to Functions. Wiley Interdiscip. Rev. RNA 2018, 9, 1–17. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z. Efficient Backsplicing Produces Translatable Circular MRNAs. RNA 2015, 21, 172–179. [Google Scholar] [CrossRef]
- Di Timoteo, G.; Dattilo, D.; Centrón-Broco, A.; Colantoni, A.; Guarnacci, M.; Rossi, F.; Incarnato, D.; Oliviero, S.; Fatica, A.; Morlando, M.; et al. Modulation of CircRNA Metabolism by M6A Modification. Cell Rep. 2020, 31. [Google Scholar] [CrossRef]
- Hong, X.; Song, R.; Song, H.; Zheng, T.; Wang, J.; Liang, Y.; Qi, S.; Lu, Z.; Song, X.; Jiang, H.; et al. PTEN Antagonises Tcl1/HnRNPK-Mediated G6PD Pre-MRNA Splicing Which Contributes to Hepatocarcinogenesis. Gut 2014, 63, 1635–1647. [Google Scholar] [CrossRef]
- Hu, Z.; Dong, L.; Li, S.; Li, Z.; Qiao, Y.; Li, Y.; Ding, J.; Chen, Z.; Wu, Y.; Wang, Z.; et al. Splicing Regulator P54(Nrb) /Non-POU Domain-Containing Octamer-Binding Protein Enhances Carcinogenesis Through Oncogenic Isoform Switch of MYC Box-Dependent Interacting Protein 1 in Hepatocellular Carcinoma. Hepatology 2020, 72, 548–568. [Google Scholar] [CrossRef]
- Zhang, T.; Huang, X.-H.; Dong, L.; Hu, D.; Ge, C.; Zhan, Y.-Q.; Xu, W.-X.; Yu, M.; Li, W.; Wang, X.; et al. PCBP-1 Regulates Alternative Splicing of the CD44 Gene and Inhibits Invasion in Human Hepatoma Cell Line HepG2 Cells. Mol. Cancer 2010, 9, 72. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.L.; Pan, Q.; Zhang, R.N.; Shen, F.; Yan, S.Y.; Sun, C.; Xu, Z.J.; Chen, Y.W.; Fan, J.G. Disease-Specific MiR-34a as Diagnostic Marker of Nonalcoholic Steatohepatitis in a Chinese Population. World J. Gastroenterol. 2016, 22, 9844–9852. [Google Scholar] [CrossRef]
- Zhang, X.; Hua, L.; Yan, D.; Zhao, F.; Liu, J.; Zhou, H.; Liu, J.; Wu, M.; Zhang, C.; Chen, Y.; et al. Overexpression of PCBP2 Contributes to Poor Prognosis and Enhanced Cell Growth in Human Hepatocellular Carcinoma. Oncol. Rep. 2016, 36, 3456–3464. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Liu, J.; Wang, C.; Su, Y. NONO Promotes Hepatocellular Carcinoma Progression by Enhancing Fatty Acids Biosynthesis through Interacting with ACLY MRNA. Cancer Cell Int. 2020, 20, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Bai, Y.; Yang, X.; Lin, J.; Yang, X.; Wang, D.; He, L.; Zheng, Y.; Zhao, H. Construction and Comprehensive Analysis of a CeRNA Network to Reveal Potential Prognostic Biomarkers for Hepatocellular Carcinoma. Cancer Cell Int. 2019, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Qian, X.; Peng, L.-X.; Jiang, Y.; Hawke, D.H.; Zheng, Y.; Xia, Y.; Lee, J.-H.; Cote, G.; Wang, H.; et al. A Splicing Switch from Ketohexokinase-C to Ketohexokinase-A Drives Hepatocellular Carcinoma Formation. Nat. Cell Biol. 2016, 18, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; He, S.; Ma, X.; Zhang, M.; Zhuang, J.; Wang, G.; Ye, Y.; Xia, W. RBMX Contributes to Hepatocellular Carcinoma Progression and Sorafenib Resistance by Specifically Binding and Stabilizing BLACAT1. Am. J. Cancer Res. 2020, 10, 3644–3665. [Google Scholar]
- Liang, R.; Lin, Y.; Ye, J.-Z.; Yan, X.-X.; Liu, Z.-H.; Li, Y.-Q.; Luo, X.-L.; Ye, H.-H. High Expression of RBM8A Predicts Poor Patient Prognosis and Promotes Tumor Progression in Hepatocellular Carcinoma. Oncol. Rep. 2017, 37, 2167–2176. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bangru, S.; Arif, W.; Seimetz, J.; Bhate, A.; Chen, J.; Rashan, E.H.; Carstens, R.P.; Anakk, S.; Kalsotra, A. Alternative Splicing Rewires Hippo Signaling Pathway in Hepatocytes to Promote Liver Regeneration. Nat. Struct. Mol. Biol. 2018, 25, 928–939. [Google Scholar] [CrossRef]
- Hyun, J.; Sun, Z.; Ahmadi, A.R.; Bangru, S.; Chembazhi, U.V.; Du, K.; Chen, T.; Tsukamoto, H.; Rusyn, I.; Kalsotra, A.; et al. Epithelial Splicing Regulatory Protein 2–Mediated Alternative Splicing Reprograms Hepatocytes in Severe Alcoholic Hepatitis. J. Clin. Invest. 2020, 130, 2129–2145. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, X.; Zhang, X.; Chen, R. Identification of Important Long Non-Coding RNAs and Highly Recurrent Aberrant Alternative Splicing Events in Hepatocellular Carcinoma through Integrative Analysis of Multiple RNA-Seq Datasets. Mol. Genet. Genom. 2016, 291, 1035–1051. [Google Scholar] [CrossRef]
- Kong, Q.; Fan, Q.; Ma, X.; Li, J.; Ma, R. CircRNA CircUGGT2 Contributes to Hepatocellular Carcinoma Development via Regulation of the MiR-526b-5p/RAB1A Axis. Cancer Manag. Res. 2020, 12, 10229–10241. [Google Scholar] [CrossRef]
- Ding, B.; Fan, W.; Lou, W. Hsa_circ_0001955 Enhances In Vitro Proliferation, Migration, and Invasion of HCC Cells through MiR-145-5p/NRAS Axis. Mol. Ther. Nucleic Acids 2020, 22, 445–455. [Google Scholar] [CrossRef]
- Zhang, M.; Xu, K.; Fu, L.; Wang, Q.; Chang, Z.; Zou, H.; Zhang, Y.; Li, Y. Revealing Epigenetic Factors of CircRNA Expression by Machine Learning in Various Cellular Contexts. iScience 2020, 23, 101842. [Google Scholar] [CrossRef]
- Xu, T.; Wang, L.; Jia, P.; Song, X.; Zhao, Z. An Integrative Transcriptomic and Methylation Approach for Identifying Differentially Expressed Circular RNAs Associated with DNA Methylation Change. Biomedicines 2021, 9, 657. [Google Scholar] [CrossRef] [PubMed]
- Llorian, M.; Schwartz, S.; Clark, T.A.; Hollander, D.; Tan, L.-Y.; Spellman, R.; Gordon, A.; Schweitzer, A.C.; de la Grange, P.; Ast, G.; et al. Position-Dependent Alternative Splicing Activity Revealed by Global Profiling of Alternative Splicing Events Regulated by PTB. Nat. Struct. Mol. Biol. 2010, 17, 1114–1123. [Google Scholar] [CrossRef] [PubMed]
- Warzecha, C.C.; Jiang, P.; Amirikian, K.; Dittmar, K.A.; Lu, H.; Shen, S.; Guo, W.; Xing, Y.; Carstens, R.P. An ESRP-Regulated Splicing Programme Is Abrogated during the Epithelial-Mesenchymal Transition. EMBO J. 2010, 29, 3286–3300. [Google Scholar] [CrossRef]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for Functional Genomics Data Sets—Update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef]
- Colaprico, A.; Silva, T.C.; Olsen, C.; Garofano, L.; Cava, C.; Garolini, D.; Sabedot, T.S.; Malta, T.M.; Pagnotta, S.M.; Castiglioni, I.; et al. TCGAbiolinks: An R/Bioconductor Package for Integrative Analysis of TCGA Data. Nucleic Acids Res. 2016, 44, e71. [Google Scholar] [CrossRef]
- Lan, P.-H.; Liu, Z.-H.; Pei, Y.-J.; Wu, Z.-G.; Yu, Y.; Yang, Y.-F.; Liu, X.; Che, L.; Ma, C.-J.; Xie, Y.-K.; et al. Landscape of RNAs in Human Lumbar Disc Degeneration. Oncotarget 2016, 7, 63166–63176. [Google Scholar] [CrossRef]
- Fu, C.; Lv, R.; Xu, G.; Zhang, L.; Bi, J.; Lin, L.; Liu, X.; Huo, R. Circular RNA Profile of Infantile Hemangioma by Microarray Analysis. PLoS ONE 2017, 12, e0187581. [Google Scholar] [CrossRef] [PubMed]
- Glažar, P.; Papavasileiou, P.; Rajewsky, N. CircBase: A Database for Circular RNAs. RNA 2014, 20, 1666–1670. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Ray, D.; Kazan, H.; Cook, K.B.; Weirauch, M.T.; Najafabadi, H.S.; Li, X.; Gueroussov, S.; Albu, M.; Zheng, H.; Yang, A.; et al. A Compendium of RNA-Binding Motifs for Decoding Gene Regulation. Nature 2013, 499, 172. [Google Scholar] [CrossRef]
- ENCODE Project Consortium. An Integrated Encyclopedia of DNA Elements in the Human Genome. Nature 2012, 489, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.A.; Hitz, B.C.; Sloan, C.A.; Chan, E.T.; Davidson, J.M.; Gabdank, I.; Hilton, J.A.; Jain, K.; Baymuradov, U.K.; Narayanan, A.K.; et al. The Encyclopedia of DNA Elements (ENCODE): Data Portal Update. Nucleic Acids Res. 2018, 46, D794–D801. [Google Scholar] [CrossRef]
- Xiao, R.; Chen, J.-Y.; Liang, Z.; Luo, D.; Chen, G.; Lu, Z.J.; Chen, Y.; Zhou, B.; Li, H.; Du, X.; et al. Pervasive Chromatin-RNA Binding Protein Interactions Enable RNA-Based Regulation of Transcription. Cell 2019, 178, 107–121.e18. [Google Scholar] [CrossRef]
- Thorvaldsdóttir, H.; Robinson, J.T.; Mesirov, J.P. Integrative Genomics Viewer (IGV): High-Performance Genomics Data Visualization and Exploration. Brief. Bioinform. 2013, 14, 178–192. [Google Scholar] [CrossRef] [PubMed]
- Hinrichs, A.S.; Karolchik, D.; Baertsch, R.; Barber, G.P.; Bejerano, G.; Clawson, H.; Diekhans, M.; Furey, T.S.; Harte, R.A.; Hsu, F.; et al. The UCSC Genome Browser Database: Update 2006. Nucleic Acids Res. 2006, 34, D590–D598. [Google Scholar] [CrossRef]
- Menyhárt, O.; Nagy, Á.; Győrffy, B. Determining Consistent Prognostic Biomarkers of Overall Survival and Vascular Invasion in Hepatocellular Carcinoma. R. Soc. Open Sci. 2018, 5, 181006. [Google Scholar] [CrossRef]
- Anaya, J. OncoLnc: Linking TCGA Survival Data to MRNAs, MiRNAs, and LncRNAs. PeerJ Comput. Sci. 2016, 2016. [Google Scholar] [CrossRef]
- Warzecha, C.C.; Sato, T.K.; Nabet, B.; Hogenesch, J.B.; Carstens, R.P. ESRP1 and ESRP2 Are Epithelial Cell-Type-Specific Regulators of FGFR2 Splicing. Mol. Cell 2009, 33, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wang, J.; Deng, F.; Yan, Z.; Xia, Y.; Wang, Z.; Ye, J.; Deng, Y.; Zhang, Z.; Qiao, M.; et al. TqPCR: A Touchdown QPCR Assay with Significantly Improved Detection Sensitivity and Amplification Efficiency of SYBR Green QPCR. PLoS ONE 2015, 10, e0132666. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| circRNA | mRNA | Expression Alignment | Number of Pairs |
|---|---|---|---|
| ↑ | ↑ | Concordant pairs | 88 |
| ↑ | ↓ | Discordant pairs | 33 |
| ↑ | x | circRNA-only | 66 |
| ↓ | ↓ | Concordant pairs | 11 |
| ↓ | ↑ | Discordant pairs | 36 |
| ↓ | x | circRNA-only | 39 |
| All | Concordant Pairs | Discordant Pairs | circRNA-Only | ||||
|---|---|---|---|---|---|---|---|
| Gene Sym. | Adj. p-Value | Gene Sym. | Adj. p-Value | Gene Sym. | Adj. p-Value | Gene Sym. | Adj. p-Value |
| Upregulated circRNAs, upstream of splice site | |||||||
| ESRP2 | 1.32 × 10−4 | ESRP2 | 5.78 × 10−3 | RBM8A | 6.82 × 10−3 | NONO | 9.28 × 10−6 |
| NONO | 2.67 × 10−4 | PPRC1 | 4.54 × 10−2 | PPRC1 | 3.12 × 10−2 | HNRNPK | 2.89 × 10−4 |
| PCBP2 | 3.61 × 10−4 | RBM6 | 4.69 × 10−2 | RBM4B | 3.76 × 10−2 | SRSF4 * | 1.19 × 10−3 |
| PPRC1 | 9.13 × 10−4 | RBM4 | 4.52 × 10−2 | PCBP1 | 2.42 × 10−3 | ||
| YTHDC1 | 1.03 × 10−2 | SAMD4A | 4.96 × 10−2 | RBMX | 1.39 × 10−2 | ||
| RBM4 | 1.22 × 10−2 | HNRNPH2 | 1.54 × 10−2 | ||||
| RBMX | 1.48 × 10−2 | FXR2 | 1.87 × 10−2 | ||||
| ACO1 | 1.64 × 10−2 | ESRP2 | 2.39 × 10−2 | ||||
| RBM5 | 1.81 × 10−2 | PCBP2 | 2.49 × 10−2 | ||||
| RBM4B | 2.45 × 10−2 | SRSF9 | 3.67 × 10−2 | ||||
| HNRNPH2 | 2.84 × 10−2 | ||||||
| SRSF4 * | 3.69 × 10−2 | ||||||
| Upregulated circRNAs, downstream of splice site | |||||||
| RBMX | 9.51 × 10−3 | x | x | RBM4B | 2.90 × 10−2 | x | x |
| HNRNPK | 2.41 × 10−2 | ||||||
| SAMD4A | 3.84 × 10−2 | ||||||
| Downregulated circRNAs, upstream of splice site | |||||||
| x | x | EIF4B | 6.26 × 10−3 | x | x | x | x |
| Downregulated circRNAs, downstream of splice site | |||||||
| HNRNPR * | 9.20 × 10−3 | x | x | x | x | PABPC5 | 1.89 × 10−2 |
| Gene Symbol | circRNA | Experiment | Experiment ID |
|---|---|---|---|
| MSFD12 | hsa_circ_0048492 | NONO eCLIP K562 | ENCSR861PAR |
| ANKRD11 | hsa_circ_0040921 | NONO eCLIP K562 | ENCSR861PAR |
| FGFR1 | hsa_circ_0008016 | PCBP2 eCLIP HepG2 | ENCSR339FUY |
| ZMIZ1 | hsa_circ_0092313 | PCBP2 eCLIP HepG2 | ENCSR339FUY |
| DCAF8 | hsa_circ_0014879 | HNRNPK eCLIP HepG2 | ENCSR828ZID |
| LIN52 | hsa_circ_0000554 | PCBP1 ChIP-seq HepG2 | ENCSR872EVQ |
| DENND3 | hsa_circ_0001827 | PCBP1 ChIP-seq HepG2 | ENCSR872EVQ |
| DENND3 | hsa_circ_0001828 | PCBP1 ChIP-seq HepG2 | ENCSR872EVQ |
| DCAF8 | hsa_circ_0014879 | PCBP1 ChIP-seq HepG2 | ENCSR872EVQ |
| LIN52 | hsa_circ_0000554 | NONO ChIP-seq HepG2 | ENCSR923UTX |
| DENND3 | hsa_circ_0001827 | NONO ChIP-seq HepG2 | ENCSR923UTX |
| DENND3 | hsa_circ_0001828 | NONO ChIP-seq HepG2 | ENCSR923UTX |
| DENND3 | hsa_circ_0001827 | HNRNPK ChIP-seq HepG2 | ENCSR519QAA |
| DENND3 | hsa_circ_0001828 | HNRNPK ChIP-seq HepG2 | ENCSR519QAA |
| Gene Symbol | Fold-Change | FDR |
|---|---|---|
| SAMD4A | 0.62 | 2.26 × 10−8 |
| ESRP2 | 0.65 | 1.53 × 10−8 |
| ACO1 | 0.77 | 1.85 × 10−3 |
| FXR2 | 0.82 | 4.89 × 10−4 |
| PCBP1 | 1.20 | 2.24 × 10−4 |
| PCBP2 | 1.20 | 1.25 × 10−3 |
| EIF4B | 1.22 | 4.01 × 10−3 |
| RBM4B | 1.27 | 3.40 × 10−4 |
| SFRS9 | 1.29 | 9.36 × 10−6 |
| RBM5 | 1.33 | 4.59 × 10−8 |
| RBM4 | 1.39 | 8.15 × 10−7 |
| HNRNPK | 1.40 | 1.36 × 10−12 |
| RBM6 | 1.45 | 8.67 × 10−10 |
| HNRNPR | 1.47 | 6.28 × 10−14 |
| RBM8A | 1.64 | 3.87 × 10−15 |
| RBMX | 1.71 | 3.40 × 10−20 |
| NONO | 1.76 | 2.83 × 10−14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Razpotnik, R.; Nassib, P.; Kunej, T.; Rozman, D.; Režen, T. Identification of Novel RNA Binding Proteins Influencing Circular RNA Expression in Hepatocellular Carcinoma. Int. J. Mol. Sci. 2021, 22, 7477. https://doi.org/10.3390/ijms22147477
Razpotnik R, Nassib P, Kunej T, Rozman D, Režen T. Identification of Novel RNA Binding Proteins Influencing Circular RNA Expression in Hepatocellular Carcinoma. International Journal of Molecular Sciences. 2021; 22(14):7477. https://doi.org/10.3390/ijms22147477
Chicago/Turabian StyleRazpotnik, Rok, Petra Nassib, Tanja Kunej, Damjana Rozman, and Tadeja Režen. 2021. "Identification of Novel RNA Binding Proteins Influencing Circular RNA Expression in Hepatocellular Carcinoma" International Journal of Molecular Sciences 22, no. 14: 7477. https://doi.org/10.3390/ijms22147477
APA StyleRazpotnik, R., Nassib, P., Kunej, T., Rozman, D., & Režen, T. (2021). Identification of Novel RNA Binding Proteins Influencing Circular RNA Expression in Hepatocellular Carcinoma. International Journal of Molecular Sciences, 22(14), 7477. https://doi.org/10.3390/ijms22147477