
The Enigma of Eosinophil Degranulation

 , , and
, , and
Abstract
1. Introduction
2. Molecular Regulation of Eosinophil Degranulation
3. The Close Relationship between Degranulation and dsDNA Release in EET Formation
4. Agonists and Methods Used in the Assessment of Eosinophil Degranulation

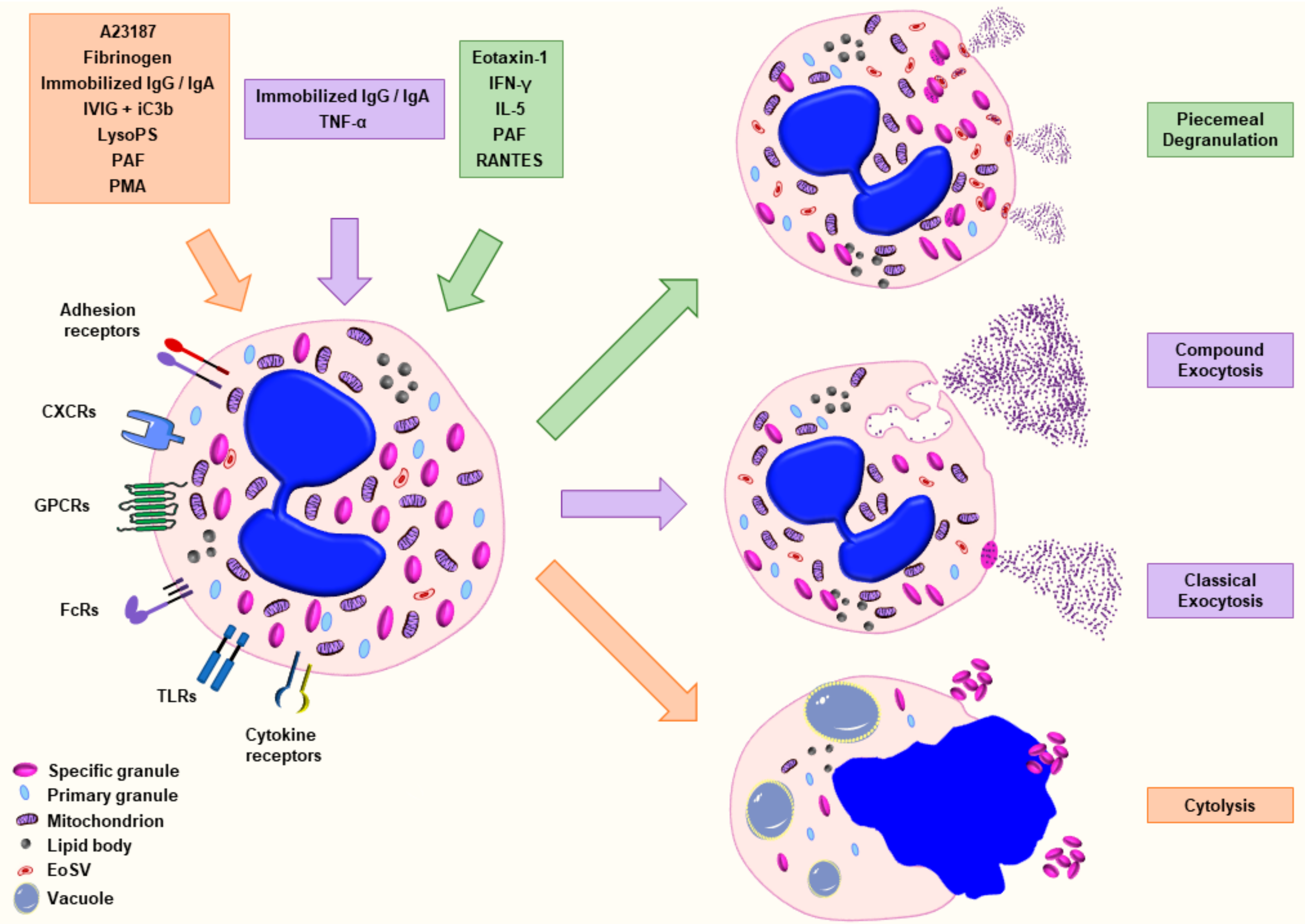{kind=link}
{kind=link}
| Stimulation/Agonists | Degranulation Type | References |
|---|---|---|
| A23187 | Cytolysis | [55,60,100] |
| C5a (+ IFN-γ * or IL-5 * or GM-CSF *,+) | --- | [4] * [44] + [94] + [98] |
| CCL11 (Eotaxin-1) (+ IFN-γ * or IL-5 *,+) | Piecemeal degranulation (PMD) | [4] * [20,39,76] [77] + [78,79,80] |
| Fibrinogen | Cytolysis | [101] |
| IFN-γ | Piecemeal degranulation (PMD) | [34,93,97,98] |
| IL-33 | --- | [37,66,67,68,104] |
| IL-5 | Piecemeal degranulation (PMD) | [66,67,68,71,72] |
| Immobilized IgG or IgA | Cytolysis | [55,102] |
| Exocytosis | [102] | |
| Ionomycin | --- | [4,33,37] |
| IVIG + iC3b | Cytolysis | [23] |
| lysoPS | Cytolysis | [60] |
| fMLP | --- | [67,96] |
| PAF (+ IL-5 * or IFN-γ * or IL-2 ¥ or GM-CSF £) | Cytolysis | [60] |
| Piecemeal degranulation (PMD) | [4] * [14] ¥ [20,33,37,92] [94] £ | |
| --- | [93] | |
| PMA | Cytolysis | [55] |
| --- | [4,41,96] | |
| RANTES | Piecemeal degranulation (PMD) | [20] |
| TNF-α | Compound exocytosis | [20,39,78,79,82] |
| Measured/Targeted Protein | Method | References |
|---|---|---|
| Eosinophil Peroxidase (EPX) | Colorimetric assay | [4,41,92,98] |
| Confocal laser scanning microscopy (CSLM) /Immunofluorescence staining | [4,68] | |
| ELISA | [33,37,38,93,97,105] | |
| Immunohistochemistry /Ultrastructural cytochemistry | [38,72] | |
| Mass spectrometry analysis | [101] | |
| Eosinophil Cationic Protein (ECP) | Confocal laser scanning microscopy (CSLM) /Immunofluorescence staining | [68] |
| Immunohistochemistry | [94] | |
| Pharmacia UniCap | [94] | |
| Radioimmunoassay | [76,82] | |
| Eosinophil-derived Neurotoxin (EDN) | Confocal laser scanning microscopy (CSLM) /Immunofluorescence staining | [93] |
| ELISA | [60,66,67,93,104] | |
| Radioimmunoassay | [80,100,102] | |
| Major Basic Protein (MBP-1) | Confocal laser scanning microscopy (CSLM) /Immunofluorescence staining | [97,101] |
| Dot blot assay | [37] | |
| Immunohistochemistry | [38] | |
| Immunonanogold electron microscopy (EM) | [106] | |
| Mass spectrometry analysis | [101] | |
| Cell Surface Upregulation of Surrogate Degranulation Marker CD63 | Flow cytometry (FACS) | [4,44,68,98,105] |
| CD63 Immunolabeling | Confocal laser scanning microscopy (CSLM) /Immunofluorescence staining | [33,37,98] |
| Immunonanogold electron microscopy (EM) | [20,78,93] | |
| Granzyme-B | Confocal laser scanning microscopy (CSLM) /Immunofluorescence staining | [68] |
| Charcot-Leyden Protein (CLC) | Confocal laser scanning microscopy (CSLM) /Immunofluorescence staining | [93,107] |
| Immunonanogold electron microscopy (EM) | [71,72,107] | |
| Radioimmunoassay | [100] | |
| IFN-γ | Immunonanogold electron microscopy (EM) | [79] |
| IL-4 | ELISA | [77] |
| Immunofluorescence staining | [77] | |
| Immunonanogold electron microscopy (EM) | [108] | |
| Granule Density/ Ultrastructural analysis | Transmission electron microscopy (TEM) | [20,22,55,68,71,72,78,79,96,101,102,106] |
| Phase-contrast microscopy | [55] | |
| Qa-SNARE syntaxin-17 (STX17) | Immunonanogold electron microscopy (EM) | [39] |
| Transmission electron microscopy (TEM) | [39] | |
| RANTES (CCL5) | Confocal laser scanning microscopy (CSLM) /Immunofluorescence staining | [34,97,98] |
| ELISA | [98] | |
| Immunonanogold electron microscopy (EM) | [97] |
5. The Implication of Charcot–Leyden Crystal Protein in Degranulation
6. Function and Clinical Relevance of Eosinophil-Derived Granule Proteins, Cytokines and Chemokines
7. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Marichal, T.; Mesnil, C.; Bureau, F. Homeostatic Eosinophils: Characteristics and Functions. Front. Med. 2017, 4, 101. [Google Scholar] [CrossRef]
- Simon, H.-U.; Yousefi, S.; Germic, N.; Arnold, I.C.; Haczku, A.; Karaulov, A.V.; Simon, D.; Rosenberg, H.F. The Cellular Functions of Eosinophils: Collegium Internationale Allergologicum (CIA) Update 2020. Int. Arch. Allergy Immunol. 2019, 181, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, H.F.; Druey, K.M. Modeling asthma: Pitfalls, promises, and the road ahead. J. Leukoc. Biol. 2018, 104, 41–48. [Google Scholar] [CrossRef]
- Germic, N.; Fettrelet, T.; Stojkov, D.; Hosseini, A.; Horn, M.; Karaulov, A.; Simon, D.; Yousefi, S.; Simon, H.-U. The Release Kinetics of Eosinophil Peroxidase and Mitochondrial DNA Is Different in Association with Eosinophil Extracellular Trap Formation. Cells 2021, 10, 306. [Google Scholar] [CrossRef]
- Ramirez, G.A.; Yacoub, M.-R.; Ripa, M.; Mannina, D.; Cariddi, A.; Saporiti, N.; Ciceri, F.; Castagna, A.; Colombo, G.; Dagna, L. Eosinophils from Physiology to Disease: A Comprehensive Review. BioMed Res. Int. 2018, 2018, 1–28. [Google Scholar] [CrossRef]
- Bainton, D.F.; Farquhar, M.G. Segregation and Packaging of Granule Enzymes in Eosinophilic Leukocytes. J. Cell Biol. 1970, 45, 54–73. [Google Scholar] [CrossRef]
- Melo, R.C.N.; Weller, P.F. Contemporary understanding of the secretory granules in human eosinophils. J. Leukoc. Biol. 2018, 104, 85–93. [Google Scholar] [CrossRef]
- Giembycz, M.A.; Lindsay, M.A. Pharmacology of the Eosinophil. Pharmacol. Rev. 1999, 51, 213–340. [Google Scholar] [PubMed]
- Davoine, F.; Lacy, P. Eosinophil Cytokines, Chemokines, and Growth Factors: Emerging Roles in Immunity. Front. Immunol. 2014, 5, 570. [Google Scholar] [CrossRef]
- Gigon, L.; Yousefi, S.; Karaulov, A.; Simon, H.-U. Mechanisms of toxicity mediated by neutrophil and eosinophil granule proteins. Allergol. Int. 2021, 70, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Bandeira-Melo, C.; Bozza, P.; Weller, P.F. The cellular biology of eosinophil eicosanoid formation and function. J. Allergy Clin. Immunol. 2002, 109, 393–400. [Google Scholar] [CrossRef]
- Kita, H. Eosinophils: Multifunctional and Distinctive Properties. Int. Arch. Allergy Immunol. 2013, 161, 3–9. [Google Scholar] [CrossRef]
- Matthews, A.N.; Friend, D.S.; Zimmermann, N.; Sarafi, M.N.; Luster, A.D.; Pearlman, E.; Wert, S.E.; Rothenberg, M.E. Eotaxin is required for the baseline level of tissue eosinophils. Proc. Natl. Acad. Sci. USA 1998, 95, 6273–6278. [Google Scholar] [CrossRef]
- Simon, H.-U.; Plötz, S.; Simon, D.; Seitzer, U.; Braathen, L.R.; Menz, G.; Straumann, A.; Dummer, R.; Levi-Schaffer, F. Interleukin-2 primes eosinophil degranulation in hypereosinophilia and Wells’ syndrome. Eur. J. Immunol. 2003, 33, 834–839. [Google Scholar] [CrossRef]
- Spencer, L.A.; Bonjour, K.; Melo, R.C.N.; Weller, P.F. Eosinophil Secretion of Granule-Derived Cytokines. Front. Immunol. 2014, 5, 496. [Google Scholar] [CrossRef]
- Dvorak, H.F.; Dvorak, A.M. Basophilic Leucocytes: Structure, Function and Role in Disease. Clin. Haematol. 1975, 4, 651–683. [Google Scholar] [CrossRef]
- Dvorak, A.M.; Monahan, R.A.; Osage, J.E.; Dickersin, G.R. Crohn’s disease: Transmission electron microscopic studies. II. Immunologic Inflammatory Response. Alterations of Mast Cells, Basophils, Eosinophils, and the Microvasculature. Hum. Pathol. 1980, 11, 606–619. [Google Scholar] [CrossRef]
- Dvorak, A.M. Basophils and mast cells: Piecemeal degranulation in situ and ex vivo: A possible mechanism for cytokine-induced function in disease. Immunol. Ser. 1992, 57, 169–271. [Google Scholar]
- Karawajczyk, M.; Seveus, L.; Garcia, R.; Björnsson, E.; Peterson, C.G.B.; Roomans, G.M.; Venge, P. Piecemeal Degranulation of Peripheral Blood Eosinophils: A Study of Allergic Subjects during and out of the Pollen Season. Am. J. Respir. Cell Mol. Biol. 2000, 23, 521–529. [Google Scholar] [CrossRef]
- Melo, R.C.; Perez, S.A.; Spencer, L.A.; Dvorak, A.M.; Weller, P.F. Intragranular Vesiculotubular Compartments are Involved in Piecemeal Degranulation by Activated Human Eosinophils. Traffic 2005, 6, 866–879. [Google Scholar] [CrossRef]
- Persson, C.G.; Erjefält, J.S. Eosinophil lysis and free granules: An in vivo paradigm for cell activation and drug development. Trends Pharmacol. Sci. 1997, 18, 117–123. [Google Scholar] [CrossRef]
- Erjefält, J.S.; Andersson, M.; Greiff, L.; Korsgren, M.; Gizycki, M.; Jeffery, P.K.; Persson, C.G. Cytolysis and piecemeal degranulation as distinct modes of activation of airway mucosal eosinophils. J. Allergy Clin. Immunol. 1998, 102, 286–294. [Google Scholar] [CrossRef]
- Radonjic-Hoesli, S.; Wang, X.; de Graauw, E.; Stoeckle, C.; Styp-Rekowska, B.; Hlushchuk, R.; Simon, D.; Spaeth, P.J.; Yousefi, S.; Simon, H.-U. Adhesion-induced eosinophil cytolysis requires the receptor-interacting protein kinase 3 (RIPK3)–mixed lineage kinase-like (MLKL) signaling pathway, which is counterregulated by autophagy. J. Allergy Clin. Immunol. 2017, 140, 1632–1642. [Google Scholar] [CrossRef] [PubMed]
- Logan, M.R.; Odemuyiwa, S.O.; Moqbel, R. Understanding exocytosis in immune and inflammatory cells: The molecular basis of mediator secretion. J. Allergy Clin. Immunol. 2003, 111, 923–932. [Google Scholar] [CrossRef]
- De Toledo, G.A.; Fernandez, J.M. Compound versus multigranular exocytosis in peritoneal mast cells. J. Gen. Physiol. 1990, 95, 397–409. [Google Scholar] [CrossRef]
- Hide, I.; Bennett, J.P.; Pizzey, A.; Boonen, G.; Bar-Sagi, D.; Gomperts, B.D.; Tatham, P.E. Degranulation of individual mast cells in response to Ca2+ and guanine nucleotides: An all-or-none event. J. Cell Biol. 1993, 123, 585–593. [Google Scholar] [CrossRef]
- Scepek, S.; Moqbel, R.; Lindau, M. Compound exocytosis and cumulative degranulation by eosinophils and their role in parasite killing. Parasitol. Today 1994, 10, 276–278. [Google Scholar] [CrossRef]
- Lollike, K.; Lindau, M.; Calafat, J.; Borregaard, N. Compound exocytosis of granules in human neutrophils. J. Leukoc. Biol. 2002, 71, 973–980. [Google Scholar] [CrossRef]
- Morgenstern, E. The formation of compound granules from different types of secretory organelles in human platelets (dense granules and alpha-granules). A cryofixation/-substitution study using serial sections. Eur. J. Cell Biol. 1995, 68, 183–190. [Google Scholar]
- Atkinson, E.A.; Gerrard, J.M.; Hildes, G.E.; Greenberg, A.H. Studies of the Mechanism of Natural Killer (NK) Degranulation and Cytotoxicity. J. Leukoc. Biol. 1990, 47, 39–48. [Google Scholar] [CrossRef]
- Acharya, K.R.; Ackerman, S.J. Eosinophil Granule Proteins: Form and Function. J. Biol. Chem. 2014, 289, 17406–17415. [Google Scholar] [CrossRef]
- Coughlin, J.J.; Odemuyiwa, S.O.; Davidson, C.E.; Moqbel, R. Differential expression and activation of Rab27A in human eosinophils: Relationship to blood eosinophilia. Biochem. Biophys. Res. Commun. 2008, 373, 382–386. [Google Scholar] [CrossRef]
- Kim, J.D.; Willetts, L.; Ochkur, S.; Srivastava, N.; Hamburg, R.; Shayeganpour, A.; Seabra, M.C.; Lee, J.J.; Moqbel, R.; Lacy, P. An essential role for Rab27a GTPase in eosinophil exocytosis. J. Leukoc. Biol. 2013, 94, 1265–1274. [Google Scholar] [CrossRef]
- Lacy, P.; Logan, M.R.; Bablitz, B.; Moqbel, R. Fusion protein vesicle-associated membrane protein 2 is implicated in IFN-γ–induced piecemeal degranulation in human eosinophils from atopic individuals. J. Allergy Clin. Immunol. 2001, 107, 671–678. [Google Scholar] [CrossRef]
- Logan, M.R.; Lacy, P.; Bablitz, B.; Moqbel, R. Expression of eosinophil target SNAREs as potential cognate receptors for vesicle-associated membrane protein-2 in exocytosis. J. Allergy Clin. Immunol. 2002, 109, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Logan, M.R.; Lacy, P.; Odemuyiwa, S.O.; Steward, M.; Davoine, F.; Kita, H.; Moqbel, R. A critical role for vesicle-associated membrane protein-7 in exocytosis from human eosinophils and neutrophils. Allergy 2006, 61, 777–784. [Google Scholar] [CrossRef]
- Willetts, L.; Felix, L.C.; Jacobsen, E.A.; Puttagunta, L.; Condjella, R.M.; Zellner, K.R.; Ochkur, S.I.; Kim, J.D.; Luo, H.; Lee, N.A.; et al. Vesicle-associated membrane protein 7-mediated eosinophil degranulation promotes allergic airway inflammation in mice. Commun. Biol. 2018, 1, 83. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, E.A.; Ochkur, S.I.; Doyle, A.D.; LeSuer, W.E.; Li, W.; Protheroe, C.A.; Colbert, D.; Zellner, K.R.; Shen, H.H.; Irvin, C.G.; et al. Lung Pathologies in a Chronic Inflammation Mouse Model Are Independent of Eosinophil Degranulation. Am. J. Respir. Crit. Care Med. 2017, 195, 1321–1332. [Google Scholar] [CrossRef]
- Carmo, L.A.S.; Dias, F.F.; Malta, K.K.; Amaral, K.B.; Shamri, R.; Weller, P.F.; Melo, R.C.N. Expression and subcellular localization of the Qa-SNARE syntaxin17 in human eosinophils. Exp. Cell Res. 2015, 337, 129–135. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Dhariwala, F.A.; Rajadhyaksha, M.S. An Unusual Member of the Cdk Family: Cdk5. Cell. Mol. Neurobiol. 2008, 28, 351–369. [Google Scholar] [CrossRef]
- Odemuyiwa, S.O.; Ilarraza, R.; Davoine, F.; Logan, M.R.; Shayeganpour, A.; Wu, Y.; Majaesic, C.; Adamko, D.J.; Moqbel, R.; Lacy, P. Cyclin-dependent kinase 5 regulates degranulation in human eosinophils. Immunology 2015, 144, 641–648. [Google Scholar] [CrossRef]
- Germic, N.; Frangez, Z.; Yousefi, S.; Simon, H.-U. Regulation of the innate immune system by autophagy: Neutrophils, eosinophils, mast cells, NK cells. Cell Death Differ. 2019, 26, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Germic, N.; Hosseini, A.; Yousefi, S.; Karaulov, A.; Simon, H.-U. Regulation of eosinophil functions by autophagy. Semin. Immunopathol. 2021, 1–16. [Google Scholar] [CrossRef]
- Germic, N.; Hosseini, A.; Stojkov, D.; Oberson, K.; Claus, M.; Benarafa, C.; Calzavarini, S.; Angelillo-Scherrer, A.; Arnold, I.C.; Müller, A.; et al. ATG5 promotes eosinopoiesis but inhibits eosinophil effector functions. Blood 2021, 137, 2958–2969. [Google Scholar] [CrossRef]
- Yousefi, S.; Gold, J.A.; Andina, N.; Lee, J.J.; Kelly, A.M.; Kozlowski, E.; Schmid, I.; Straumann, A.; Reichenbach, J.; Gleich, G.J.; et al. Catapult-like release of mitochondrial DNA by eosinophils contributes to antibacterial defense. Nat. Med. 2008, 14, 949–953. [Google Scholar] [CrossRef]
- Dworski, R.; Simon, H.-U.; Hoskins, A.; Yousefi, S. Eosinophil and neutrophil extracellular DNA traps in human allergic asthmatic airways. J. Allergy Clin. Immunol. 2011, 127, 1260–1266. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Le Pham, D.; Lee, D.-H.; Lee, S.-H.; Kim, S.-H.; Park, H.-S. Biological function of eosinophil extracellular traps in patients with severe eosinophilic asthma. Exp. Mol. Med. 2018, 50, 1–8. [Google Scholar] [CrossRef]
- Simon, D.; Hoesli, S.; Roth, N.; Staedler, S.; Yousefi, S.; Simon, H.-U. Eosinophil extracellular DNA traps in skin diseases. J. Allergy Clin. Immunol. 2011, 127, 194–199. [Google Scholar] [CrossRef]
- Stojkov, D.; Amini, P.; Oberson, K.; Sokollik, C.; Duppenthaler, A.; Simon, H.-U.; Yousefi, S. ROS and glutathionylation balance cytoskeletal dynamics in neutrophil extracellular trap formation. J. Cell Biol. 2017, 216, 4073–4090. [Google Scholar] [CrossRef]
- Amini, P.; Stojkov, D.; Felser, A.; Jackson, C.B.; Courage, C.; Schaller, A.; Gelman, L.; Soriano, M.E.; Nuoffer, J.-M.; Scorrano, L.; et al. Neutrophil extracellular trap formation requires OPA1-dependent glycolytic ATP production. Nat. Commun. 2018, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, M.; Lacy, P.; Ueki, S. Eosinophil Extracellular Traps and Inflammatory Pathologies—Untangling the Web! Front. Immunol. 2018, 9, 2763. [Google Scholar] [CrossRef]
- Yousefi, S.; Stojkov, D.; Germic, N.; Simon, D.; Wang, X.; Benarafa, C.; Simon, H. Untangling “NETosis” from NETs. Eur. J. Immunol. 2019, 49, 221–227. [Google Scholar] [CrossRef]
- Morshed, M.; Yousefi, S.; Stöckle, C.; Simon, H.-U.; Simon, D. Thymic stromal lymphopoietin stimulates the formation of eosinophil extracellular traps. Allergy 2012, 67, 1127–1137. [Google Scholar] [CrossRef]
- Germic, N.; Stojkov, D.; Oberson, K.; Yousefi, S.; Simon, H. Neither eosinophils nor neutrophils require ATG5-dependent autophagy for extracellular DNA trap formation. Immunology 2017, 152, 517–525. [Google Scholar] [CrossRef]
- Ueki, S.; Melo, R.C.N.; Ghiran, I.; Spencer, L.A.; Dvorak, A.M.; Weller, P.F. Eosinophil extracellular DNA trap cell death mediates lytic release of free secretion-competent eosinophil granules in humans. Blood 2013, 121, 2074–2083. [Google Scholar] [CrossRef]
- Kano, G.; Bochner, B.S.; Zimmermann, N. Regulation of Siglec-8-induced intracellular reactive oxygen species production and eosinophil cell death by Src family kinases. Immunobiology 2017, 222, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, M.; Bulir, D.C.; Radford, K.; Kjarsgaard, M.; Huang, C.M.; Jacobsen, E.A.; Ochkur, S.I.; Catuneanu, A.; Lamothe-Kipnes, H.; Mahony, J.; et al. Sputum autoantibodies in patients with severe eosinophilic asthma. J. Allergy Clin. Immunol. 2018, 141, 1269–1279. [Google Scholar] [CrossRef]
- Muniz, V.S.; Silva, J.C.; Braga, Y.A.; Melo, R.C.; Ueki, S.; Takeda, M.; Hebisawa, A.; Asano, K.; Figueiredo, R.T.; Neves, J.S. Eosinophils release extracellular DNA traps in response to Aspergillus fumigatus. J. Allergy Clin. Immunol. 2018, 141, 571–585.e7. [Google Scholar] [CrossRef]
- Mukherjee, M.; Thomas, S.R.; Radford, K.; Dvorkin-Gheva, A.; Davydchenko, S.; Kjarsgaard, M.; Svenningsen, S.; Almas, S.; Felix, L.C.; Stearns, J.; et al. Sputum Antineutrophil Cytoplasmic Antibodies in Serum Antineutrophil Cytoplasmic Antibody–Negative Eosinophilic Granulomatosis with Polyangiitis. Am. J. Respir. Crit. Care Med. 2019, 199, 158–170. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Sim, M.S.; Lee, D.H.; Kim, C.; Choi, Y.; Park, H.; Chung, I.Y. Lysophosphatidylserine induces eosinophil extracellular trap formation and degranulation: Implications in severe asthma. Allergy 2020, 75, 3159–3170. [Google Scholar] [CrossRef] [PubMed]
- Van De Veerdonk, F.L.; Netea, M.G. New Insights in the Immunobiology of IL-1 Family Members. Front. Immunol. 2013, 4, 167. [Google Scholar] [CrossRef]
- Dunne, A.; O’Neill, L.A.J. The Interleukin-1 Receptor/Toll-Like Receptor Superfamily: Signal Transduction During Inflammation and Host Defense. Sci. STKE 2003, 2003, re3. [Google Scholar] [CrossRef]
- Schmitz, J.; Owyang, A.; Oldham, E.; Song, Y.; Murphy, E.; McClanahan, T.K.; Zurawski, G.; Moshrefi, M.; Qin, J.; Li, X.; et al. IL-33, an Interleukin-1-like Cytokine that Signals via the IL-1 Receptor-Related Protein ST2 and Induces T Helper Type 2-Associated Cytokines. Immunity 2005, 23, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Cayrol, C.; Girard, J.-P. The IL-1-like cytokine IL-33 is inactivated after maturation by caspase-1. Proc. Natl. Acad. Sci. USA 2009, 106, 9021–9026. [Google Scholar] [CrossRef] [PubMed]
- Suzukawa, M.; Koketsu, R.; Iikura, M.; Nakae, S.; Matsumoto, K.; Nagase, H.; Saito, H.; Matsushima, K.; Ohta, K.; Yamamoto, K.; et al. Interleukin-33 enhances adhesion, CD11b expression and survival in human eosinophils. Lab. Investig. 2008, 88, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Cherry, W.B.; Yoon, J.; Bartemes, K.R.; Iijima, K.; Kita, H. A novel IL-1 family cytokine, IL-33, potently activates human eosinophils. J. Allergy Clin. Immunol. 2008, 121, 1484–1490. [Google Scholar] [CrossRef] [PubMed]
- Angulo, E.L.; McKernan, E.M.; Fichtinger, P.S.; Mathur, S.K. Comparison of IL-33 and IL-5 family mediated activation of human eosinophils. PLoS ONE 2019, 14, 0217807. [Google Scholar] [CrossRef]
- Andreone, S.; Spadaro, F.; Buccione, C.; Mancini, J.; Tinari, A.; Sestili, P.; Gambardella, A.R.; Lucarini, V.; Ziccheddu, G.; Parolini, I.; et al. IL-33 Promotes CD11b/CD18-Mediated Adhesion of Eosinophils to Cancer Cells and Synapse-Polarized Degranulation Leading to Tumor Cell Killing. Cancers 2019, 11, 1664. [Google Scholar] [CrossRef]
- Pelaia, C.; Paoletti, G.; Puggioni, F.; Racca, F.; Pelaia, G.; Canonica, G.W.; Heffler, E. Interleukin-5 in the Pathophysiology of Severe Asthma. Front. Physiol. 2019, 10, 1514. [Google Scholar] [CrossRef]
- Tavernier, J.; Devos, R.; Cornelis, S.; Tuypens, T.; Van der Heyden, J.; Fiers, W.; Plaetinck, G. A human high affinity interleukin-5 receptor (IL5R) is composed of an IL5-specific α chain and a β chain shared with the receptor for GM-CSF. Cell 1991, 66, 1175–1184. [Google Scholar] [CrossRef]
- Dvorak, A.M.; Furitsu, T.; Letourneau, L.; Ishizaka, T.; Ackerman, S.J. Mature eosinophils stimulated to develop in human cord blood mononuclear cell cultures supplemented with recombinant human interleukin-5. Part I. Piecemeal degranulation of specific granules and distribution of Charcot-Leyden crystal protein. Am. J. Pathol. 1991, 138, 69–82. [Google Scholar] [PubMed]
- Dvorak, A.M.; Ackerman, S.J.; Furitsu, T.; Estrella, P.; Letourneau, L.; Ishizaka, T. Mature eosinophils stimulated to develop in human-cord blood mononuclear cell cultures supplemented with recombinant human interleukin-5. II. Vesicular transport of specific granule matrix peroxidase, a mechanism for effecting piecemeal degranulation. Am. J. Pathol. 1992, 140, 795–807. [Google Scholar]
- Kitaura, M.; Nakajima, T.; Imai, T.; Harada, S.; Combadiere, C.; Tiffany, H.L.; Murphy, P.M.; Yoshie, O. Molecular Cloning of Human Eotaxin, an Eosinophil-selective CC Chemokine, and Identification of a Specific Eosinophil Eotaxin Receptor, CC Chemokine Receptor 3. J. Biol. Chem. 1996, 271, 7725–7730. [Google Scholar] [CrossRef]
- Ogilvie, P.; Bardi, G.; Clark-Lewis, I.; Baggiolini, M.; Uguccioni, M. Eotaxin is a natural antagonist for CCR2 and an agonist for CCR5. Blood 2001, 97, 1920–1924. [Google Scholar] [CrossRef]
- Zlotnik, A.; Yoshie, O. The Chemokine Superfamily Revisited. Immunity 2012, 36, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Kampen, G.T.; Stafford, S.; Adachi, T.; Jinquan, T.; Quan, S.; Grant, J.A.; Skov, P.S.; Poulsen, L.K.; Alam, R. Eotaxin induces degranulation and chemotaxis of eosinophils through the activation of ERK2 and p38 mitogen-activated protein kinases. Blood 2000, 95, 1911–1917. [Google Scholar] [CrossRef] [PubMed]
- Bandeira-Melo, C.; Sugiyama, K.; Woods, L.J.; Weller, P.F. Cutting Edge: Eotaxin Elicits Rapid Vesicular Transport-Mediated Release of Preformed IL-4 from Human Eosinophils. J. Immunol. 2001, 166, 4813–4817. [Google Scholar] [CrossRef] [PubMed]
- Carmo, L.A.S.; Bonjour, K.; Ueki, S.; Neves, J.S.; Liu, L.; Spencer, L.A.; Dvorak, A.M.; Weller, P.F.; Melo, R.C.N. CD63 is tightly associated with intracellular, secretory events chaperoning piecemeal degranulation and compound exocytosis in human eosinophils. J. Leukoc. Biol. 2016, 100, 391–401. [Google Scholar] [CrossRef]
- Carmo, L.A.S.; Bonjour, K.; Spencer, L.A.; Weller, P.F.; Melo, R.C.N. Single-Cell Analyses of Human Eosinophils at High Resolution to Understand Compartmentalization and Vesicular Trafficking of Interferon-Gamma. Front. Immunol. 2018, 9, 1542. [Google Scholar] [CrossRef]
- El-Shazly, A.; Masuyama, K.; Nakano, K.; Eura, M.; Samejima, Y.; Ishikawa, T. Human Eotaxin Induces Eosinophil-Derived Neurotoxin Release from Normal Human Eosinophils. Int. Arch. Allergy Immunol. 1998, 117, 55–58. [Google Scholar] [CrossRef]
- Keystone, E.C.; Ware, C.F.; El-Gabalawy, H.; Guenther, L.C.; Bernstein, C.N. Tumor Necrosis Factor and Anti-Tumor Necrosis Factor Therapies. J. Rheumatol. Suppl. 2010, 85, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Uings, I.; Puxeddu, I.; Temkin, V.; Smith, S.J.; Fattah, D.; Ray, K.P.; Levi-Schaffer, F. Effects of dexamethasone on TNF-alpha-induced release of cytokines from purified human blood eosinophils. Clin. Mol. Allergy 2005, 3, 5. [Google Scholar] [CrossRef] [PubMed]
- Kankaanranta, H.; Ilmarinen, P.; Zhang, X.; Adcock, I.M.; Lahti, A.; Barnes, P.J.; Giembycz, M.A.; Lindsay, M.A.; Moilanen, E. Tumour Necrosis Factor-α Regulates Human Eosinophil Apoptosis via Ligation of TNF-Receptor 1 and Balance between NF-κB and AP-1. PLoS ONE 2014, 9, 90298. [Google Scholar] [CrossRef]
- Wardlaw, A.J.; Moqbel, R.; Cromwell, O.; Kay, A.B. Platelet-activating factor. A potent chemotactic and chemokinetic factor for human eosinophils. J. Clin. Investig. 1986, 78, 1701–1706. [Google Scholar] [CrossRef] [PubMed]
- Kimani, G.; Tonnesen, M.G.; Henson, P.M. Stimulation of eosinophil adherence to human vascular endothelial cells in vitro by platelet-activating factor. J. Immunol. 1988, 140, 3161–3166. [Google Scholar]
- Martins, M.A.; Silva, P.M.; Neto, H.C.C.F.; Bozza, P.; Dias, P.M.; Lima, M.C.; Cordeiro, R.S.; Vargaftig, B.B. Pharmacological modulation of Paf-induced rat pleurisy and its role in inflammation by zymosan. Br. J. Pharmacol. 1989, 96, 363–371. [Google Scholar] [CrossRef]
- Kato, M.; Kita, H.; Tachibana, A.; Hayashi, Y.; Tsuchida, Y.; Kimura, H. Dual Signaling and Effector Pathways Mediate Human Eosinophil Activation by Platelet-Activating Factor. Int. Arch. Allergy Immunol. 2004, 134, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Honda, Z.-I.; Nakamura, M.; Miki, I.; Minami, M.; Watanabe, T.; Seyama, Y.; Okado, H.; Toh, H.; Ito, K.; Miyamoto, T.; et al. Cloning by functional expression of platelet-activating factor receptor from guinea-pig lung. Nat. Cell Biol. 1991, 349, 342–346. [Google Scholar] [CrossRef] [PubMed]
- Ishii, S.; Kuwaki, T.; Nagase, T.; Maki, K.; Tashiro, F.; Sunaga, S.; Cao, W.-H.; Kume, K.; Fukuchi, Y.; Ikuta, K.; et al. Impaired Anaphylactic Responses with Intact Sensitivity to Endotoxin in Mice Lacking a Platelet-activating Factor Receptor. J. Exp. Med. 1998, 187, 1779–1788. [Google Scholar] [CrossRef] [PubMed]
- Ukena, D.; Krogel, C.; Dent, G.; Yukawa, T.; Sybrecht, G.; Barnes, P.J. PAF-Receptors on eosinophils: Identification with a novel ligand, [3H]WEB 2086. Biochem. Pharmacol. 1989, 38, 1702–1705. [Google Scholar] [CrossRef]
- Korth, R.-M. Specific High Affinity Binding of Platelet Activating Factor to Intact Human Blood Neutrophils and Eosinophils. Int. Arch. Allergy Immunol. 1996, 110, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Dyer, K.D.; Percopo, C.M.; Xie, Z.; Yang, Z.; Kim, J.D.; Davoine, F.; Lacy, P.; Druey, K.M.; Moqbel, R.; Rosenberg, H.F. Mouse and Human Eosinophils Degranulate in Response to Platelet-Activating Factor (PAF) and LysoPAF via a PAF-Receptor–Independent Mechanism: Evidence for a Novel Receptor. J. Immunol. 2010, 184, 6327–6334. [Google Scholar] [CrossRef] [PubMed]
- Grozdanovic, M.M.; Doyle, C.B.; Liu, L.; Maybruck, B.T.; Kwatia, M.A.; Thiyagarajan, N.; Acharya, K.R.; Ackerman, S.J. Charcot-Leyden crystal protein/galectin-10 interacts with cationic ribonucleases and is required for eosinophil granulogenesis. J. Allergy Clin. Immunol. 2020, 146, 377–389.e10. [Google Scholar] [CrossRef]
- Simon, H.-U.; Weber, M.; Becker, E.; Zilberman, Y.; Blaser, K.; Levi-Schaffer, F. Eosinophils Maintain Their Capacity to Signal and Release Eosinophil Cationic Protein Upon Repetitive Stimulation with the Same Agonist. J. Immunol. 2000, 165, 4069–4075. [Google Scholar] [CrossRef]
- Yazdanbakhsh, M.; Eckmann, C.; Koenderman, L.; Verhoeven, A.; Roos, D. Eosinophils do respond to fMLP. Blood 1987, 70, 379–383. [Google Scholar] [CrossRef]
- Malm-Erjefält, M.; Persson, C.G.A.; Erjefält, J.S. Degranulation Status of Airway Tissue Eosinophils in Mouse Models of Allergic Airway Inflammation. Am. J. Respir. Cell Mol. Biol. 2001, 24, 352–359. [Google Scholar] [CrossRef]
- Lacy, P.; Mahmudi-Azer, S.; Bablitz, B.; Hagen, S.C.; Velazquez, J.R.; Man, S.F.; Moqbel, R. Rapid mobilization of intracellularly stored RANTES in response to interferon-gamma in human eosinophils. Blood 1999, 94, 23–32. [Google Scholar] [CrossRef]
- Mahmudi-Azer, S.; Downey, G.P.; Moqbel, R. Translocation of the tetraspanin CD63 in association with human eosinophil mediator release. Blood 2002, 99, 4039–4047. [Google Scholar] [CrossRef]
- Kita, H.; Ohnishi, T.; Okubo, Y.; Weiler, D.; Abrams, J.S.; Gleich, G.J. Granulocyte/macrophage colony-stimulating factor and interleukin 3 release from human peripheral blood eosinophils and neutrophils. J. Exp. Med. 1991, 174, 745–748. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, T.; Ackerman, S.J.; Reed, C.E.; Peters, M.S.; Dunnette, S.L.; Gleich, G.J. Calcium ionophore A23187 calcium-dependent cytolytic degranulation in human eosinophils. J. Immunol. 1985, 135, 1349–1356. [Google Scholar]
- Coden, M.E.; Loffredo, L.F.; Walker, M.T.; Jeong, B.M.; Nam, K.; Bochner, B.S.; Abdala-Valencia, H.; Berdnikovs, S. Fibrinogen Is a Specific Trigger for Cytolytic Eosinophil Degranulation. J. Immunol. 2019, 204, 438–448. [Google Scholar] [CrossRef]
- Weiler, C.R.; Kita, H.; Hukee, M.; Gleich, G.J. Eosinophil viability during immunoglobulin-induced degranulation. J. Leukoc. Biol. 1996, 60, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.M.; Kim, H.J.; Kim, S.M.; Jung, Y.; Park, S.W.; Chung, I.Y. Lysophosphatidylserine receptor P2Y10: A G protein-coupled receptor that mediates eosinophil degranulation. Clin. Exp. Allergy 2018, 48, 990–999. [Google Scholar] [CrossRef] [PubMed]
- Suzukawa, M.; Iikura, M.; Koketsu, R.; Nagase, H.; Tamura, C.; Komiya, A.; Nakae, S.; Matsushima, K.; Ohta, K.; Yamamoto, K.; et al. An IL-1 Cytokine Member, IL-33, Induces Human Basophil Activation via Its ST2 Receptor. J. Immunol. 2008, 181, 5981–5989. [Google Scholar] [CrossRef]
- Arnold, I.C.; Artola-Borán, M.; De Lara, P.T.; Kyburz, A.; Taube, C.; Ottemann, K.; Broek, M.V.D.; Yousefi, S.; Simon, H.-U.; Müller, A. Eosinophils suppress Th1 responses and restrict bacterially induced gastrointestinal inflammation. J. Exp. Med. 2018, 215, 2055–2072. [Google Scholar] [CrossRef] [PubMed]
- Dias, F.F.; Amaral, K.B.; Malta, K.K.; Silva, T.P.; Rodrigues, G.S.C.; Rosa, F.M.; Rodrigues, G.O.L.; Costa, V.V.; Chiarini-Garcia, H.; Weller, P.F.; et al. Identification of Piecemeal Degranulation and Vesicular Transport of MBP-1 in Liver-Infiltrating Mouse Eosinophils During Acute Experimental Schistosoma mansoni Infection. Front. Immunol. 2018, 9, 3019. [Google Scholar] [CrossRef]
- Melo, R.C.N.; Wang, H.; Silva, T.P.; Imoto, Y.; Fujieda, S.; Fukuchi, M.; Miyabe, Y.; Hirokawa, M.; Ueki, S.; Weller, P.F. Galectin-10, the protein that forms Charcot-Leyden crystals, is not stored in granules but resides in the peripheral cytoplasm of human eosinophils. J. Leukoc. Biol. 2020, 108, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Melo, R.C.N.; Spencer, L.A.; Perez, S.A.C.; Ghiran, I.; Dvorak, A.M.; Weller, P.F. Human Eosinophils Secrete Preformed, Granule-Stored Interleukin-4 Through Distinct Vesicular Compartments. Traffic 2005, 6, 1047–1057. [Google Scholar] [CrossRef]
- Ueki, S.; Tokunaga, T.; Melo, R.C.N.; Saito, H.; Honda, K.; Fukuchi, M.; Konno, Y.; Takeda, M.; Yamamoto, Y.; Hirokawa, M.; et al. Charcot-Leyden crystal formation is closely associated with eosinophil extracellular trap cell death. Blood 2018, 132, 2183–2187. [Google Scholar] [CrossRef]
- Persson, E.K.; Verstraete, K.; Heyndrickx, I.; Gevaert, E.; Aegerter, H.; Percier, J.-M.; Deswarte, K.; Verschueren, K.H.G.; Dansercoer, A.; Gras, D.; et al. Protein crystallization promotes type 2 immunity and is reversible by antibody treatment. Science 2019, 364, eaaw4295. [Google Scholar] [CrossRef]
- Gevaert, E.; Delemarre, T.; De Volder, J.; Zhang, N.; Holtappels, G.; De Ruyck, N.; Persson, E.; Heyndrickx, I.; Verstraete, K.; Aegerter, H.; et al. Charcot-Leyden crystals promote neutrophilic inflammation in patients with nasal polyposis. J. Allergy Clin. Immunol. 2020, 145, 427–430.e4. [Google Scholar] [CrossRef]
- Rodríguez-Alcázar, J.F.; Ataide, M.; Engels, G.; Schmitt-Mabmunyo, C.; Garbi, N.; Kastenmüller, W.; Latz, E.; Franklin, B.S. Charcot–Leyden Crystals Activate the NLRP3 Inflammasome and Cause IL-1β Inflammation in Human Macrophages. J. Immunol. 2019, 202, 550–558. [Google Scholar] [CrossRef]
- Melo, R.C.N.; Liu, L.; Xenakis, J.J.; Spencer, L.A. Eosinophil-derived cytokines in health and disease: Unraveling novel mechanisms of selective secretion. Allergy 2013, 68, 274–284. [Google Scholar] [CrossRef]
- Geering, B.; Stoeckle, C.; Conus, S.; Simon, H.-U. Living and dying for inflammation: Neutrophils, eosinophils, basophils. Trends Immunol. 2013, 34, 398–409. [Google Scholar] [CrossRef]
- Mattes, J.; Foster, P.S. Regulation of eosinophil migration and Th2 cell function by IL-5 and eotaxin. Curr. Drug Target. Inflamm. Allergy 2003, 2, 169–174. [Google Scholar] [CrossRef]
- Ikutani, M.; Yanagibashi, T.; Ogasawara, M.; Tsuneyama, K.; Yamamoto, S.; Hattori, Y.; Kouro, T.; Itakura, A.; Nagai, Y.; Takaki, S.; et al. Identification of Innate IL-5–Producing Cells and Their Role in Lung Eosinophil Regulation and Antitumor Immunity. J. Immunol. 2011, 188, 703–713. [Google Scholar] [CrossRef] [PubMed]
- Carretero, R.; Sektioglu, I.M.; Garbi, N.; Salgado, O.C.; Beckhove, P.; Hämmerling, G.J. Eosinophils orchestrate cancer rejection by normalizing tumor vessels and enhancing infiltration of CD8+ T cells. Nat. Immunol. 2015, 16, 609–617. [Google Scholar] [CrossRef]
- Lucarini, V.; Ziccheddu, G.; Macchia, I.; La Sorsa, V.; Peschiaroli, F.; Buccione, C.; Sistigu, A.; Sanchez, M.; Andreone, S.; D’Urso, M.T.; et al. IL-33 restricts tumor growth and inhibits pulmonary metastasis in melanoma-bearing mice through eosinophils. Oncoimmunology 2017, 6, 1317420. [Google Scholar] [CrossRef] [PubMed]
- Reichman, H.; Itan, M.; Rozenberg, P.; Yarmolovski, T.; Brazowski, E.; Varol, C.; Gluck, N.; Shapira, S.; Arber, N.; Qimron, U.; et al. Activated Eosinophils Exert Antitumorigenic Activities in Colorectal Cancer. Cancer Immunol. Res. 2019, 7, 388–400. [Google Scholar] [CrossRef] [PubMed]
- Simson, L.; Ellyard, J.I.; Dent, L.A.; Matthaei, K.I.; Rothenberg, M.E.; Foster, P.S.; Smyth, M.J.; Parish, C. Regulation of Carcinogenesis by IL-5 and CCL11: A Potential Role for Eosinophils in Tumor Immune Surveillance. J. Immunol. 2007, 178, 4222–4229. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, S.; Konishi, Y.; Nishio, Y.; Fujikawa-Adachi, K.; Tominaga, A. Antitumor Activity of Eosinophils Activated by IL-5 and Eotaxin against Hepatocellular Carcinoma. DNA Cell Biol. 2004, 23, 549–560. [Google Scholar] [CrossRef] [PubMed]
- Hollande, C.; Boussier, J.; Ziai, J.; Nozawa, T.; Bondet, V.; Phung, W.; Lu, B.; Duffy, D.; Paradis, V.; Mallet, V.; et al. Inhibition of the dipeptidyl peptidase DPP4 (CD26) reveals IL-33-dependent eosinophil-mediated control of tumor growth. Nat. Immunol. 2019, 20, 257–264. [Google Scholar] [CrossRef]
- Moy, J.N.; Gleich, G.J.; Thomas, L.L. Noncytotoxic activation of neutrophils by eosinophil granule major basic protein. Effect on superoxide anion generation and lysosomal enzyme release. J. Immunol. 1990, 145, 2626–2632. [Google Scholar]
- O’Donnell, M.C.; Ackerman, S.J.; Gleich, G.J.; Thomas, L.L. Activation of basophil and mast cell histamine release by eosinophil granule major basic protein. J. Exp. Med. 1983, 157, 1981–1991. [Google Scholar] [CrossRef]
- Lotfi, R.; Lotze, M.T. Eosinophils induce DC maturation, regulating immunity. J. Leukoc. Biol. 2007, 83, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Morgan, R.K.; Costello, R.W.; Durcan, N.; Kingham, P.J.; Gleich, G.J.; McLean, W.G.; Walsh, M.-T. Diverse Effects of Eosinophil Cationic Granule Proteins on IMR-32 Nerve Cell Signaling and Survival. Am. J. Respir. Cell Mol. Biol. 2005, 33, 169–177. [Google Scholar] [CrossRef]
- Butterworth, A.E.; Wassom, D.L.; Gleich, G.J.; Loegering, D.A.; David, J.R. Damage to schistosomula of Schistosoma mansoni induced directly by eosinophil major basic protein. J. Immunol. 1979, 122, 221–229. [Google Scholar] [PubMed]
- Lehrer, R.I.; Szklarek, D.; Barton, A.; Ganz, T.; Hamann, K.J.; Gleich, G.J. Antibacterial properties of eosinophil major basic protein and eosinophil cationic protein. J. Immunol. 1989, 142, 4428–4434. [Google Scholar]
- Soragni, A.; Yousefi, S.; Stoeckle, C.; Soriaga, A.B.; Sawaya, M.R.; Kozlowski, E.; Schmid, I.; Radonjic-Hoesli, S.; Boutet, S.; Williams, G.J.; et al. Toxicity of Eosinophil MBP Is Repressed by Intracellular Crystallization and Promoted by Extracellular Aggregation. Mol. Cell 2015, 57, 1011–1021. [Google Scholar] [CrossRef]
- Peterson, K.A.; Gleich, G.J.; Limaye, N.S.; Crispin, H.; Robson, J.; Fang, J.; Saffari, H.; Clayton, F.; Leiferman, K.M. Eosinophil granule major basic protein 1 deposition in eosinophilic esophagitis correlates with symptoms independent of eosinophil counts. Dis. Esophagus 2019, 32. [Google Scholar] [CrossRef]
- Piliponsky, A.M.; Gleich, G.J.; Bar, I.; Levi-Schaffer, F. Effects of eosinophils on mast cells: A new pathway for the perpetuation of allergic inflammation. Mol. Immunol. 2002, 38, 1369–1372. [Google Scholar] [CrossRef]
- Ackerman, S.J.; Gleich, G.J.; Loegering, D.A.; Richardson, B.A.; Butterworth, A.E. Comparative Toxicity of Purified Human Eosinophil Granule Cationic Proteins for Schistosomula of Schistosoma Mansoni. Am. J. Trop. Med. Hyg. 1985, 34, 735–745. [Google Scholar] [CrossRef]
- Venge, P.; Byström, J.; Carlson, M.; Hâkansson, L.; Karawacjzyk, M.; Peterson, C.; Sevéus, L.; Trulson, A. Eosinophil cationic protein (ECP): Molecular and biological properties and the use of ECP as a marker of eosinophil activation in disease. Clin. Exp. Allergy 1999, 29, 1172–1186. [Google Scholar] [CrossRef]
- Bystrom, J.; Amin, K.; Bishop-Bailey, D. Analysing the eosinophil cationic protein—A clue to the function of the eosinophil granulocyte. Respir. Res. 2011, 12, 10. [Google Scholar] [CrossRef]
- Niccoli, G.; Ferrante, G.; Cosentino, N.; Conte, M.; Belloni, F.; Marino, M.; Bacà, M.; Montone, R.A.; Sabato, V.; Schiavino, D.; et al. Eosinophil cationic protein: A new biomarker of coronary atherosclerosis. Atherosclerosis 2010, 211, 606–611. [Google Scholar] [CrossRef] [PubMed]
- Lind, L.; Siegbahn, A.; Lindahl, B.; Stenemo, M.; Sundstrom, J.; Ärnlöv, J. Discovery of New Risk Markers for Ischemic Stroke Using a Novel Targeted Proteomics Chip. Stroke 2015, 46, 3340–3347. [Google Scholar] [CrossRef] [PubMed]
- Amber, K.T.; Chernyavsky, A.; Agnoletti, A.F.; Cozzani, E.; Grando, S.A. Mechanisms of pathogenic effects of eosinophil cationic protein and eosinophil-derived neurotoxin on human keratinocytes. Exp. Dermatol. 2018, 27, 1322–1327. [Google Scholar] [CrossRef]
- Domachowske, J.B.; Bonville, C.A.; Dyer, K.D.; Rosenberg, H.F. Evolution of antiviral activity in the ribonuclease A gene superfamily: Evidence for a specific interaction between eosinophil-derived neurotoxin (EDN/RNase 2) and respiratory syncytial virus. Nucleic Acids Res. 1998, 26, 5327–5332. [Google Scholar] [CrossRef]
- Rugeles, M.T.; Trubey, C.M.; Bedoya, V.I.; Pinto, L.A.; Oppenheim, J.J.; Rybak, S.M.; Shearer, G.M. Ribonuclease is partly responsible for the HIV-1 inhibitory effect activated by HLA alloantigen recognition. AIDS 2003, 17, 481–486. [Google Scholar] [CrossRef]
- Liu, J.; Li, Y.-H.; Xue, C.-F.; Ding, J.; Gong, W.-D.; Zhao, Y.; Huang, Y.-X. Targeted ribonuclease can inhibit replication of hepatitis B virus. World J. Gastroenterol. 2003, 9, 295–299. [Google Scholar] [CrossRef]
- Tsuda, T.; Maeda, Y.; Nishide, M.; Koyama, S.; Hayama, Y.; Nojima, S.; Takamatsu, H.; Okuzaki, D.; Kinehara, Y.; Kato, Y.; et al. Eosinophil-derived neurotoxin enhances airway remodeling in eosinophilic chronic rhinosinusitis and correlates with disease severity. Int. Immunol. 2018, 31, 33–40. [Google Scholar] [CrossRef]
- DeChatelet, L.; Migler, R.; Shirley, P.; Muss, H.; Szejda, P.; Bass, D. Comparison of intracellular bactericidal activities of human neutrophils and eosinophils. Blood 1978, 52, 609–617. [Google Scholar] [CrossRef]
- Klebanoff, S.J.; Coombs, R.W. Virucidal Effect of Stimulated Eosinophils on Human Immunodeficiency Virus Type 1. AIDS Res. Hum. Retrovir. 1996, 12, 25–29. [Google Scholar] [CrossRef]
- Jong, E.C.; Mahmoud, A.A.; Klebanoff, S.J. Peroxidase-mediated toxicity to schistosomula of Schistosoma mansoni. J. Immunol. 1981, 126, 468–471. [Google Scholar] [PubMed]
- Jong, E.C.; Klebanoff, S.J. Eosinophil-mediated mammalian tumor cell cytotoxicity: Role of the peroxidase system. J. Immunol. 1980, 124, 1949–1953. [Google Scholar]
- Slungaard, A.; Mahoney, J.R. Bromide-dependent toxicity of eosinophil peroxidase for endothelium and isolated working rat hearts: A model for eosinophilic endocarditis. J. Exp. Med. 1991, 173, 117–126. [Google Scholar] [CrossRef]
- Wright, B.L.; Doyle, A.D.; Shim, K.P.; Pai, R.K.; Barshow, S.M.; Horsley-Silva, J.L.; Luo, H.; Rank, M.A.; Jacobsen, E.A.; Katzka, D.A.; et al. Image Analysis of Eosinophil Peroxidase Immunohistochemistry for Diagnosis of Eosinophilic Esophagitis. Dig. Dis. Sci. 2021, 66, 775–783. [Google Scholar] [CrossRef]
- Wright, B.L.; Ochkur, S.I.; Olson, N.S.; Shim, K.P.; Jacobsen, E.A.; Rank, M.A.; Dellon, E.S.; Lee, J.J. Normalized serum eosinophil peroxidase levels are inversely correlated with esophageal eosinophilia in eosinophilic esophagitis. Dis. Esophagus 2018, 31. [Google Scholar] [CrossRef] [PubMed]
- Malm-Erjefalt, M.; Greiff, L.; Ankerst, J.; Andersson, M.; Wallengren, J.; Cardell, L.-O.; Rak, S.; Persson, C.G.A.; Erjefalt, J.S. Circulating eosinophils in asthma, allergic rhinitis, and atopic dermatitis lack morphological signs of degranulation. Clin. Exp. Allergy 2005, 35, 1334–1340. [Google Scholar] [CrossRef]
- Rajamanickam, A.; Munisankar, S.; Bhootra, Y.; Dolla, C.K.; Nutman, T.B.; Babu, S. Elevated Systemic Levels of Eosinophil, Neutrophil, and Mast Cell Granular Proteins in Strongyloides Stercoralis Infection that Diminish following Treatment. Front. Immunol. 2018, 9, 207. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.; Straumann, A.; Yousefi, S.; Simon, H.-U.; Radonjic-Hösli, S. Active eosinophilic esophagitis is characterized by epithelial barrier defects and eosinophil extracellular trap formation. Allergy 2015, 70, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Thakral, D.; Agarwal, P.; Saran, R.K.; Saluja, S. Significance of Charcot Leyden crystals in liver cytology-A case report. Diagn. Cytopathol. 2015, 43, 392–394. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Tutumi, T.; Watanabe, K. Middle ear effusion and fungi. Ann. Otol. Rhinol. Laryngol. 2012, 121, 609–614. [Google Scholar] [CrossRef]
- Correll, D.P.; Luzi, S.A.; Nelson, B.L. Allergic Fungal Sinusitis. Head Neck Pathol. 2014, 9, 488–491. [Google Scholar] [CrossRef] [PubMed]
- Charatcharoenwitthaya, P.; Apisarnthanarak, P.; Pongpaibul, A.; Boonyaarunnate, T. Eosinophilic pseudotumour of the liver. Liver Int. 2011, 32, 311. [Google Scholar] [CrossRef]
- Fjaerli, H.-O.; Bukholm, G.; Krog, A.; Skjaeret, C.; Holden, M.; Nakstad, B. Whole blood gene expression in infants with respiratory syncytial virus bronchiolitis. BMC Infect. Dis. 2006, 6, 175. [Google Scholar] [CrossRef] [PubMed]
- De Re, V.; Simula, M.P.; Caggiari, L.; Orzes, N.; Spina, M.; Da Ponte, A.; De Appollonia, L.; Dolcetti, R.; Canzonieri, V.; Cannizzaro, R. Proteins specifically hyperexpressed in a coeliac disease patient with aberrant T cells. Clin. Exp. Immunol. 2007, 148, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Udell, I.J.; Gleich, G.J.; Allansmith, M.R.; Ackerman, S.J.; Abelson, M.B. Eosinophil Granule Major Basic Protein and Charcot-Leyden Crystal Protein in Human Tears. Am. J. Ophthalmol. 1981, 92, 824–828. [Google Scholar] [CrossRef]
- Dor, P.J.; Ackerman, S.J.; Gleich, G.J. Charcot-Leyden crystal protein and eosinophil granule major basic protein in sputum of patients with respiratory diseases. Am. Rev. Respir. Dis. 1984, 130, 1072–1077. [Google Scholar] [CrossRef]
- Ghafouri, B.; Irander, K.; Lindbom, J.; Tagesson, C.; Lindahl, M. Comparative Proteomics of Nasal Fluid in Seasonal Allergic Rhinitis. J. Proteome Res. 2006, 5, 330–338. [Google Scholar] [CrossRef]
- Hirszel, P.; Cashell, A.W.; Whelan, T.V.; Dolan, R.; Yoshihashi, A. Urinary Charcot-Leyden Crystals in the Hypereosinophilic Syndrome With Acute Renal Failure. Am. J. Kidney Dis. 1988, 12, 319–322. [Google Scholar] [CrossRef]
- Baines, K.J.; Simpson, J.L.; Wood, L.; Scott, R.J.; Fibbens, N.L.; Powell, H.; Cowan, D.C.; Taylor, D.R.; Cowan, J.O.; Gibson, P.G. Sputum gene expression signature of 6 biomarkers discriminates asthma inflammatory phenotypes. J. Allergy Clin. Immunol. 2014, 133, 997–1007. [Google Scholar] [CrossRef]
- Chua, J.C.; Douglass, J.A.; Gillman, A.; O’Hehir, R.E.; Meeusen, E.N. Galectin-10, a Potential Biomarker of Eosinophilic Airway Inflammation. PLoS ONE 2012, 7, 42549. [Google Scholar] [CrossRef]
- Martin, L.B.; Kita, H.; Leiferman, K.M.; Gleich, G.J. Eosinophils in Allergy: Role in Disease, Degranulation, and Cytokines. Int. Arch. Allergy Immunol. 1996, 109, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Ying, S.; Humbert, M.; Barkans, J.; Corrigan, C.J.; Pfister, R.; Menz, G.; Larche, M.; Robinson, D.S.; Durham, S.R.; Kay, A.B. Expression of IL-4 and IL-5 mRNA and protein product by CD4+ and CD8+ T cells, eosinophils, and mast cells in bronchial biopsies obtained from atopic and nonatopic (intrinsic) asthmatics. J. Immunol. 1997, 158, 3539–3544. [Google Scholar]
- Desreumaux, P.; Janin, A.; Colombel, J.F.; Prin, L.; Plumas, J.; Emilie, D.; Torpier, G.; Capron, A.; Capron, M. Interleukin 5 messenger RNA expression by eosinophils in the intestinal mucosa of patients with coeliac disease. J. Exp. Med. 1992, 175, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Dubucquoi, S.; Desreumaux, P.; Janin, A.; Klein, O.; Goldman, M.; Tavernier, J.; Capron, A.; Capron, M. Interleukin 5 synthesis by eosinophils: Association with granules and immunoglobulin-dependent secretion. J. Exp. Med. 1994, 179, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Desreumaux, P.; Janin, A.; Dubucquoi, S.; Copin, M.C.; Torpier, G.; Capron, A.; Capron, M.; Prin, L. Synthesis of Interleukin-5 by Activated Eosinophils in Patients with Eosinophilic Heart Diseases. Blood 1993, 82, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Almas, S.; Fayad, N.; Srivastava, O.; Siddique, M.; Touret, N.; Lacy, P. Cytokine trafficking of IL-9 and IL-13 through TfnRc + vesicles in activated human eosinophils. J. Leukoc. Biol. 2021, 109, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, R.; Osuka, K.; Aoyama, M.; Miyachi, S.; Takayasu, M. Expressions of Eotaxin-3, Interleukin-5, and Eosinophil-Derived Neurotoxin in Chronic Subdural Hematoma Fluids. J. Neurotrauma 2018, 35, 2242–2249. [Google Scholar] [CrossRef]
- Kienzl, M.; Hasenoehrl, C.; Valadez-Cosmes, P.; Maitz, K.; Sarsembayeva, A.; Sturm, E.; Heinemann, A.; Kargl, J.; Schicho, R. IL-33 reduces tumor growth in models of colorectal cancer with the help of eosinophils. OncoImmunology 2020, 9, 1776059. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.-H.; Liao, B.; Liu, K.; Liu, Y.-H. Effect of RNA interference therapy on the mice eosinophils CCR3 gene and granule protein in the murine model of allergic rhinitis. Asian Pac. J. Trop. Med. 2014, 7, 226–230. [Google Scholar] [CrossRef]
- Zhu, X.-H.; Liao, B.; Xu, Y.; Liu, K.; Huang, Y.; Huang, Q.-L.; Liu, Y.-H. Downregulation of mouse CCR3 by lentiviral shRNA inhibits proliferation and induces apoptosis of mouse eosinophils. Mol. Med. Rep. 2016, 15, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Knuplez, E.; Curcic, S.; Theiler, A.; Bärnthaler, T.; Trakaki, A.; Trieb, M.; Holzer, M.; Heinemann, A.; Zimmermann, R.; Sturm, E.M.; et al. Lysophosphatidylcholines inhibit human eosinophil activation and suppress eosinophil migration in vivo. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2020, 1865, 158686. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fettrelet, T.; Gigon, L.; Karaulov, A.; Yousefi, S.; Simon, H.-U. The Enigma of Eosinophil Degranulation. Int. J. Mol. Sci. 2021, 22, 7091. https://doi.org/10.3390/ijms22137091
Fettrelet T, Gigon L, Karaulov A, Yousefi S, Simon H-U. The Enigma of Eosinophil Degranulation. International Journal of Molecular Sciences. 2021; 22(13):7091. https://doi.org/10.3390/ijms22137091
Chicago/Turabian StyleFettrelet, Timothée, Lea Gigon, Alexander Karaulov, Shida Yousefi, and Hans-Uwe Simon. 2021. "The Enigma of Eosinophil Degranulation" International Journal of Molecular Sciences 22, no. 13: 7091. https://doi.org/10.3390/ijms22137091
APA StyleFettrelet, T., Gigon, L., Karaulov, A., Yousefi, S., & Simon, H.-U. (2021). The Enigma of Eosinophil Degranulation. International Journal of Molecular Sciences, 22(13), 7091. https://doi.org/10.3390/ijms22137091