
Endothelial Progenitor Cell-Derived Extracellular Vesicles: Potential Therapeutic Application in Tissue Repair and Regeneration



Abstract
1. Introduction
2. EV Isolation and Characterization
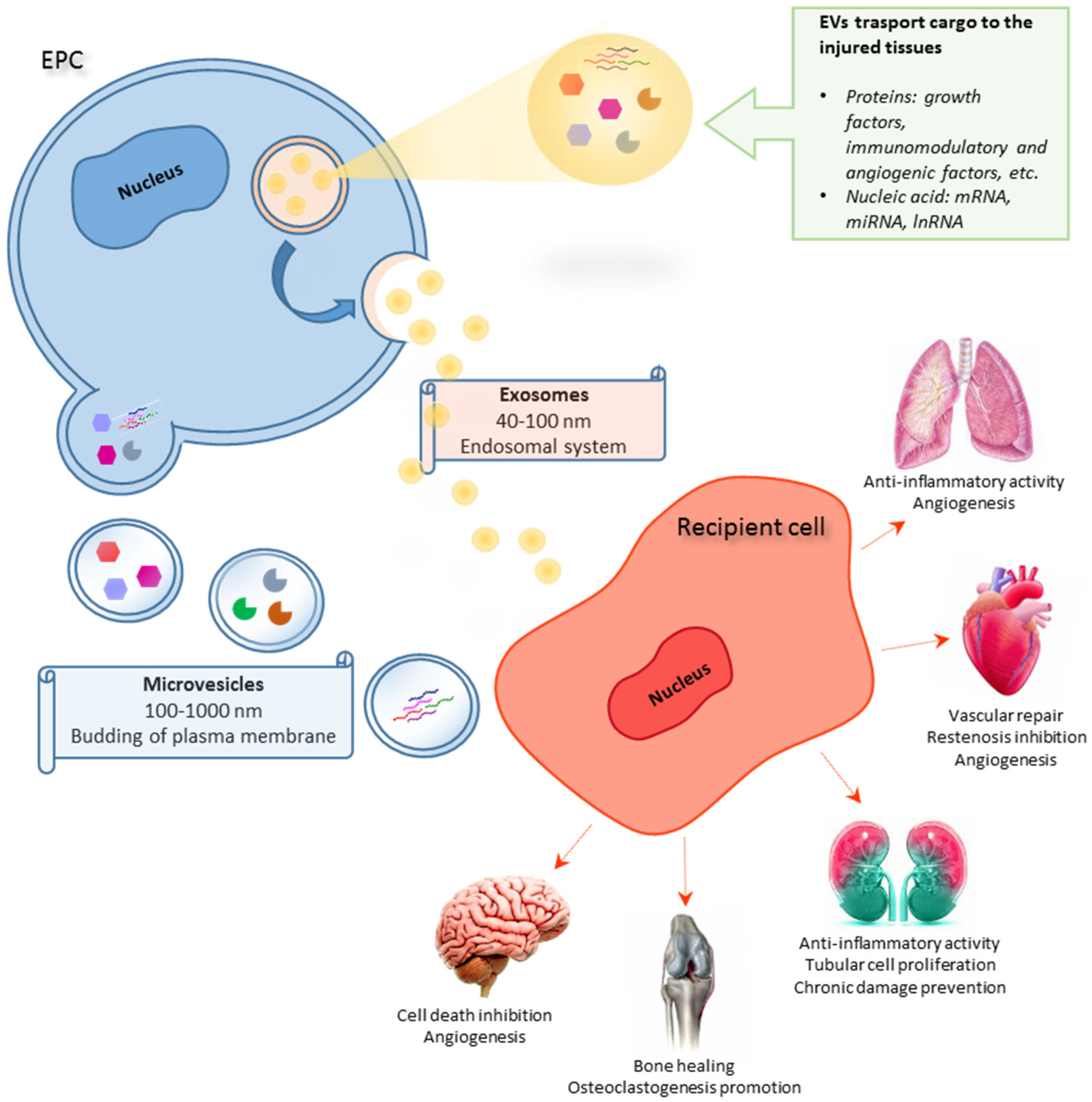
3. The Role of EPC-Derived EVs in Tissue Repair and Regeneration
3.1. EPC-Derived EVs in Cardiovascular Repair
3.2. EPC-Derived EVs in the Treatment of Diabetes
3.3. EPC-Derived EVs in Kidney Pathology
3.4. EPC-Derived EVs in Bone Healing
3.5. EPC-Derived EVs in Brain Regeneration
3.6. EPC-Derived EVs in Lung Repair
4. Modified EPC-Derived EVs as a Therapeutic Strategy in Tissue Repair and Regeneration
5. Challenges and Future Perspective
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Urbich, C.; Dimmeler, S. Endothelial progenitor cells: Characterization and role in vascular biology. Circ. Res. 2004, 95, 343–353. [Google Scholar] [CrossRef]
- Asahara, T.; Murohara, T.; Sullivan, A.; Silver, M.; Van Der Zee, R.; Li, T.; Witzenbichler, B.; Schatteman, G.; Isner, J.M. Isolation of putative progenitor endothelial cells for angiogenesis. Science 1997, 275, 964–966. [Google Scholar] [CrossRef]
- Kalka, C.; Masuda, H.; Takahashi, T.; Kalka-Moll, W.M.; Silver, M.; Kearney, M.; Li, T.; Isner, J.M.; Asahara, T. Transplantation of ex vivo expanded endothelial progenitor cells for therapeutic neovascularization. Proc. Natl. Acad. Sci. USA 2000, 97, 3422–3427. [Google Scholar] [CrossRef]
- Loomans, C.; Wan, H.; De Crom, R.; Van Haperen, R.; De Boer, H.; Leenen, P.; Drexhage, H.; Rabelink, T.; Van Zonneveld, A.; Staal, F. Angiogenic Murine Endothelial Progenitor Cells Are Derived From a Myeloid Bone Marrow Fraction and Can Be Identified by Endothelial NO Synthase Expression. Arter. Thromb. Vasc. Biol. 2006, 26, 1760–1767. [Google Scholar] [CrossRef] [PubMed]
- Ratajczak, J.; Kucia, M.; Mierzejewska, K.; Marlicz, W.; Pietrzkowski, Z.; Wojakowski, W.; Greco, N.J.; Tendera, M.; Ratajczak, M.Z. Paracrine Proangiopoietic Effects of Human Umbilical Cord Blood-Derived Purified CD133+Cells—Implications for Stem Cell Therapies in Regenerative Medicine. Stem Cells Dev. 2013, 22, 422–430. [Google Scholar] [CrossRef]
- Hur, J.; Yoon, C.-H.; Kim, H.-S.; Choi, J.-H.; Kang, H.-J.; Hwang, K.-K.; Oh, B.-H.; Lee, M.-M.; Park, Y.-B. Characterization of Two Types of Endothelial Progenitor Cells and Their Different Contributions to Neovasculogenesis. Arter. Thromb. Vasc. Biol. 2004, 24, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Abdelgawad, M.E.; Desterke, C.; Uzan, G.; Naserian, S. Single-cell transcriptomic profiling and characterization of endothelial progenitor cells: New approach for finding novel markers. Stem Cell Res. Ther. 2021, 12, 1–16. [Google Scholar] [CrossRef]
- Bouvard, C.; Gafsou, B.; Dizier, B.; Galy-Fauroux, I.; Lokajczyk, A.; Boisson-Vidal, C.; Fischer, A.-M.; Helley, D. α6-Integrin Subunit Plays a Major Role in the Proangiogenic Properties of Endothelial Progenitor Cells. Arter. Thromb. Vasc. Biol. 2010, 30, 1569–1575. [Google Scholar] [CrossRef]
- Ingram, D.A.; Mead, L.E.; Tanaka, H.; Meade, V.; Fenoglio, A.; Mortell, K.; Pollok, K.; Ferkowicz, M.J.; Gilley, D.; Yoder, M.C. Identification of a novel hierarchy of endothelial progenitor cells using human peripheral and umbilical cord blood. Blood 2004, 104, 2752–2760. [Google Scholar] [CrossRef]
- Yong, T.; Zhang, X.; Bie, N.; Zhang, H.; Zhang, X.; Li, F.; Hakeem, A.; Hu, J.; Gan, L.; Santos, H.A.; et al. Tumor exosome-based nanoparticles are efficient drug carriers for chemotherapy. Nat. Commun. 2019, 10, 1–16. [Google Scholar] [CrossRef]
- Taylor, D.D.; Gercel-Taylor, C. The origin, function, and diagnostic potential of RNA within extracellular vesicles present in human biological fluids. Front. Genet. 2013, 4, 142. [Google Scholar] [CrossRef]
- Mol, E.A.; Goumans, M.-J.; Doevendans, P.A.; Sluijter, J.P.; Vader, P. Higher functionality of extracellular vesicles isolated using size-exclusion chromatography compared to ultracentrifugation. Nanomed. Nanotechnol. Biol. Med. 2017, 13, 2061–2065. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, A.-K.; De Miroschedji, K.; Doeppner, T.R.; Börger, V.; Ruesing, J.; Rebmann, V.; Durst, S.; Jansen, S.; Bremer, M.; Behrmann, E.; et al. Precipitation with polyethylene glycol followed by washing and pelleting by ultracentrifugation enriches extracellular vesicles from tissue culture supernatants in small and large scales. J. Extracell. Vesicles 2018, 7, 1528109. [Google Scholar] [CrossRef]
- De Jong, O.G.; Van Balkom, B.W.M.; Schiffelers, R.M.; Bouten, C.; Verhaar, M. Extracellular Vesicles: Potential Roles in Regenerative Medicine. Front. Immunol. 2014, 5, 608. [Google Scholar] [CrossRef]
- Konoshenko, M.Y.; Lekchnov, E.A.; Vlassov, A.V.; Laktionov, P.P. Isolation of Extracellular Vesicles: General Methodologies and Latest Trends. BioMed Res. Int. 2018, 2018, 1–27. [Google Scholar] [CrossRef]
- Livshits, M.A.; Khomyakova, E.; Evtushenko, E.G.; Lazarev, V.N.; Kulemin, N.; Semina, S.E.; Generozov, E.V.; Govorun, V.M. Isolation of exosomes by differential centrifugation: Theoretical analysis of a commonly used protocol. Sci. Rep. 2015, 5, 17319. [Google Scholar] [CrossRef]
- Bryzgunova, O.E.; Zaripov, M.M.; Skvortsova, T.E.; Lekchnov, E.A.; Grigor’Eva, A.E.; Zaporozhchenko, I.A.; Morozkin, E.S.; Ryabchikova, E.I.; Yurchenko, Y.B.; Voitsitskiy, V.E.; et al. Comparative Study of Extracellular Vesicles from the Urine of Healthy Individuals and Prostate Cancer Patients. PLoS ONE 2016, 11, e0157566. [Google Scholar] [CrossRef]
- Tataruch-Weinert, D.; Musante, L.; Kretz, O.; Holthofer, H. Urinary extracellular vesicles for RNA extraction: Optimization of a protocol devoid of prokaryote contamination. J. Extracell. Vesicles 2016, 5, 30281. [Google Scholar] [CrossRef]
- Cheruvanky, A.; Zhou, H.; Pisitkun, T.; Kopp, J.B.; Knepper, M.A.; Yuen, P.S.T.; Star, R.A. Rapid isolation of urinary exosomal biomarkers using a nanomembrane ultrafiltration concentrator. Am. J. Physiol. Physiol. 2007, 292, F1657–F1661. [Google Scholar] [CrossRef]
- Rood, I.M.; Deegens, J.K.; Merchant, M.L.; Tamboer, W.P.; Wilkey, D.W.; Wetzels, J.F.; Klein, J.B. Comparison of three methods for isolation of urinary microvesicles to identify biomarkers of nephrotic syndrome. Kidney Int. 2010, 78, 810–816. [Google Scholar] [CrossRef]
- Gámez-Valero, A.; Monguió-Tortajada, M.; Carreras-Planella, L.; Franquesa, M.; Beyer, K.; Borràs, F.E. Size-Exclusion Chromatography-based isolation minimally alters Extracellular Vesicles’ characteristics compared to precipitating agents. Sci. Rep. 2016, 6, srep33641. [Google Scholar] [CrossRef]
- Salih, M.; Zietse, R.; Hoorn, E.J. Urinary extracellular vesicles and the kidney: Biomarkers and beyond. Am. J. Physiol. Physiol. 2014, 306, F1251–F1259. [Google Scholar] [CrossRef]
- Taylor, D.D.; Shah, S. Methods of isolating extracellular vesicles impact down-stream analyses of their cargoes. Methods 2015, 87, 3–10. [Google Scholar] [CrossRef]
- Van Deun, J.; Mestdagh, P.; Sormunen, R.; Cocquyt, V.; Vermaelen, K.; Vandesompele, J.; Bracke, M.; De Wever, O.; Hendrix, A. The impact of disparate isolation methods for extracellular vesicles on downstream RNA profiling. J. Extracell. Vesicles 2014, 18, 3. [Google Scholar] [CrossRef]
- Xing, C.; Yin, P.; Peng, Z.; Zhang, H. Engineering Mono-Chalcogen Nanomaterials for Omnipotent Anticancer Applications: Progress and Challenges. Adv. Heal. Mater. 2020, 9. [Google Scholar] [CrossRef]
- Zaborowski, M.P.; Balaj, L.; Breakefield, X.O.; Lai, C.P.-K. Extracellular Vesicles: Composition, Biological Relevance, and Methods of Study. Bioscience 2015, 65, 783–797. [Google Scholar] [CrossRef]
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef]
- Yuana, Y.; Sturk, A.; Nieuwland, R. Extracellular vesicles in physiological and pathological conditions. Blood Rev. 2013, 27, 31–39. [Google Scholar] [CrossRef]
- Sokolova, V.; Ludwig, A.-K.; Hornung, S.; Rotan, O.; Horn, P.A.; Epple, M.; Giebel, B. Characterisation of exosomes derived from human cells by nanoparticle tracking analysis and scanning electron microscopy. Colloids Surfaces B Biointerfaces 2011, 87, 146–150. [Google Scholar] [CrossRef]
- Mendt, M.; Rezvani, K.; Shpall, E. Mesenchymal stem cell-derived exosomes for clinical use. Bone Marrow Transplant. 2019, 54, 789–792. [Google Scholar] [CrossRef]
- Van Camp, G. Cardiovascular disease prevention. Acta Clin. Belg. 2014, 69, 407–411. [Google Scholar] [CrossRef]
- Kaushik, K.; Das, A. Endothelial progenitor cell therapy for chronic wound tissue regeneration. Cytotherapy 2019, 21, 1137–1150. [Google Scholar] [CrossRef]
- Alvarez-Erviti, L.; Seow, Y.; Yin, H.; Betts, C.; Lakhal, S.; Wood, M.J.A. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat. Biotechnol. 2011, 29, 341–345. [Google Scholar] [CrossRef]
- El-Andaloussi, S.; Lee, Y.; Lakhal-Littleton, S.; Li, J.; Seow, Y.; Gardiner, C.; Alvarez-Erviti, L.; Sargent, I.L.; Wood, M.J.A. Exosome-mediated delivery of siRNA in vitro and in vivo. Nat. Protoc. 2012, 7, 2112–2126. [Google Scholar] [CrossRef]
- Siontis, G.C.M.; Stefanini, G.G.; Mavridis, D.; Siontis, K.C.; Alfonso, F.; Pérez-Vizcayno, M.J.; Byrne, R.; Kastrati, A.; Meier, B.; Salanti, G.; et al. Percutaneous coronary interventional strategies for treatment of in-stent restenosis: A network meta-analysis. Lancet 2015, 386, 655–664. [Google Scholar] [CrossRef]
- Stanziale, S.F.; Wholey, M.H.; Boules, T.N.; Selzer, F.; Makaroun, M.S. Determining In-Stent Stenosis of Carotid Arteries by Duplex Ultrasound Criteria. J. Endovasc. Ther. 2005, 12, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Kong, J.; Wang, F.; Zhang, J.; Cui, Y.; Pan, L.; Zhang, W.; Wen, J.; Liu, P. Exosomes of Endothelial Progenitor Cells Inhibit Neointima Formation After Carotid Artery Injury. J. Surg. Res. 2018, 232, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Baron, M.; Boulanger, C.M.; Staels, B.; Tailleux, A. Cell-derived microparticles in atherosclerosis: Biomarkers and targets for pharmacological modulation? J. Cell. Mol. Med. 2011, 16, 1365–1376. [Google Scholar] [CrossRef] [PubMed]
- Georgescu, A.; Alexandru, N.; Popov, D.; Amuzescu, M.; Andrei, E.; Zamfir, C.; Maniu, H.; Badila, A. Chronic venous insufficiency is associated with elevated level of circulating microparticles. J. Thromb. Haemost. 2009, 7, 1566–1575. [Google Scholar] [CrossRef] [PubMed]
- Alexandru, N.; Andrei, E.; Safciuc, F.; Dragan, E.; Balahura, A.M.; Badila, E.; Georgescu, A. Intravenous Administration of Allogenic Cell-Derived Microvesicles of Healthy Origins Defends Against Atherosclerotic Cardiovascular Disease Development by a Direct Action on Endothelial Progenitor Cells. Cells 2020, 9, 423. [Google Scholar] [CrossRef]
- Gallet, R.; Dawkins, J.; Valle, J.; Simsolo, E.; De Couto, G.; Middleton, R.; Tseliou, E.; Luthringer, D.; Kreke, M.; Smith, R.R.; et al. Exosomes secreted by cardiosphere-derived cells reduce scarring, attenuate adverse remodelling, and improve function in acute and chronic porcine myocardial infarction. Eur. Heart J. 2017, 38, 201–211. [Google Scholar] [CrossRef]
- Ibrahim, A.G.-E.; Cheng, K.; Marbán, E. Exosomes as Critical Agents of Cardiac Regeneration Triggered by Cell Therapy. Stem Cell Rep. 2014, 2, 606–619. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.W.; Wang, L.L.; Zaman, S.; Gordon, J.; Arisi, M.F.; Venkataraman, C.M.; Chung, J.J.; Hung, G.; Gaffey, A.C.; Spruce, L.A.; et al. Sustained release of endothelial progenitor cell-derived extracellular vesicles from shear-thinning hydrogels improves angiogenesis and promotes function after myocardial infarction. Cardiovasc. Res. 2018, 114, 1029–1040. [Google Scholar] [CrossRef]
- Lin, F.; Zeng, Z.; Song, Y.; Li, L.; Wu, Z.; Zhang, X.; Li, Z.; Ke, X.; Hu, X. YBX-1 mediated sorting of miR-133 into hypoxia/reoxygenation-induced EPC-derived exosomes to increase fibroblast angiogenesis and MEndoT. Stem Cell Res. Ther. 2019, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ubil, E.; Duan, J.; Pillai, I.; Rosa-Garrido, M.; Wu, Y.; Bargiacchi, F.; Lu, Y.; Stanbouly, S.; Huang, J.; Rojas, M.; et al. Mesenchymal–endothelial transition contributes to cardiac neovascularization. Nat. Cell Biol. 2014, 514, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Okayama, K.; Azuma, J.; Dosaka, N.; Iekushi, K.; Sanada, F.; Kusunoki, H.; Iwabayashi, M.; Rakugi, H.; Taniyama, Y.; Morishita, R. Hepatocyte Growth Factor Reduces Cardiac Fibrosis by Inhibiting Endothelial-Mesenchymal Transition. Hypertension 2012, 59, 958–965. [Google Scholar] [CrossRef]
- Ke, X.; Yang, D.; Liang, J.; Wang, X.; Wu, S.; Wang, X.; Hu, C. Human Endothelial Progenitor Cell-Derived Exosomes Increase Proliferation and Angiogenesis in Cardiac Fibroblasts by Promoting the Mesenchymal–Endothelial Transition and Reducing High Mobility Group Box 1 Protein B1 Expression. DNA Cell Biol. 2017, 36, 1018–1028. [Google Scholar] [CrossRef]
- Castoldi, G.; di Gioia, C.R.; Bombardi, C.; Catalucci, D.; Corradi, B.; Gualazzi, M.G.; Leopizzi, M.; Mancini, M.; Zerbini, G.; Condorelli, G.; et al. MiR-133a regulates collagen 1A1: Potential role of miR-133a in myocardial fibrosis in angiotensin II-dependent hypertension. J. Cell. Physiol. 2012, 227, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, B.C.; Gao, X.-M.; Winbanks, C.E.; Boey, E.J.H.; Tham, Y.K.; Kiriazis, H.; Gregorevic, P.; Obad, S.; Kauppinen, S.; Du, X.-J.; et al. Therapeutic inhibition of the miR-34 family attenuates pathological cardiac remodeling and improves heart function. Proc. Natl. Acad. Sci. USA 2012, 109, 17615–17620. [Google Scholar] [CrossRef]
- Paschou, S.A.; Stamou, M.; Vuagnat, H.; Tentolouris, N.; Jude, E. Pain management of chronic wounds: Diabetic ulcers and beyond. Maturitas 2018, 117, 17–21. [Google Scholar] [CrossRef]
- Li, X.; Jiang, C.; Zhao, J. Human endothelial progenitor cells-derived exosomes accelerate cutaneous wound healing in diabetic rats by promoting endothelial function. J. Diabetes Complicat. 2016, 30, 986–992. [Google Scholar] [CrossRef]
- Xu, J.; Bai, S.; Cao, Y.; Liu, L.; Fang, Y.; Du, J.; Luo, L.; Chen, M.; Shen, B.; Zhang, Q. miRNA-221-3p in Endothelial Progenitor Cell-Derived Exosomes Accelerates Skin Wound Healing in Diabetic Mice. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 1259–1270. [Google Scholar] [CrossRef]
- Scioli, M.G.; Storti, G.; D’Amico, F.; Guzmán, R.R.; Centofanti, F.; Doldo, E.; Miranda, E.M.C.; Orlandi, A. Oxidative Stress and New Pathogenetic Mechanisms in Endothelial Dysfunction: Potential Diagnostic Biomarkers and Therapeutic Targets. J. Clin. Med. 2020, 9, 1995. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, C.; Hu, B.; Niu, X.; Liu, X.; Zhang, G.; Zhang, C.; Li, Q.; Wang, Y. Exosomes Derived from Human Endothelial Progenitor Cells Accelerate Cutaneous Wound Healing by Promoting Angiogenesis Through Erk1/2 Signaling. Int. J. Biol. Sci. 2016, 12, 1472–1487. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Yin, Q.; Dong, T.; Dai, F.; Qin, Y.; Ye, L.; Du, J.; Zhang, Q.; Chen, H.; Shen, B. Endothelial progenitor cell–derived exosomes ameliorate endothelial dysfunction in a mouse model of diabetes. Biomed. Pharmacother. 2020, 131, 110756. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Chen, M. The significance of atypical morphology in the changes of spectrum of postinfectious glomerulonephritis. Clin. Nephrol. 2010, 73, 173–179. [Google Scholar] [CrossRef]
- Huang, K.; Li, R.; Wei, W. Sirt1 activation prevents anti-Thy 1.1 mesangial proliferative glomerulonephritis in the rat through the Nrf2/ARE pathway. Eur. J. Pharmacol. 2018, 832, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Sawai, K.; Mori, K.; Mukoyama, M.; Sugawara, A.; Suganami, T.; Koshikawa, M.; Yahata, K.; Makino, H.; Nagae, T.; Fujinaga, Y.; et al. Angiogenic Protein Cyr61 is Expressed by Podocytes in Anti-Thy-1 Glomerulonephritis. J. Am. Soc. Nephrol. 2003, 14, 1154–1163. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Oka, M.; Shirai, M.; Igarashi, Y.; Kojima, K.; Kaneko, O.; Hamada, N.; Mera, J.; Masaoka, H.; Nagase, M. Source of reactive oxygen species in anti-Thy1 nephritis. Ren. Fail. 1998, 20, 399–405. [Google Scholar] [CrossRef]
- Kunter, U.; Rong, S.; Djuric, Z.; Boor, P.; Müller-Newen, G.; Yu, D.; Floege, J. Transplanted Mesenchymal Stem Cells Accelerate Glomerular Healing in Experimental Glomerulonephritis. J. Am. Soc. Nephrol. 2006, 17, 2202–2212. [Google Scholar] [CrossRef]
- Rampino, T.; Gregorini, M.; Bedino, G.; Piotti, G.; Gabanti, E.; Ibatici, A.; Sessarego, N.; Piacenza, C.; Balenzano, C.T.; Esposito, P.; et al. Mesenchymal stromal cells improve renal injury in anti-Thy 1 nephritis by modulating inflammatory cytokines and scatter factors. Clin. Sci. 2010, 120, 25–36. [Google Scholar] [CrossRef]
- Cantaluppi, V.; Medica, D.; Mannari, C.; Stiaccini, G.; Figliolini, F.; Dellepiane, S.; Quercia, A.D.; Migliori, M.; Panichi, V.; Giovannini, L.; et al. Endothelial progenitor cell-derived extracellular vesicles protect from complement-mediated mesangial injury in experimental anti-Thy1.1 glomerulonephritis. Nephrol. Dial. Transplant. 2014, 30, 410–422. [Google Scholar] [CrossRef]
- Zarjou, A.; Agarwal, A. Sepsis and Acute Kidney Injury. J. Am. Soc. Nephrol. 2011, 22, 999–1006. [Google Scholar] [CrossRef]
- Cribbs, S.K.; Sutcliffe, D.J.; Taylor, W.R.; Rojas, M.; Easley, K.A.; Tang, L.; Brigham, K.L.; Martin, G.S. Circulating endothelial progenitor cells inversely associate with organ dysfunction in sepsis. Intensiv. Care Med. 2012, 38, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Li, P.; Goodwin, A.J.; Cook, J.A.; Halushka, P.V.; Chang, E.; Fan, H. Exosomes from Endothelial Progenitor Cells Improve the Outcome of a Murine Model of Sepsis. Mol. Ther. 2018, 26, 1375–1384. [Google Scholar] [CrossRef]
- He, Z.; Wang, H.; Yue, L. Endothelial progenitor cells-secreted extracellular vesicles containing microRNA-93-5p confer protection against sepsis-induced acute kidney injury via the KDM6B/H3K27me3/TNF-α axis. Exp. Cell Res. 2020, 395, 112173. [Google Scholar] [CrossRef] [PubMed]
- Aguado-Fraile, E.; Ramos, E.; Conde, E.; Rodríguez, M.; Martín-Gómez, L.; Lietor, A.; Candela, Á.; Ponte, B.; Liaño, F.; García-Bermejo, M.L. A Pilot Study Identifying a Set of microRNAs As Precise Diagnostic Biomarkers of Acute Kidney Injury. PLoS ONE 2015, 10, e0127175. [Google Scholar] [CrossRef] [PubMed]
- Salminen, A.; Kaarniranta, K.; Hiltunen, M.; Kauppinen, A. Histone demethylase Jumonji D3 (JMJD3/KDM6B) at the nexus of epigenetic regulation of inflammation and the aging process. J. Mol. Med. 2014, 92, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- Cantaluppi, V.; Gatti, S.; Medica, D.; Figliolini, F.; Bruno, S.; Deregibus, M.C.; Sordi, A.; Biancone, L.; Tetta, C.; Camussi, G. Microvesicles derived from endothelial progenitor cells protect the kidney from ischemia-reperfusion injury by microRNA-dependent reprogramming of resident renal cells. Kidney Int. 2012, 82, 412–427. [Google Scholar] [CrossRef] [PubMed]
- Ranghino, A.; Cantaluppi, V.; Grange, C.; Vitillo, L.; Fop, F.; Biancone, L.; Deregibus, M.; Tetta, C.; Segoloni, G.; Camussi, G. Endothelial Progenitor Cell-Derived Microvesicles Improve Neovascularization in a Murine Model of Hindlimb Ischemia. Int. J. Immunopathol. Pharmacol. 2012, 25, 75–85. [Google Scholar] [CrossRef]
- Marsell, R.; Einhorn, T.A. The biology of fracture healing. Injury 2011, 42, 551–555. [Google Scholar] [CrossRef]
- Loi, F.; Córdova, L.A.; Pajarinen, J.; Lin, T.-H.; Yao, Z.; Goodman, S.B. Inflammation, fracture and bone repair. Bone 2016, 86, 119–130. [Google Scholar] [CrossRef]
- Teitelbaum, S.L. Osteoclasts: What do they do and how do they do it? Am. J. Pathol. 2007, 170, 427–435. [Google Scholar] [CrossRef]
- Pang, H.; Wu, X.-H.; Fu, S.-L.; Luo, F.; Zhang, Z.-H.; Hou, T.-Y.; Li, Z.-Q.; Chang, Z.-Q.; Yu, B.; Xu, J.-Z. Co-culture with endothelial progenitor cells promotes survival, migration, and differentiation of osteoclast precursors. Biochem. Biophys. Res. Commun. 2013, 430, 729–734. [Google Scholar] [CrossRef] [PubMed]
- Bates, B.D.; Godbout, C.; Ramnaraign, D.J.; Schemitsch, E.; Nauth, A. Delayed Endothelial Progenitor Cell Therapy Promotes Bone Defect Repair in a Clinically Relevant Rat Model. Stem Cells Int. 2017, 2017, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Qadir, A.S.; Um, S.; Lee, H.; Baek, K.; Seo, B.M.; Lee, G.; Kim, G.-S.; Woo, K.M.; Ryoo, H.-M.; Baek, J.-H. miR-124 Negatively Regulates Osteogenic Differentiation and In vivo Bone Formation of Mesenchymal Stem Cells. J. Cell Biochem. 2015, 116, 730–742. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Weng, Y.; He, F.; Liang, D.; Cai, L. LncRNA MALAT-1 competitively regulates miR-124 to promote EMT and development of non-small-cell lung cancer. Anti-Cancer Drugs 2018, 29, 628–636. [Google Scholar] [CrossRef]
- Cui, Y.; Fu, S.; Sun, D.; Xing, J.; Hou, T.; Wu, X. EPC-derived exosomes promote osteoclastogenesis through LncRNA-MALAT1. J. Cell Mol. Med. 2019, 23, 3843–3854. [Google Scholar] [CrossRef]
- Dhahri, D.; Sato-Kusubata, K.; Ohki-Koizumi, M.; Nishida, C.; Tashiro, Y.; Munakata, S.; Shimazu, H.; Salama, Y.; Eiamboonsert, S.; Nakauchi, H.; et al. Fibrinolytic crosstalk with endothelial cells expands murine mesenchymal stromal cells. Blood 2016, 128, 1063–1075. [Google Scholar] [CrossRef]
- Qin, Y.; Zhang, C. Endothelial progenitor cell-derived extracellular vesicle-meditated cell-to-cell communication regulates the proliferation and osteoblastic differentiation of bone mesenchymal stromal cells. Mol. Med. Rep. 2017, 16, 7018–7024. [Google Scholar] [CrossRef]
- Li, W.; Zhu, S.; Hu, J. Bone Regeneration Is Promoted by Orally Administered Bovine Lactoferrin in a Rabbit Tibial Distraction Osteogenesis Model. Clin. Orthop. Relat. Res. 2015, 473, 2383–2393. [Google Scholar] [CrossRef]
- Ma, L.; Zheng, L.W.; Sham, M.H.; Cheung, L.K. Uncoupled angiogenesis and osteogenesis in nicotine-compromised bone healing. J. Bone Miner. Res. 2010, 25, 1305–1313. [Google Scholar] [CrossRef]
- Jia, Y.; Zhu, Y.; Qiu, S.; Xu, J.; Chai, Y. Exosomes secreted by endothelial progenitor cells accelerate bone regeneration during distraction osteogenesis by stimulating angiogenesis. Stem Cell Res. Ther. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Ma, X.; Wang, J.; Li, J.; Ma, C.; Chen, S.; Lei, W.; Yang, Y.; Liu, S.; Bihl, J.; Chen, C. Loading MiR-210 in Endothelial Progenitor Cells Derived Exosomes Boosts Their Beneficial Effects on Hypoxia/Reoxygeneation-Injured Human Endothelial Cells via Protecting Mitochondrial Function. Cell. Physiol. Biochem. 2018, 46, 664–675. [Google Scholar] [CrossRef]
- Persidsky, Y.; Ramirez, S.H.; Haorah, J.; Kanmogne, G.D. Blood–brain Barrier: Structural Components and Function Under Physiologic and Pathologic Conditions. J. Neuroimmune Pharmacol. 2006, 1, 223–236. [Google Scholar] [CrossRef]
- Ruck, T.; Bittner, S.; Epping, L.; Herrmann, A.M.; Meuth, S.G. Isolation of Primary Murine Brain Microvascular Endothelial Cells. J. Vis. Exp. 2014, e52204. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Shao, M.; Liu, H.; Chen, J.; Hu, J.; Zhu, L.; Liu, F.; Wang, D.; Zou, Y.; Xiong, Y.; et al. Fibroblast growth factor 21 enhances angiogenesis and wound healing of human brain microvascular endothelial cells by activating PPARγ. J. Pharmacol. Sci. 2019, 140, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Deregibus, M.C.; Cantaluppi, V.; Calogero, R.; Lo Iacono, M.; Tetta, C.; Biancone, L.; Bruno, S.; Bussolati, B.; Camussi, G. Endothelial progenitor cell–derived microvesicles activate an angiogenic program in endothelial cells by a horizontal transfer of mRNA. Blood 2007, 110, 2440–2448. [Google Scholar] [CrossRef] [PubMed]
- Zeng, W.; Lei, Q.; Ma, J.; Gao, S.; Ju, R. Endothelial Progenitor Cell-Derived Microvesicles Promote Angiogenesis in Rat Brain Microvascular Endothelial Cells In vitro. Front. Cell. Neurosci. 2021, 15, 638351. [Google Scholar] [CrossRef] [PubMed]
- Das, C.K.; Jena, B.C.; Banerjee, I.; Das, S.; Parekh, A.; Bhutia, S.K.; Mandal, M. Exosome as a Novel Shuttle for Delivery of Therapeutics across Biological Barriers. Mol. Pharm. 2019, 16, 24–40. [Google Scholar] [CrossRef] [PubMed]
- Garbuzova-Davis, S.; Kurien, C.; Haller, E.; Eve, D.J.; Navarro, S.; Steiner, G.; Mahendrasah, A.; Hailu, S.; Khatib, M.; Boccio, K.J.; et al. Human Bone Marrow Endothelial Progenitor Cell Transplantation into Symptomatic ALS Mice Delays Disease Progression and Increases Motor Neuron Survival by Repairing Blood-Spinal Cord Barrier. Sci. Rep. 2019, 9, 1–20. [Google Scholar] [CrossRef]
- Garbuzova-Davis, S.; Willing, A.E.; Ehrhart, J.; Wang, L.; Sanberg, P.R.; Borlongan, C.V. Cell-Free Extracellular Vesicles Derived from Human Bone Marrow Endothelial Progenitor Cells as Potential Therapeutics for Microvascular Endothelium Restoration in ALS. Neuromolecular Med. 2020, 22, 503–516. [Google Scholar] [CrossRef]
- Voswinckel, R.; Ziegelhoeffer, T.; Heil, M.; Kostin, S.; Breier, G.; Mehling, T.; Haberberger, R.; Clauss, M.; Gaumann, A.; Schaper, W.; et al. Circulating Vascular Progenitor Cells Do Not Contribute to Compensatory Lung Growth. Circ. Res. 2003, 93, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Liu, Z.; Hu, L.; Gu, W.; Zhu, L. Exosomes derived from endothelial progenitor cells ameliorate acute lung injury by transferring miR-126. Exp. Cell Res. 2018, 370, 13–23. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, P.; Goodwin, A.J.; Cook, J.A.; Halushka, P.V.; Chang, E.; Zingarelli, B.; Fan, H. Exosomes from endothelial progenitor cells improve outcomes of the lipopolysaccharide-induced acute lung injury. Crit. Care 2019, 23, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chong, M.; Ng, W.K.; Chan, J.K.Y. Concise Review: Endothelial Progenitor Cells in Regenerative Medicine: Applications and Challenges. STEM CELLS Transl. Med. 2016, 5, 530–538. [Google Scholar] [CrossRef] [PubMed]
- Mulcahy, L.; Pink, R.C.; Carter, D.R.F. Routes and mechanisms of extracellular vesicle uptake. J. Extracell. Vesicles 2014, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, J.; Chen, S.; Wu, P.; Xu, S.; Wang, C.; Shi, H.; Bihl, J. miR-137 boosts the neuroprotective effect of endothelial progenitor cell-derived exosomes in oxyhemoglobin-treated SH-SY5Y cells partially via COX2/PGE2 pathway. Stem Cell Res. Ther. 2020, 11, 1–18. [Google Scholar] [CrossRef]
- Smrt, R.D.; Szulwach, K.E.; Pfeiffer, R.; Li, X.; Guo, W.; Pathania, M.; Teng, Z.-Q.; Luo, Y.; Peng, J.; Bordey, A.; et al. MicroRNA miR-137 Regulates Neuronal Maturation by Targeting Ubiquitin Ligase Mind Bomb-1. Stem Cells 2010, 28, 1060–1070. [Google Scholar] [CrossRef]
- He, E.; Lozano, M.A.G.; Stringer, S.; Watanabe, K.; Sakamoto, K.; Oudsten, F.D.; Koopmans, F.; Giamberardino, S.N.; Hammerschlag, A.; Cornelisse, L.N.; et al. MIR137 schizophrenia-associated locus controls synaptic function by regulating synaptogenesis, synapse maturation and synaptic transmission. Hum. Mol. Genet. 2018, 27, 1879–1891. [Google Scholar] [CrossRef]
- Liu, X.-L.; Wang, G.; Song, W.; Yang, W.-X.; Hua, J.; Lyu, L. microRNA-137 promotes endothelial progenitor cell proliferation and angiogenesis in cerebral ischemic stroke mice by targeting NR4A2 through the Notch pathway. J. Cell. Physiol. 2018, 233, 5255–5266. [Google Scholar] [CrossRef]
- Ayer, R.; Jadhav, V.; Sugawara, T.; Zhang, J.H. The Neuroprotective Effects of Cyclooxygenase-2 Inhibition in a Mouse Model of Aneurysmal Subarachnoid Hemorrhage. Neurosurg. Med. Ethics 2011, 111, 145–149. [Google Scholar] [CrossRef]
- Wu, X.; Peng, K.; Huang, H.; Li, Z.; Xiang, W.; Deng, W.; Liu, L.; Li, W.; Zhang, T. MiR-21b-3p protects NS2OY cells against oxygen-glucose deprivation/reperfusion-induced injury by down-regulating cyclooxygenase-2. Am. J. Transl. Res. 2019, 11, 3007–3017. [Google Scholar]
- Air, E.L.; Kissela, B.M. Diabetes, the Metabolic Syndrome, and Ischemic Stroke: Epidemiology and possible mechanisms. Diabetes Care 2007, 30, 3131–3140. [Google Scholar] [CrossRef]
- Wang, J.; Chen, S.; Zhang, W.; Chen, Y.; Bihl, J.C. Exosomes from miRNA-126-modified endothelial progenitor cells alleviate brain injury and promote functional recovery after stroke. CNS Neurosci. Ther. 2020, 26, 1255–1265. [Google Scholar] [CrossRef]
- Yi, M.; Wu, Y.; Long, J.; Liu, F.; Liu, Z.; Zhang, Y.-H.; Sun, X.-P.; Fan, Z.-X.; Gao, J.; Si, J.; et al. Exosomes secreted from osteocalcin-overexpressing endothelial progenitor cells promote endothelial cell angiogenesis. Am. J. Physiol. Physiol. 2019, 317, C932–C941. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-W.; Hennessy, R.R.; Khosla, S.; Lennon, R.; Loeffler, D.; Sun, T.; Liu, Z.; Park, K.-H.; Wang, F.-L.; Lerman, L.O.; et al. Circulating osteogenic endothelial progenitor cell counts: New biomarker for the severity of coronary artery disease. Int. J. Cardiol. 2017, 227, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Zhou, M.; Lu, Z.; Li, H.; Wang, Y.; Sun, L.; Gao, M.; Wei, M.; Jia, W. Serum levels of osteocalcin are inversely associated with the metabolic syndrome and the severity of coronary artery disease in Chinese men. Clin. Endocrinol. 2011, 75, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Bonomini, M.; Giardinelli, A.; Morabito, C.; Di Silvestre, S.; Di Cesare, M.; Di Pietro, N.; Sirolli, V.; Formoso, G.; Amoroso, L.; Mariggiò, M.A.; et al. Calcimimetic R-568 and Its Enantiomer S-568 Increase Nitric Oxide Release in Human Endothelial Cells. PLoS ONE 2012, 7, e30682. [Google Scholar] [CrossRef]
- Chen, G.; Li, P.; Liu, Z.; Zeng, R.; Ma, X.; Chen, Y.; Xu, H.; Li, Z. Enrichment of miR-126 enhances the effects of endothelial progenitor cell-derived microvesicles on modulating MC3T3-E1 cell function via Erk1/2-Bcl-2 signalling pathway. Prion 2019, 13, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Kashte, S.; Gupta, M.; Rodriguez, H.C.; Gautam, S.S.; Kadam, S. Mesenchymal stem cells and exosome therapy for COVID-19: Current status and future perspective. Hum. Cell. 2020, 33, 907–918. [Google Scholar] [CrossRef]
- Nassar, W.; El-Ansary, M.; Sabry, D.; Mostafa, M.A.; Fayad, T.; Kotb, E.; Temraz, M.; Saad, A.-N.; Essa, W.; Adel, H. Umbilical cord mesenchymal stem cells derived extracellular vesicles can safely ameliorate the progression of chronic kidney diseases. Biomater. Res. 2016, 20, 21. [Google Scholar] [CrossRef]
- Adlerz, K.; Patel, D.; Rowley, J.; Ng, K.; Ahsan, T. Strategies for scalable manufacturing and translation of MSC-derived extracellular vesicles. Stem Cell Res. 2020, 48, 101978. [Google Scholar] [CrossRef] [PubMed]
- Adjei, I.M.; Blanka, S. Modulation of the Tumor Microenvironment for Cancer Treatment: A Biomaterials Approach. J. Funct. Biomater. 2015, 6, 81–103. [Google Scholar] [CrossRef]
- Ader, M.; Tanaka, E.M. Modeling human development in 3D culture. Curr. Opin. Cell Biol. 2014, 31, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Tabata, Y. Biomaterial technology for tissue engineering applications. J. R. Soc. Interface 2009, 6, S311–S324. [Google Scholar] [CrossRef] [PubMed]
- Paganini, C.; Palmiero, U.C.; Pocsfalvi, G.; Touzet, N.; Bongiovanni, A.; Arosio, P. Scalable Production and Isolation of Extracellular Vesicles: Available Sources and Lessons from Current Industrial Bioprocesses. Biotechnol. J. 2019, 14, e1800528. [Google Scholar] [CrossRef] [PubMed]
- Yan, I.K.; Shukla, N.; Borrelli, D.A.; Patel, T. Use of a Hollow Fiber Bioreactor to Collect Extracellular Vesicles from Cells in Culture. Methods Mol. Biol. 2018, 1740, 35–41. [Google Scholar] [CrossRef]
- Colao, I.; Corteling, R.; Bracewell, D.; Wall, I. Manufacturing Exosomes: A Promising Therapeutic Platform. Trends Mol. Med. 2018, 24, 242–256. [Google Scholar] [CrossRef]
- Takasugi, M. Emerging roles of extracellular vesicles in cellular senescence and aging. Aging Cell 2018, 17, e12734. [Google Scholar] [CrossRef]
- Ramasubramanian, L.; Kumar, P.; Wang, A. Engineering Extracellular Vesicles as Nanotherapeutics for Regenerative Medicine. Biomolecules 2019, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Antimisiaris, S.G.; Mourtas, S.; Marazioti, A. Exosomes and Exosome-Inspired Vesicles for Targeted Drug Delivery. Pharmaceutics 2018, 10, 218. [Google Scholar] [CrossRef]
- Man, K.; Brunet, M.Y.; Jones, M.-C.; Cox, S.C. Engineered Extracellular Vesicles: Tailored-Made Nanomaterials for Medical Applications. Nanomaterials 2020, 10, 1838. [Google Scholar] [CrossRef] [PubMed]
- Schiffelers, R.; Kooijmans, S.; Vader, P.; Dommelen, V.; Solinge, V. Exosome mimetics: A novel class of drug delivery systems. Int. J. Nanomed. 2012, 7, 1525–1541. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Li, T.-S.; Tang, J.; Cheng, K. Response by Luo et al. to Letter Regarding Article, “Fabrication of Synthetic Mesenchymal Stem Cells for the Treatment of Acute Myocardial Infarction in Mice.”. Circ. Res. 2017, 120, e48–e49. [Google Scholar] [CrossRef]
- Tang, E.; Giusti, C.; Baum, G.L.; Gu, S.; Pollock, E.; Kahn, A.E.; Roalf, D.R.; Moore, T.M.; Ruparel, K.; Gur, R.C.; et al. Developmental increases in white matter network controllability support a growing diversity of brain dynamics. Nat. Commun. 2017, 8, 1252. [Google Scholar] [CrossRef]


{kind=link}
| Pathology | Experimental Model | EV Type | EV Isolation | EV Factors | EV Modification | Effects of EVs | Ref. |
|---|---|---|---|---|---|---|---|
| Restenosis | In-stent restenosis model in rats | Exosomes | Polymer precipitation | Not specified | None | Promotion of EC repair and reduction of SMC migration and proliferation | [37] |
| Atherosclerosis | Hypertensive- hyperlipidemic hamster model | Microvesicles | Immuno precipitation | miR-10a, miR-21, miR-126 miR-146a | None | Increase in circulating late EPCs and vascular repair | [40] |
| Diabetic atherosclerotic model in mice | Exosomes | Polymer precipitation | miR-21a-5p, miR-223-3p, miR-155-5p, miR-29a-3p | None | Reduction in atherosclerotic plaques and inflammatory factors | [55] | |
| Myocardial infarction | Myocardial infarction model in rats | Not specified | Polymer precipitation | Not specified | None | Restoration of vascular structure and enhancement of hemodynamic function | [43] |
| Myocardial fibrosis | In vitro model of H/R-injured rat fibroblasts | Exosomes | Ultracentrifugation | miR-133 | None | Increase in angiogenesis and MEndoT | [44] |
| Stroke | In vitro H/R-injured human ECs | Exosomes | Ultracentrifugation | miR-210 | Loading of miR-210 | Increase in angiogenesis, improvement in mitochondrial functions, reduction of apoptosis and ROS level | [84] |
| In vitro oxyHb-injured. SH-SY5Y | Exosomes | Ultracentrifugation | miR-137 | Loading of miR-137 | Prevention of cell apoptosis and mitochondrial dysfunction | [98] | |
| Diabetic stroke model in mice | Exosomes | Immuno precipitation | miR-126 | Enrichment of miR-126 | Reduction of the infarcted area, increase in cerebral blood flow, promotion of angiogenesis and neurogenesis | [105] | |
| Wound healing | Diabetic cutaneous ulcers in mice | Exosomes | Polymer precipitation | miR-221-3p | None | Stimulation of angiogenesis and enhancing wound healing | [52] |
| Glomerulonephritis | Anti-thy1.1 glomerulonephritis model in rats; in vitro anti-thy1.1-treated rat mesangial cells | Not specified | Ultracentrifugation | Factor H, CD55, CD59 mRNA | None | Inhibition of complement-mediated injury; reduction in cell death | [62] |
| Acute kidney injury | AKI model in mice; in vitro LPS-treated HK2 | Not specified | Ultracentrifugation | miR-93-5p | None | Amelioration of multiple organ injury, inflammation and apoptosis | [66] |
| AKI model in rats | Microvesicles | Ultracentrifugation | miR-126 and miR-296 | None | Tubular cell proliferation, reduction of apoptosis and leukocyte infiltration and prevention of chronic damage | [69] | |
| Bone defects | Femur fracture model in mice; mouse BMMs in vitro | Exosomes | Ultrafiltration | Lnc- MALAT1, ITGB1 | None | Promotion of bone healing and osteoclastogenesis | [78] |
| Mouse BMSCs | Not specified | Ultracentrifugation | Not specified | None | Inhibition of osteogenesis and increase in BMSC proliferation | [80] | |
| DO model in rats; in vitro wound healing assay in HUVECs | Exosomes | Ultrafiltration | miR-126 | None | Promotion of bone regeneration and angiogenesis and cell proliferation | [83] | |
| Mouse MC3T3-E1 in vitro | Microvesicles | Ultracentrifugation | miR-126 | Enrichment of miR-126 | Reduction of apoptosis and increase in proliferation and migration | [110] | |
| Brain damage | Rats BMECs in vitro | Microvesicles | Ultracentrifugation | miR-210 | None | Promotion of proliferation, migration and tube formation | [89] |
| Amyotrophic lateral sclerosis | ALS plasma-treated mBECs | Exosomes | Polymer precipitation | Not specified | None | Prevention of cell death | [92] |
| Acute lung injury | ALI model in rats; in vitro LPS-injured HUVECs | Exosomes | Ultracentrifugation | miR-126 | None | Reduction of interstitial edema, alveolar wall thickness and inflammatory cell number; induction of cell proliferation, migration and tube formation | [94] |
| Lung injury model in rats; human SAECs in vitro | Exosomes | Polymer precipitation | miR-126 | None | Reduction of inflammation and permeability | [95] | |
| Endothelial dysfunction | RAOECs | Exosomes | Polymer precipitation | OCN | OCN overexpression | Stimulation of angiogenesis | [106] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terriaca, S.; Fiorelli, E.; Scioli, M.G.; Fabbri, G.; Storti, G.; Cervelli, V.; Orlandi, A. Endothelial Progenitor Cell-Derived Extracellular Vesicles: Potential Therapeutic Application in Tissue Repair and Regeneration. Int. J. Mol. Sci. 2021, 22, 6375. https://doi.org/10.3390/ijms22126375
Terriaca S, Fiorelli E, Scioli MG, Fabbri G, Storti G, Cervelli V, Orlandi A. Endothelial Progenitor Cell-Derived Extracellular Vesicles: Potential Therapeutic Application in Tissue Repair and Regeneration. International Journal of Molecular Sciences. 2021; 22(12):6375. https://doi.org/10.3390/ijms22126375
Chicago/Turabian StyleTerriaca, Sonia, Elena Fiorelli, Maria Giovanna Scioli, Giulia Fabbri, Gabriele Storti, Valerio Cervelli, and Augusto Orlandi. 2021. "Endothelial Progenitor Cell-Derived Extracellular Vesicles: Potential Therapeutic Application in Tissue Repair and Regeneration" International Journal of Molecular Sciences 22, no. 12: 6375. https://doi.org/10.3390/ijms22126375
APA StyleTerriaca, S., Fiorelli, E., Scioli, M. G., Fabbri, G., Storti, G., Cervelli, V., & Orlandi, A. (2021). Endothelial Progenitor Cell-Derived Extracellular Vesicles: Potential Therapeutic Application in Tissue Repair and Regeneration. International Journal of Molecular Sciences, 22(12), 6375. https://doi.org/10.3390/ijms22126375