Overexpression of ABCB1 Transporter Confers Resistance to mTOR Inhibitor WYE-354 in Cancer Cells


 ,
, 
Abstract
1. Introduction
2. Results
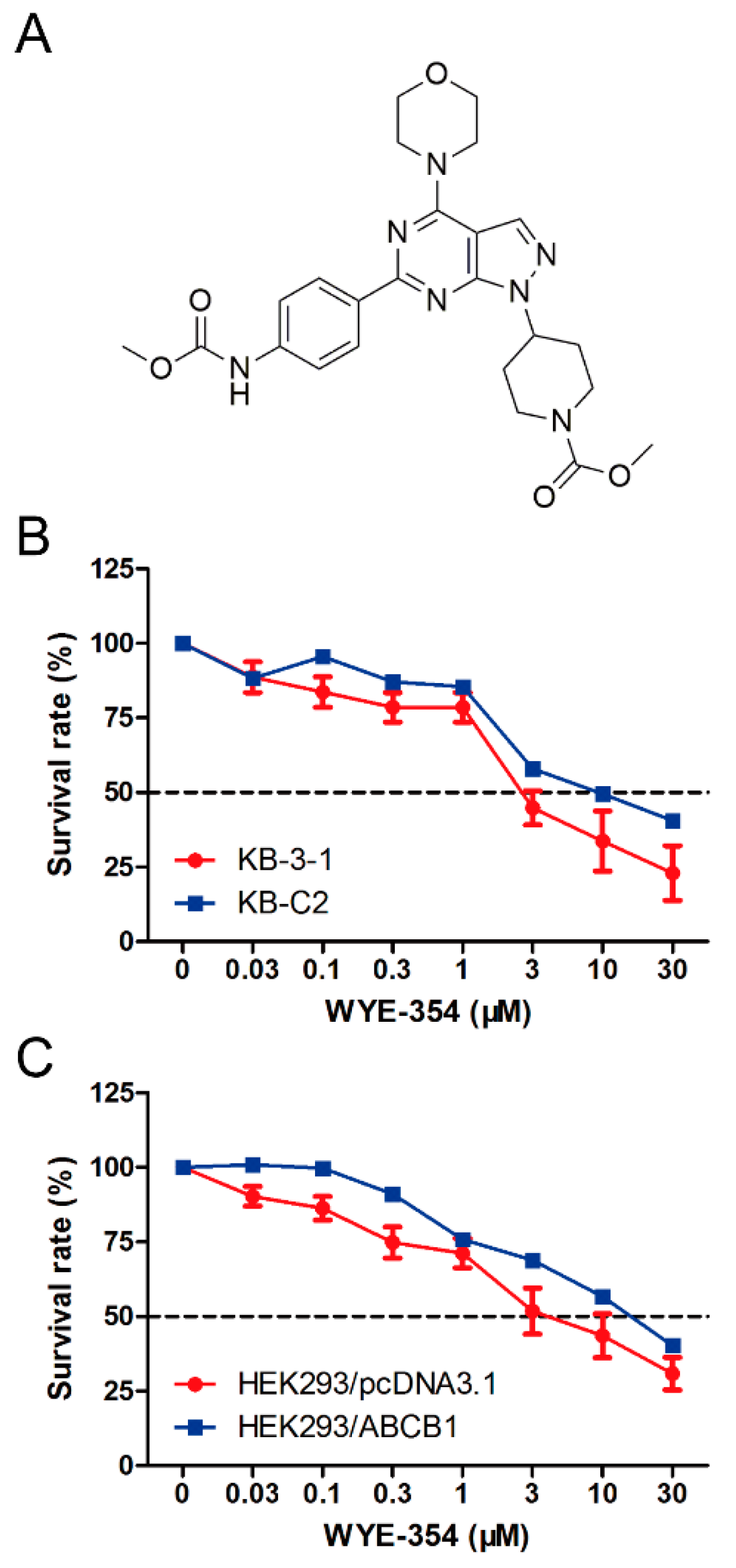
2.1. The Survival Rate of ABCB1-Overexpressing Cells Treated with WYE-354 Was Higher than Those of Sensitive Cells
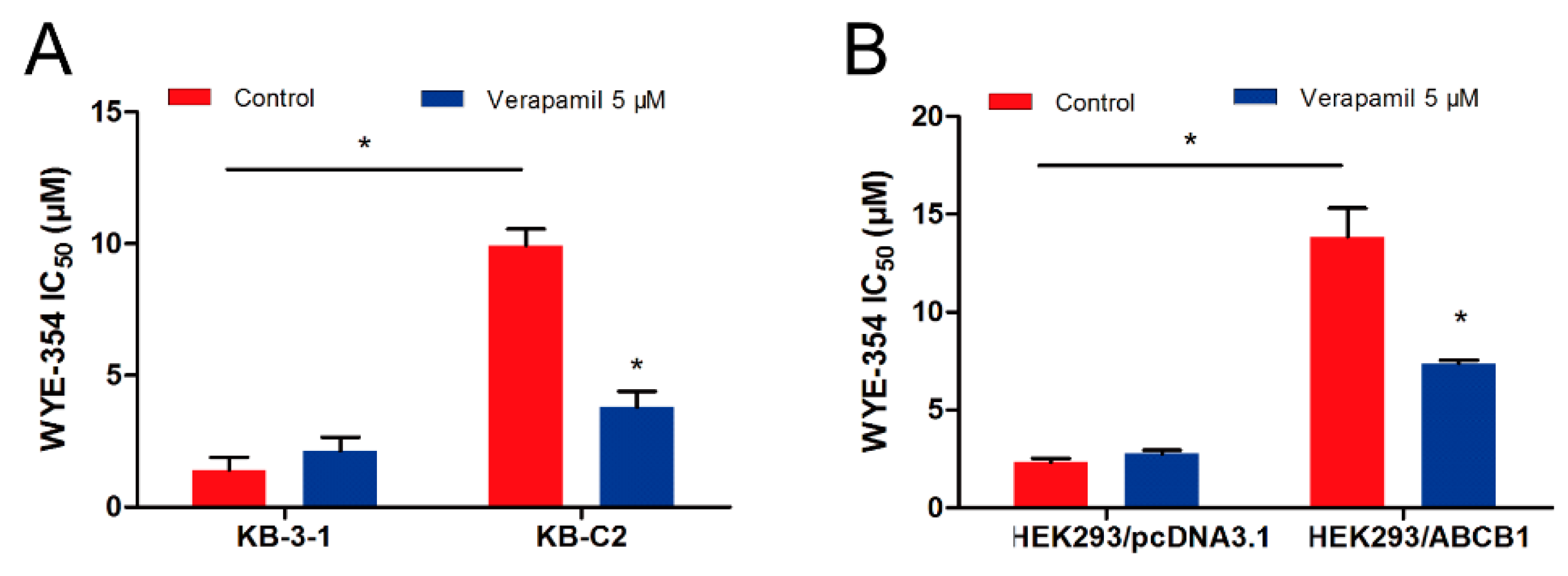
2.2. Verapamil, an ABCB1 Inhibitor, Sensitized the Efficacy of WYE-354 in ABCB1-Mediated MDR Cells
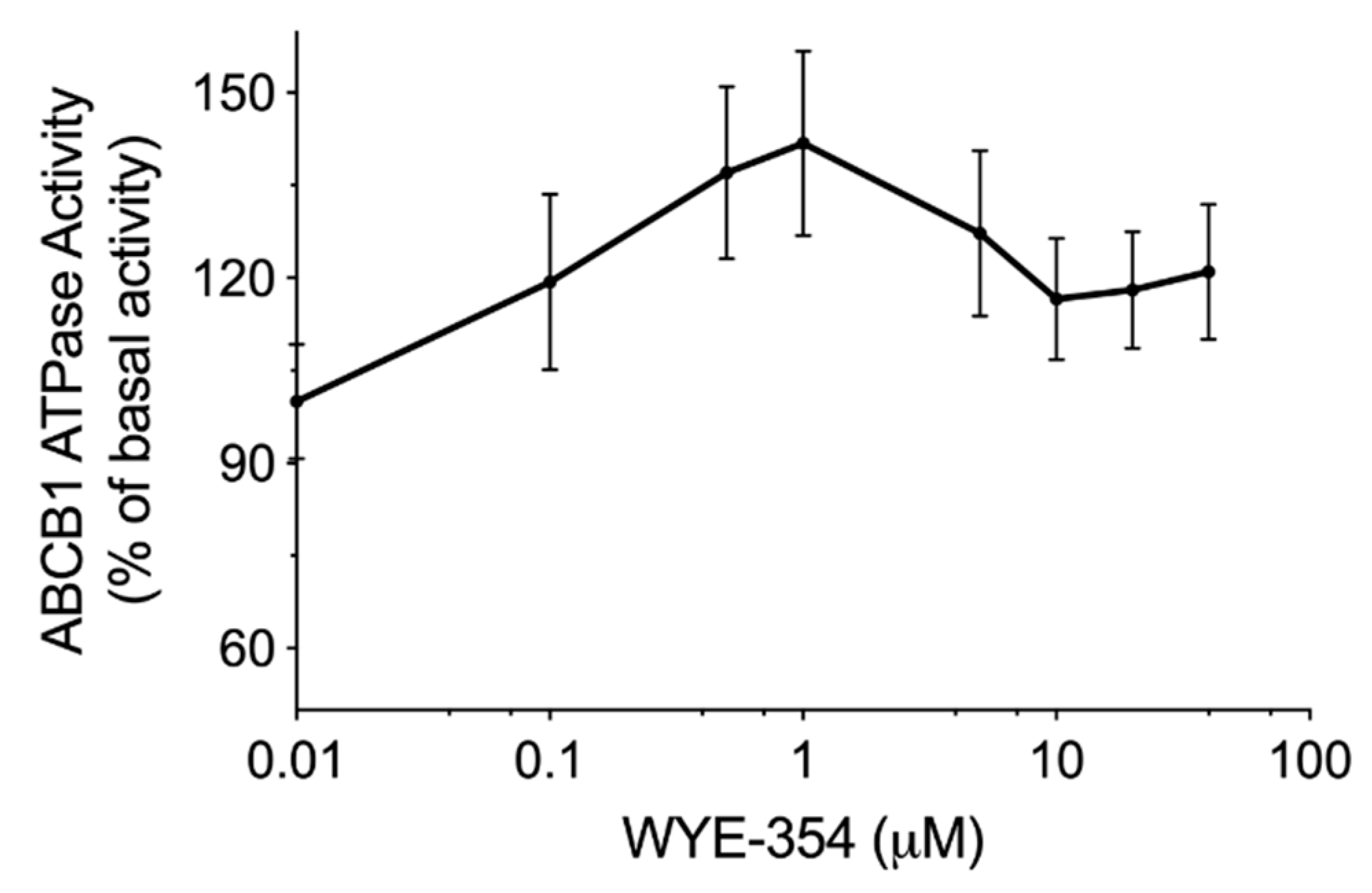
2.3. WYE-354 Stimulated ABCB1 ATPase Activity
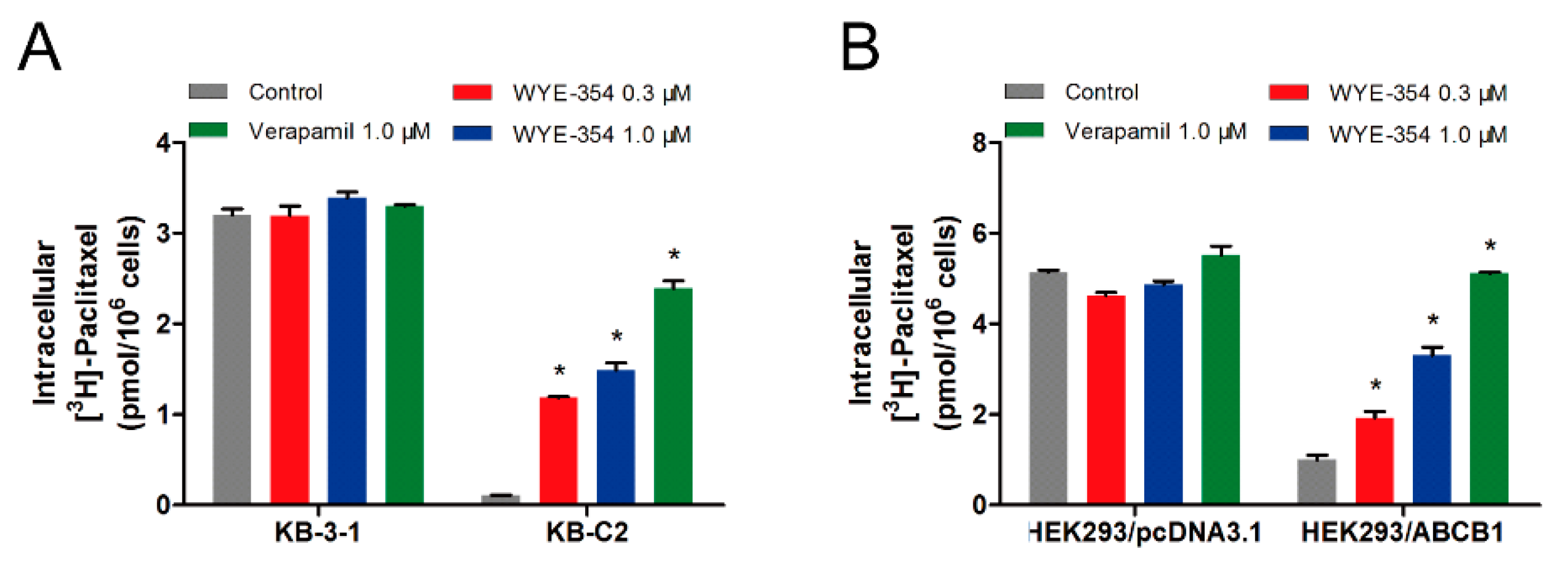
2.4. WYE-354 Increased the ABCB1-Mediated Transport of [3H]-Paclitaxel
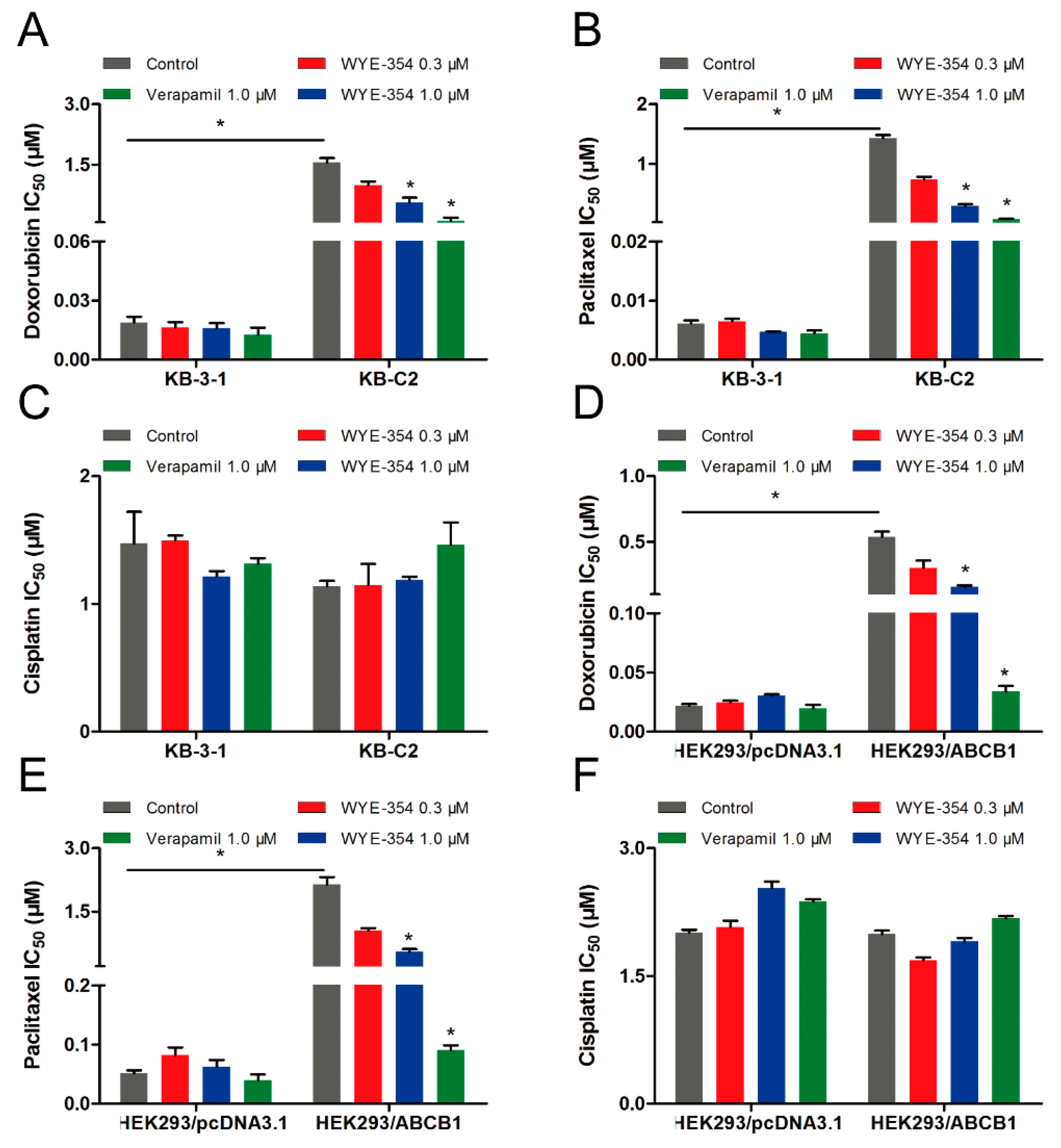
2.5. Substrate-Drugs Co-Treated with WYE-354 Decreased the Survival Rates of ABCB1-Medified MDR Cells
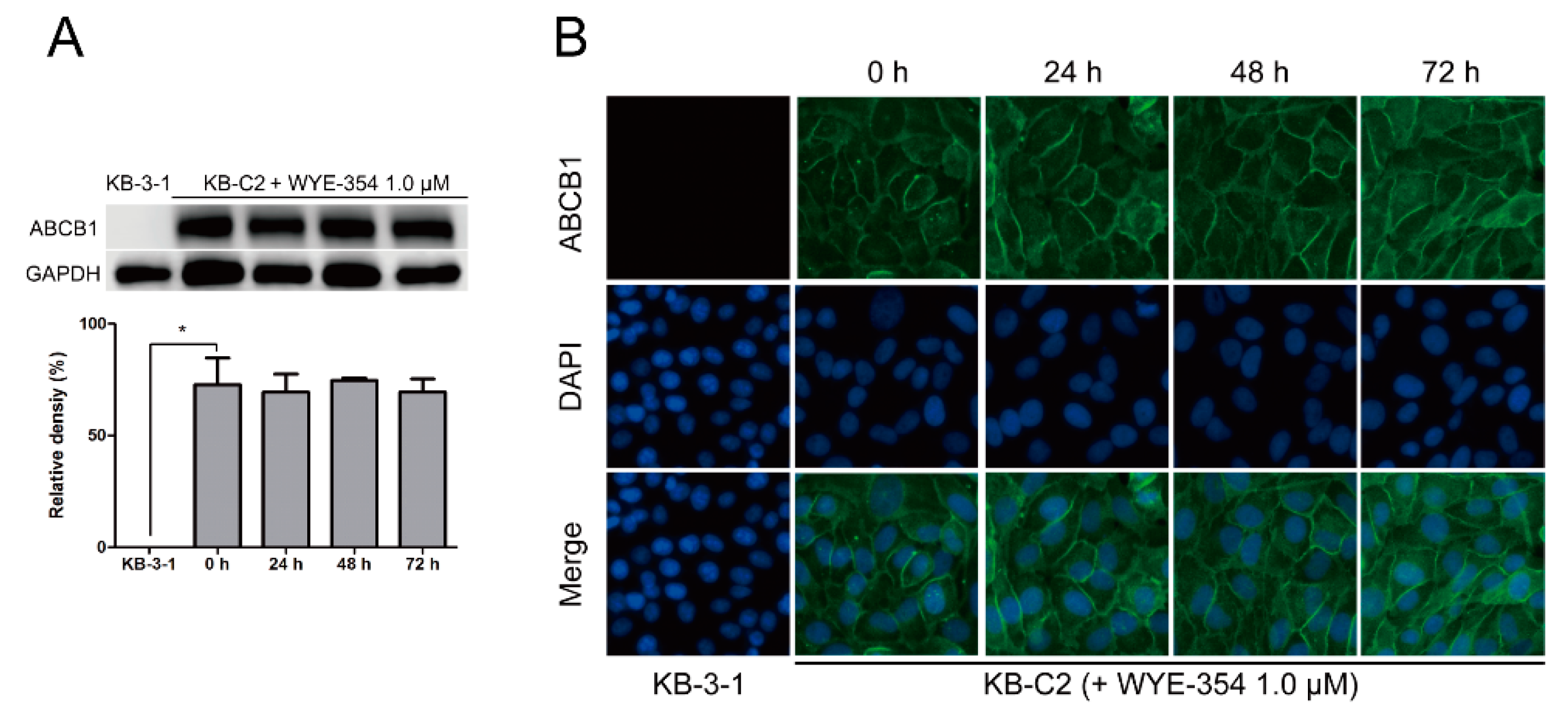
2.6. WYE-354 Did Not Affect the Protein Expression or Subcellular Localization of ABCB1
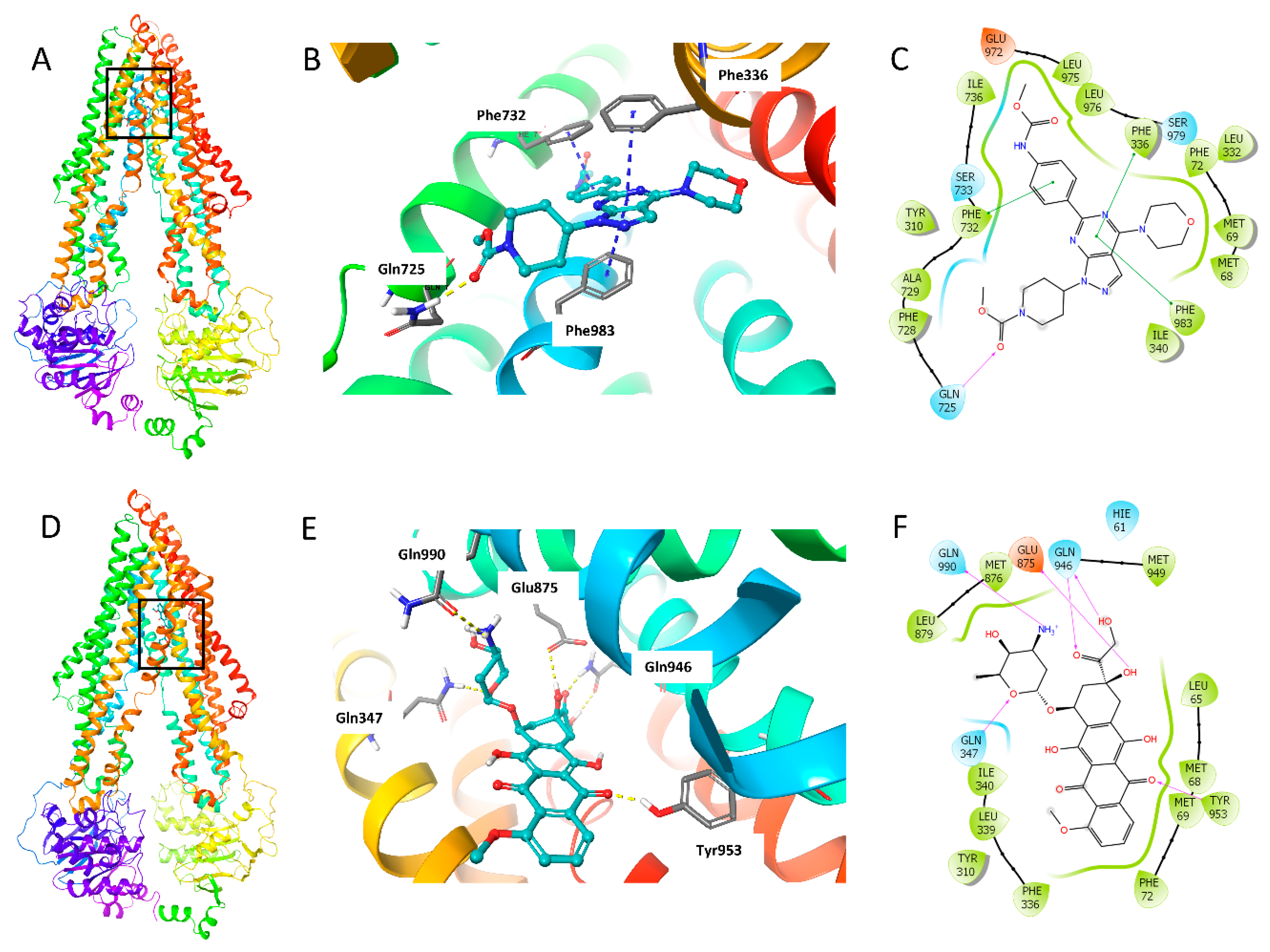
2.7. Molecular Docking Analysis of WYE-354 with Human ABCB1 Model
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Lines and Cell Culture
4.3. Cell Viability Assay
4.4. [3H]-Paclitaxel Accumulation Assay
4.5. ABCB1 ATPase Assay
4.6. Western Blotting Analysis
4.7. Immunofluorescence Assay
4.8. Molecular Modeling of Human ABCB1 Model
4.9. Statistics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Amawi, H.; Sim, H.-M.; Tiwari, A.K.; Ambudkar, S.V.; Shukla, S. ABC Transporter-Mediated Multidrug-Resistant Cancer. In Drug Transporters in Drug Disposition, Effects and Toxicity; Liu, X., Pan, G., Eds.; Springer: Singapore, 2019; pp. 549–580. [Google Scholar] [CrossRef]
- Kumar, A.; Jaitak, V. Natural products as multidrug resistance modulators in cancer. Eur. J. Med. Chem. 2019, 176, 268–291. [Google Scholar] [CrossRef] [PubMed]
- Malmström, A.; Łysiak, M.; Åkesson, L.; Jakobsen, I.; Mudaisi, M.; Milos, P.; Hallbeck, M.; Fomichov, V.; Broholm, H.; Grunnet, K.; et al. ABCB1 single-nucleotide variants and survival in patients with glioblastoma treated with radiotherapy concomitant with temozolomide. Pharm. J. 2019. [Google Scholar] [CrossRef] [PubMed]
- Al-Ali, A.A.A.; Nielsen, R.B.; Steffansen, B.; Holm, R.; Nielsen, C.U. Nonionic surfactants modulate the transport activity of ATP-binding cassette (ABC) transporters and solute carriers (SLC): Relevance to oral drug absorption. Int. J. Pharm. 2019, 566, 410–433. [Google Scholar] [CrossRef] [PubMed]
- Sinha, B.K.; Perera, L.; Cannon, R.E. Reversal of drug resistance by JS-K and nitric oxide in ABCB1- and ABCG2-expressing multi-drug resistant human tumor cells. Biomed. Pharmacother. 2019, 120, 109468. [Google Scholar] [CrossRef]
- Sachs, J.; Döhl, K.; Weber, A.; Bonus, M.; Ehlers, F.; Fleischer, E.; Klinger, A.; Gohlke, H.; Pietruszka, J.; Schmitt, L.; et al. Novel 3,4-Dihydroisocoumarins Inhibit Human P-gp and BCRP in Multidrug Resistant Tumors and Demonstrate Substrate Inhibition of Yeast Pdr5. Front. Pharmacol. 2019, 10. [Google Scholar] [CrossRef]
- Kayouka, M.; Hamade, A.; Saliba, E.; Najjar, F.; Landy, D.; Greige-Gerges, H. P-glycoprotein modulates oleanolic acid effects in hepatocytes cancer cells and zebrafish embryos. Chem. Biol. Interact. 2020, 315, 108892. [Google Scholar] [CrossRef]
- Dönmez, Y.; Akhmetova, L.; İşeri, Ö.D.; Kars, M.D.; Gündüz, U. Effect of MDR modulators verapamil and promethazine on gene expression levels of MDR1 and MRP1 in doxorubicin-resistant MCF-7 cells. Cancer Chemother. Pharmacol. 2011, 67, 823–828. [Google Scholar] [CrossRef]
- Larrivée, B.; Averill, D.A. Modulation of Adriamycin cytotoxicity and transport in drug-sensitive and multidrug-resistant Chinese hamster ovary cells by hyperthermia and cyclosporin A. Cancer Chemother. Pharmacol. 2000, 45, 219–230. [Google Scholar] [CrossRef]
- Han, W.; Shi, L.; Ren, L.; Zhou, L.; Li, T.; Qiao, Y.; Wang, H. A nanomedicine approach enables co-delivery of cyclosporin A and gefitinib to potentiate the therapeutic efficacy in drug-resistant lung cancer. Signal Transduct. Target. Ther. 2018, 3, 16. [Google Scholar] [CrossRef]
- Bhat, U.G.; Winter, M.A.; Pearce, H.L.; Beck, W.T. A structure-function relationship among reserpine and yohimbine analogues in their ability to increase expression of mdr1 and P-glycoprotein in a human colon carcinoma cell line. Mol. Pharmacol. 1995, 48, 682–689. [Google Scholar]
- Shaheen, A.; Afridi, W.A.; Mahboob, S.; Sana, M.; Zeeshan, N.; Ismat, F.; Mirza, O.; Iqbal, M.; Rahman, M. Reserpine Is the New Addition into the Repertoire of AcrB Efflux Pump Inhibitors. Mol. Biol. 2019, 53, 596–605. [Google Scholar] [CrossRef]
- Dastvan, R.; Mishra, S.; Peskova, Y.B.; Nakamoto, R.K.; Mchaourab, H.S. Mechanism of allosteric modulation of P-glycoprotein by transport substrates and inhibitors. Science 2019, 364, 689–692. [Google Scholar] [CrossRef]
- Ledwitch, K.V.; Gibbs, M.E.; Barnes, R.W.; Roberts, A.G. Cooperativity between verapamil and ATP bound to the efflux transporter P-glycoprotein. Biochem. Pharmacol. 2016, 118, 96–108. [Google Scholar] [CrossRef]
- Aygül, A. The importance of efflux systems in antibiotic resistance and efflux pump inhibitors in the management of resistance. Mikrobiyoloji Bul. 2015, 49, 278–291. [Google Scholar] [CrossRef]
- Karar, J.; Maity, A. PI3K/AKT/mTOR Pathway in Angiogenesis. Front. Mol. Neurosci. 2011, 4. [Google Scholar] [CrossRef]
- Porta, C.; Paglino, C.; Mosca, A. Targeting PI3K/Akt/mTOR Signaling in Cancer. Front. Oncol. 2014, 4. [Google Scholar] [CrossRef]
- Lapointe, S.; Mason, W.; MacNeil, M.; Harlos, C.; Tsang, R.; Sederias, J.; Luchman, H.A.; Weiss, S.; Rossiter, J.P.; Tu, D.; et al. A phase I study of vistusertib (dual mTORC1/2 inhibitor) in patients with previously treated glioblastoma multiforme: A CCTG study. Investig. New Drugs 2019. [Google Scholar] [CrossRef]
- Counts, B.R.; Hardee, J.P.; Fix, D.K.; VanderVeen, B.N.; Montalvo, R.N.; Carson, J.A. Cachexia Disrupts Diurnal Regulation of Activity, Feeding, and Muscle mTORC1 in Mice. Med. Sci. Sports Exerc. 2019. [Google Scholar] [CrossRef]
- Sarma, P.; Ennis, K.; Plas, D.R. CSIG-10. INVESTIGATING THE S6K1 AND S6K2 IN PTEN-DEFICIENT GLIOBLASTOMA. Neuro Oncol. 2018, 20, vi45. [Google Scholar] [CrossRef][Green Version]
- Fang, Y.; Vilella-Bach, M.; Bachmann, R.; Flanigan, A.; Chen, J. Phosphatidic Acid-Mediated Mitogenic Activation of mTOR Signaling. Science 2001, 294, 1942–1945. [Google Scholar] [CrossRef]
- Zou, Z.; Zhang, J.; Zhang, H.; Liu, H.; Li, Z.; Cheng, D.; Chen, J.; Liu, L.; Ni, M.; Zhang, Y.; et al. 3-Methyladenine can depress drug efflux transporters via blocking the PI3K–AKT–mTOR pathway thus sensitizing MDR cancer to chemotherapy. J. Drug Target. 2014, 22, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Scherbakova, E.A.; Rybalkina, E.Y.; Stromskaya, T.P.; Stavrovskaya, A.A. Participation of mTOR in the regulation of multidrug resistance of tumor cells. Biochem. (Mosc.) Suppl. Ser. A Membr. Cell Biol. 2009, 3, 184–189. [Google Scholar] [CrossRef]
- Weber, H.; Leal, P.; Stein, S.; Kunkel, H.; García, P.; Bizama, C.; Espinoza, J.A.; Riquelme, I.; Nervi, B.; Araya, J.C.; et al. Rapamycin and WYE-354 suppress human gallbladder cancer xenografts in mice. Oncotarget 2015, 6, 31877–31888. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Toral-Barza, L.; Shi, C.; Zhang, W.-G.; Lucas, J.; Shor, B.; Kim, J.; Verheijen, J.; Curran, K.; Malwitz, D.J.; et al. Biochemical, Cellular, and In vivo Activity of Novel ATP-Competitive and Selective Inhibitors of the Mammalian Target of Rapamycin. Cancer Res. 2009, 69, 6232–6240. [Google Scholar] [CrossRef]
- Zhou, H.; Luo, Y.; Huang, S. Updates of mTOR Inhibitors. Anti-Cancer Agents Med. Chem. (Former. Curr. Med. Chem. Anti Cancer Agents) 2010, 10, 571–581. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, Y.-R.; Wang, S.; Zhao, S. Autophagy inhibition sensitizes WYE-354-induced anti-colon cancer activity in vitro and in vivo. Tumor Biol. 2016, 37, 11743–11752. [Google Scholar] [CrossRef]
- Sami, A.; Karsy, M. Targeting the PI3K/AKT/mTOR signaling pathway in glioblastoma: Novel therapeutic agents and advances in understanding. Tumor Biol. 2013, 34, 1991–2002. [Google Scholar] [CrossRef]
- Ibrahim, S.M.; Bakhashab, S.; Ilyas, A.M.; Pushparaj, P.N.; Karim, S.; Khan, J.A.; Abuzenadah, A.M.; Chaudhary, A.G.; Al-Qahtani, M.H.; Ahmed, F. WYE-354 restores Adriamycin sensitivity in multidrug-resistant acute myeloid leukemia cell lines. Oncol. Rep. 2019, 41, 3179–3188. [Google Scholar] [CrossRef]
- Sharom, F.J. Shedding light on drug transport: Structure and function of the P-glycoprotein multidrug transporter (ABCB1)This paper is one of a selection of papers published in this Special Issue, entitled CSBMCB—Membrane Proteins in Health and Disease. Biochem. Cell Biol. 2006, 84, 979–992. [Google Scholar] [CrossRef]
- Broccatelli, F.; Carosati, E.; Neri, A.; Frosini, M.; Goracci, L.; Oprea, T.I.; Cruciani, G. A Novel Approach for Predicting P-Glycoprotein (ABCB1) Inhibition Using Molecular Interaction Fields. J. Med. Chem. 2011, 54, 1740–1751. [Google Scholar] [CrossRef]
- Dhaliwal, A.K.; Mohan, A.; Gill, K.S. Comparative analysis of ABCB1 reveals novel structural and functional conservation between monocots and dicots. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Ravna, A.W.; Sylte, I.; Sager, G. Molecular model of the outward facing state of the human P-glycoprotein (ABCB1), and comparison to a model of the human MRP5 (ABCC5). Theor. Biol. Med Model. 2007, 4, 33. [Google Scholar] [CrossRef] [PubMed]
- Sauna, Z.E.; Kim, I.-W.; Ambudkar, S.V. Genomics and the mechanism of P-glycoprotein (ABCB1). J. Bioenerg. Biomembr. 2007, 39, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Ji, N.; Yang, Y.; Cai, C.-Y.; Wang, J.-Q.; Lei, Z.-N.; Wu, Z.-X.; Cui, Q.; Yang, D.-H.; Chen, Z.-S.; Kong, D. Midostaurin Reverses ABCB1-Mediated Multidrug Resistance, an in vitro Study. Front. Oncol. 2019, 9, 514. [Google Scholar] [CrossRef]
- Sauna, Z.E.; Ambudkar, S.V. About a switch: How P-glycoprotein (ABCB1) harnesses the energy of ATP binding and hydrolysis to do mechanical work. Mol. Cancer Ther. 2007, 6, 13–23. [Google Scholar] [CrossRef]
- Chufan, E.E.; Sim, H.-M.; Ambudkar, S.V. Chapter Three-Molecular Basis of the Polyspecificity of P-Glycoprotein (ABCB1): Recent Biochemical and Structural Studies. In Advances in Cancer Research; Schuetz, J.D., Ishikawa, T., Eds.; Academic Press: Cambridge, MA, USA, 2015; Volume 125, pp. 71–96. [Google Scholar]
- Kim, R.B. Drugs as P-glycoprotein substrates, inhibitors, and inducers. Drug Metab. Rev. 2002, 34, 47–54. [Google Scholar] [CrossRef]
- Hu, T.; To, K.K.W.; Wang, L.; Zhang, L.; Lu, L.; Shen, J.; Chan, R.L.Y.; Li, M.; Yeung, J.H.K.; Cho, C.H. Reversal of P-glycoprotein (P-gp) mediated multidrug resistance in colon cancer cells by cryptotanshinone and dihydrotanshinone of Salvia miltiorrhiza. Phytomedicine 2014, 21, 1264–1272. [Google Scholar] [CrossRef]
- Summers, M.A.; Moore, J.L.; McAuley, J.W. Use of Verapamil as a Potential P-Glycoprotein Inhibitor in a Patient with Refractory Epilepsy. Ann. Pharmacother. 2004, 38, 1631–1634. [Google Scholar] [CrossRef]
- Tuijnenburg, P.; aan de Kerk, D.J.; Jansen, M.H.; Morris, B.; Lieftink, C.; Beijersbergen, R.L.; van Leeuwen, E.M.; Kuijpers, T.W. High-throughput compound screen reveals mTOR inhibitors as potential therapeutics to reduce (auto) antibody production by human plasma cells. Eur. J. Immunol. 2019, 50, 73–85. [Google Scholar] [CrossRef]
- Levy, E.S.; Samy, K.E.; Lamson, N.G.; Whitehead, K.A.; Kroetz, D.L.; Desai, T.A. Reversible inhibition of efflux transporters by hydrogel microdevices. Eur. J. Pharm. Biopharm. 2019, 145, 76–84. [Google Scholar] [CrossRef]
- Lyall, R.M.; Hwang, J.; Cardarelli, C.; FitzGerald, D.; Akiyama, S.-I.; Gottesman, M.M.; Pastan, I. Isolation of Human KB Cell Lines Resistant to Epidermal Growth Factor-Pseudomonas Exotoxin Conjugates. Cancer Res. 1987, 47, 2961–2966. [Google Scholar] [PubMed]
- Fung, K.L.; Pan, J.; Ohnuma, S.; Lund, P.E.; Pixley, J.N.; Kimchi-Sarfaty, C.; Ambudkar, S.V.; Gottesman, M.M. MDR1 Synonymous Polymorphisms Alter Transporter Specificity and Protein Stability in a Stable Epithelial Monolayer. Cancer Res. 2014, 74, 598–608. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, S.A.; Reiisi, S.; Ghiasi Tabari, P.; Shekari, A.; Aliakbari, F.; Azadfallah, E.; Elahian, F. Broad blocking of MDR efflux pumps by acetylshikonin and acetoxyisovalerylshikonin to generate hypersensitive phenotype of malignant carcinoma cells. Sci. Rep. 2018, 8, 3446. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-Y.; Zhang, Y.-K.; Wang, Y.-J.; Gupta, P.; Zeng, L.; Xu, M.; Wang, X.-Q.; Yang, D.-H.; Chen, Z.-S. Osimertinib (AZD9291), a Mutant-Selective EGFR Inhibitor, Reverses ABCB1-Mediated Drug Resistance in Cancer Cells. Molecules 2016, 21, 1236. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-K.; Zhang, G.-N.; Wang, Y.-J.; Patel, B.A.; Talele, T.T.; Yang, D.-H.; Chen, Z.-S. Bafetinib (INNO-406) reverses multidrug resistance by inhibiting the efflux function of ABCB1 and ABCG2 transporters. Sci. Rep. 2016, 6, 25694. [Google Scholar] [CrossRef] [PubMed]
- Alam, A.; Kowal, J.; Broude, E.; Roninson, I.; Locher, K.P. Structural insight into substrate and inhibitor discrimination by human P-glycoprotein. Science 2019, 363, 753–756. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | IC50 ± SD a (RF b) (μM) | |
|---|---|---|
| WYE-354 | WYE-354 + Verapamil 5 µM | |
| KB-3-1 | 1.410 ± 0.481 (1.00) | 2.135 ± 0.522 (1.51) |
| KB-C2 | 9.937 ± 0.625 (7.05) | 3.788 ± 0.604 (2.69) * |
| HEK293/pcDNA3.1 | 2.319 ± 0.208 (1.00) | 2.764 ± 0.179 (1.19) |
| HEK293/ABCB1 | 13.84 ± 1.465 (5.97) | 7.372 ± 0.165 (3.18) * |
| Treatment | IC50 ± SD a (RF b) (μM) | |
|---|---|---|
| KB-3-1 | KB-C2 | |
| Doxorubicin | 0.019 ± 0.004 (1.00) | 1.549 ± 0.158 (81.52) |
| +WYE-354 (0.3 µM) | 0.016 ± 0.004 (0.84) | 0.983 ± 0.140 (51.74) * |
| +WYE-354 (1 µM) | 0.016 ± 0.004 (0.84) | 0.563 ± 0.164 (29.63) * |
| +Verapamil (1 µM) | 0.013 ± 0.005 (0.68) | 0.095 ± 0.012 (5.00) * |
| Paclitaxel | 0.006 ± 0.001 (1.00) | 1.432 ± 0.075 (238.67) |
| +WYE-354 (0.3 µM) | 0.006 ± 0.002 (1.00) | 0.736 ± 0.069 (122.67) * |
| +WYE-354 (1 µM) | 0.005 ± 0.001 (0.83) | 0.296 ± 0.041 (49.33) * |
| +Verapamil (1 µM) | 0.004 ± 0.001 (0.67) | 0.075 ± 0.010 (12.50) * |
| Cisplatin | 1.472 ± 0.351 (1.00) | 1.139 ± 0.060 (0.77) |
| +WYE-354 (0.3 µM) | 1.498 ± 0.054 (1.02) | 1.148 ± 0.232 (0.78) |
| +WYE-354 (1 µM) | 1.214 ± 0.061 (0.82) | 1.187 ± 0.037 (0.81) |
| +Verapamil (1 µM) | 1.314 ± 0.063 (0.89) | 1.462 ± 0.306 (1.02) |
| Treatment | IC50 ± SD a (RF b) (μM) | |
|---|---|---|
| HEK293/pcDNA3.1 | HEK293/ABCB1 | |
| Doxorubicin | 0.022 ± 0.002 (1.00) | 0.539 ± 0.054 (24.50) |
| +WYE-354 (0.3 µM) | 0.023 ± 0.002 (1.04) | 0.302 ± 0.077 (13.73) * |
| +WYE-354 (1 µM) | 0.030 ± 0.001 (1.36) | 0.157 ± 0.014 (7.14) * |
| +Verapamil (1 µM) | 0.020 ± 0.004 (0.91) | 0.034 ± 0.006 (1.54) * |
| Paclitaxel | 0.051 ± 0.007 (1.00) | 2.144 ± 0.245 (42.04) |
| +WYE-354 (0.3 µM) | 0.082 ± 0.019 (1.61) | 1.048 ± 0.087 (20.55) * |
| +WYE-354 (1 µM) | 0.063 ± 0.016 (1.23) | 0.549 ± 0.087 (10.76) * |
| +Verapamil (1 µM) | 0.040 ± 0.013 (0.78) | 0.091 ± 0.011 (1.78) * |
| Cisplatin | 2.008 ± 0.055 (1.00) | 1.990 ± 0.063 (0.99) |
| +WYE-354 (0.3 µM) | 2.075 ± 0.106 (1.03) | 1.686 ± 0.046 (0.84) |
| +WYE-354 (1 µM) | 2.531 ± 0.111 (1.26) | 1.907 ± 0.056 (0.95) |
| +Verapamil (1 µM) | 2.375 ± 0.353 (1.18) | 2.179 ± 0.037 (1.08) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Yang, D.-H.; Yang, Y.; Wang, J.-Q.; Cai, C.-Y.; Lei, Z.-N.; Teng, Q.-X.; Wu, Z.-X.; Zhao, L.; Chen, Z.-S. Overexpression of ABCB1 Transporter Confers Resistance to mTOR Inhibitor WYE-354 in Cancer Cells. Int. J. Mol. Sci. 2020, 21, 1387. https://doi.org/10.3390/ijms21041387
Wang J, Yang D-H, Yang Y, Wang J-Q, Cai C-Y, Lei Z-N, Teng Q-X, Wu Z-X, Zhao L, Chen Z-S. Overexpression of ABCB1 Transporter Confers Resistance to mTOR Inhibitor WYE-354 in Cancer Cells. International Journal of Molecular Sciences. 2020; 21(4):1387. https://doi.org/10.3390/ijms21041387
Chicago/Turabian StyleWang, Jingqiu, Dong-Hua Yang, Yuqi Yang, Jing-Quan Wang, Chao-Yun Cai, Zi-Ning Lei, Qiu-Xu Teng, Zhuo-Xun Wu, Linguo Zhao, and Zhe-Sheng Chen. 2020. "Overexpression of ABCB1 Transporter Confers Resistance to mTOR Inhibitor WYE-354 in Cancer Cells" International Journal of Molecular Sciences 21, no. 4: 1387. https://doi.org/10.3390/ijms21041387
APA StyleWang, J., Yang, D.-H., Yang, Y., Wang, J.-Q., Cai, C.-Y., Lei, Z.-N., Teng, Q.-X., Wu, Z.-X., Zhao, L., & Chen, Z.-S. (2020). Overexpression of ABCB1 Transporter Confers Resistance to mTOR Inhibitor WYE-354 in Cancer Cells. International Journal of Molecular Sciences, 21(4), 1387. https://doi.org/10.3390/ijms21041387