The Flagellar Gene Regulates Biofilm Formation and Mussel Larval Settlement and Metamorphosis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
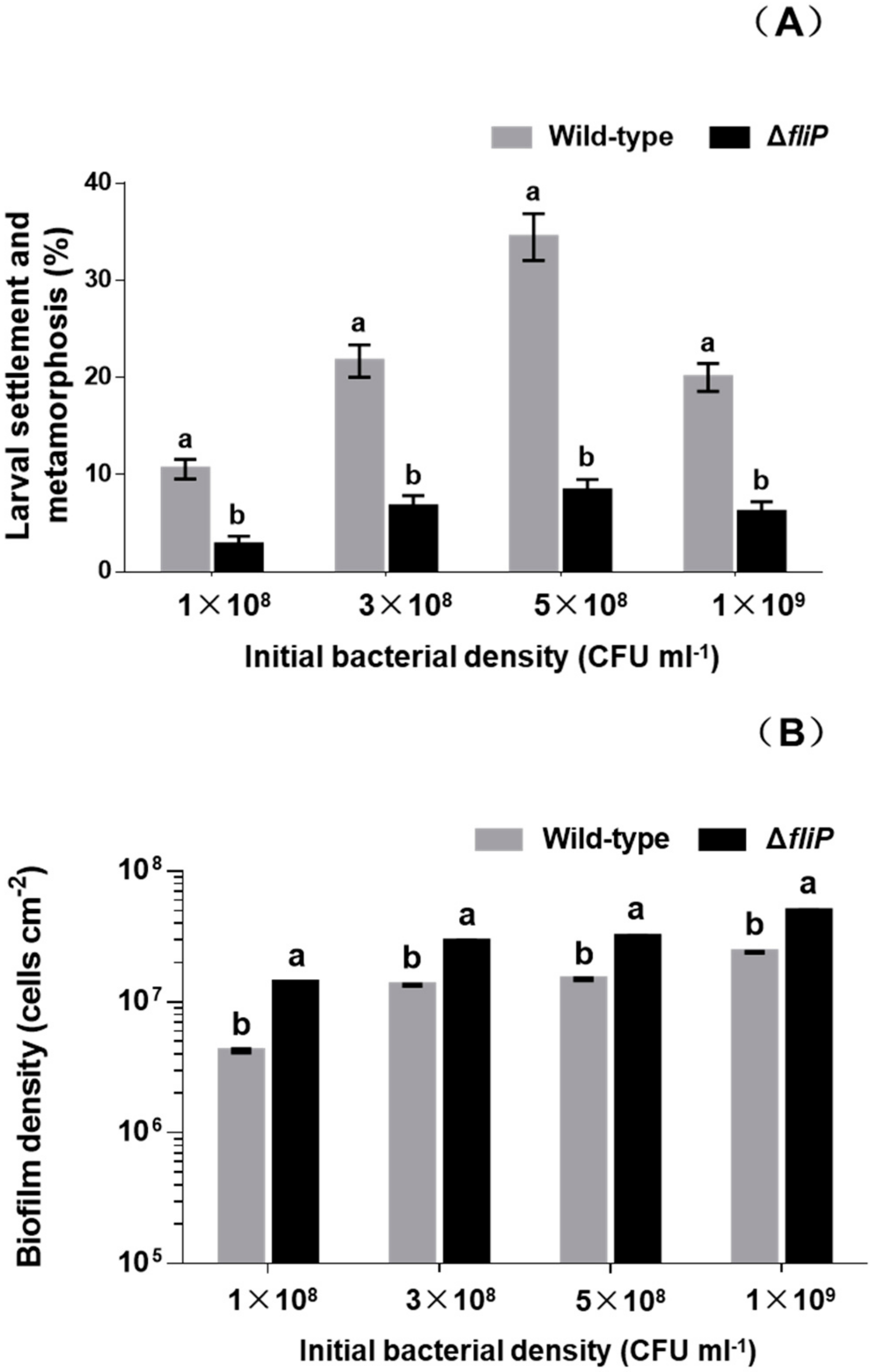
2.1. Deletion of fliP Gene Reduced Mussel Larval Settlement and Metamorphosis
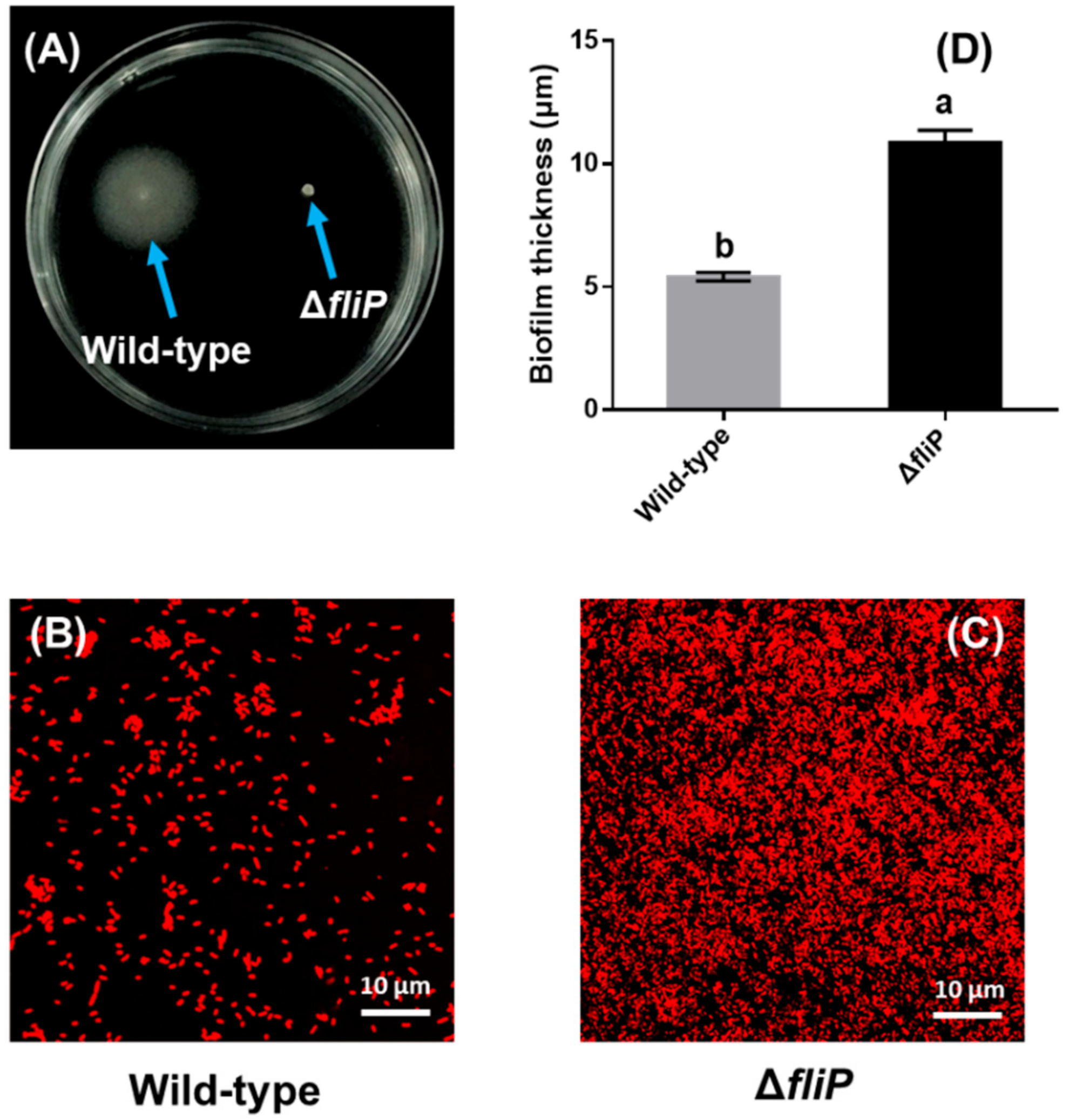
2.2. Effects of fliP Gene Deletion on Flagellum Morphology and Structure to P. marina
2.3. Increased Biofilm Forming Ability after fliP Deletion
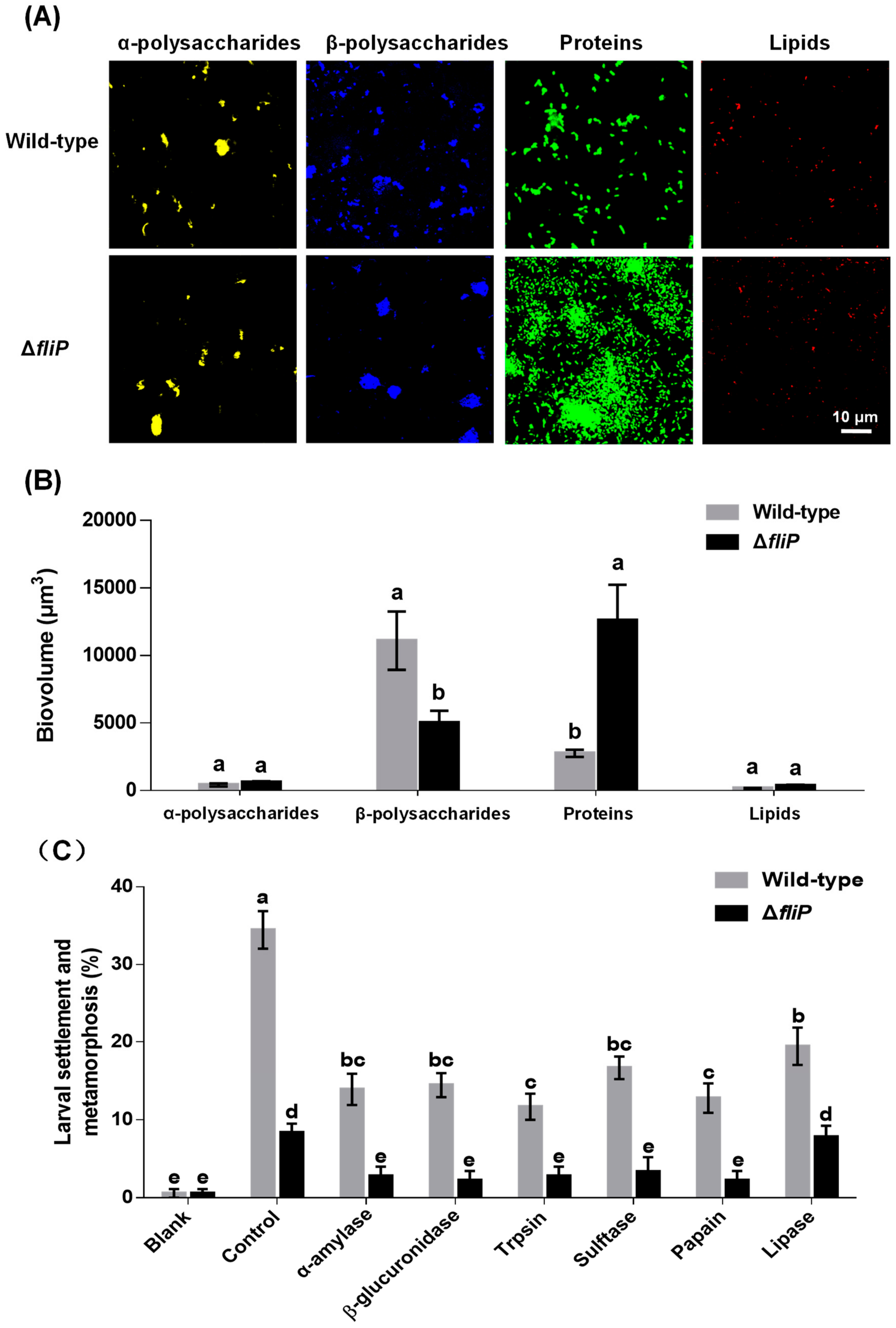
2.4. Variance of Biofilm EPS and Inducing Activities by Enzyme Treatment
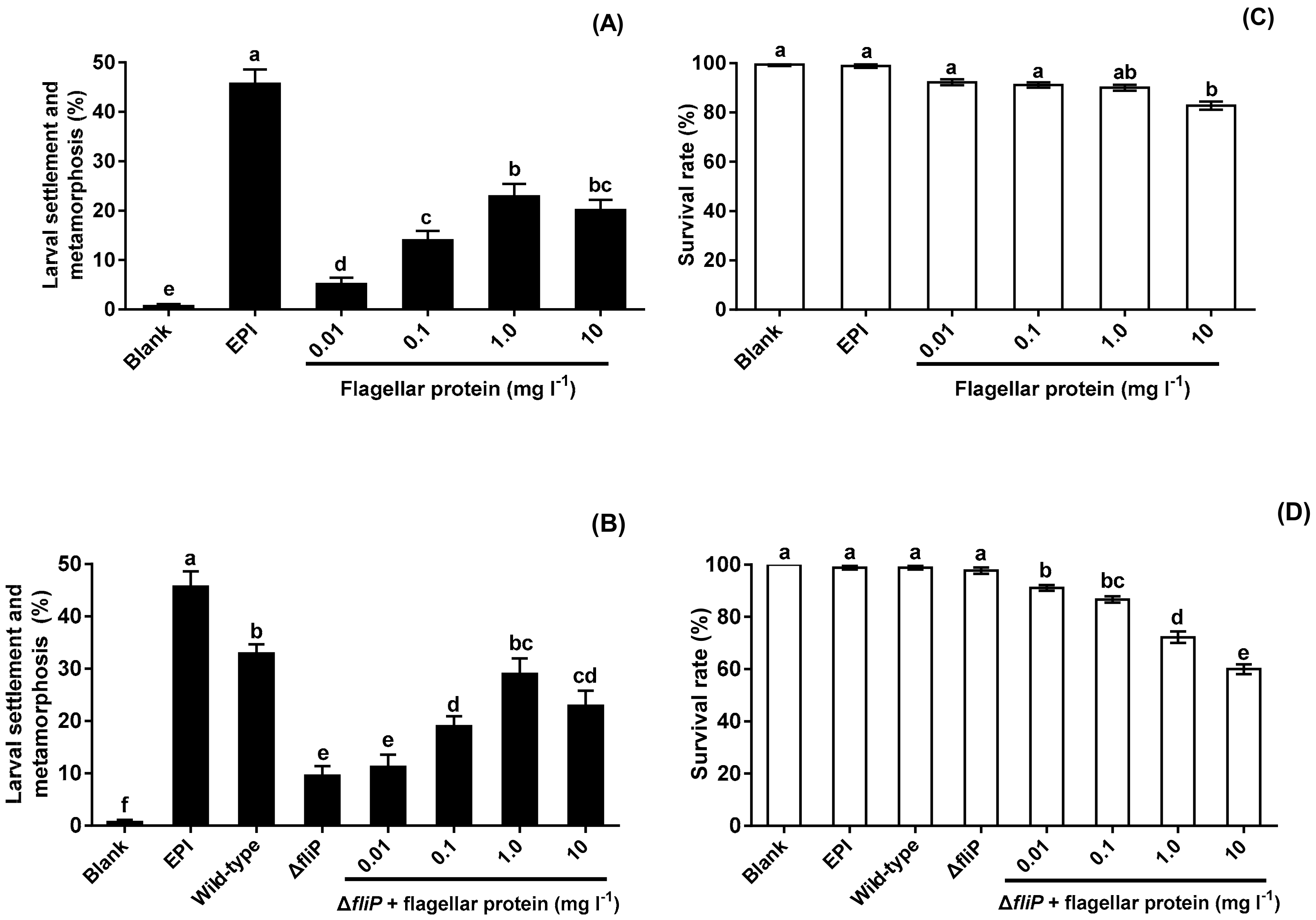
2.5. Effects of Flagellar Protein Extracted on Inducing Activities
3. Discussion
4. Materials and Methods
4.1. Strains, Plasmids, and Culture Conditions
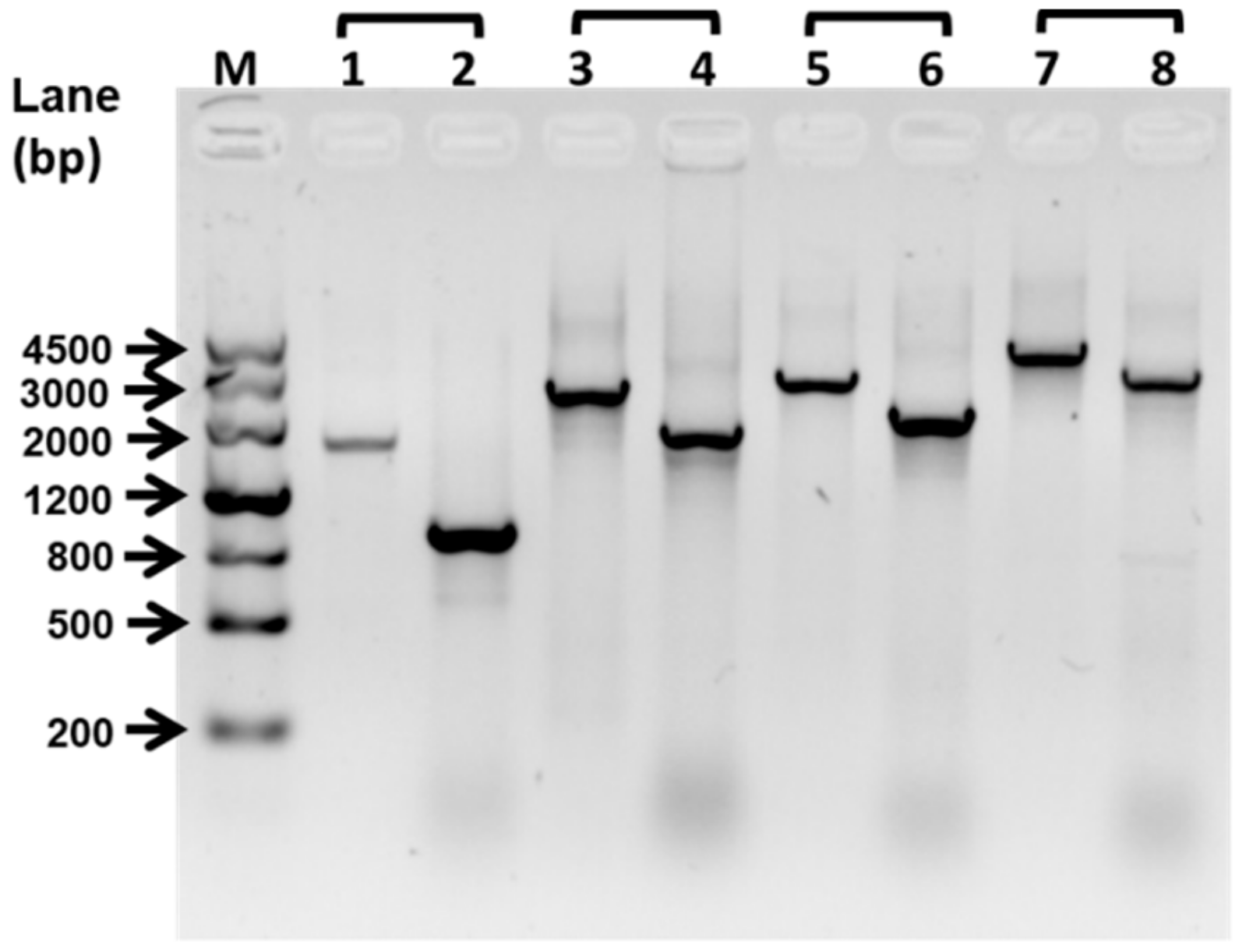
4.2. Construction of ΔfliP Strain
4.3. The Acquisition of Larvae
4.4. The Bioassay of Larval Settlement and Metamorphosis
4.5. Swimming Motility Determination
4.6. Transmission Electron Microscopy (TEM)
4.7. Detection of Bacterial Density of Biofilms
4.8. The Inductive Effect of Extracted Flagellar Proteins
4.9. Analysis of Confocal Laser Scanning Microscopy (CLSM) Images of Biofilms
4.10. Data Statistical analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dang, H.; Lovell, C.R. Microbial surface colonization and biofilm development in marine environments. Microbiol. Mol. Biol. Rev. 2016, 80, 91–138. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Dobretsov, S. Inhibition and induction of marine biofouling by biofilms. In Marine And Industrial Biofouling; Flemming, H.C., Murthy, P.S., Venkatesan, R., Cooksey, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 293–313. [Google Scholar]
- Hadfield, M.G. Biofilms and marine invertebrate larvae: What bacteria produce that larvae use to choose settlement sites. Annu. Rev. Mar. Sci. 2011, 3, 453–470. [Google Scholar] [CrossRef]
- Salta, M.; Capretto, L.; Carugo, D.; Wharton, J.A.; Stokes, K.R. Life under flow: A novel microfluidic device for the assessment of anti-biofilm technologies. Biomicrofluidics 2013, 7, 2879–2893. [Google Scholar] [CrossRef] [PubMed]
- Qian, P.Y.; Dahms, H.U. A triangle model: Environmental changes affect biofilms that affect larval settlement. In Marine and Industrial Biofouling; Flemming, H.C., Murthy, P.S., Venkatesan, R., Cooksey, K., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 315–328. [Google Scholar]
- Lema, K.A.; Constancias, F.; Rice, S.A.; Hadfield, M.G. High bacterial diversity in nearshore and oceanic biofilms and their influence on larval settlement by Hydroides elegans (Polychaeta). Environ. Microbiol. 2019, 21, 3472–3488. [Google Scholar] [CrossRef] [PubMed]
- Fulaz, S.; Vitale, S.; Quinn, L.; Casey, E. Nanoparticle–biofilm interactions: The role of the EPS matrix. Trends. Microbiol. 2019, 27, 915–926. [Google Scholar] [CrossRef] [PubMed]
- Railkin, A.; Tatiana, G.; Oleg, M. Marine Biofouling: Colonization Processes and Defenses; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Zhang, W.; Ding, W.; Li, Y.X.; Tam, C.; Bougouffa, S.; Wang, R.; Pei, B.; Chiang, H.; Leung, P.; Lu, Y.; et al. Marine biofilms constitute a bank of hidden microbial diversity and functional potential. Nat. Commun. 2019, 10, 517. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wuertz, S. Bacteria and archaea on earth and their abundance in biofilms. Nat. Rev. Microbiol. 2019, 17, 247–260. [Google Scholar] [CrossRef]
- Gauthier, G.; Gauthier, M.; Christen, R. Phylogenetic analysis of the genera Alteromonas, Shewanella and Moritella using genes-coding for small-subunit ribosomal-RNA sequences and division of the genus Alteromonas into 2 genera, Alteromonas (Emended) and Pseudoalteromonas gen-nov, and proposal of 12 new species combinations. Inter. J. Syst. Bacteriol. 1995, 45, 755–761. [Google Scholar]
- Holmström, C.; Egan, S.; Franks, A.; McCloy, S.; Kjelleberg, S. Antifouling activities expressed by marine surface associated Pseudoalteromonas species. FEMS Microbiol. Ecol. 2002, 41, 47–58. [Google Scholar] [CrossRef]
- Yang, J.L.; Shen, P.J.; Liang, X.; Li, Y.F.; Bao, W.Y.; Li, J.L. Larval settlement and metamorphosis of the mussel Mytilus coruscus in response to monospecific bacterial biofilms. Biofouling 2013, 29, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Shikuma, N.J.; Pilhofer, M.; Weiss, G.L.; Hadfield, M.G.; Jensen, G.J.; Newman, D.K. Marine tubeworm metamorphosis induced by arrays of bacterial phage tail–like structures. Science 2014, 343, 529–533. [Google Scholar] [CrossRef] [PubMed]
- Ericson, C.F.; Eisenstein, F.; Medeiros, J.M.; Malter, K.E.; Cavalcanti, G.S.; Zeller, R.W.; Newman, D.K.; Pilhofer, M.; Shikuma, N.J. A contractile injection system stimulates tubeworm metamorphosis by translocating a proteinaceous effector. Elife 2019, 8, e46845. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Callahan, S.; Hadfield, M.G. Recruitment in the sea: Bacterial genes required for inducing larval settlement in a polychaete worm. Sci. Rep. 2012, 2, 228. [Google Scholar] [CrossRef] [PubMed]
- Shikuma, N.J.; Antoshechkin, I.; Medeiros, J.M.; Pilhofer, M.; Neman, D.K. Stepwise metamorphosis of the tubeworm Hydroides elegans is mediated by a bacterial inducer and MAPK signaling. Proc. Natl. Acad. Sci. USA 2016, 113, 10097–10102. [Google Scholar] [CrossRef]
- Wang, C.; Bao, W.Y.; Gu, Z.Q.; Li, Y.F.; Liang, X.; Ling, Y.; Cai, S.L.; Shen, H.D.; Yang, J.L. Larval settlement and metamorphosis of the mussel Mytilus coruscus in response to natural biofilms. Biofouling 2012, 28, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.F.; Guo, X.P.; Yang, J.L.; Liang, X.; Bao, W.Y.; Shen, P.J.; Shi, Z.Y.; Li, J.L. Effects of bacterial biofilms on settlement of plantigrades of the mussel Mytilus coruscus. Aquaculture 2014, 433, 434–441. [Google Scholar] [CrossRef]
- Zeng, Z.S.; Guo, X.P.; Li, B.Y.; Wang, P.X.; Cai, X.S.; Tian, X.P.; Zhang, S.; Yang, J.L.; Wang, X.X. Characterization of self-generated variants in Pseudoalteromonas lipolytica biofilm with increased antifouling activities. Appl. Microbiol. Biotechnol. 2015, 99, 10127–10139. [Google Scholar] [CrossRef]
- Zeng, Z.; Guo, X.P.; Cai, X.; Wang, P.; Li, B.; Yang, J.L.; Wang, X. Pyomelanin from Pseudoalteromonas lipolytica reduces biofouling. Microb. Biotechnol. 2017, 10, 1718–1731. [Google Scholar] [CrossRef]
- Peng, L.H.; Liang, X.; Guo, X.P.; Yoshida, A.; Osatomi, K.; Yang, J.L. Complete genome of Pseudoalteromonas marina ECSMB14103, a mussel settlement-inducing bacterium isolated from the East China Sea. Mar. Genom. 2018, 41, 46–49. [Google Scholar] [CrossRef]
- Wieczorek, S.K.; Todd, C.D. Inhibition and facilitation of settlement of epifaunal marine invertebrate larvae by microbial biofilm cues. Biofouling 1998, 12, 81–118. [Google Scholar] [CrossRef]
- Son, K.; Brumley, D.R.; Stocker, R. Live from under the lens: Exploring microbial motility with dynamic imaging and microfluidics. Nat. Rev. Microbiol. 2015, 13, 761–775. [Google Scholar] [CrossRef] [PubMed]
- Macnab, R.M. How bacteria assemble flagella. Annu. Rev. Microbiol. 2003, 57, 77–100. [Google Scholar] [CrossRef]
- O’Toole, G.A.; Kolter, R. Flagellar and twitching motility are necessary for Pseudomonas aeruginosa biofilm development. Mol. Microbiol. 1998, 30, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Watnick, P.I.; Lauriano, C.M.; Klose, K.E.; Croal, L.; Kolter, R. The absence of a flagellum leads to altered colony morphology, biofilm development and virulence in Vibrio cholerae O139. Mol. Microbiol. 2001, 39, 223–235. [Google Scholar] [CrossRef]
- Lemon, K.P.; Higgins, D.E.; Kolter, R. Flagellar motility is critical for Listeria monocytogenes biofilm formation. J. Bacteriol. 2007, 189, 4418–4424. [Google Scholar] [CrossRef] [PubMed]
- Barker, C.S.; Meshcheryakova, I.V.; Inoue, T.; Samatey, F.A. Assembling flagella in Salmonella mutant strains producing a type III export apparatus without FliO. J. Bacteriol. 2014, 196, 4001–4011. [Google Scholar] [CrossRef]
- Guttenplan, S.B.; Blair, K.M.; Kearns, D.B. The EpsE flagellar clutch is bifunctional and synergizes with EPS biosynthesis to promote Bacillus subtilis biofilm formation. PLoS Genet. 2010, 6, e1001243. [Google Scholar] [CrossRef]
- Subramanian, S.B.; Kearns, D.B. Functional regulators of bacterial flagella. Annu. Rev. Microbiol. 2019, 73, 225–246. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef]
- Koo, H.; Allan, R.N.; Howlin, R.P.; Stoodley, P.; Hall-Stoodley, L. Targeting microbial biofilms: current and prospective therapeutic strategies. Nat. Rev. Microbiol. 2017, 15, 740–755. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q.L.; Li, Y.; Zhang, Y.J.; Zhou, Z.M.; Zhang, W.X.; Chen, X.L.; Zhang, X.Y.; Zhou, B.C.; Wang, L.; Zhang, Y.Z. Comparative genomics reveals a deep-sea sediment-adapted life style of Pseudoalteromonas sp. SM9913. ISME J. 2011, 5, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.S.; Peng, L.H.; Guo, X.P.; Yoshida, A.; Osatomi, K.; Li, Y.F.; Yang, J.L.; Liang, X. Complete genome of Pseudoalteromonas atlantica ECSMB14104, a Gammaproteobacterium inducing mussel settlement. Mar. Genom. 2019, 46, 54–57. [Google Scholar] [CrossRef]
- Dahms, H.; Dobretsov, S.; Qian, P. The effect of bacterial and diatom biofilms on the settlement of the bryozoan Bugula Neritina. J. Exp. Mar. Biol. Ecol. 2004, 313, 191–209. [Google Scholar] [CrossRef]
- Huggett, M.J.; Williamson, J.E.; de Nys, R.; Kjelleberg, S.; Steinberg, P.D. Larval settlement of the common Australian sea urchin Heliocidaris erythrogramma in response to bacteria from the surface of coralline algae. Oecologia 2006, 149, 604–619. [Google Scholar] [CrossRef]
- Tebben, J.; Tapiolas, D.M.; Motti, C.A.; Abrego, D.; Negri, A.P.; Blackall, L.L.; Steinberg, P.D.; Harder, T. Induction of larval metamorphosis of the coral Acropora millepora by tetrabromopyrrole isolated from a Pseudoalteromonas bacterium. PLoS ONE 2011, 6, e19082. [Google Scholar] [CrossRef]
- Sneed, J.M.; Sharp, K.H.; Ritchie, K.B.; Paul, V.J. The chemical cue tetrabromopyrrole from a biofilm bacterium induces settlement of multiple Caribbean corals. Proc. Biol. Sci. 2014, 281, 130–143. [Google Scholar] [CrossRef]
- Sun, J.J.; Liang, X.; Guo, X.P.; Chen, Y.R.; Ding, D.W.; Zhang, D.M.; Yang, J.L. Effects of culture media on the biofilm formation and subsequent settlement of Mytilus coruscus. J. Fish. China 2016, 40, 229–1238. [Google Scholar]
- Bao, W.Y.; Yang, J.L.; Satuito, C.G.; Kitamura, H. Larval metamorphosis of the mussel Mytilus galloprovincialis in response to Alteromonas sp. 1: Evidence for two chemical cues? Mar. Biol. 2007, 152, 657–666. [Google Scholar] [CrossRef]
- Kirchman, D.; Graham, S.; Reish, D.; Mitchell, R. Lectins may mediate in the settlement and metamorphosis of Janua (Dexiospira) brasiliensis Grube (Polychaeta: Spirorbidae). Mar. Biol. Lett. 1982, 3, 131–142. [Google Scholar]
- Maki, J.S.; Mitchell, R. Involvement of lectins in the settlement and metamorphosis of marine invertebrate larvae. Bull. Mar. Sci. 1985, 37, 675–683. [Google Scholar]
- Khandeparker, L.; Anil, A.C.; Raghukumar, S. Barnacle larval destination: Piloting possibilities by bacteria and lectin interaction. J. Exp. Mar. Biol. Ecol. 2003, 289, 1–13. [Google Scholar] [CrossRef]
- Brennan, C.A.; Hunt, J.R.; Kremer, N.; Krasity, B.C.; Apicella, M.A.; McFall-Ngai, M.J.; Ruby, E.G. A model symbiosis reveals a role for sheathed-flagellum rotation in the release of immunogenic lipopolysaccharide. Elife 2014, 3, e01579. [Google Scholar] [CrossRef]
- Aschtgen, M.S.; Brennan, C.A.; Nikolakakis, K.; Cohen, S.; McFall-Ngai, M.; Ruby, E.G. Insights into flagellar function and mechanism fromthe squid–vibrio symbiosis. NPJ Biofilms. Microbi. 2019, 5, 32. [Google Scholar] [CrossRef] [PubMed]
- Young, G.M.; Badger, J.L.; Miller, V.L. Motility is required to initiate host cell invasion by Yersinia enterocolitica. Infect. Immun. 2000, 68, 4323–4326. [Google Scholar] [CrossRef]
- Hayashi, F.; Smith, K.D.; Ozinsky, A.; Hawn, T.R.; Yi, E.C.; Goodlett, D.R.; Eng, J.K.; Akira, S.; Underhill, D.M.; Aderem, A. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature 2001, 410, 1099–1103. [Google Scholar] [CrossRef]
- Aschtgen, M.-S.; Wetzel, K.; Goldman, W.; McFall-Ngai, M.; Ruby, E. Vibrio fischeri-derived outer membrane vesicles trigger host development. Cell. Microbiol. 2016, 4, 288–499. [Google Scholar] [CrossRef] [PubMed]
- Dubilier, N. H2S-a settlement cue or a toxic substance for Capitella sp. I larvae? Biol. Bull. 1988, 174, 30–38. [Google Scholar] [CrossRef]
- Yang, J.L.; Satuito, C.G.; Bao, W.Y.; Kitamura, H. Induction of metamorphosis of pediveliger larvae of the mussel Mytilus galloprovincialis Lamarck, 1819 using neuroactive compounds, KCl, NH4Cl and organic solvents. Biofouling 2008, 24, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Chen, K.; Li, Y.F.; Bao, W.Y.; Yoshida, A.; Osatomi, K.; Yang, J.L. An α2-adrenergic receptor is involved in larval metamorphosis in the mussel, Mytilus coruscus. Biofouling 2019, 35, 986–996. [Google Scholar] [CrossRef] [PubMed]
- Harder, T.; Dobretsov, S.; Qian, P.Y. Waterborne polar macromolecules act as algal antifoulants in the seaweed Ulva reticulata. Mar. Ecol. Prog. Ser. 2004, 274, 133–141. [Google Scholar] [CrossRef]
- Jaffar, N.; Ishikawa, Y.; Mizuno, K.; Okinaga, T.; Maeda, T. Mature biofilm degradation by potential probiotics: Aggregatibacter actinomycetemcomitans versus lactobacillus spp. PLoS ONE 2016, 11, e0159466. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, G.F.; Fleet, G.H.; Lyons, M.J.; Walker, R.A. Method for the isolation of highly purified Salmonella flagellins. J. Clin. Microbiol. 1985, 22, 1040–1044. [Google Scholar] [CrossRef] [PubMed]
- González-Machado, C.; Capita1, R.; Riesco-Peláez, F.; Alonso-Calleja, C. Visualization and quantification of the cellular and extracellular components of Salmonella Agona biofilms at different stages of development. PLoS ONE 2018, 13, e0200011. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, X.; Zhang, X.-K.; Peng, L.-H.; Zhu, Y.-T.; Yoshida, A.; Osatomi, K.; Yang, J.-L. The Flagellar Gene Regulates Biofilm Formation and Mussel Larval Settlement and Metamorphosis. Int. J. Mol. Sci. 2020, 21, 710. https://doi.org/10.3390/ijms21030710
Liang X, Zhang X-K, Peng L-H, Zhu Y-T, Yoshida A, Osatomi K, Yang J-L. The Flagellar Gene Regulates Biofilm Formation and Mussel Larval Settlement and Metamorphosis. International Journal of Molecular Sciences. 2020; 21(3):710. https://doi.org/10.3390/ijms21030710
Chicago/Turabian StyleLiang, Xiao, Xiu-Kun Zhang, Li-Hua Peng, You-Ting Zhu, Asami Yoshida, Kiyoshi Osatomi, and Jin-Long Yang. 2020. "The Flagellar Gene Regulates Biofilm Formation and Mussel Larval Settlement and Metamorphosis" International Journal of Molecular Sciences 21, no. 3: 710. https://doi.org/10.3390/ijms21030710
APA StyleLiang, X., Zhang, X.-K., Peng, L.-H., Zhu, Y.-T., Yoshida, A., Osatomi, K., & Yang, J.-L. (2020). The Flagellar Gene Regulates Biofilm Formation and Mussel Larval Settlement and Metamorphosis. International Journal of Molecular Sciences, 21(3), 710. https://doi.org/10.3390/ijms21030710