Effect of S-Allyl –L-Cysteine on MCF-7 Cell Line 3-Mercaptopyruvate Sulfurtransferase/Sulfane Sulfur System, Viability and Apoptosis



Abstract
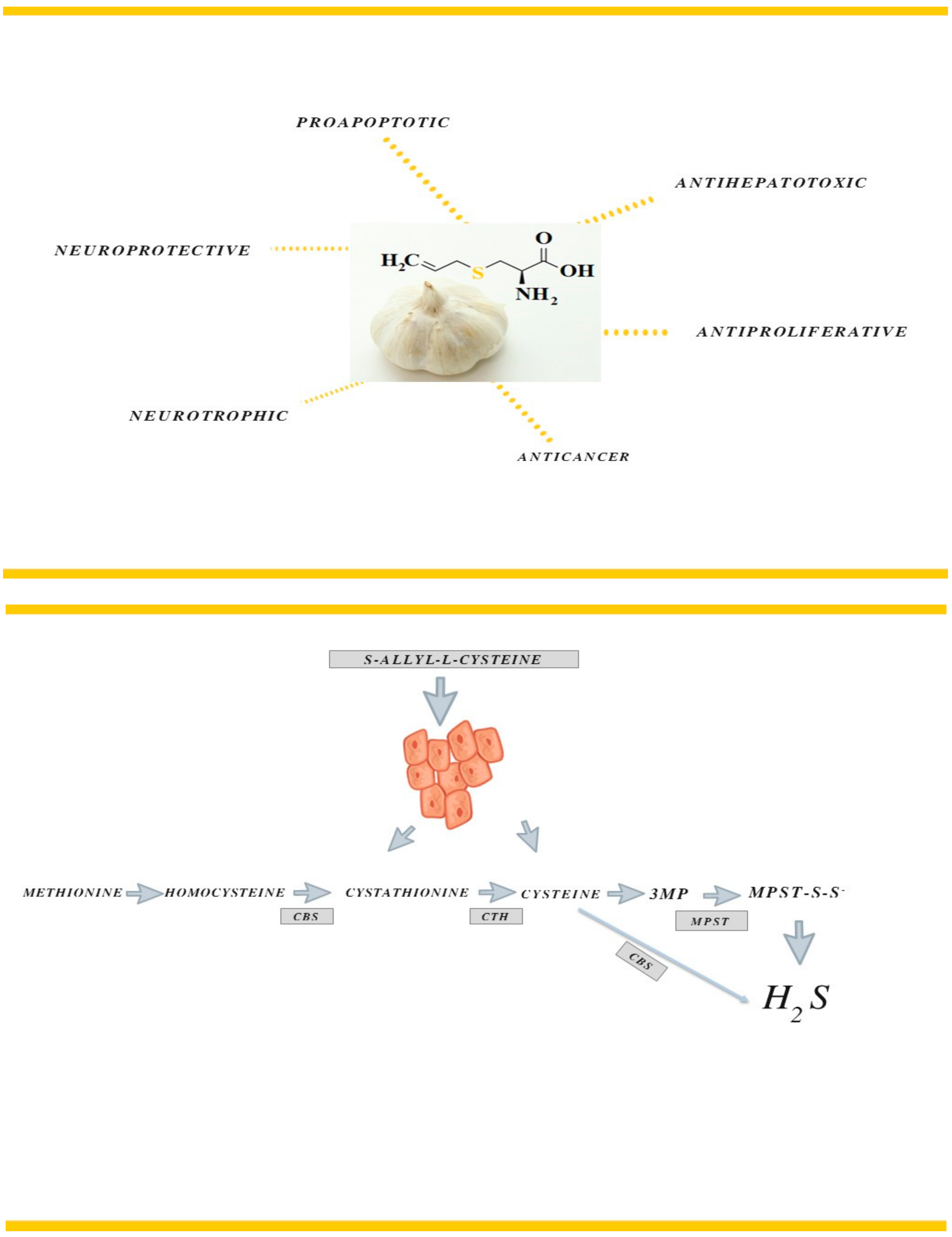
1. Introduction
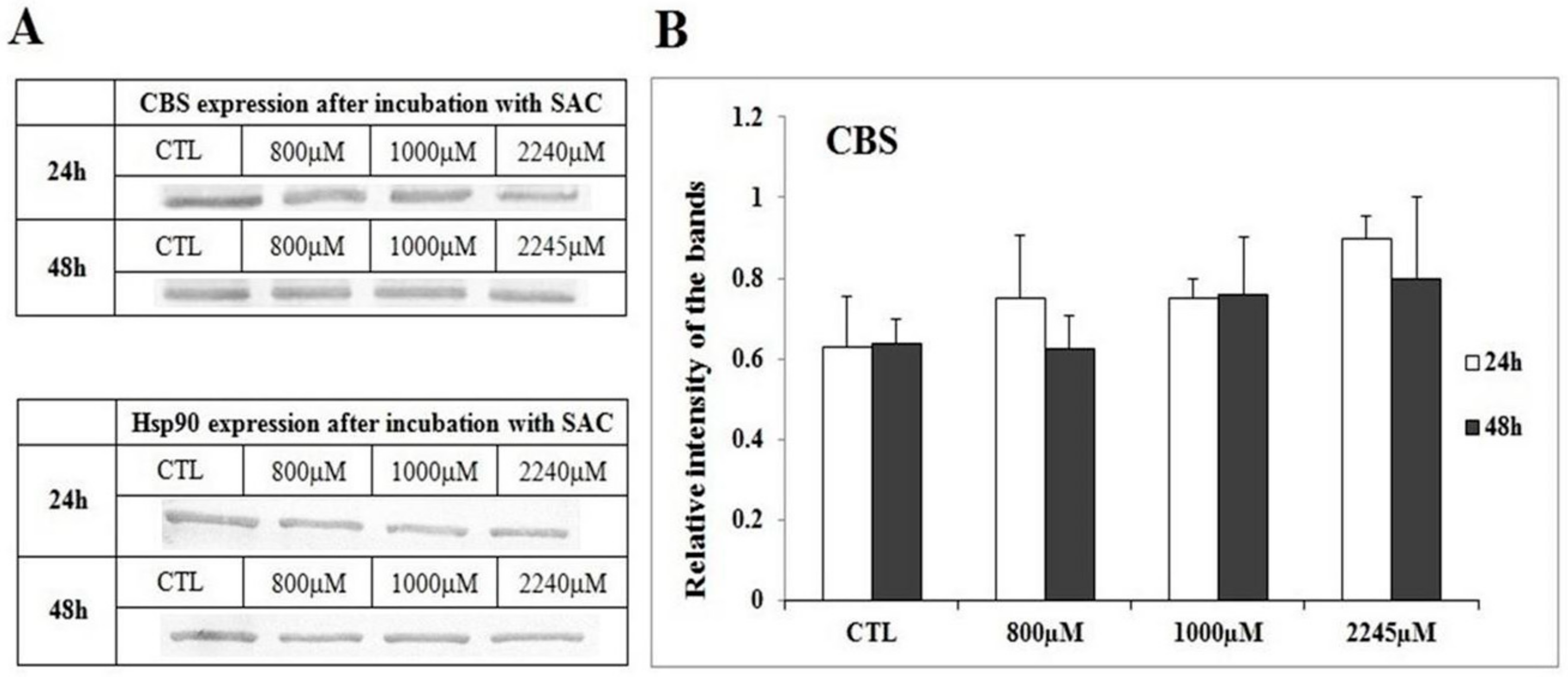
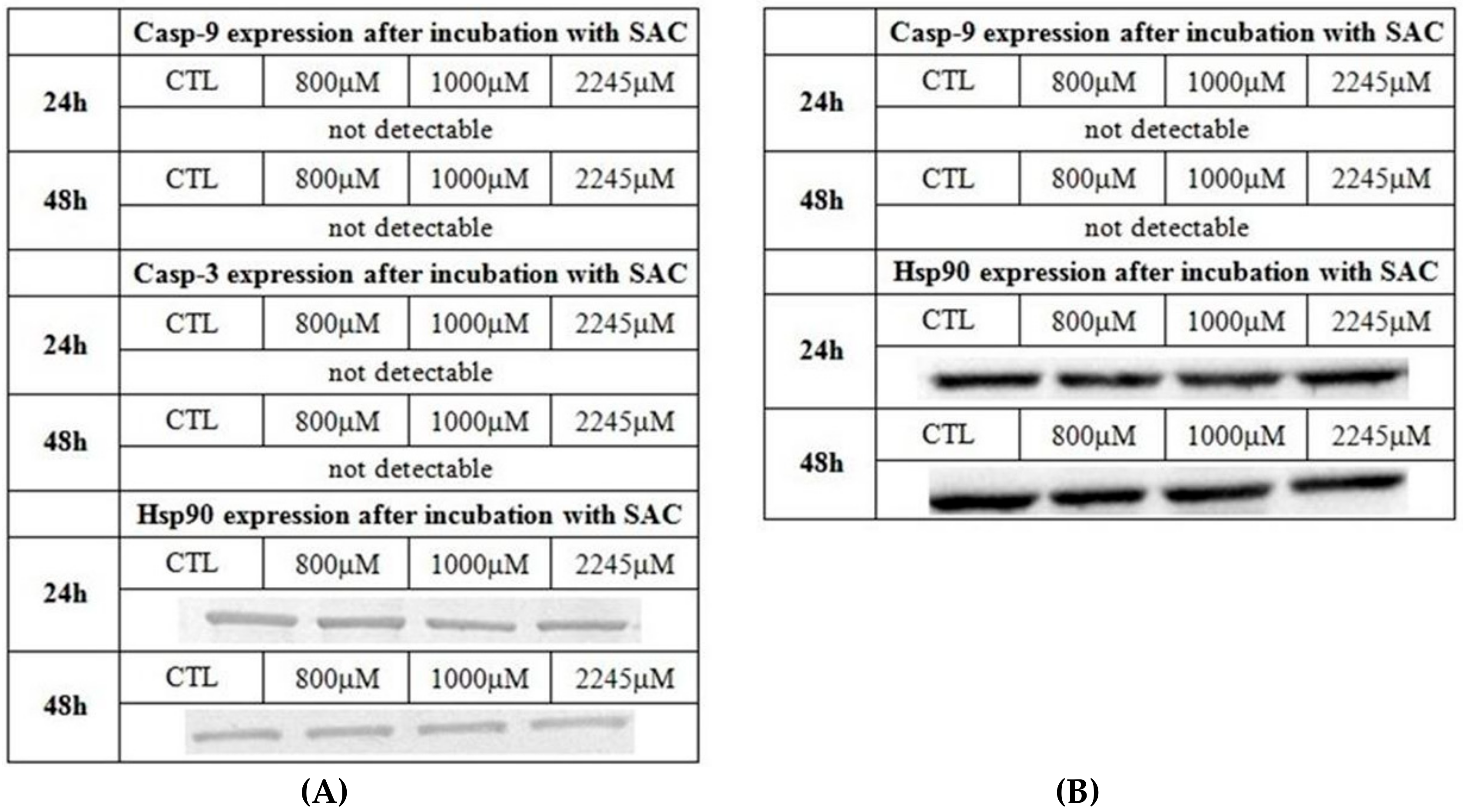
2. Results
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture
4.3. Cell Treatment
4.4. Cytotoxicity of Reagents
4.5. Cell Viability by MTT Assay
4.6. Flow Cytometry (FC)
4.7. Western Blot Analysis
4.8. Densitometric Evaluation of Western Blot bands
4.9. Enzymes Assay
4.9.1. Cell Homogenization
4.9.2. MPST Activity
4.9.3. CTH Activity
4.9.4. Sulfane Sulfur
4.9.5. Protein
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AGE | Aged garlic extract |
| ATCC | American Type Culture Collections |
| casp-3 | caspase-3 |
| casp-9 | caspase-9 |
| CBS | cystathionine β-lyase |
| CTH | γ-cystathionase |
| CTL | control |
| DTT | D,L-dithiothreitol |
| NEM | N-ethylmaleimide |
| FC | flow cytometry |
| Gi | growth inhibition |
| Hsp90 | heat shock protein 90 |
| LDH | L-lactic dehydrogenase |
| MGL | methionine γ-lyase |
| MPST | 3-mercaptopyruvate sulfurtransferase |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide |
| NADH | β-Nicotinamide adenine dinucleotide reduced disodium salt hydrate |
| PA | pyruvate |
| PCA | perchloric acid |
| PLP | pyridoxal phosphate |
| SAC | S-allyl- L-cysteine |
| SAM | S-adenosyl-L-methionine |
| SD | standard deviation |
| WB | Western blot |
| 3MP | 3-mercaptopyruvate |
References
- Colín-González, A.L.; Santana, R.A.; Silva-Islas, C.A.; Chánez-Cárdenas, M.E.; Santamaría, A.; Maldonado, P.D. The Antioxidant Mechanisms Underlying the Aged Garlic Extract- and S-Allylcysteine-Induced Protection. Oxidative Med. Cell. Longev. 2012, 907162, 16. [Google Scholar] [CrossRef]
- DeSantis, C.E.; Ma, J.; Goding Sauer, A.; Newman, L.A.; Jemal, A. Breast cancer statistics, 2017, racial disparity in mortality by state. Cancer J. Clin. 2017, 67, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Cave, D.D.; Desiderio, V.; Mosca, L.; Ilisso, C.P.; Mele, L.; Caraglia, M.; Cacciapuoti, G.; Porcelli, M. S-Adenosylmethionine-mediated apoptosis is potentiated by autophagy inhibition induced by chloroquine in human breast cancer cells. J. Cell Physiol. 2018, 233, 1370–1383. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Li, Y.N.; Wang, F.; Zhang, W.M.; Geng, X. S-adenosylmethionine inhibits the growth of cancer cells by reversing the hypomethylation status of c-myc and H-ras in human gastric cancer and colon cancer. Int. J. Biol. Sci. 2010, 6, 784–795. [Google Scholar] [CrossRef] [PubMed]
- Mosca, L.; Pagano, M.; Ilisso, C.P.; Cave, D.D.; Desiderio, V.; Mele, L.; Caraglia, M.; Cacciapuoti, G.; Porcelli, M. AdoMet triggers apoptosis in head and neck squamous cancer by inducing ER stress and potentiates cell sensitivity to cisplatin. J. Cell Physiol. 2019, 234, 13277–132916. [Google Scholar] [CrossRef] [PubMed]
- Javed, H.; Khan, M.M.; Khan, A.; Vaibhav, K.; Ahmad, A.; Khuwaja, G.; Ahmed, M.E.; Raza, S.S.; Ashafaq, M.; Tabassum, R.; et al. S-allyl cysteine attenuates oxidative stress associated cognitive impairment and neurodegeneration in mouse model of streptozotocin-induced experimental dementia of Alzheimer’s type. Brain Res. 2011, 10, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.N.; Kang, M.; Lee, S.; Oh, J.J.; Hong, S.K.; Lee, S.E.; Byun, S.S. Anticancer effect of S-allyl-L-cysteine via induction of apoptosis in human bladder cancer cells. Oncol. Letters. 2018, 15, 623–629. [Google Scholar] [CrossRef]
- Bhuiyan, A.I.; Papajani, V.T.; Paci, M.; Melino, S. Glutathione-Garlic Sulfur Conjugates: Slow Hydrogen Sulfide Releasing Agents for Therapeutic Applications. Molecules 2015, 20, 1731–1750. [Google Scholar] [CrossRef]
- Welch, C.; Wuarin, L.; Sidell, N. Antiproliferative effect of the garlic compound S-allyl cysteine on human neuroblastoma cells in vitro. Cancer Lett. 1992, 63, 211–219. [Google Scholar] [CrossRef]
- Chu, Q.; Ling, M.T.; Feng, H.; Cheung, H.W.; Tsao, S.W.; Wang, X.; Wong, Y.C. A novel anticancer effect of garlic derivatives: Inhibition of cancer cell invasion through restoration of E-cadherin expression. Carcinogenesis 2006, 27, 2180–2189. [Google Scholar] [CrossRef]
- Pai, M.H.; Kuo, Y.H.; Chiang, E.P.; Tang, F.Y. S-Allylcysteine inhibits tumour progression and the epithelial-mesenchymal transition in a mouse xenograft model of oral cancer. Br. J. Nutr. 2012, 14, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.T.; Guo, D.Y.; Cheng, Q.; Geng, W.; Ling, C.C.; Li, C.X.; Liu, X.B.; Ma, Y.Y.; Lo, C.M.; Poon, R.T.; et al. A garlic derivative, S-allylcysteine (SAC), suppresses proliferation and metastasis of hepatocellular carcinoma. PLoS ONE 2012, 7, e31655. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.S.; Feng, J.G.; Zhang, D.; Zhang, B.; Luo, M.; Su, D.; Lin, N.M. S-allylcysteine, a garlic derivative, suppresses proliferation and induces apoptosis in human ovarian cancer cells in vitro. Acta Pharmacol. Sin. 2014, 35, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Nagahara, N. Multiple role of 3-mercaptopyruvate sulfurtransferase: Antioxidative function, H2S and polysulfide production and possible SOx production. Br. J. Pharmacol. 2018, 175, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Bronowicka-Adamska, P.; Bentke, A.; Wróbel, M. Hydrogen sulfide generation from L-cysteine in the human glioblastoma-astrocytoma U-87 MG and neuroblastoma SHSY5Y cell lines. Acta Biochim. Pol. 2017, 64, 171–176. [Google Scholar] [PubMed]
- Sen, S.; Kawahara, B.; Gupta, D.; Tsai, R.; Khachatryan, M.; Roy-Chowdhuri, S.; Bose, S.; Yoon, A.; Faull, K.; Farias-Eisner, R.; et al. Role of cystathionine b-synthase in human breast cancer. Free Radic. Biol. Med. 2015, 86, 228–238. [Google Scholar] [CrossRef]
- Sen, S.; Kawahara, B.; Mahata, S.K.; Tsai, R.; Yoon, A.; Hwang, L.; Hu-Moore, K.; Villanueva, C.; Vajihuddin, A.; Parameshwar, P.; et al. Cystathionine: A novel on cometabolite in human breast cancer. Arch. Biochem. Biophys. 2016, 15, 95–102. [Google Scholar] [CrossRef]
- Pokrovsky, V.S.; Anisimova, N.Y.; Davydov, D.Z.; Bazhenov, S.V.; Bulushova, N.V.; Zavilgelsk, G.B.; Kotova, V.Y.; Manukhov, I.V. Methionine Gamma Lyase from Clostridium sporogenes Increases the Anticancer Efficacy of Doxorubicin on A549 Cancer Cells In Vitro and Human Cancer Xenografts. Methods Mol. Biol. 2019, 1866, 243–261. [Google Scholar]
- Lasota, M.; Klein, A.; Balwierz, W. Cytostatic and cytotoxic effects of tyrphostin AG1296 on RMS cells. Contemp. Oncol. (Pozn) 2012, 16, 1–5. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Wong, R.S.Y. Apoptosis in cancer: From pathogenesis to treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.; Watari, H.; AbuAlmaaty, A.; Ohba, Y.; Sakuragi, N. Apoptosis and Molecular Targeting Therapy in Cancer. J. BioMed. Biotechnol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Bidkar, A.B.; Sanapui, P.; Ghosh, S.S. Efficient induction of apoptosis in cancer cells by paclitaxel-loaded selenium nanoparticles. Nanomedicine 2017, 12, 2641–2651. [Google Scholar] [CrossRef] [PubMed]
- Nam, G.H.; Jo, K.J.; Park, Y.S.; Kawk, H.W.; Kim, S.Y.; Kim, Y.M. In vitro and in vivo Induction of p53-Dependent Apoptosis by Extract of Euryale ferox Salisb in A549 Human Caucasian Lung Carcinoma Cancer Cells Is Mediated Through Akt Signaling Pathway. Front. Oncol. 2019, 9, 406. [Google Scholar] [CrossRef] [PubMed]
- Looi, C.Y.; Arya, A.; Cheah, F.K.; Muharram, B.; Leong, K.H.; Mohamad, K.; Wong, W.F.; Rai, N.; Mustafa, M.R. Induction of Apoptosis in Human Breast Cancer Cells via Caspase Pathway by Vernodalin Isolated from Centratherum anthelminticum. PLoS ONE 2013, 8, e56643. [Google Scholar] [CrossRef]
- Sivakumaran, N.; Samarakoon, S.R.; Adhikari, A.; Ediriweera, M.K.; Tennekoon, K.H.; Malavige, N.; Thabrew, I.; Shrestha, R.L.S. Cytotoxic and Apoptotic Effects of Govaniadine Isolated from Corydalis govaniana Wall. Roots on Human Breast Cancer (MCF-7) Cells. Biomed. Res. Int. 2018, 2018, 1–11. [Google Scholar] [CrossRef]
- Oladghaffari, M.; Islamian, J.P.; Baradaran, B.; Monfared, A.S.; Farajollahi, A.; Shanehbandi, D.; Mohammadi, M. High Efficiency Apoptosis Induction in Breast Cancer Cell Lines by MLN4924/2DG Co-Treatment. Asian Pac. J. Cancer Prev. 2015, 16, 5471–5476. [Google Scholar] [CrossRef]
- Yang, H.L.; Chen, C.S.; Chang, W.H.; Lu, F.J.; Lai, Y.C.; Chen, C.C.; Hseu, T.H.; Kuo, C.T.; Hseu, Y.C. Growth inhibition and induction of apoptosis in MCF-7 breast cancer cells by Antrodia camphorate. Cancer Letters. 2006, 231, 215–227. [Google Scholar] [CrossRef]
- Tor, Y.S.; Yazan, L.S.; Foo, J.B.; Wibowo, A.; Ismail, N.; Cheah, Y.K.; Abdullah, R.; Ismail, M.; Ismail, I.S.; Yeap, S.K. Induction of Apoptosis in MCF-7 Cells via Oxidative Stress Generation, Mitochondria-Dependent and Caspase-Independent Pathway by Ethyl Acetate Extract of Dillenia suffruticosa and Its Chemical Profile. PLoS ONE 2015, 10, e0127441. [Google Scholar] [CrossRef]
- Pilco-Ferreto, N.; Calaf, G.M. Influence of doxorubicin on apoptosis and oxidative stress in breast cancer cell lines. Int. J. Oncol. 2016, 49, 753–762. [Google Scholar] [CrossRef]
- Shunsuke, K.; Gu, J.; Honda, T.; McDonnell, T.J.; Swisher, S.G.; Roth, J.A.; Fang, B. Deficiency of caspase-3 in MCF7 cells blocks Bax-mediated nuclear fragmentation but not cell death. Clin Cancer Res. 2001, 7, 1474–1480. [Google Scholar]
- Węsierska-Gądek, J.; Hackl, S.; Zulehner, N.; Maurer, M.; Komina, O. Reconstitution of human MCF-7 breast cancer cells with caspase-3 does not sensitize them to action of CDK inhibitors. J. Cell Biochem. 2011, 112, 273–288. [Google Scholar] [CrossRef] [PubMed]
- Turner, C.; Devitt, A.; Parker, K.; MacFarlane, M.; Giuliano, M.; Cohen, G.M.; Gregory, C.D. Macrophage-mediated clearance of cells undergoing caspase-3-independent death. Cell Death Differ. 2003, 10, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Rosado, J.O.; Salvador, M.; Bonatto, D. Importance of the trans- sulfuration pathway in cancer prevention and promotion. Mol. Cell Biochem. 2007, 301, 1–12. [Google Scholar] [CrossRef]
- Gapter, L.A.; Yuin, O.Z.; Ng, K.Y. S-Allylcysteine reduces breast tumor cell adhesion and invasion. Biochem. Biophys. Res. Commun. 2008, 367, 446–451. [Google Scholar] [CrossRef]
- Tang, F.Y.; Chiang, E.P.; Pai, M.H. Consumption of S-allylcysteine inhibits the growth of human non-small-cell lung carcinoma in a mouse xenograft model. J. Agric. Food Chem. 2010, 58, 11156–11164. [Google Scholar] [CrossRef]
- You, J.; Shi, X.; Liang, H.; Ye, J.; Wang, L.; Han, H.; Fang, H.; Kang, W.; Wang, T. Cystathionine- γ-lyase promotes process of breast cancer in association with STAT3 signaling pathway. Oncotarget 2017, 8, 65677–65686. [Google Scholar] [CrossRef]
- Augsburger, F.; Szabo, C. Potential role of the 3-mercaptopyruvate sulfurtransferase (3-MST)—hydrogen sulfide (H2S) pathway in cancer cells. Pharmacol. Res. 2018, 27, 104083. [Google Scholar] [CrossRef]
- Gallegos-Arreola, M.P.; Figuera-Villanueva, L.E.; Ramos-Silva, A.; Salas-González, E.; Puebla-Pérez, A.M.; Peralta-Leal, V.; García-Ortiz, J.E.; Dávalos-Rodríguez, I.P.; Zúñiga-González, G.M. The association between the 844ins68 polymorphism in the CBS gene and breast cancer. Arch. Med. Sci. 2014, 10, 1214–1224. [Google Scholar] [CrossRef]
- Ramasamy, S.; Singh, S.; Taniere, P.; Langman, M.J.S.; Eggo, M.C. Sulfide-detoxifying enzymes in the human colon are decreased in cancer and upregulated in differentiation. Am. J. Physiol Gastrointest Liver Physiol 2006, 291, G288–G296. [Google Scholar] [CrossRef]
- Gai, J.W.; Wahafu, W.; Guo, H.; Liu, M.; Wang, X.C.; Xiao, Y.X.; Zhang, L.; Xin, Z.C.; Jin, J. Further evidence of endogenous hydrogen sulfide as a mediator of relaxation in human and rat bladder. Asian J. Androl. 2013, 15, 692–696. [Google Scholar] [CrossRef] [PubMed]
- Zuhra, K.; Tomé, C.S.; Masi, L.; Giardina, G.; Paulini, G.; Malagrinò, F.; Forte, E.; Vicente, J.B.; Giuffrè, A. N-Acetylcysteine Serves as Substrate of 3-Mercaptopyruvate Sulfurtransferase and Stimulates Sulfide Metabolism in Colon Cancer. Cells 2019, 8, 828. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Ding, L.; Xie, Z.Z.; Yang, Y.; Whiteman, M.; Moore, P.K.; Bian, J.S. A Review of Hydrogen Sulfide Synthesis, Metabolism, and Measurement: Is Modulation of Hydrogen Sulfide a Novel Therapeutic for Cancer? Antioxidants Redox Signaling 2019, 31, 1–38. [Google Scholar]
- Shinkai, Y.; Kumagai, Y. Sulfane Sulfur in Toxicology: A Novel Defense System Against Electrophilic Stress. Toxicol. Sci. 2019, 170, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Greer, S.; Honeywell, R.; Geletu, M.; Arulanandam, R.; Raptis, L. Housekeeping genes; expression levels may change with density of cultured cells. J. Immunol. Methods 2010, 15, 76–79. [Google Scholar] [CrossRef]
- Pick, E.; Kluger, Y.; Giltnane, J.M.; Moeder, C.H.; Camp, R.L.; Rimm, D.R.; Kluger, H.M. High HSP90 Expression Is Associated with Decreased Survival in Breast Cancer. Cancer Res. 2007, 1, 2932–2937. [Google Scholar] [CrossRef]
- Valentine, W.N.; Frankelfeld, J.K. 3-Mercaptopyuruvate sulfurtransferase (EC 2.8.1.2): A simple assay adapted to human blood cells. Clin. Chim. Acta 1974, 14, 205–210. [Google Scholar] [CrossRef]
- Wróbel, M.; Jurkowska, H.; Śliwa, L.; Srebro, Z. Sulfurtransferases and cyanide detoxification in mouse liver, kidney, and brain. Toxicol. Mech. Methods 2004, 14, 331–337. [Google Scholar] [CrossRef]
- Matsuo, Y.; Greenberg, D.M. A crystalline enzyme that cleaves homoserine and cystathionine. J. Biol. Chem. 1958, 230, 545–560. [Google Scholar]
- Czubak, J.; Wróbel, M.; Jurkowska, H. Cystathionine γ-lyase (EC 4.4.1.1): An enzymatic assay of α-ketobutyrate using lactate dehydrogenase. Acta Biol. Cracov. Ser. Zool. 2002, 44, 113–117. [Google Scholar]
- Wood, L. Sulfane sulfur. Methods Enzymol. 1987, 143, 25–29. [Google Scholar] [PubMed]
- Lowry, O.H.; Rosenbrough, N.J.; Farr, A.L.; Randall, R.I. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [PubMed]
- Bradford, M.M. A rapid and sensitive for the quantitation of microgram quantitites of protein utilizing the principle of protein-dye binding. Analytical Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 24 h | 48 h | |
|---|---|---|
| % of Cytotoxicity | ||
| 800 µM | 0 | 0 |
| 1000 µM | 0 | 0 |
| 2245 µM | 0.2 | 0 |
| MCF-7 Cell Line 24 h | MPST | CTH | Sulfane Sulfur |
|---|---|---|---|
| nmol·106·cells−1·min−1 | nmol·106·cells−1 | ||
| CTL | 2714 ± 904 | 25 ± 9 | 826 ± 249 |
| 800 µM | 3042 ± 310 | 11 ± 6* | 949 ± 279 |
| 1000 µM | 3636 ± 818 | 5 ± 1* | 999 ± 198 |
| 2245 µM | 1828 ± 389 *Δ | 25 ± 16 | 497 ± 100 *Δ |
| MCF-7 Cells 48 h | MPST | CTH | Sulfane Sulfur |
|---|---|---|---|
| nmol·106·cells−1·min−1 | nmol·106·cells−1 | ||
| CTL | 3261 ± 798 | 30 ± 10 | 929 ± 194 |
| 800 µM | 2580 ± 761 | 50 ± 10* | 1731 ± 426* |
| 1000 µM | 4229 ± 830* | 10 ± 5* | 1905 ± 430* |
| 2245 µM | 2019 ± 382 Δ | 20 ± 1 | 571 ± 116*Δ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bronowicka-Adamska, P.; Bentke, A.; Lasota, M.; Wróbel, M. Effect of S-Allyl –L-Cysteine on MCF-7 Cell Line 3-Mercaptopyruvate Sulfurtransferase/Sulfane Sulfur System, Viability and Apoptosis. Int. J. Mol. Sci. 2020, 21, 1090. https://doi.org/10.3390/ijms21031090
Bronowicka-Adamska P, Bentke A, Lasota M, Wróbel M. Effect of S-Allyl –L-Cysteine on MCF-7 Cell Line 3-Mercaptopyruvate Sulfurtransferase/Sulfane Sulfur System, Viability and Apoptosis. International Journal of Molecular Sciences. 2020; 21(3):1090. https://doi.org/10.3390/ijms21031090
Chicago/Turabian StyleBronowicka-Adamska, Patrycja, Anna Bentke, Małgorzata Lasota, and Maria Wróbel. 2020. "Effect of S-Allyl –L-Cysteine on MCF-7 Cell Line 3-Mercaptopyruvate Sulfurtransferase/Sulfane Sulfur System, Viability and Apoptosis" International Journal of Molecular Sciences 21, no. 3: 1090. https://doi.org/10.3390/ijms21031090
APA StyleBronowicka-Adamska, P., Bentke, A., Lasota, M., & Wróbel, M. (2020). Effect of S-Allyl –L-Cysteine on MCF-7 Cell Line 3-Mercaptopyruvate Sulfurtransferase/Sulfane Sulfur System, Viability and Apoptosis. International Journal of Molecular Sciences, 21(3), 1090. https://doi.org/10.3390/ijms21031090