A Transcriptomic Insight into the Impact of Colon Cancer Cells on Mast Cells


Abstract
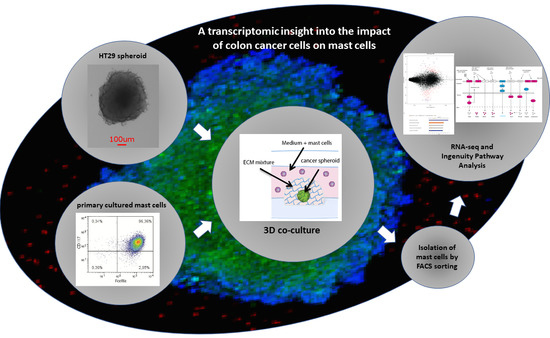
1. Introduction
2. Results
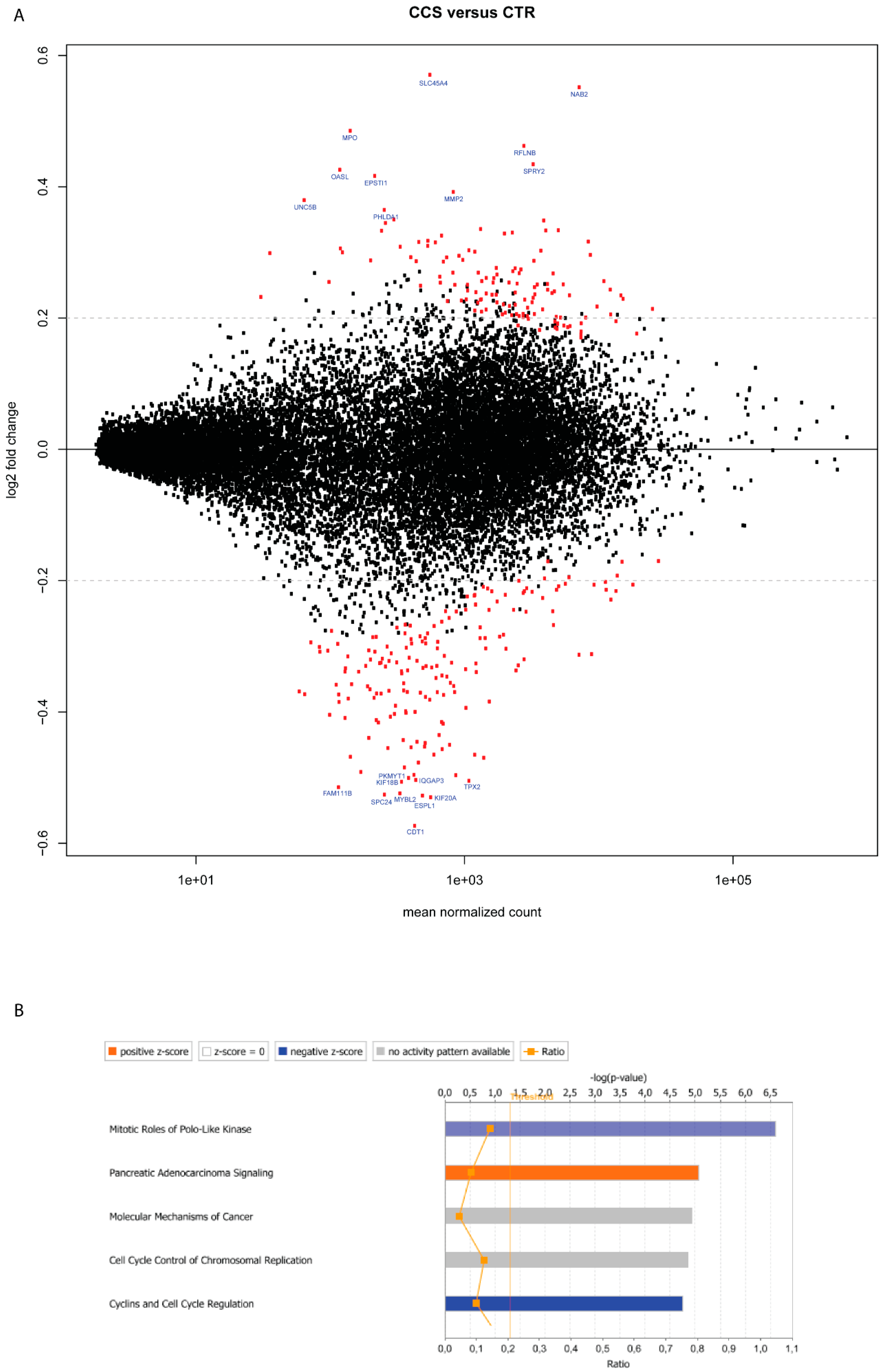
2.1. Deregulated Genes in Colon Cancer-Cocultured MCs
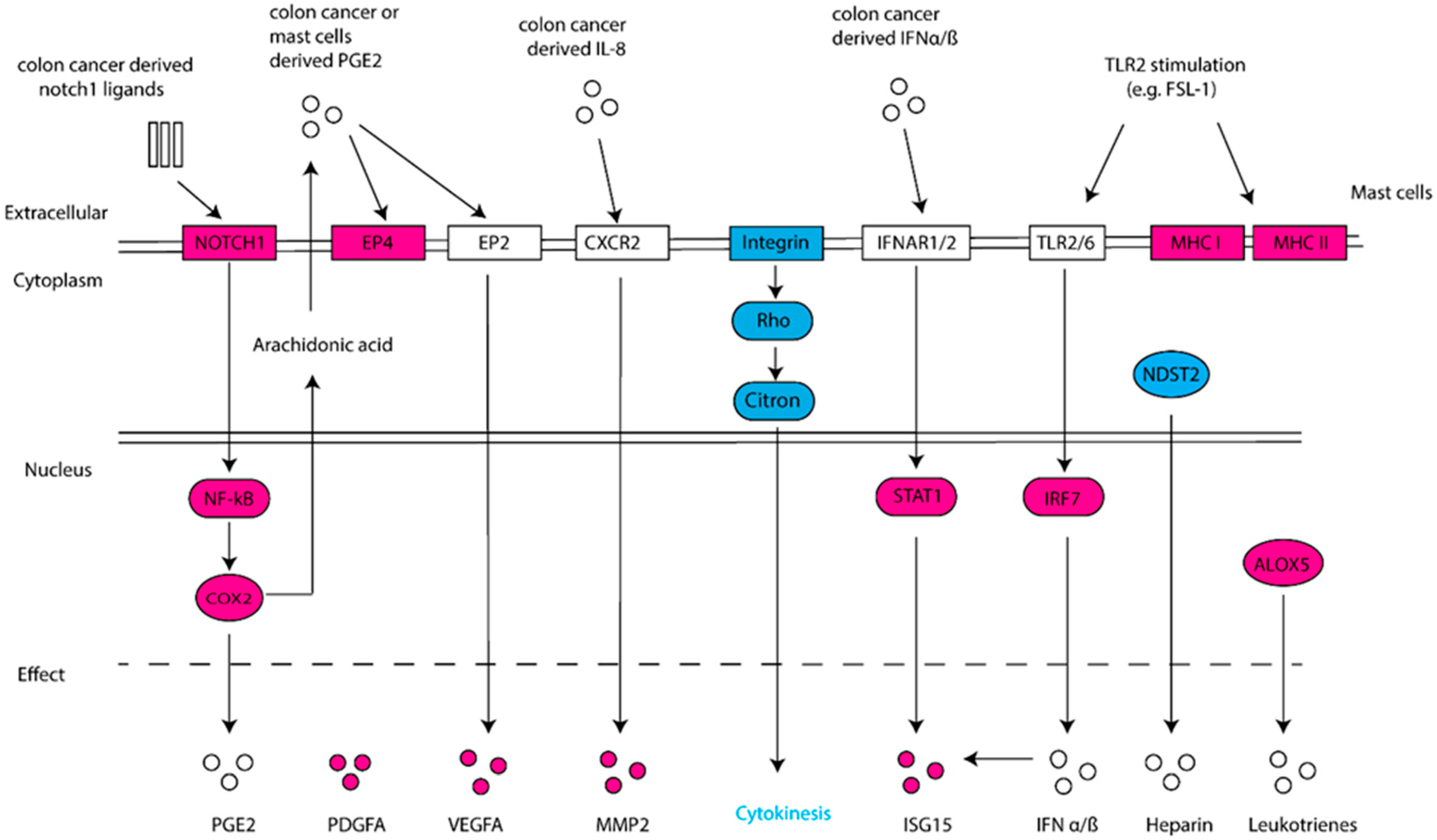
2.2. Gene Deregulation for Extracellular Mediators in Colon Cancer-Cocultured MCs
2.3. Gene Deregulation for Membrane Receptors in Colon Cancer-Cocultured MCs
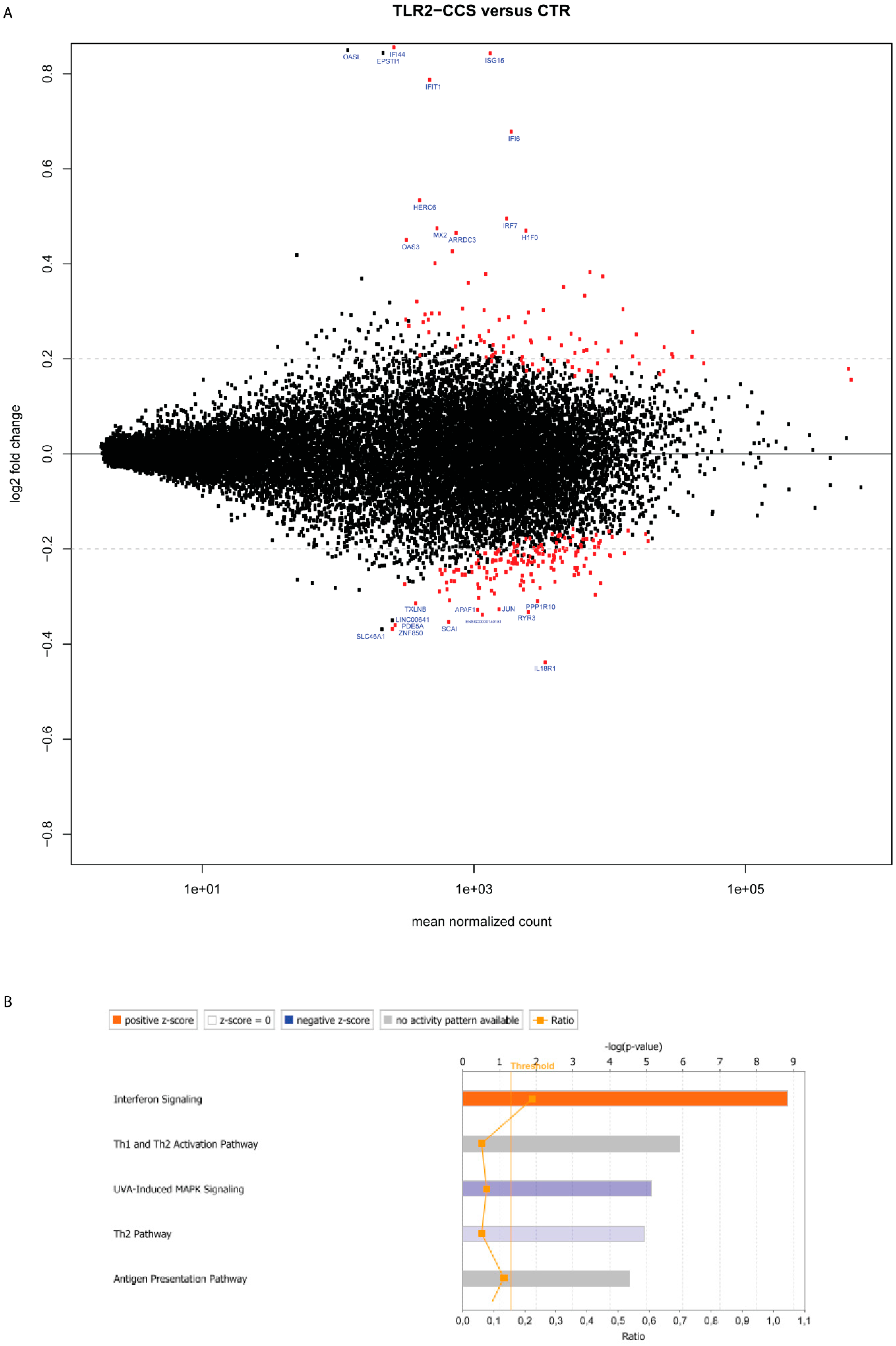
2.4. Deregulated Genes in TLR2-Primed MCs Cocultured with Colon Cancer Cells
2.5. Gene Deregulation for Extracellular Mediators in TLR2-Primed MCs Cocultured with Colon Cancer Cells
2.6. Gene Deregulation for Membrane Receptors in TLR2-Primed MCs Cocultured with Colon Cancer Cells
3. Discussion
4. Materials and Methods
4.1. Generation of Primary Human MCs
4.2. Colon Cancer Cell Lines
4.3. Colon Cancer Spheroids
4.4. Isolation and Purification of MCs
4.5. Transcriptome Analysis
4.6. Detecting Differential Gene Expression
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oldford, S.A.; Marshall, J.S. Mast cells as targets for immunotherapy of solid tumors. Mol. Immunol. 2015, 63, 113–124. [Google Scholar] [CrossRef]
- Varricchi, G.; Galdiero, M.R.; Loffredo, S.; Marone, G.; Iannone, R.; Granata, F. Are mast cells MASTers in Cancer? Front. Immunol. 2017, 8, 424. [Google Scholar] [CrossRef] [PubMed]
- Kalesnikoff, J.; Galli, S.J. New developments in mast cell biology. Nat. Immunol. 2008, 9, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Ko, E.A.; Sanders, K.M.; Zhou, T. A transcriptomic insight into the impacts of mast cells in lung, breast, and colon cancers. Oncoimmunology 2017, 6, e1360457. [Google Scholar] [CrossRef]
- Acikalin, M.F.; Oner, U.; Topcu, I.; Yasar, B.; Kiper, H.; Colak, E. Tumour angiogenesis and mast cell density in the prognostic assessment of colorectal carcinomas. Dig. Liver Dis. 2005, 37, 162–169. [Google Scholar] [CrossRef]
- Ammendola, M.; Rosario, S.; Giuseppe, S.; Giuseppe, D.; Severino, M.; Eustachio, R.; Rosa, P.; Ilaria, M.; Marica, C.; Angelo, V.; et al. Correlation between serum tryptase, mast cells positive to tryptase and microvascular density in colo-rectal cancer patients: Possible biological-clinical significance. PLoS ONE 2014, 9, e99512. [Google Scholar] [CrossRef] [PubMed]
- Gulubova, M.; Vlaykova, T. Prognostic significance of mast cell number and microvascular density for the survival of patients with primary colorectal cancer. J. Gastroenterol. Hepatol. 2009, 24, 1265–1275. [Google Scholar] [CrossRef]
- Malfettone, A.; Silvestris, N.; Saponaro, C.; Ranieri, G.; Russo, A.; Caruso, S.; Popescu, O.; Simone, G.; Paradiso, A.; Mangia, A. High density of tryptase-positive mast cells in human colorectal cancer: A poor prognostic factor related to protease-activated receptor 2 expression. J. Cell. Mol. Med. 2013, 17, 1025–1037. [Google Scholar] [CrossRef]
- Suzuki, S.; Ichikawa, Y.; Nakagawa, K.; Kumamoto, T.; Mori, R.; Matsuyama, R.; Takeda, K.; Ota, M.; Tanaka, K.; Tamura, T.; et al. High infiltration of mast cells positive to tryptase predicts worse outcome following resection of colorectal liver metastases. BMC Cancer 2015, 15, 840. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Linmei, L.; Renren, S.; Xueting, L.; Junyan, Z.; Zehong, Z.; Zhuofang, H.; Ailin, T. Mast cell targeted chimeric toxin can be developed as an adjunctive therapy in colon cancer treatment. Toxins (Basel) 2016, 8, 71. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.Y.; Fan, Y.; Luo, H.S.; Shen, Z.X.; Guo, Y.; Zhao, L.J. Prognostic significance of cell infiltrations of immunosurveillance in colorectal cancer. World J. Gastroenterol. 2005, 11, 1210–1214. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, H.J.; Hansen, U.; Chrostensen, I.J.; Reimert, C.M.; Brünner, N.; Moesgaard, F. Independent prognostic value of eosinophil and mast cell infiltration in colorectal cancer tissue. J. Pathol. 1999, 189, 487–495. [Google Scholar] [CrossRef]
- Visciano, C.; Liotti, F.; Prevete, N.; Cali, G.; Franco, R.; Collina, F.; de Paulis, A.; Marone, G.; Santoro, M.; Melillo, R.M. Mast cells induce epithelial-to-mesenchymal transition and stem cell features in human thyroid cancer cells through an IL-8-Akt-Slug pathway. Oncogene 2015, 34, 5175–5186. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Andrade, G.Y.; Ibarra-Sánchez, A.; González, D.; Lamas, M.; González-Espinosa, C. Immunoglobulin E induces VEGF production in mast cells and potentiates their pro-tumorigenic actions through a Fyn kinase-dependent mechanism. J. Hematol. Oncol. 2013, 6, 56. [Google Scholar] [CrossRef]
- Aoki, M.; Pawankar, R.; Niimi, Y.; Kawana, S. Mast cells in basal cell carcinoma express VEGF, IL-8 and RANTES. Int. Arch. Allergy Immunol. 2003, 130, 216–223. [Google Scholar] [CrossRef]
- Grützkau, A.; Krüger-Krasagakes, S.; Baumeister, H.; Schwarz, C.; Kögel, H.; Welker, P.; Lippert, U.; Henz, B.M.; Möller, A. Synthesis, storage, and release of vascular endothelial growth factor/vascular permeability factor (VEGF/VPF) by human mast cells: Implications for the biological significance of VEGF206. Mol. Biol. Cell 1998, 9, 875–884. [Google Scholar] [CrossRef]
- Coussens, L.M.; Raymond, W.W.; Bergers, G.; Laig-Webster, M.; Behrendtsen, O.; Werb, Z.; Caughey, G.H.; Hanahan, D. Inflammatory mast cells up-regulate angiogenesis during squamous epithelial carcinogenesis. Genes Dev. 1999, 13, 1382–1397. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Lei, Z.; Zhang, G.M.; Li, D.; Song, C.; Li, B.; Liu, Y.; Yuan, Y.; Unkeless, J.; Xiong, H.; et al. SCF-mediated mast cell infiltration and activation exacerbate the inflammation and immunosuppression in tumor microenvironment. Blood 2008, 112, 1269–1279. [Google Scholar] [CrossRef] [PubMed]
- Zaiss, D.M.; van Loosdregt, J.; Gorlani, A.; Bekker, C.P.; Gröne, A.; Sibilia, M.; van Bergen en Henegouwen, P.M.; Roovers, R.C.; Coffer, P.J.; Sijts, A.J. Amphiregulin enhances regulatory T cell-suppressive function via the epidermal growth factor receptor. Immunity 2013, 38, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Shikotra, A.; Ohri, C.M.; Green, R.H.; Waller, D.A.; Bradding, P. Mast cell phenotype, TNFα expression and degranulation status in non-small cell lung cancer. Sci. Rep. 2016, 6, 38352. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed]
- Schmetzer, O.; Valentin, P.; Smorodchenko, A.; Domenis, R.; Gri, G.; Siebenhaar, F.; Metz, M.; Maurer, M. A novel method to generate and culture human mast cells: Peripheral CD34+ stem cell-derived mast cells (PSCMCs). J. Immunol. Methods 2014, 413, 62–68. [Google Scholar] [CrossRef]
- Yu, Y.; Blokhuis, B.; Derks, Y.; Kumari, S.; Garssen, J.; Redegeld, F. Human mast cells promote colon cancer growth via bidirectional crosstalk: Studies in 2D and 3D coculture models. OncoImmunology 2018, 7, e1504729. [Google Scholar] [CrossRef] [PubMed]
- Said, A.H.; Raufman, J.P.; Xie, G. The role of matrix metalloproteinases in colorectal cancer. Cancers (Basel) 2014, 6, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Rymaszewski, A.L.; Tate, E.; Yimbesalu, J.P.; Gelman, A.E.; Jarzembowski, J.A.; Zhang, H.; Pritchard, K.A.; Vikis, H. The role of neutrophil myeloperoxidase in models of lung tumor development. Cancers (Basel) 2014, 6, 1111–1127. [Google Scholar] [CrossRef] [PubMed]
- de Neergaard, M.; Kim, J.; Villadsen, R.; Fridriksdottir, A.J.; Rank, F.; Timmermans-Wielenga, V.; Langerød, A.; Børresen-Dale, A.L.; Petersen, O.W.; Rønnov-Jessen, L. Epithelial-stromal interaction 1 (EPSTI1) substitutes for peritumoral fibroblasts in the tumor microenvironment. Am. J. Pathol. 2010, 176, 1229–1240. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.R.; Elodie, C.; Lisa, K.; Remy, N.; Simone, B.; Thierry, L.; Yves, A.; Jennifer, S.; François, R.; Bruno, G. Deregulation of rab and rab effector genes in bladder cancer. PLoS ONE 2012, 7, e39469. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; McQueen, J.; Cuschieri, L.; Vogel, J.; Measday, V. Spc24 and Stu2 promote spindle integrity when DNA replication is stalled. Mol. Biol. Cell 2007, 18, 2805–2816. [Google Scholar] [CrossRef]
- Musa, J.; Aynaud, M.-M.; Mirabeau, O.; Delattre, O.; Grünewald, T.G.P. MYBL2 (B-Myb): A central regulator of cell proliferation, cell survival and differentiation involved in tumorigenesis. Cell Death Dis. 2017, 8, e2895. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Bian, M.; Xin, G.; Deng, Z.; Luo, J.; Guo, X.; Chen, H.; Wang, Y.; Jiang, Q.; Zhang, C. TPX2 phosphorylation maintains metaphase spindle length by regulating microtubule flux. J. Cell Biol. 2015, 210, 373–383. [Google Scholar] [CrossRef]
- Sibilano, R.; Gaudenzio, N.; DeGorter, M.K.; Reber, L.L.; Hernandez, J.D.; Starkl, P.M.; Zurek, O.W.; Tsai, M.; Zahner, S.; Montgomery, S.B.; et al. A TNFRSF14-FcɛRI-mast cell pathway contributes to development of multiple features of asthma pathology in mice. Nat. Commun. 2016, 7, 13696. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.Z.; Upadhyay, V.; Prabhakar, B.; Maker, A.V. Shedding LIGHT (TNFSF14) on the tumor microenvironment of colorectal cancer liver metastases. J. Transl. Med. 2013, 11, 70. [Google Scholar] [CrossRef] [PubMed]
- Maker, A.V. Precise identification of immunotherapeutic targets for solid malignancies using clues within the tumor microenvironment—Evidence to turn on the LIGHT. OncoImmunology 2015, 5, e1069937. [Google Scholar] [CrossRef]
- Sainz, B., Jr.; Martín, B.; Tatari, M.; Heeschen, C.; Guerra, S. ISG15 is a critical microenvironmental factor for pancreatic cancer stem cells. Cancer Res. 2014, 74, 7309–7320. [Google Scholar] [CrossRef] [PubMed]
- Feng, C.; Beller, E.M.; Bagga, S.; Boyce, J.A. Human mast cells express multiple EP receptors for prostaglandin E2 that differentially modulate activation responses. Blood 2006, 107, 3243–3250. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Chan, C.M.; Wong, S.C.; Au, T.C.; Ho, W.S.; Chan, A.K.; Chan, A.S.; Ma, B.B.; Chan, A.T. Jagged 2 silencing inhibits motility and invasiveness of colorectal cancer cell lines. Oncol. Lett. 2016, 12, 5193–5198. [Google Scholar] [CrossRef] [PubMed]
- Holcombe, R.F.; Marsh, J.L.; Waterman, M.L.; Lin, F.; Milovanovic, T.; Truong, T. Expression of Wnt ligands and Frizzled receptors in colonic mucosa and in colon carcinoma. Mol. Pathol. 2002, 55, 220–226. [Google Scholar] [CrossRef]
- Mehlen, P.; Llambi, F. Role of netrin-1 and netrin-1 dependence receptors in colorectal cancers. Br. J. Cancer 2005, 93, 1–6. [Google Scholar] [CrossRef]
- Nowarski, R.; Jackson, R.; Gagliani, N.; de Zoete, M.R.; Palm, N.W.; Bailis, W.; Low, J.S.; Harman, C.C.; Graham, M.; Elinav, E.; et al. Epithelial IL-18 equilibrium controls barrier function in colitis. Cell 2015, 163, 1444–1456. [Google Scholar] [CrossRef]
- Sime, W.; Lunderius-Andersson, C.; Enoksson, M.; Rousselle, P.; Tryggvason, K.; Nilsson, G.; Harvima, I.; Patarroyo, M. Human mast cells adhere to and migrate on epithelial and vascular basement membrane laminins LM-332 and LM-511 via alpha3beta1 integrin. J. Immunol. 2009, 183, 4657–4665. [Google Scholar] [CrossRef]
- Edelson, B.T.; Li, Z.; Pappan, L.K.; Zutter, M.M. Mast cell-mediated inflammatory responses require the α2β1 integrin. Blood 2004, 103, 2214–2220. [Google Scholar] [CrossRef] [PubMed]
- Gurish, M.F.; Austen, K.F. Developmental origin and functional specialization of mast cell subsets. Immunity 2012, 37, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Dahlin, J.S.; Malinovschi, A.; Öhrvik, H.; Sandelin, M.; Janson, C.; Alving, K.; Hallgren, J. Lin-CD34hi CD117int/hi FcεRI+ cells in human blood constitute a rare population of mast cell progenitors. Blood 2016, 127, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Dubey, S.; Varney, M.L.; Dave, B.J.; Singh, R.K. IL-8 directly enhanced endothelial cell survival, proliferation, and matrix metalloproteinases production and regulated angiogenesis. J. Immunol. 2003, 170, 3369–3376. [Google Scholar] [CrossRef] [PubMed]
- Thirumangalakudi, L.; Yin, L.; Rao, H.V.; Grammas, P. IL-8 induces expression of matrix metalloproteinases, cell cycle and pro-apoptotic proteins, and cell death in cultured neurons. J. Alzheimers Dis. 2007, 11, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Iwanaga, K.; Nakamura, T.; Maeda, S.; Aritake, K.; Hori, M.; Urade, Y.; Ozaki, H.; Murata, T. Mast cell-derived prostaglandin D2 inhibits colitis and colitis-associated colon cancer in mice. Cancer Res. 2014, 74, 3011–3019. [Google Scholar] [CrossRef] [PubMed]
- Hawcroft, G.; Ko, C.W.; Hull, M.A. Prostaglandin E2-EP4 receptor signalling promotes tumorigenic behaviour of HT-29 human colorectal cancer cells. Oncogene 2007, 26, 3006–3019. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; DuBois, R.N. Inflammatory mediator prostaglandin E2 in colorectal cancer. Cancer J. 2013, 19, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Majid, R.M.; Marshall, J.S. Prostaglandin E2 induces degranulation-independent production of vascular endothelial growth factor by human mast cells. J. Immunol. 2004, 172, 1227–1236. [Google Scholar] [CrossRef] [PubMed]
- Detoraki, A.; Staiano, R.I.; Granata, F.; Giannattasio, G.; Prevete, N.; de Paulis, A.; Ribatti, D.; Genovese, A.; Triggiani, M.; Marone, G. Vascular endothelial growth factors synthesized by human lung mast cells exert angiogenic effects. J. Allergy Clin. Immunol. 2009, 123, 1142. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, L. Complexity of danger: The diverse nature of damage-associated molecular patterns. J. Biol. Chem. 2014, 289, 35237–35245. [Google Scholar] [CrossRef] [PubMed]
- Han, H.G.; Moon, H.W.; Jeon, Y.J. ISG15 in cancer: Beyond ubiquitin-like protein. Cancer Lett. 2018, 438, 52–62. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.D. ISG15: A double edged sword in cancer. Oncoimmunology 2015, 4, e1052935. [Google Scholar] [CrossRef] [PubMed]
- Reich, N.; Evans, B.; Levy, D.; Fahey, D.; Knight, E., Jr.; Darnell, J.E., Jr. Interferon-induced transcription of a gene encoding a 15-kDa protein depends on an upstream enhancer element. Proc. Natl. Acad. Sci. USA 1987, 84, 6394–6398. [Google Scholar] [CrossRef]
- Lotfi-Emran, S.; Ward, B.R.; Le, Q.T.; Pozez, A.L.; Manjili, M.H.; Woodfolk, J.A.; Schwartz, L.B. Human mast cells present antigen to autologous CD4 + T cells. J. Allergy Clin. Immunol. 2018, 141, 311–321.e10. [Google Scholar] [CrossRef] [PubMed]
- Kambayashi, T.; Allenspach, E.J.; Chang, J.T.; Zou, T.; Shoag, J.E.; Reiner, S.L.; Caton, A.J.; Koretzky, G.A. Inducible MHC class II expression by mast cells supports effector and regulatory T cell activation. J. Immunol. 2009, 182, 4686–4695. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Gaudenzio, N. Human mast cells as antigen-presenting cells: When is this role important in vivo? J. Allergy Clin. Immunol. 2018, 141, 92–93. [Google Scholar] [CrossRef]
- Gessani, S.; Conti, L.; Del Cornò, M.; Belardelli, F. Type I interferons as regulators of human antigen presenting cell functions. Toxins 2014, 6, 1696–1723. [Google Scholar] [CrossRef] [PubMed]
- Qiao, G.; Qin, J.; Kunda, N.; Calata, J.F.; Mahmud, D.L.; Gann, P.; Fu, Y.X.; Rosenberg, S.A.; Prabhakar, B.S.; Maker, A.V. LIGHT elevation enhances immune eradication of colon cancer metastases. Cancer Res. 2017, 77, 1880–1891. [Google Scholar] [CrossRef]
- Timmins, N.E.; Nielsen, L.K. Generation of multicellular tumor spheroids by the hanging-drop method. Methods Mol. Med. 2007, 140, 141–151. [Google Scholar] [PubMed]
- Berens, E.B.; Holy, J.M.; Riegel, A.T.; Wellstein, A. A cancer cell spheroid assay to assess invasion in a 3D Setting. J. Vis. Exp. 2015, e53409. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Name | Gene Expression ‡ | Log2FC | Adjusted p Value |
|---|---|---|---|---|
| upregulated | ||||
| SLC45A4 | Solute carrier family 45 member 4 | 1.07 | 0.57 | **** |
| NAB2 | Nuclear polyadenylated RNA-binding protein | 40.76 | 0.55 | **** |
| MPO | Myeloperoxidase | 0.49 | 0.49 | **** |
| RFLNB | Refilin-B | 18.00 | 0.46 | **** |
| SPRY2 | Protein sprouty homolog 2 | 19.00 | 0.43 | **** |
| OASL | 2′-5′-oligoadenylate synthase-like protein | 0.21 | 0.43 | **** |
| EPSTI1 | Epithelial-stromal interaction protein 1 | 0.39 | 0.42 | **** |
| MMP2 | Matrix metalloproteinase-2 | 3.23 | 0.39 | **** |
| UNC5B | Netrin receptor UNC5B | 0.20 | 0.38 | **** |
| PHLDA1 | Pleckstrin homology-like domain family A member 1 | 0.54 | 0.36 | **** |
| downregulated | ||||
| CDT1 | DNA replication factor | 3.34 | −0.57 | **** |
| KIF20A | Kinesin-like protein KIF20A | 3.54 | −0.53 | **** |
| ESPL1 | Separin | 1.27 | −0.53 | **** |
| SPC24 | Kinetochore protein Spc24 | 2.20 | −0.53 | **** |
| MYBL2 | Myb-related protein B | 2.33 | −0.52 | **** |
| KIF18B | Kinesin-like protein KIF18B | 1.66 | −0.51 | **** |
| FAM111B | Protein FAM111B | 0.68 | −0.51 | **** |
| TPX2 | Targeting protein for Xklp2 | 7.41 | −0.50 | **** |
| IQGAP3 | Ras GTPase-activating-like protein IQGAP3 | 1.59 | −0.50 | **** |
| PKMYT1 | Membrane-associated tyrosine- and threonine-specific cdc2-inhibitory kinase | 1.88 | −0.50 | **** |
| Gene Symbol | Name | Cytolocation | Gene Expression * | Log2FC | Adjusted p Value |
|---|---|---|---|---|---|
| MMP2 | Matrix metalloproteinase-2 | Extracellular | 3.23 | 0.39 | **** |
| VEGFA | Vascular endothelial growth factor A | Extracellular | 2.51 | 0.23 | * |
| PDGFA | Platelet-derived growth factor subunit A | Extracellular | 3.84 | 0.29 | ** |
| TNFSF14 | Tumor necrosis factor ligand superfamily member 14 | Extracellular | 2.04 | 0.31 | ** |
| ISG15 | Interferon-stimulated gene 15 | Extracellular | 25.01 | 0.34 | ** |
| NPTX1 | Neuronal pentraxin-1 | Extracellular | 1.66 | −0.38 | *** |
| PTGS2 | Prostaglandin G/H synthase 2 | Cytoplasm | 1.42 | 0.35 | ** |
| RHOB | Rho-related GTP-binding protein RhoB | Cytoplasm | 35.05 | −0.22 | * |
| CIT | Citron Rho-interacting kinase | Cytoplasm | 1.32 | −0.37 | *** |
| RELA | Nuclear factor NF-kappa-B p65 subunit | Nucleus | 20.11 | 0.24 | * |
| STAT1 | Signal transducer and activator of transcription 1-alpha/beta | Nucleus | 14.78 | 0.27 | ** |
| NOTCH1 | Neurogenic locus notch homolog protein 1 | Membrane | 10.18 | 0.35 | **** |
| PTGER4 | Prostaglandin E2 receptor EP4 subtype | Membrane | 1.16 | 0.29 | * |
| FZD1 | Frizzled-1 | Membrane | 3.09 | 0.32 | * |
| UNC5B | Netrin receptor UNC5B | Membrane | 0.20 | 0.38 | *** |
| IL18RAP | Interleukin-18 receptor accessory protein | Membrane | 0.56 | 0.25 | * |
| P2RY11 | P2Y purinoceptor 11 | Membrane | 2.06 | 0.33 | ** |
| ITGA3 | Integrin alpha-3 | Membrane | 14.56 | −0.21 | ** |
| ITGA2 | Integrin alpha-2 | Membrane | 1.19 | −0.27 | * |
| HMMR | Hyaluronan mediated motility receptor | Membrane | 2.19 | −0.44 | **** |
| TJP2 | Tight junction protein ZO-2 | Membrane | 11.75 | −0.22 | * |
| Gene Symbol | Name | Gene Expression ‡ | Log2FC | Adjusted p Value |
|---|---|---|---|---|
| upregulated | ||||
| IFI44 | Interferon-induced protein 44 | 0.90 | 0.86 | **** |
| ISG15 | Interferon-stimulated gene 15 | 25.01 | 0.84 | **** |
| IFIT1 | Interferon-induced protein with tetratricopeptide repeats 1 | 2.05 | 0.79 | **** |
| IFI6 | Interferon alpha-inducible protein 6 | 14.49 | 0.68 | **** |
| HERC6 | Probable E3 ubiquitin-protein ligase HERC6 | 1.24 | 0.53 | **** |
| IRF7 | Interferon regulatory factor 7 | 13.80 | 0.49 | **** |
| H1F0 | Histone H1.0 | 21.75 | 0.47 | **** |
| MX2 | Interferon-induced GTP-binding protein Mx2 | 0.79 | 0.47 | **** |
| ARRDC3 | Arrestin domain-containing protein 3 | 3.42 | 0.46 | **** |
| OAS3 | 2′-5′-oligoadenylate synthase 3 | 0.07 | 0.45 | **** |
| downregulated | ||||
| IL18R1 | Interleukin-18 receptor 1 | 26.24 | −0.44 | **** |
| ZNF850 | Zinc finger protein 850 | 1.10 | −0.37 | ** |
| PDE5A | cGMP-specific 3′,5′-cyclic phosphodiesterase | 0.98 | −0.36 | ** |
| SCAI | Protein SCAI | 1.75 | −0.35 | *** |
| ENSG00000140181 | N/A | 5.13 | −0.34 | *** |
| APAF1 | Apoptotic protease-activating factor 1 | 4.32 | −0.33 | ** |
| JUN | Transcription factor AP-1 | 16.81 | −0.33 | ** |
| RYR3 | Ryanodine receptor 3 | 5.47 | −0.33 | **** |
| PPP1R10 | Serine/threonine-protein phosphatase 1 regulatory subunit 10 | 17.85 | −0.31 | **** |
| TXLNB | Beta-taxilin | 2.86 | −0.31 | * |
| Gene Symbol | Name | Cytolocation | Gene Expression * | Log2FC | Adjusted p Value |
|---|---|---|---|---|---|
| ISG15 | Interferon-stimulated gene 15 | Extracellular | 25.01 | 0.84 | **** |
| MMP2 | Matrix metalloproteinase-2 | Extracellular | 3.23 | 0.31 | ** |
| TNFSF14 | Tumor necrosis factor ligand superfamily member 14 | Extracellular | 2.04 | 0.27 | * |
| NDST2 | Bifunctional heparan sulfate N-deacetylase/N-sulfotransferase 2 | Cytoplasm | 8.66 | −0.21 | * |
| ALOX5 | Arachidonate 5-lipoxygenase | Nucleus | 35.70 | 0.21 | * |
| IRF7 | Interferon regulatory factor 7 | Nucleus | 13.79 | 0.49 | **** |
| STAT1 | Signal transducer and activator of transcription 1-alpha/beta | Nucleus | 14.78 | 0.35 | **** |
| HLA-B | HLA class I histocompatibility antigen, B-51 alpha chain | Membrane | 127.04 | 0.25 | ** |
| HLA-C | HLA class I histocompatibility antigen, Cw-12 alpha chain | Membrane | 148.74 | 0.21 | * |
| HLA-DPA1 | HLA class II histocompatibility antigen, DP alpha 1 chain | Membrane | 6.69 | 0.21 | * |
| HLA-DRA | HLA class II histocompatibility antigen, DR alpha chain | Membrane | 4.57 | 0.28 | * |
| CD74 | HLA class II histocompatibility antigen gamma chain | Membrane | 17.18 | 0.30 | *** |
| ITGB7 | Integrin beta-7 | Membrane | 7.11 | 0.23 | * |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Y.; Blokhuis, B.R.; Garssen, J.; Redegeld, F.A. A Transcriptomic Insight into the Impact of Colon Cancer Cells on Mast Cells. Int. J. Mol. Sci. 2019, 20, 1689. https://doi.org/10.3390/ijms20071689
Yu Y, Blokhuis BR, Garssen J, Redegeld FA. A Transcriptomic Insight into the Impact of Colon Cancer Cells on Mast Cells. International Journal of Molecular Sciences. 2019; 20(7):1689. https://doi.org/10.3390/ijms20071689
Chicago/Turabian StyleYu, Yingxin, Bart R. Blokhuis, Johan Garssen, and Frank A. Redegeld. 2019. "A Transcriptomic Insight into the Impact of Colon Cancer Cells on Mast Cells" International Journal of Molecular Sciences 20, no. 7: 1689. https://doi.org/10.3390/ijms20071689
APA StyleYu, Y., Blokhuis, B. R., Garssen, J., & Redegeld, F. A. (2019). A Transcriptomic Insight into the Impact of Colon Cancer Cells on Mast Cells. International Journal of Molecular Sciences, 20(7), 1689. https://doi.org/10.3390/ijms20071689