Miscanthus x giganteus Stem Versus Leaf-Derived Lignins Differing in Monolignol Ratio and Linkage


 , , ,
, , , 
Abstract

1. Introduction
2. Results and Discussion
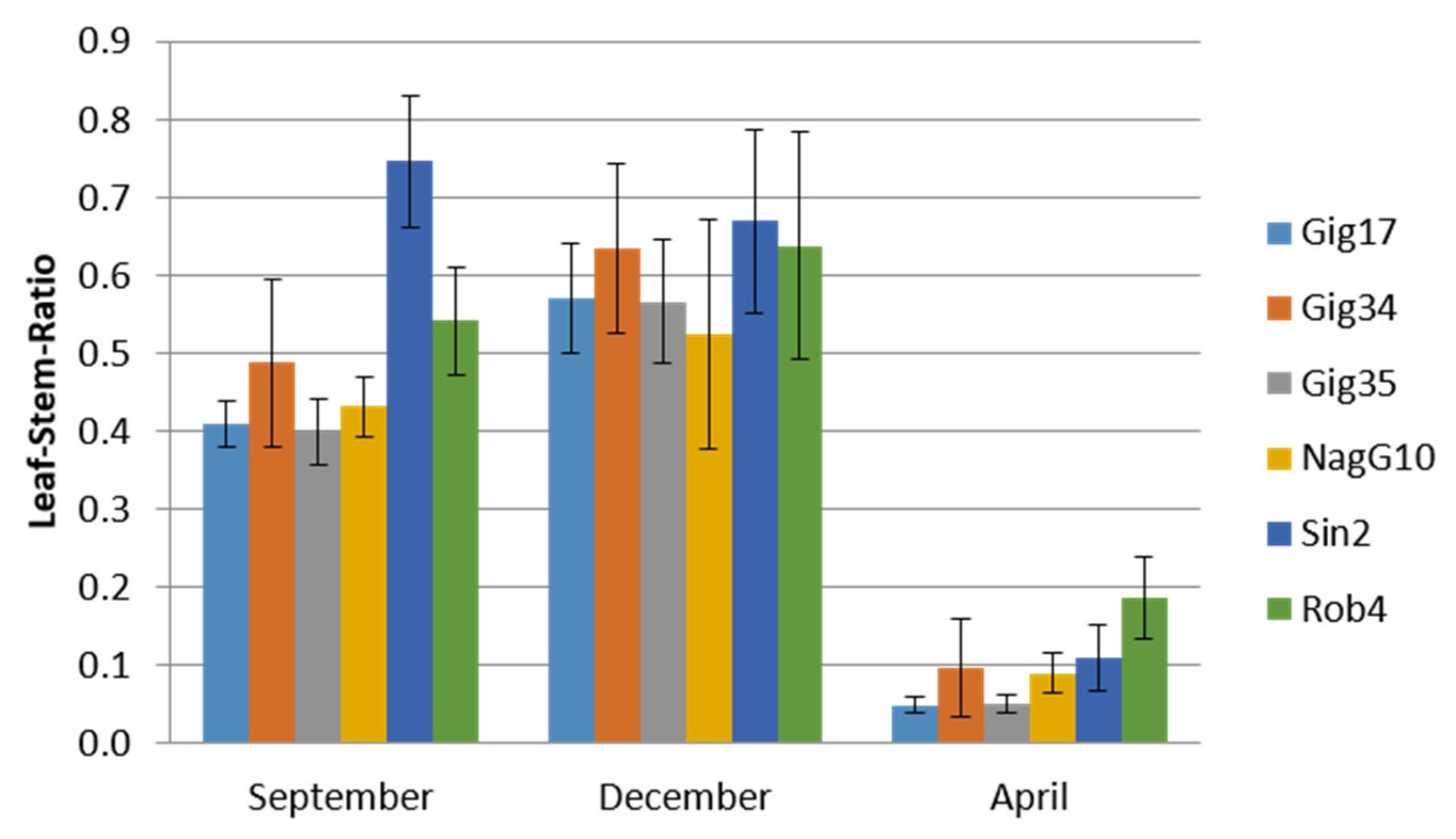
2.1. Miscanthus Crop Analysis
2.2. NREL Compositional Analysis
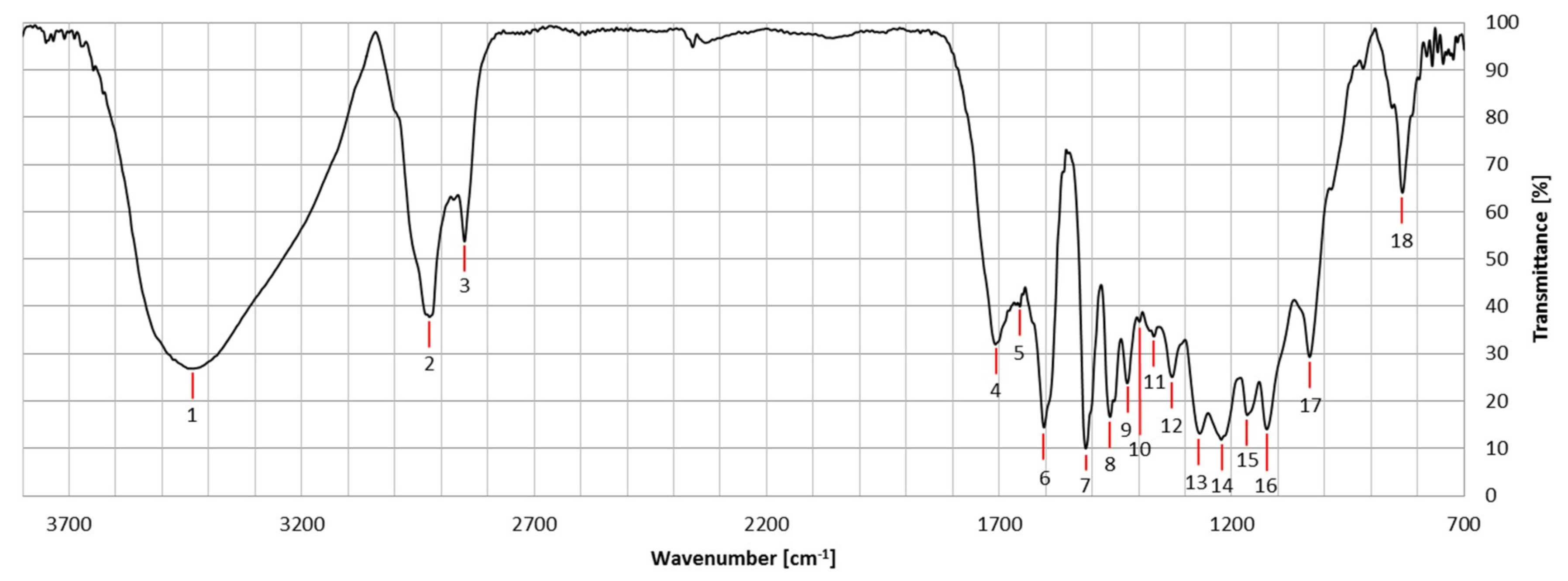
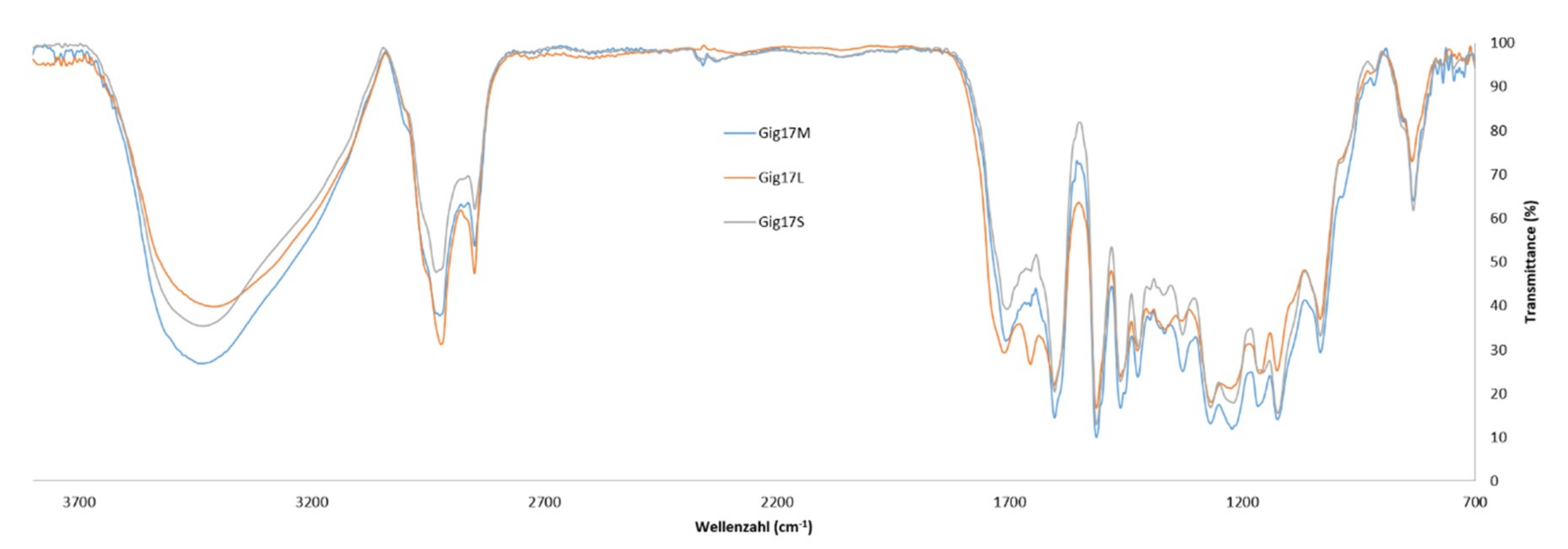
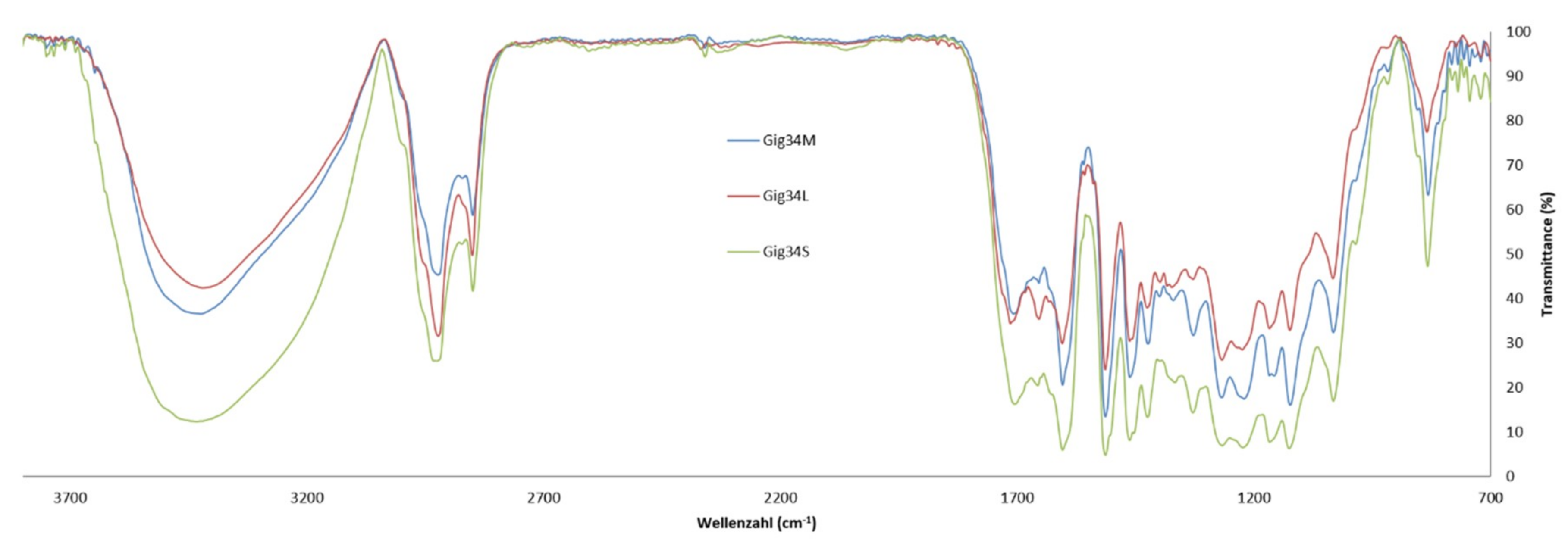
2.3. FTIR Spectroscopy
2.4. Size Exclusion Chromatography
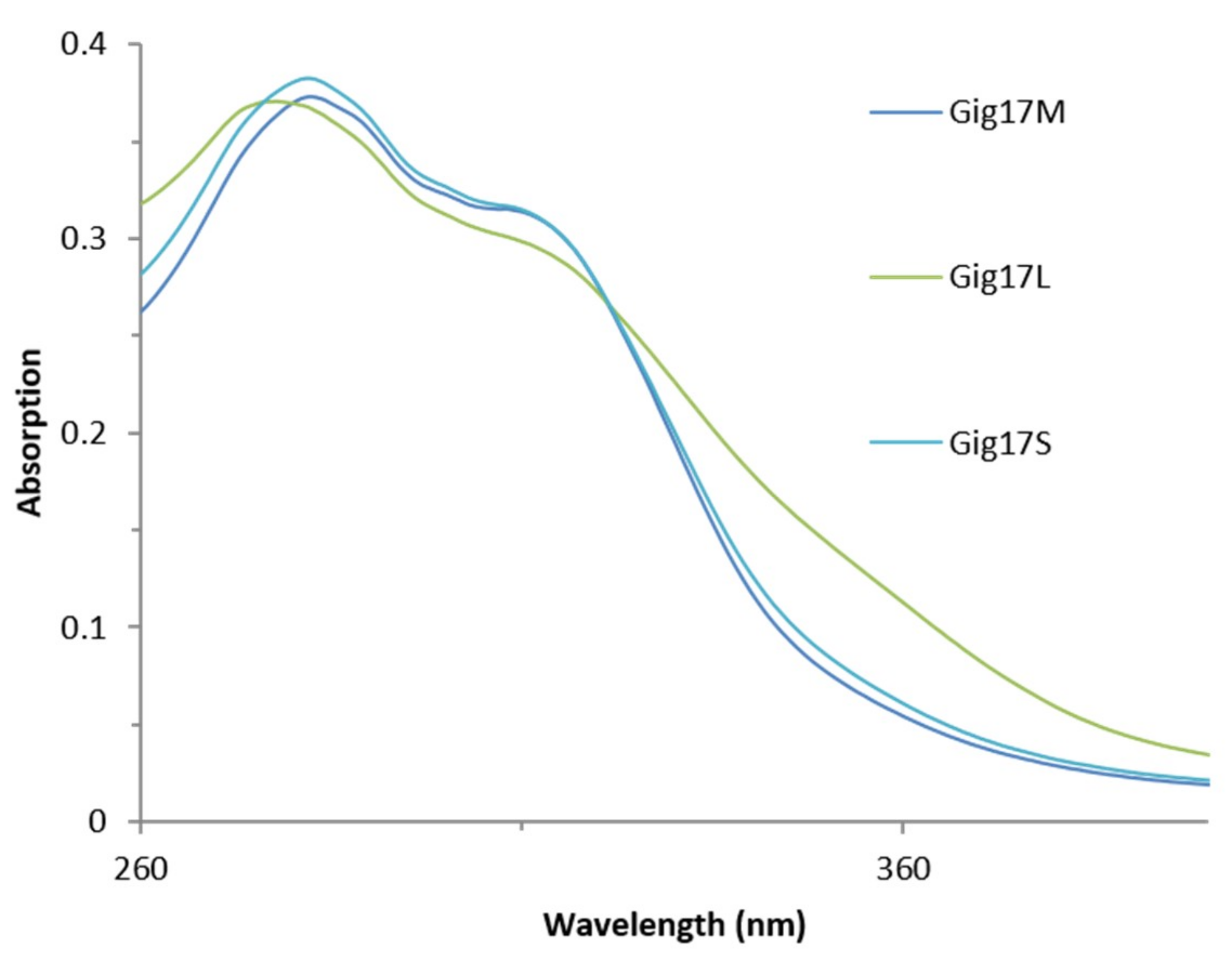
2.5. UV-Vis Spectroscopy
2.6. Thermogravimetric Analysis
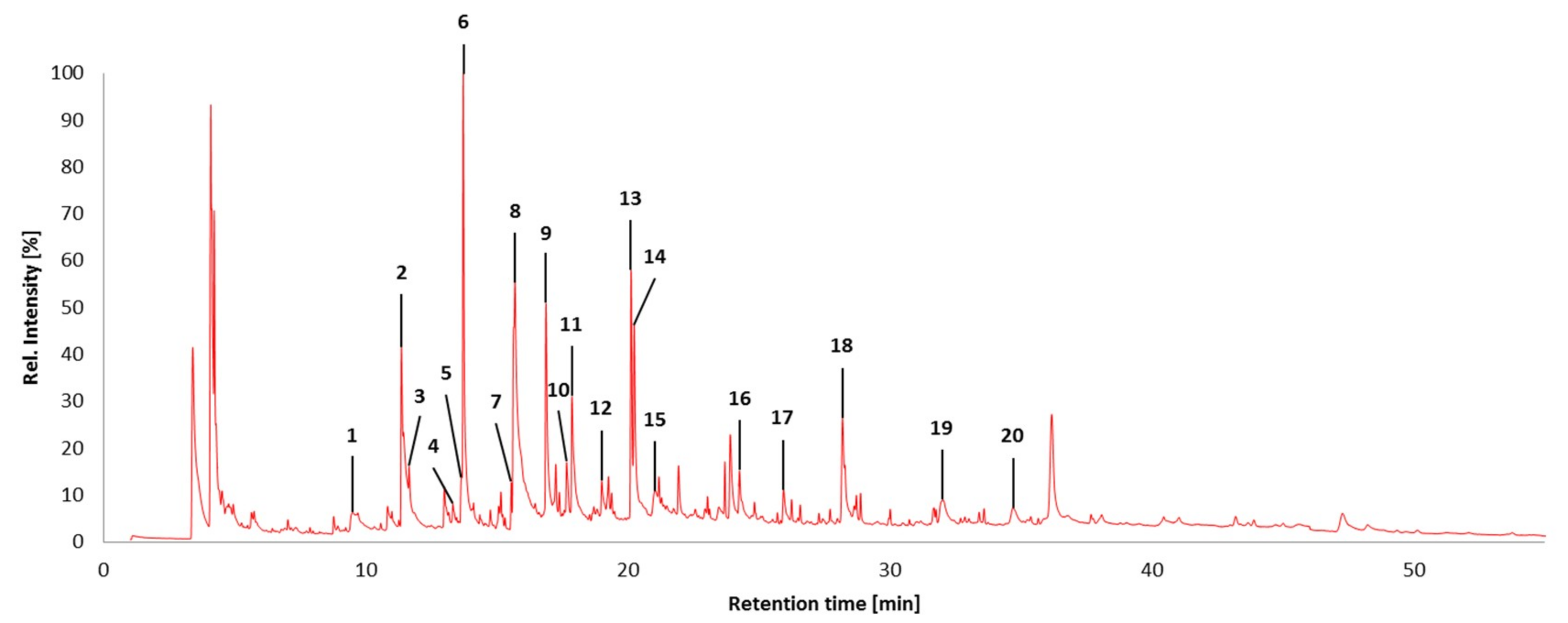
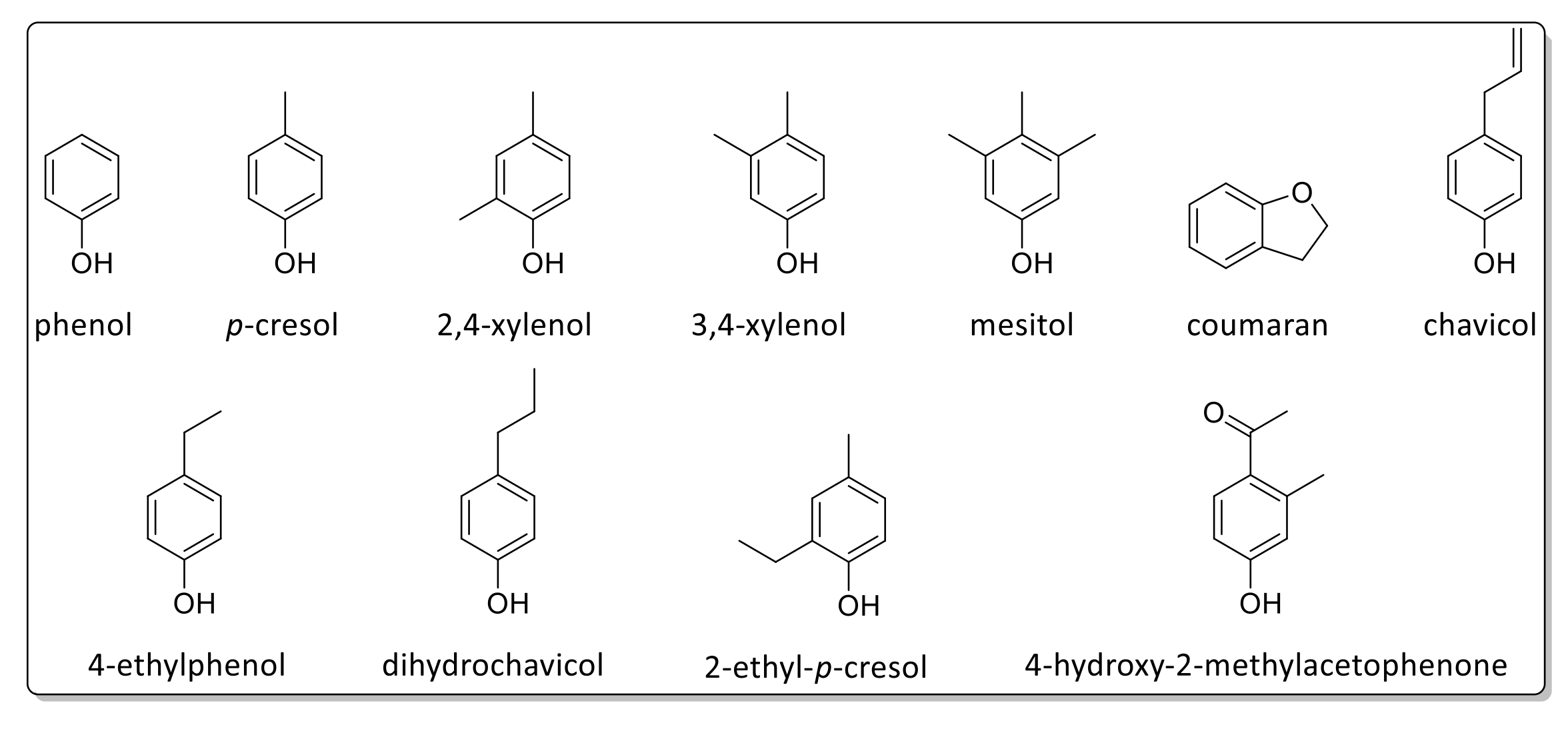
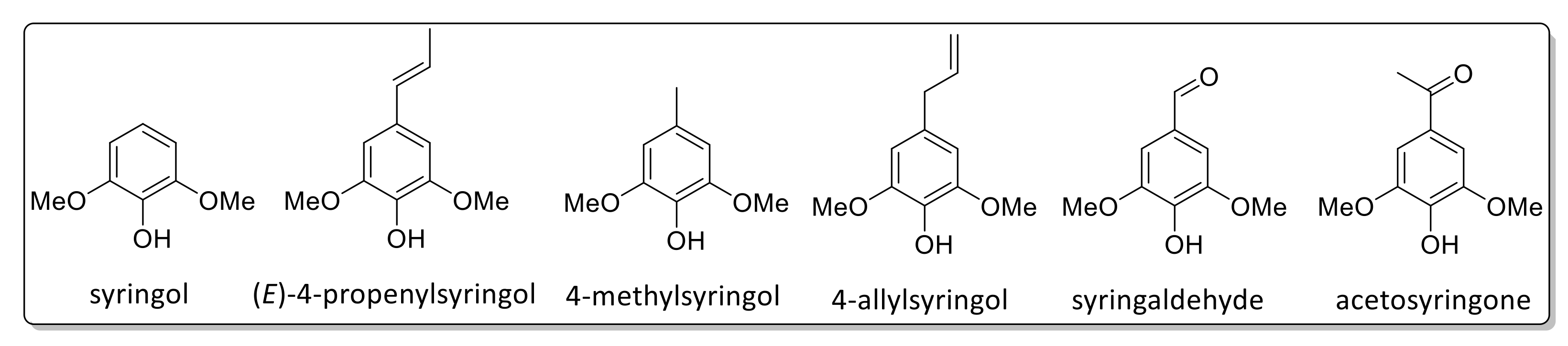
2.7. Pyrolysis Gas Chromatography/Mass Spectrometry
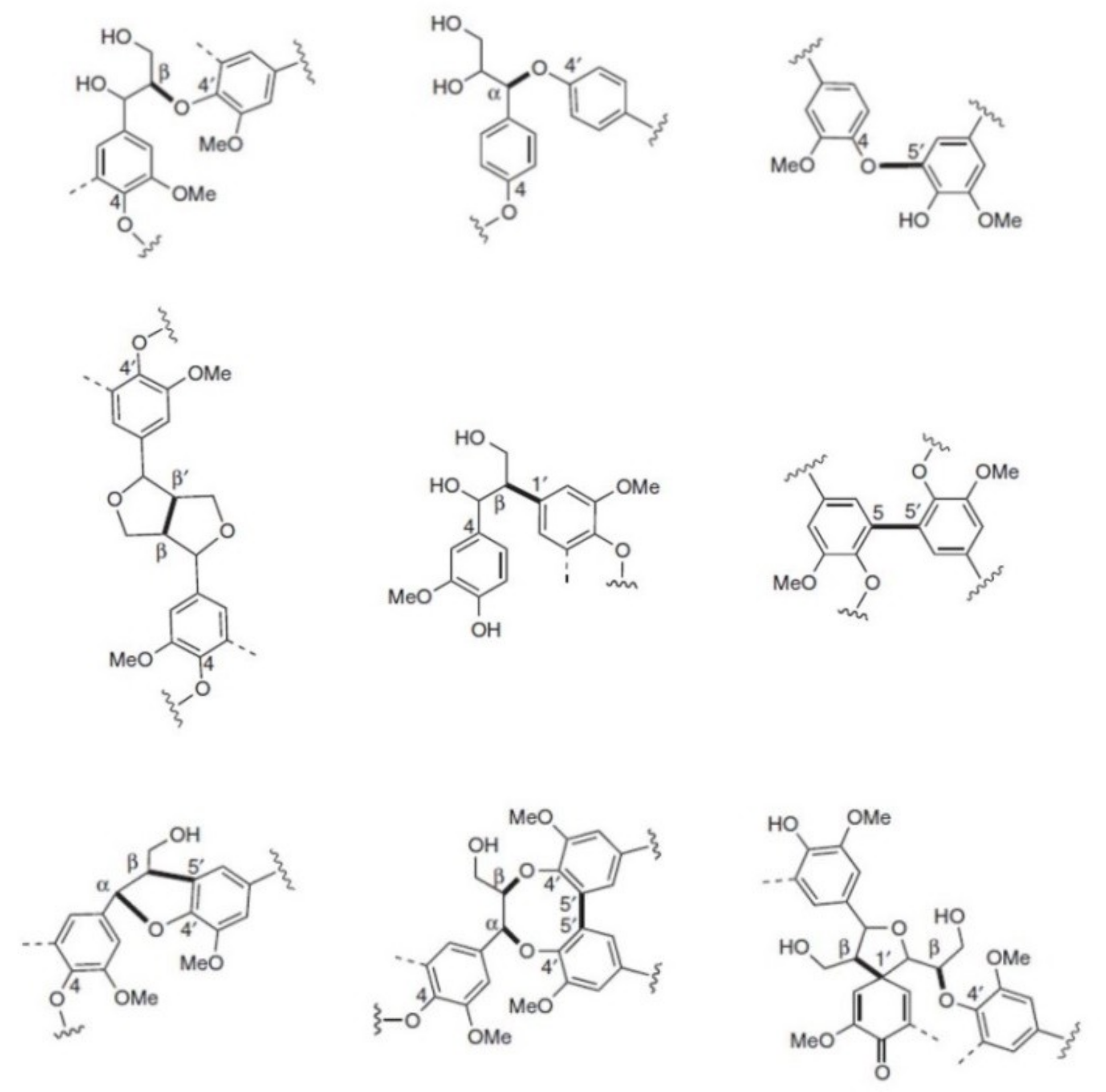
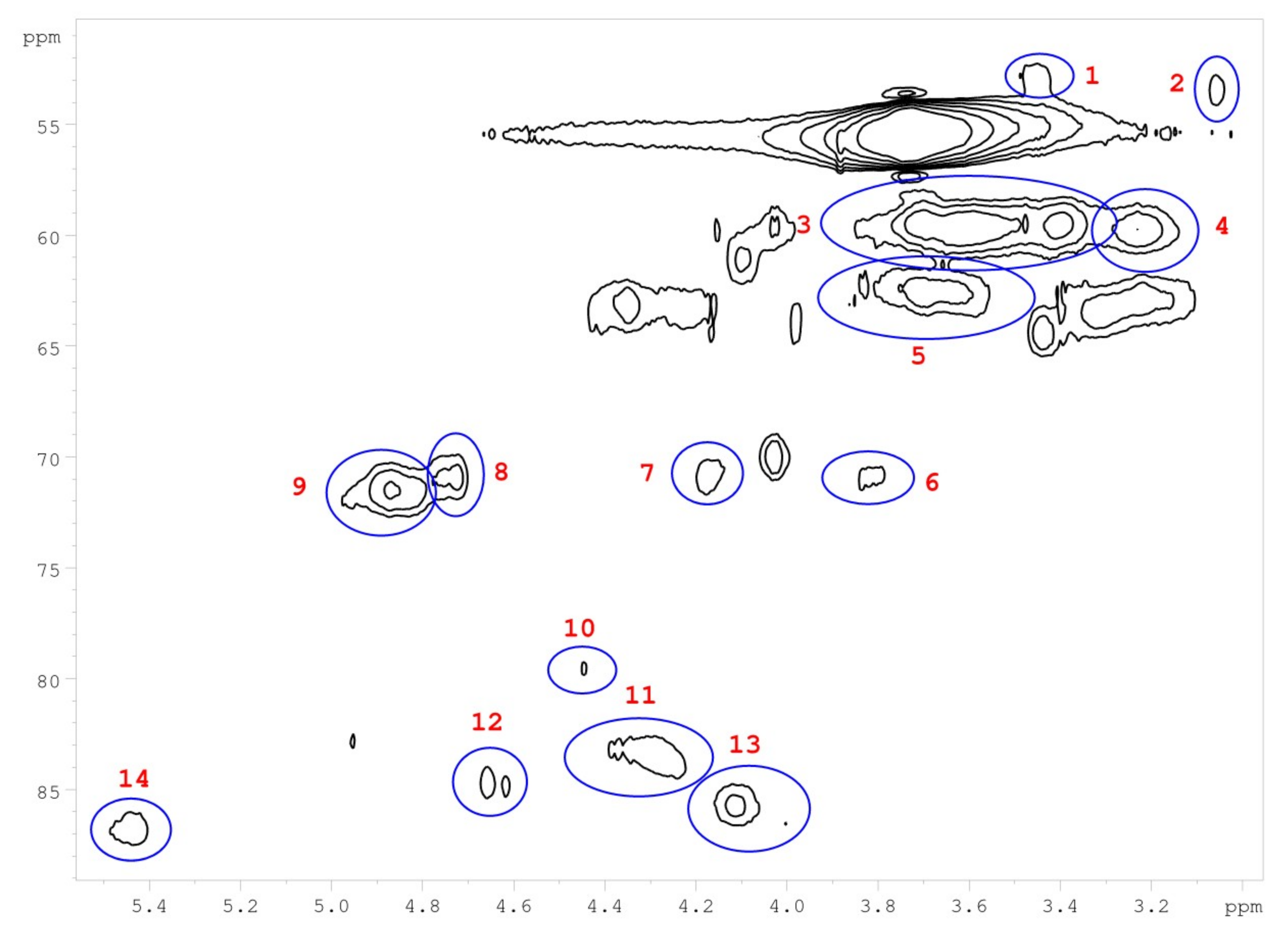
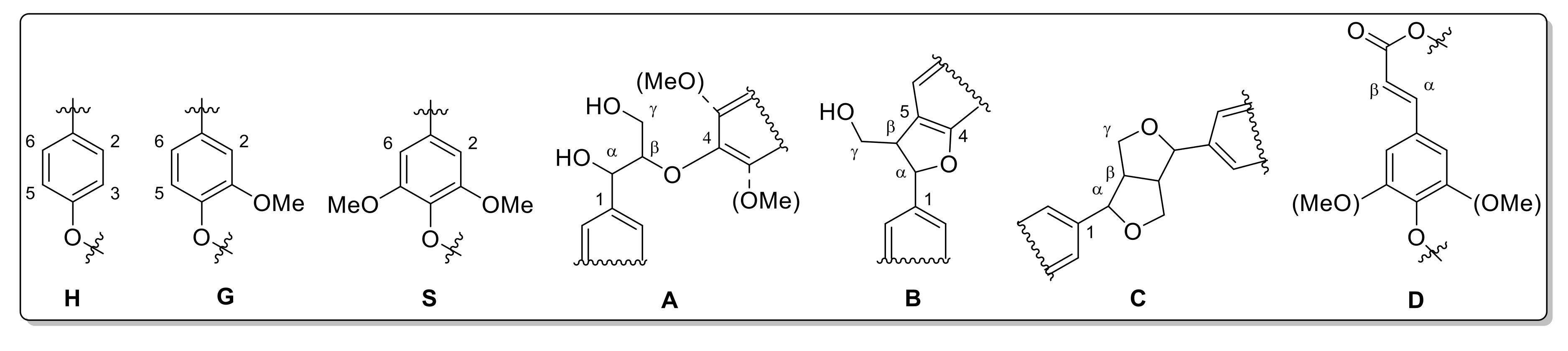
2.8. Nuclear Magnetic Resonance
3. Materials and Methods
3.1. Miscanthus Field Trial Lignin Isolation via Organosolv Pulping
3.2. Lignin Isolation via Organosolv Pulping
3.3. Lignin Purity, Ash, and Sugar Content via NREL Measurements
3.4. Size-Exclusion Chromatography
3.5. UV-Vis Spectroscopy
3.6. Fourier Transform Infrared Spectroscopy
3.7. Multivariate Data Analysis
3.8. Nuclear Magnetic Resonance Spectroscopy
3.9. Thermogravimetric Analysis
3.10. Pyrolysis-Gas Chromatography/Mass Spectrometry
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Emmerling, C.; Pude, R. Introducing Miscanthus to the greening measures of the EU Common Agricultural Policy. GCB Bioenergy 2017, 9, 274–279. [Google Scholar] [CrossRef]
- Lewandowski, I.; Clifton-Brown, J.; Trindade, L.M.; van der Linden, G.C.; Schwarz, K.-U.; Müller-Sämann, K.; Anisimov, A.; Chen, C.-L.; Dolstra, O.; Donnison, I.S.; et al. Progress on optimizing miscanthus biomass production for the European bioeconomy: Results of the EU FP7 project OPTIMISC. Front. Plant Sci. 2016, 7, 1620. [Google Scholar] [CrossRef] [PubMed]
- McCalmont, J.P.; Hastings, A.; McNamara, N.P.; Richter, G.M.; Robson, P.; Donnison, I.S.; Clifton-Brown, J. Environmental costs and benefits of growing Miscanthus for bioenergy in the UK. GCB Bioenergy 2017, 9, 489–507. [Google Scholar] [CrossRef] [PubMed]
- Kellogg, E.A. C4 photosynthesis. Curr. Biol. 2013, 23, R594–R599. [Google Scholar] [CrossRef] [PubMed]
- Villaverde, J.J.; Ligero, P.; Vega, A. Miscanthus x giganteus as a Source Of Biobased Products Through Organosolv Fractionation: A Mini Review. TOAJ 2010, 4, 102–110. [Google Scholar] [CrossRef]
- Teese, P. Intraspecific variation for CO2 compensation point and differential growth among variants in a C3-C4 intermediate plant. Oecologia 1995, 102, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Felten, D.; Fr€oba, N.; Fries, J.; Emmerling, C. Energy balances and CO2-mitigation potentials of bioenergy crops (Miscanthus, rapeseed, maize) based on farming conditions in Western Germany. Renew. Energy 2013, 55, 160–174. [Google Scholar] [CrossRef]
- Felten, D.; Emmerling, C. Accumulation of Miscanthus-derived carbon in soils in relation to soil depth and duration of land-use under commercial farming conditions. J. Plant Nutr. Soil Sci. 2012, 175, 661–670. [Google Scholar] [CrossRef]
- Kiesel, A.; Nunn, C.; Iqbal, Y.; Van der Weijde, T.; Wagner, M.; Özgüven, M.; Tarakanov, I.; Kalinina, O.; Trindade, L.M.; Clifton-Brown, J.; et al. Site-Specific Management of Miscanthus Genotypes for Combustion and anaerobic digestion: A comparison of energy yields. Front. Plant Sci. 2017, 8, 347. [Google Scholar] [CrossRef]
- Brosse, N.; Dufour, A.; Meng, X.; Sun, Q.; Ragauskas, A. Miscanthus: A fast growing crop for biofuels and chemicals production. Biofuels Bioprod. Biorefin. 2012, 6, 580–598. [Google Scholar] [CrossRef]
- Jørgensen, U. Genotypic variation in dry matter accumulation and content of N, K and Cl in Miscanthus in Denmark. Biomass Bioenergy 1997, 12, 155–169. [Google Scholar] [CrossRef]
- Clifton-Brown, J.C.; Lewandowski, I. Frosttoleranz der Rhizome verschiedener Miscanthus Genotypen. Mitteilungen der Gesellschaft fuer Pflanzenbauwissenschaften 1998, 11, 225–226. [Google Scholar]
- Eppel-Hotz, A.; Jodl, S.; Kuhn, W.; Marzini, K.; Munzer, W. Miscanthus: New cultivars and results of research experiments for improving the establishment rate. In Biomass for Energy. In Proceedings of the 10th European Biomass Conference, Würzburg, Germany, 8–11 July 1998; pp. 780–786. [Google Scholar]
- Greef, J.M.; Deuter, M.; Jung, C.; Schondelmaier, J. Genetic diversity of European Miscanthus species revealed by AFLP fingerprinting. Genet. Resour. Crop Evol. 1997, 44, 185–195. [Google Scholar] [CrossRef]
- Pude, R.; Treseler, C.H.; Noga, G. Morphological, chemical and technical parameters of Miscanthus genotypes. J. Appl. Bot. 2004, 78, 58–63. [Google Scholar]
- Pude, R.; Treseler, C.H.; Trettin, R.; Noga, G. Suitability of Miscanthus genotypes for lightweight concrete. Die Bodenkultur 2005, 56, 61–69. [Google Scholar]
- Dahmen, N.; Lewandowski, I.; Zibek, S.; Weidtmann, A. Integrated lignocellulosic value chains in a growing bioeconomy: Status quo and perspectives. GCB Bioenerg. 2019, 11, 107–117. [Google Scholar] [CrossRef]
- Clifton-Brown, J.; Breuer, J.; Jones, M.B. Carbon mitigation by the energy crop Miscanthus. Glob. Chang. Biol. 2007, 13, 2296–2307. [Google Scholar] [CrossRef]
- Clifton-Brown, J.C.; Lewandowski, I.; Andersson, B.; Basch, G.; Christian, D.G.; Kjeldsen, J.B.; Jørgensen, U.; Mortensene, J.V.; Riched, A.B.; Schwarz, K.U.; et al. Performance of 15 Miscanthus Genotypes at Five Sites in Europe. Agron. J. 2011, 93, 1013–1019. [Google Scholar] [CrossRef]
- Kamm, B.; Kamm, M.; Hirth, T.; Schulze, M. Lignocelluloses Based Chemical Products and Product Family Trees. In Biorefineries-Industrial Processes and Products; Kamm, M., Kamm, B., Gruber, P.C., Eds.; Wiley-VCH: Weinheim, Germany, 2006; pp. 97–150. ISBN 3-527-31027-4. [Google Scholar]
- Kamm, B.; Gruber, P.R.; Kamm, M. Biorefineries-Industrial Processes and Products. In Ullmann’s Encyclopedia of Industrial Chemistry; Wiley-VCH: Weinheim, Germany, 2016; ISBN 9783527306732. [Google Scholar]
- Alzagameem, A.; El Khaldi-Hansen, B.; Kamm, B.; Schulze, M. Lignocellulosic biomass for energy, biofuels, biomaterials, and chemicals. In Biomass and Green Chemistry, 1st ed.; Vaz, S., Jr., Ed.; Springer International Publishing: Basel, Switzerland, 2018; pp. 95–132. ISBN 978-3-319-66736-2. [Google Scholar] [CrossRef]
- Kraska, T.; Kleinschmidt, B.; Weinand, J.; Pude, R. Cascading use of Miscanthus as growing substrate in soilless cultivation of vegetables (tomatoes, cucumbers) and subsequent direct combustion. Sci. Hortic. 2018, 235, 205–213. [Google Scholar] [CrossRef]
- Winzer, F.; Kraska, T.; Elsenberger, C.; Kötter, T.; Pude, R. Biomass from fruit trees for combined energy and food production. Biomass Bioenergy 2017, 107, 279–286. [Google Scholar] [CrossRef]
- Hemansi Gupta, R.; Yadav, G.; Kumar, G.; Yadav, A.; Saini, J.K.; Kuhad, R.C. Second Generation Bioethanol Production: The State of Art. In Sustainable Approaches for Biofuels Production Technologies. Biofuel and Biorefinery Technologies; Srivastava, N., Srivastava, M., Mishra, P., Upadhyay, S., Ramteke, P., Gupta, V., Eds.; Springer International Publishing: Heidelberg, Germany, 2019; Volume 7, pp. 121–146. [Google Scholar]
- de Vrije, T.; de Haas, G.G.; Tan, G.B.; Keijsers, E.R.P.; Claassen, P.A.M. Pretreatment of Miscanthus for hydrogen production by Thermotoga elfii. Int. J. Hydrogen Energy 2002, 27, 1381–1390. [Google Scholar] [CrossRef]
- Kjeldsen, J.B.; Joergensen, U.; Kristensen, E.F. Thatching: Use of Miscanthus; DJF Rapport; Danmarks Jordbrugsforskning: Foulum, Denmark, 1999. [Google Scholar]
- Wagenaar, B.M.; van den Heuvel, E. Co-combustion of Miscanthus in a pulverised coal combustor: Experiments in a droptube furnace. Biomass Bioenergy 1997, 12, 185–197. [Google Scholar] [CrossRef]
- Khelfa, A.; Sharypov, V.; Finqueneisel, G.; Weber, J.V. Catalytic pyrolysis and gasification of Miscanthus Giganteus: Haematite (Fe2O3) a versatile catalyst. J. Anal. Appl. Pyrolysis 2009, 84, 84–88. [Google Scholar] [CrossRef]
- Velasquez, J.A.; Ferrando, F.; Farriol, X.; Salvado, J. Binderless fiberboard from steam exploded Miscanthus sinensis. Wood Sci. Technol. 2003, 37, 269–278. [Google Scholar] [CrossRef]
- Vega, A.; Bao, M.; Lamas, J. Application of factorial design to the modelling of organosolv delignification of Miscanthus sinensis (elephant grass) with phenol and dilute acid solutions. Bioresour. Technol. 1997, 61, 1–7. [Google Scholar] [CrossRef]
- El Hage, R.; Brosse, N.; Chrusciel, L.; Sanchez, C.; Sannigrahi, P.; Ragauskas, A. Characterization of milled wood lignin and ethanol organosolv lignin from miscanthus. Polym. Degrad. Stab. 2009, 94, 1632–1638. [Google Scholar] [CrossRef]
- El Hage, R.; Perrin, D.; Brosse, N. Effect of the pre-treatment severity on the antioxidant properties of ethanol organosolv Miscanthus × giganteus lignin. Nat. Resour. 2012, 3, 29–34. [Google Scholar] [CrossRef]
- El Hage, R.; Brosse, N.; Sannigrahi, P.; Ragauskas, A. Effects of process severity on the chemical structure of Miscanthus ethanol organosolv lignin. Polym. Degrad. Stab. 2010, 95, 997–1003. [Google Scholar] [CrossRef]
- Chan, J.M.W.; Bauer, S.; Sorek, H.; Sreekumar, S.; Wang, K.; Toste, F.D. Studies on the vanadium-catalyzed nonoxidative depolymerization of Miscanthus giganteus-derived lignin. ACS Catal. 2013, 3, 1369–1377. [Google Scholar] [CrossRef]
- Luo, H.; Klein, I.M.; Jiang, Y.; Zhu, H.; Liu, B.; Kenttämaa, H.I.; Abu-Omar, M.M. Total Utilization of Miscanthus Biomass, Lignin and Carbohydrates, Using Earth Abundant Nickel Catalyst. ACS Sustain. Chem. Eng. 2016, 4, 2316–2322. [Google Scholar] [CrossRef]
- Vanderghem, C.; Richel, A.; Jacquet, N.; Blecker, C.; Paquot, M. Impact of formic/acetic acid and ammonia pre-treatments on chemical structure and physico-chemical properties of Miscanthus x giganteus lignins. Polym. Degrad. Stab. 2011, 96, 1761–1770. [Google Scholar] [CrossRef]
- Timilsena, Y.P.; Abeywickrama, C.J.; Rakshit, S.K.; Brosse, N. Effect of different pretreatments on delignification pattern and enzymatic hydrolysability of miscanthus, oil palm biomass and typha grass. Bioresour. Technol. 2013, 135, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Chen, X.; Shi, C.; Li, M.; Guan, Y.; Yeon, C.; Yamada, Y.T.; Sacks, E.J.; Peng, J. Determination of hemicellulose, cellulose and lignin content using visible and near infrared spectroscopy in Miscanthus sinensis. Bioresour. Technol. 2017, 241, 603–609. [Google Scholar] [CrossRef] [PubMed]
- van der Weijde, T.; Kiesel, A.; Iqbal, Y.; Muylle, H.; Dolstra, O.; Visser, R.G.F.; Lewandowski, I.; Trindade, L.M. Evaluation of Miscanthus sinensis biomass quality as feedstock for conversion into different bioenergy products. GCB Bioenergy 2016, 9, 176–190. [Google Scholar] [CrossRef]
- Baker, P.W.; Winters, A.; Hale, M.D.C. Biodegradation of Different Genotypes of Miscanthus by Wood Rot Fungi. BioResources 2016, 11, 4379–4391. [Google Scholar] [CrossRef]
- Ion, S.; Opris, C.; Cojocaru, B.; Tudorache, M.; Zgura, I.; Galca, A.C.; Bodescu, A.M.; Enache, M.; Maria, G.; Parvulescu, V.I. One-Pot Enzymatic Production of Lignin-Composites. Front. Chem. 2018, 6, 124. [Google Scholar] [CrossRef] [PubMed]
- Sonnenberg, A.M.; Baars, J.J.P.; Visser, M.H.M.; Lavrijssen, B.; Hendrickx, P.M. Evaluation of Shiitake Strains (Lentinula edodes) on Selective Lignin Degradation in Miscanthus × giganteus; Wageningen UR, Plant Breeding: Wageningen, The Netherlands, 2016. [Google Scholar]
- da Costa, R.M.F.; Lee, S.J.; Allison, G.G.; Hazen, S.P.; Winters, A.; Bosch, M. Genotype, development and tissue-derived variation of cell-wall properties in the lignocellulosic energy crop Miscanthus. Ann. Bot. 2014, 114, 1265–1277. [Google Scholar] [CrossRef]
- Iqbal, Y.; Lewandowski, I. Inter-annual variation in biomass combustion quality traits over five years in fifteen Miscanthus genotypes in south Germany. Fuel Process. Technol. 2014, 121, 47–55. [Google Scholar] [CrossRef]
- Ralph, J.; Landucci, L. NMR of lignins. In Lignin and Lignans: Advances in Chemistry; Taylor & Francis: Boca Raton, FL, USA, 2010; pp. 137–244. [Google Scholar]
- Lupoi, J.S.; Singh, S.; Parthasarathi, R.; Simmons, B.A.; Henry, R.J. Recent innovations in analytical methods for the qualitative and quantitative assessment of lignin. Renew. Sustain. Energy. Rev. 2015, 49, 871–906. [Google Scholar] [CrossRef]
- Tarasov, D.; Leitch, M.; Fatehi, P. Lignin–carbohydrate complexes: Properties, applications, analyses, and methods of extraction: A review. Biotechnol. Biofuels 2018, 11, 269. [Google Scholar] [CrossRef]
- Li, M.; Pu, Y.; Ragauskas, A.J. Current Understanding of the Correlation of Lignin Structure with Biomass Recalcitrance. Front. Chem. 2016, 4, 45. [Google Scholar] [CrossRef] [PubMed]
- Lang, J.M.; Shrestha, U.M.; Dadmun, M. The Effect of Plant Source on the Properties of lignin-Based Polyurethanes. Front. Energy Res. 2018, 6, 4. [Google Scholar] [CrossRef]
- Klein, S.E.; Rumpf, J.; Kusch, P.; Albach, R.; Rehahn, M.; Witzleben, S.; Schulze, M. Utilization of Unmodified Kraft Lignin for the Preparation of Highly Flexible and Transparent Polyurethane Coatings. RSC Adv. 2018, 8, 40765–40777. [Google Scholar] [CrossRef]
- Klein, S.E.; Rumpf, J.; Rehahn, M.; Witzleben, S.; Schulze, M. Biobased Flexible Polyurethane Coatings Prepared from Kraft Lignin: One-Pot Synthesis and Antioxidant Activity. J. Coat. Technol. Res. 2019, in press. [Google Scholar]
- Witzler, M.; Alzagameem, A.; Bergs, M.; El Khaldi-Hansen, B.; Klein, S.E.; Hielscher, D.; Kamm, B.; Kreyenschmidt, J.; Tobiasch, E.; Schulze, M. Lignin-Derived Biomaterials for Drug Release and Tissue Engineering. Molecules 2018, 23, 1885. [Google Scholar] [CrossRef] [PubMed]
- Groenewold, G.S.; Johnson, K.M.; Fox, S.C.; Rae, C.; Zarzana, C.A.; Kersten, B.R.; Rowe, S.M.; Westover, T.L.; Gresham, G.L.; Emerson, R.M. Pyrolysis Two-Dimensional GC-MS of Miscanthus Biomass: Quantitative Measurement using an Internal Standard Method. Energy Fuels 2017, 31, 1620–1630. [Google Scholar] [CrossRef]
- Bottcher, A.; Cesarino, I.; dos Santos, A.B.; Vicentini, R.; Mayer, J.L.S.; Vanholme, R.; Morreel, K.; Goeminne, G.; Moura, J.C.M.S.; Nobile, P.M.; et al. Lignification in sugarcane: Biochemical characterization gene discovery, and expression analysis in two genotypes contrasting for lignin content. Plant Physiol. 2013, 163, 1539–1557. [Google Scholar] [CrossRef] [PubMed]
- Del Río, J.C.; Rencoret, J.; Prinsen, P.; Martinez, A.T.; Ralph, J.; Gutierrez, A. Structural characterization of wheat straw lignin as revealed by analytical pyrolysis, 2D-NMR, and reductive cleavage methods. J. Agric. Food Chem. 2012, 60, 5922–5935. [Google Scholar] [CrossRef] [PubMed]
- Villaverde, J.J.; Li, J.; Ek, M.; Ligero, P.; Vega, A.D. Native lignin structure of Miscanthus × giganteus and its changes during acetic and formic acid fractionation. J. Agric. Food Chem. 2009, 57, 6262–6270. [Google Scholar] [CrossRef] [PubMed]
- Boerjan, W.; Ralph, J.; Baucher, M. Lignin biosynthesis. Annu. Rev. Plant Biol. 2003, 54, 519–546. [Google Scholar] [CrossRef]
- Kukkola, E.M.; Koutaniemi, S.; Pollanen, E.; Gustafsson, M.; Karhunen, P.; Lundell, T.K.; Saranpaa, P.; Kilpelainen, I.; Teeri, T.H.; Fagerstedt, K.V. The dibenzodioxocin lignin substructure is abundant in the inner part of the secondary wall in Norway spruce and silver birch xylem. Planta 2004, 218, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Argyropoulos, D.S.; Jurasek, L.; Kristofova, L.; Xia, Z.; Sun, Y.; Palus, E. Abundance and reactivity of dibenzodioxocins in softwood lignin. J. Agric. Food Chem. 2002, 50, 658–666. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Dai, G.; Yang, H.; Luo, Z. Lignocellulosic biomass pyrolysis mechanism: A state-of-the-art review. Prog. Energy Combust. Sci. 2017, 62, 33–86. [Google Scholar] [CrossRef]
- Li, Z.; Zhao, C.; Zha, Y.; Wan, C.; Si, S.; Liu, F.; Zhang, R.; Li, F.; Yu, B.; Yi, Z.; et al. The minor wall-networks between monolignols and interlinked-phenolics predominantly affect biomass enzymatic digestibility in Miscanthus. PLoS ONE 2014, 9, e105115. [Google Scholar] [CrossRef] [PubMed]
- Lan, W.; Yue, F.; Rencoret, J.; Carlos del Río, J.; Boerjan, W.; Lu, F.; Ralph, J. Elucidating Tricin-Lignin Structures: Assigning Correlations in HSQC Spectra of Monocot Lignins. Polymers 2018, 10, 916–922. [Google Scholar] [CrossRef]
- Monakhova, Y.; Diehl, B.W.K.; Do, X.T.; Witzleben, S.; Schulze, M. Novel method for the determination of average molecular weight of natural polymers based on 2D DOSY NMR and chemometrics: Example of heparin. J. Pharm. Biomed. Anal. 2018, 149, 128–132. [Google Scholar] [CrossRef] [PubMed]
- Boeriu, C.G.; Bravo, D.; Gosselink, R.J.A.; van Dam, J.E.G. Characterisation of structure-dependent functional properties of Lignin with infrared spectroscopy. Ind. Crops Prod. 2004, 20, 205–218. [Google Scholar] [CrossRef]
- Chen, H.; Ferrari, C.; Angiuli, M.; Yao, J.; Raspi, C.; Bramanti, E. Qualitative and quantitative analysis of wood samples by Fourier transform infrared spectroscopy and multivariate analysis. Carbohydr. Polym. 2010, 82, 772–778. [Google Scholar] [CrossRef]
- Christou, C.; Agapiou, A.; Kokkinofta, R. Use of FTIR spectroscopy and chemometrics for the classification of carobs origin. J. Adv. Res. 2018, 10, 1–8. [Google Scholar] [CrossRef]
- Wahid, R.; Nielsen, S.F.; Hernandez, V.M.; Ward, A.J.; Gislum, R.; Jørgensen, U.; Møller, H.B. Methane production potential from Miscanthus sp: Effect of harvesting time, genotypes and plant fractions. Biosyst. Eng. 2015, 133, 71–80. [Google Scholar] [CrossRef]
- Bergs, M. Einfluss von Miscanthus-Genotyp und Erntezeit auf Gehalt und Struktur von Lignin aus Organosolv-Verfahren. Ph.D. Thesis, Rheinische Friedrich-Wilhelms-Universität Bonn, Bonn, Germany, 20 December 2018. [Google Scholar]
- Hafez, I.; Hassan, E.B. Rapid liquefaction of giant miscanthus feedstock in ethanol–water system for production of biofuels. Energy Conv. Manag. 2015, 91, 219–224. [Google Scholar] [CrossRef]
- Scagline-Mellor, S.; Griggs, T.; Skousen, J.; Wolfrum, E.; Holásková, I. Switchgrass and Giant Miscanthus Biomass and Theoretical Ethanol Production from Reclaimed Mine Lands. Bioenergy Res. 2018, 11, 562–573. [Google Scholar] [CrossRef]
- Sørensen, A.; Teller, P.J.; Hilstrøm, T.; Ahring, B.K. Hydrolysis of Miscanthus for bioethanol production using dilute acid presoaking combined with wet explosion pre-treatment and enzymatic treatment. Bioresour. Technol. 2008, 99, 6602–6607. [Google Scholar] [CrossRef] [PubMed]
- Cha, Y.-L.; An, G.H.; Yang, J.; Moon, Y.-H.; Yu, G.-D.; Ahn, J.-W. Bioethanol production from Miscanthus using thermotolerant Saccharomyces cerevisiae mbc 2 isolated from the respiration-deficient mutants. Renew. Energy 2015, 80, 259–265. [Google Scholar] [CrossRef]
- Yang, H.; Yan, R.; Chen, H.; Lee, D.H.; Zheng, C. Characteristics of hemicellulose, cellulose and lignin pyrolysis. Fuel 2007, 86, 1781–1788. [Google Scholar] [CrossRef]
- Schorr, D.; Diouf, P.N.; Stevanovic, T. Evaluation of industrial lignins for biocomposites production. Ind. Crops Prod. 2014, 52, 65–73. [Google Scholar] [CrossRef]
- Bauer, S.; Sorek, H.; Mitchell, V.D.; Ibáñez, A.B.; Wemmer, D.E. Characterization of Miscanthus giganteus lignin isolated by ethanol organosolv process under reflux condition. J. Agric. Food Chem. 2012, 60, 8203–8212. [Google Scholar] [CrossRef]
- Hansen, B.; Kusch, P.; Schulze, M.; Kamm, B. Qualitative and quantitative analysis of lignin produced from beech wood by different conditions of the Organosolv process. J. Polym. Environ. 2016, 24, 85–97. [Google Scholar] [CrossRef]
- Hansen, B.; Kamm, B.; Schulze, M. Qualitative and quantitative analysis of lignins from different sources and isolation methods for an application as a biobased chemical resource and polymeric material. In Analytical Techniques and Methods for Biomass Products; Vaz, S., Jr., Seidl, P., Eds.; Springer: Berlin, Germany, 2017; pp. 15–44. ISBN 978-3-319-41414-0. [Google Scholar]
- Brandt, A.; Chen, L.; van Dongen, B.E.; Welton, T.; Hallett, J.P. Structural changes in lignins isolated using an acidic ionic liquid water mixture. Green Chem. 2015, 17, 5019–5034. [Google Scholar] [CrossRef]
- Alzagameem, A.; El Khladi-Hansen, B.; Büchner, D.; Larkins, M.; Kamm, B.; Witzleben, S.; Schulze, M. Environmentally Benign Lignin-based Antioxidants isolated from Lignocellulose Feedstock. Molecules 2018, 23, 2664. [Google Scholar] [CrossRef]
- Fenner, R.A.; Lephardt, J.O. Examination of the thermal decomposition of kraft pine lignin by Fourier transform infrared evolved gas analysis. J. Agric. Food Chem. 1981, 29, 846–849. [Google Scholar] [CrossRef]
- Norambuena, M.; Vidal, C.; Carrasco, L.; Contreras, D.; Mendonça, R.T. Alkaline-catalyzed Modification of Organosolv Lignin: Optimization of Experimental Conditions. In Proceedings of the 5rd Nordic Wood Biorefinery Conference (NWBC 2015), Helsinki, Finland, 20–22 October 2015. [Google Scholar]
- Domínguez, C.; Oliet, M.; Gilarranz, M.A.; Rodríguez, F. Thermal stability and pyrolysis kinetics of organosolv lignins obtained from Eucalyptus globulus. Ind. Crops Prod. 2008, 27, 150–156. [Google Scholar] [CrossRef]
- Wittkowski, R.; Ruther, J.; Drinda, H.; Rafiei-Taghanaki, F. Formation of Smoke Flavor Compounds by Thermal Lignin Degradation. ACS Symp. Ser. 1992, 490, 232–243. [Google Scholar]
- El-Saied, H.; Nada, A.M.A. The thermal behaviour of lignins from wasted black pulping liquors. Polym. Degrad. Stab. 1993, 40, 417–421. [Google Scholar] [CrossRef]
- Glasser, W.G.; Jain, R.K. Lignin derivatives. I. Alkanoates. Holzforschung 1993, 47, 225–233. [Google Scholar] [CrossRef]
- Hodgson, E.M.; Nowakowski, D.J.; Shield, I.; Riche, A.; Bridgwater, A.V.; Clifton-Brown, J.C.; Donnison, I.S. Variation in Miscanthus chemical composition and implications for conversion by pyrolysis and thermo-chemical bio-refining for fuels and chemicals. Bioresour. Technol. 2011, 102, 3411–3418. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, E.M.; Lister, S.J.; Bridgwater, A.V.; Clifton-Brown, J.; Donnison, I.S. Genotypic and environmentally derived variation in the cell wall composition of Miscanthus in relation to its use as a biomass feedstock. Biomass Bioenergy 2010, 34, 652–660. [Google Scholar] [CrossRef]
- Lourenço, A.; Gominho, J.; Pereira, H. Chemical characterization of lignocellulosic materials by analytical pyrolysis. In Analytical Pyrolysis, 1st ed.; Kusch, P., Ed.; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Du, X.; Pérez-Boada, M.; Fernández, C.; Rencoret, J.; del Río, J.C.; Jiménez-Barbero, J.; Li, J.; Gutiérrez, A.; Martínez, A.T. Analysis of lignin-carbohydrate and lignin-lignin linkages after hydrolase treatment of xylan-lignin, glucomannan-lignin and glucan-lignin complexes from spruce wood. Planta 2014, 239, 1079–1090. [Google Scholar] [CrossRef]
- Obama, P.; Ricochon, G.; Muniglia, L.; Brosse, N. Combination of enzymatic hydrolysis and ethanol organosolv pretreatments: Effect of lignin structures, delignification yields and cellulose-to-glucose con-version. Bioresour. Technol. 2012, 112, 156–163. [Google Scholar] [CrossRef]
- Grisel, R.J.H.; van der Waal, J.C.; de Jong, E.; Huijgen, W.J.J. Acid catalysed alcoholysis of wheat straw: Towards second generation furan-derivatives. Catal. Today 2014, 223, 3–10. [Google Scholar] [CrossRef]
- Toledano, A.; Serrano, L.; Labidi, J. Improving base catalyzed lignin depolymerization by avoiding lignin repolymerization. Fuel 2014, 116, 617–624. [Google Scholar] [CrossRef]
- Determination of Structural Carbohydrates and Lignin in Biomass. Available online: https://www.nrel.gov/docs/gen/fy13/42618.pdf (accessed on 10 October 2018).
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. Determination of extractives in biomass. Lab. Anal. Proced. 2008, 1617, 1–16. [Google Scholar]
- Sluiter, A.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D. Determination of Extractives in Biomass (NREL/TP-510-42619); National Renewable Energy Laboratory: Golden, CO, USA, 2008. [Google Scholar]
- Sluiter, A.; Hames, B.; Hyman, D.; Payne, C.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Wolfe, J. Determination of Total Solids in Biomass and Total Dissolved Solids in Liquid Process Samples; NREL Technical Report No. NREL/TP-510-42621; National Renewable Energy Laboratory: Golden, CO, USA, 2008; pp. 1–6. [Google Scholar]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D. Determination of Ash in Biomass (NREL/TP-510-42622); National Renewable Energy Laboratory: Golden, CO, USA, 2005. [Google Scholar]
- Cordella, C.B.Y.; Bertrand, D. SAISIR: A new general chemometric toolbox. Trends Anal. Chem. 2014, 54, 75–82. [Google Scholar] [CrossRef]
- Martens, H.; Naes, T. Multivariate calibration by data compression. In Near Infrared Technology in the Agriculture and Food Industries; William, P., Noris, K., Eds.; American Association of Cereal Chemists: St. Paul, MN, USA, 1987. [Google Scholar]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Linkage | Hard Wood H/G/S traces/25–50/50–75 | Soft Wood H/G/S 0.5–3.4/90–95/0–1 | Miscanthus H/G/S 24/49/27 |
|---|---|---|---|
| β-O-4‘ | 50–65 | 43–50 | 93 |
| A-O-4‘ | 4–8 | 6–8 | ns* |
| β-β‘ | 3–7 | 2–4 | 4 |
| β-5‘ | 4–6 | 9–12 | 3 |
| β-1‘ | 5–7 | 3–7 | traces |
| 4-O-5’ | 6–7 | 4 | ns * |
| 5-5’ | 4–10 | 10–25 | ns * |
| Genotype Harvest Time Portion | Gig17 April Leaf | Gig34 April Leaf | Gig17 April Stem | Gig34 April Stem |
|---|---|---|---|---|
| AIL (%) | 20.65 ± 0.47 | 21.06 ± 0.31 | 21.18 ± 0.01 | 20.97 ± 0.43 |
| ASL (%) | 5.06 ± 0.30 | 4.13 ± 0.05 | 4.71 ± 0.26 | 4.31 ± 0.02 |
| AIR (%) | 21.83 ± 0.51 | 22.14 ± 0.42 | 22.46 ± 0.22 | 22.31 ± 0.60 |
| Total lignin (%) | 25.52 ± 0.54 | 25.32 ± 0.48 | 26.00 ± 0.36 | 25.25 ± 0.39 |
| AISA (%) | 1.18 ± 0.10 | 1.08 ± 0.22 | 1.29 ± 0.11 | 1.01 ± 0.05 |
| Glucan (%) | 44.79 ± 1.50 | 44.98 ± 2.30 | 50.02 ± 0.63 | 50.49 ± 0.87 |
| Xylan (%) | 28.37 ± 1.77 | 29.52 ± 0.63 | 27.41 ± 2.81 | 26.23 ± 0.49 |
| Galactan (%) | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Arabinan (%) | 3.11 ± 0.05 | 4.11 ± 1.63 | 1.89 ± 0.51 | 2.04 ± 0.34 |
| Mannan (%) | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| Dry matter (%) | 92.55 | 91.25 | 92.18 | 92.16 |
| Total ash (%) | 4.53 | 6.82 | 2.50 | 3.07 |
| Number | Wave Number [cm−1] | Functional Group | Assignment |
|---|---|---|---|
| 1 | 3428 ± 60 | O-H | Stretching |
| 2 | 2926 ± 11 | C-H | Stretching |
| 3 | 2850 ± 6 | C-H | Stretching |
| 4 | 1708 ± 11 | C=O | Stretching |
| 5 | 1655 ± 4 | C=O | Stretching |
| 6 | 1605 ± 12 | Aromatic ring | Symmetric Stretching |
| 7 | 1514 ± 6 | Aromatic ring | Antisymmetric Stretching |
| 8 | 1460 ± 4 | C-H | Antisymmetric Deformation |
| 9 | 1424 ± 4 | C-H in O-CH3 | Antisymmetric Deformation (S mode) |
| 10 | 1398 ± 4 | C-H | Bending C-H in-plane |
| 11 | 1371 ± 15 | Aromatic skeleton | Deformation |
| 12 | 1331 ± 9 | Aromatic skeleton; C-O | Skeleton Stretching (S mode) |
| 13 | 1267 ± 2 | Aromatic skeleton; C-O | Skeleton Stretching (G mode) |
| 14 | 1225 ± 12 | C-C; C-O; C=O | Stretching (G mode) |
| 15 | 1166 ± 10 | C-H (in G) | Stretching |
| 16 | 1124 ± 2 | Aromatic C-H | In-plane Deformation |
| 17 | 1033 ± 2 | Aromatic C-H | In-plane Deformation |
| 18 | 834 ± 4 | Aromatic C-H | Out-of-plane Deformation (S mode) |
| SEC Results | Detector | Gig17M | Gig34M | Gig17L | Gig34L | Gig17S | Gig34S |
|---|---|---|---|---|---|---|---|
| Mn (g·mol−1) | RI | 1138.80 | 1139.10 | 967.13 | 960.96 | 1314.40 | 1206.30 |
| UV | 1077.70 | 1084.40 | 904.49 | 931.43 | 1168.90 | 1120.00 | |
| Mw (g·mol−1) | RI | 2041.40 | 2146.40 | 1758.70 | 1800.10 | 2385.00 | 2098.70 |
| UV | 1983.40 | 2050.90 | 1691.60 | 1745.00 | 2216.30 | 2033.90 | |
| PD | RI | 1.79 | 1.88 | 1.82 | 1.87 | 1.81 | 1.74 |
| UV | 1.84 | 1.89 | 1.87 | 1.87 | 1.90 | 1.82 |
| Sample | First ML (120 °C) | Second ML (350 °C) | RM | Td |
|---|---|---|---|---|
| Gig17M | 0.86% | 75.88% | 23.26% | 358.2 °C |
| Gig17L | 0.91% | 64.98% | 34.11% | 354.8 °C |
| Gig17S | 0.42% | 65.48% | 34.10% | 381.6 °C |
| Gig34M | 0.54% | 73.79% | 25.67% | 360.0 °C |
| Gig34L | 0.72% | 71.41% | 27.87% | 336.1 °C |
| Gig34S | 4.60% | 84.08% | 11.32% | 379.2 °C |
| Peak No. | Retention Time (min) | Amount (%) | Name | Assignment |
|---|---|---|---|---|
| 1 | 9.49 | 3.77 | Phenol | H |
| 2 | 11.36 | 10.07 | Guaiacol | G |
| 3 | 11.65 | 1.83 | p-Cresol | H |
| 4 | 13.32 | 0.70 | Creosol | G |
| 5 | 13.67 | 1.27 | 4-Ethylphenol | H |
| 6 | 13.72 | 12.92 | Creosol | G |
| 7 | 15.55 | 0.91 | 4-Ethylguaiacol | G |
| 8 | 15.69 | 22.08 | Coumaran | H |
| 9 | 16.874 | 8.17 | 4-Hydroxy-2-methylacetophenone | H |
| 10 | 17.66 | 1.67 | Eugenol | G |
| 11 | 17.87 | 5.68 | Syringol | S |
| 12 | 19.00 | 1.55 | Eugenol | G |
| 13 | 20.13 | 6.80 | 4-Methylsyringol | S |
| 14 | 20.24 | 6.23 | (E)-Isoeugenol | G |
| 15 | 21.05 | 1.91 | Vanillin | G |
| 16 | 24.26 | 1.68 | 4-Allylsyringol | S |
| 17 | 25.95 | 1.56 | (E)-4-Propenylsyringol | S |
| 18 | 28.20 | 6.91 | (E)-4-Propenylsyringol | S |
| 19 | 32.03 | 2.50 | Syringaldehyde | S |
| 20 | 34.70 | 1.78 | Acetosyringone | S |
| Sample | H (%) | G (%) | S (%) |
|---|---|---|---|
| Gig17M | 37.13 | 35.96 | 26.91 |
| Gig17L | 46.35 | 45.82 | 7.83 |
| Gig17S | 35.11 | 32.94 | 31.95 |
| Gig34M | 43.79 | 29.50 | 26.71 |
| Gig34L | 46.30 | 39.25 | 14.45 |
| Gig34S | 35.06 | 38.55 | 26.39 |
| Number | Integral (rel) | δ 1H (ppm) | δ 13C (ppm) | Name | Assignment |
|---|---|---|---|---|---|
| 1 | 0.0190 | 3.45 | 52.88 | B | B |
| 2 | 0.0254 | 3.06 | 53.34 | C | C |
| 3 | 0.7089 | 3.56 | 59.66 | A | A |
| 4 | 0.1322 | 3.23 | 59.71 | B | B |
| 5 | 0.1932 | 3.68 | 62.73 | A | A |
| 6 | 0.0321 | 3.82 | 71.00 | C | C |
| 7 | 0.0441 | 4.17 | 70.76 | C | C |
| 8 | 0.0561 | 4.74 | 70.95 | A | A |
| 9 | 0.1541 | 4.86 | 71.37 | A | A |
| 10 | 0.0435 | 4.47 | 79.78 | A | A |
| 11 | 0.1160 | 4.31 | 83.63 | A | A |
| 12 | 0.0361 | 4.64 | 84.75 | C | C |
| 13 | 0.1106 | 4.07 | 85.77 | A | A |
| 14 | 0.0626 | 5.45 | 86.79 | B | B |
| 15 | 0.7042 | 6.73 | 103.42 | S 2/6 | S |
| 16 | 0.0411 | 6.99 | 103.24 | S 2/6 | S |
| 17 | 0.0337 | 7.33 | 103.79 | S 2/6 | S |
| 18 | 0.4662 | 6.96 | 109.88 | G 2 | G |
| 19 | 0.1192 | 6.30 | 113.46 | D | D |
| 20 | 1.0000 | 6.71 | 114.80 | G 5 | G |
| 21 | 0.1827 | 6.79 | 118.61 | G 6 | G |
| 22 | 0.1588 | 7.12 | 127.35 | H 2/6 | H |
| 23 | 0.4032 | 7.48 | 129.07 | H 2/6 | H |
| 24 | 0.1315 | 7.43 | 143.84 | D | D |
| Sample | H | G | S |
|---|---|---|---|
| Gig17M | 23.03 | 45.05 | 31.92 |
| Gig17L | 13.65 | 67.24 | 19.11 |
| Gig17S | 15.77 | 57.67 | 26.56 |
| Gig34M | 21.17 | 53.54 | 25.29 |
| Gig34L | 14.87 | 63.43 | 21.70 |
| Gig34S | 19.07 | 54.80 | 26.13 |
| Sample | A (β-Aryl Ether) % | B (Phenylcoumaran) % | C (Resinol) % | D (Unsaturated Ester) % |
|---|---|---|---|---|
| Gig17M | 61.84 | 9.56 | 6.16 | 22.43 |
| Gig17L | 65.31 | 9.43 | 4.35 | 20.91 |
| Gig17S | 58.14 | 9.37 | 8.18 | 24.31 |
| Gig34M | 62.62 | 7.70 | 8.00 | 21.68 |
| Gig34L | 66.41 | 10.10 | 4.92 | 18.57 |
| Gig34S | 54.73 | 8.22 | 7.28 | 29.77 |
| Component | Description | Parameter |
|---|---|---|
| Eluent | THF p.a. | HPLC-grade |
| Pump | Agilent 1100 Series | Flowrate: 1.000 mL min−1 |
| Injector | Rheodyne 7725i sample loop | Injection volume: 60 µL |
| Oven | Agilent 1100 Series | Temperature: 35 °C |
| Column | 1× PSS SDV 8 × 50 mm pre-column | Particle size: 5 µm |
| 2× PSS SDV 8 × 300 mm Linear M 5µ | Particle size: 5 µm Molar weight range: 50–10,000,000 g mol−1 | |
| Detector | Agilent 1100 Series VWD | Wavelength: 280 nm |
| Calibration | PSS ReadyCal-Kit Polystyren | Calibration range: 376–2,570,000 g mol−1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bergs, M.; Völkering, G.; Kraska, T.; Pude, R.; Do, X.T.; Kusch, P.; Monakhova, Y.; Konow, C.; Schulze, M. Miscanthus x giganteus Stem Versus Leaf-Derived Lignins Differing in Monolignol Ratio and Linkage. Int. J. Mol. Sci. 2019, 20, 1200. https://doi.org/10.3390/ijms20051200
Bergs M, Völkering G, Kraska T, Pude R, Do XT, Kusch P, Monakhova Y, Konow C, Schulze M. Miscanthus x giganteus Stem Versus Leaf-Derived Lignins Differing in Monolignol Ratio and Linkage. International Journal of Molecular Sciences. 2019; 20(5):1200. https://doi.org/10.3390/ijms20051200
Chicago/Turabian StyleBergs, Michel, Georg Völkering, Thorsten Kraska, Ralf Pude, Xuan Tung Do, Peter Kusch, Yulia Monakhova, Christopher Konow, and Margit Schulze. 2019. "Miscanthus x giganteus Stem Versus Leaf-Derived Lignins Differing in Monolignol Ratio and Linkage" International Journal of Molecular Sciences 20, no. 5: 1200. https://doi.org/10.3390/ijms20051200
APA StyleBergs, M., Völkering, G., Kraska, T., Pude, R., Do, X. T., Kusch, P., Monakhova, Y., Konow, C., & Schulze, M. (2019). Miscanthus x giganteus Stem Versus Leaf-Derived Lignins Differing in Monolignol Ratio and Linkage. International Journal of Molecular Sciences, 20(5), 1200. https://doi.org/10.3390/ijms20051200