Proteomic Analysis of the Resistance Mechanisms in Sugarcane during Sporisorium scitamineum Infection



 ,
,  ,
,
Abstract
1. Introduction
2. Results
2.1. DE Analysis of Differentially Expressed Proteins in Sugarcane after S. scitamineum Inoculation
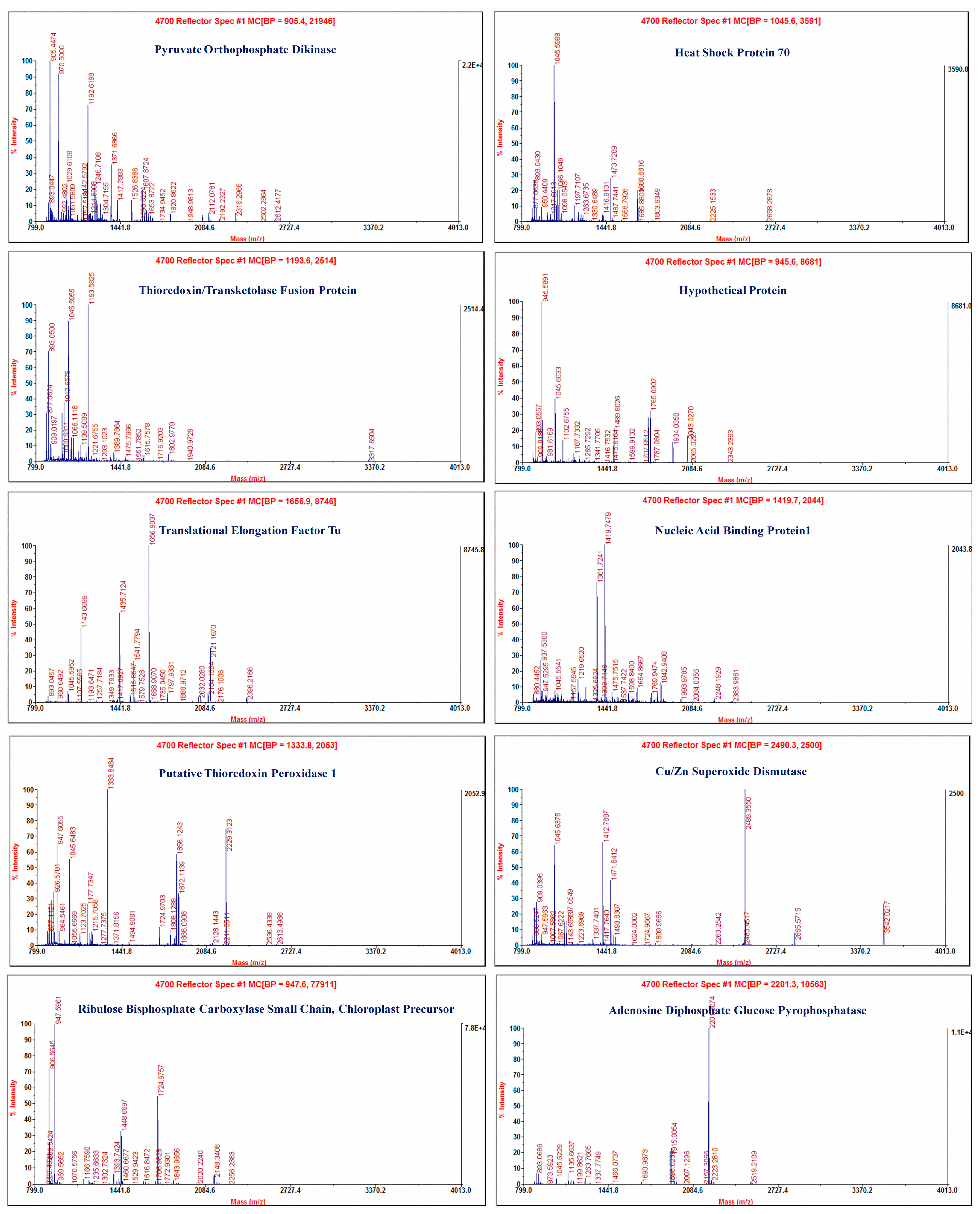
2.2. MALDI-TOF-TOF/MS Analysis of Differentially Expressed Proteins
2.3. Genes Expression Analysis by Real-Time Quantitative PCR (qRT-PCR)
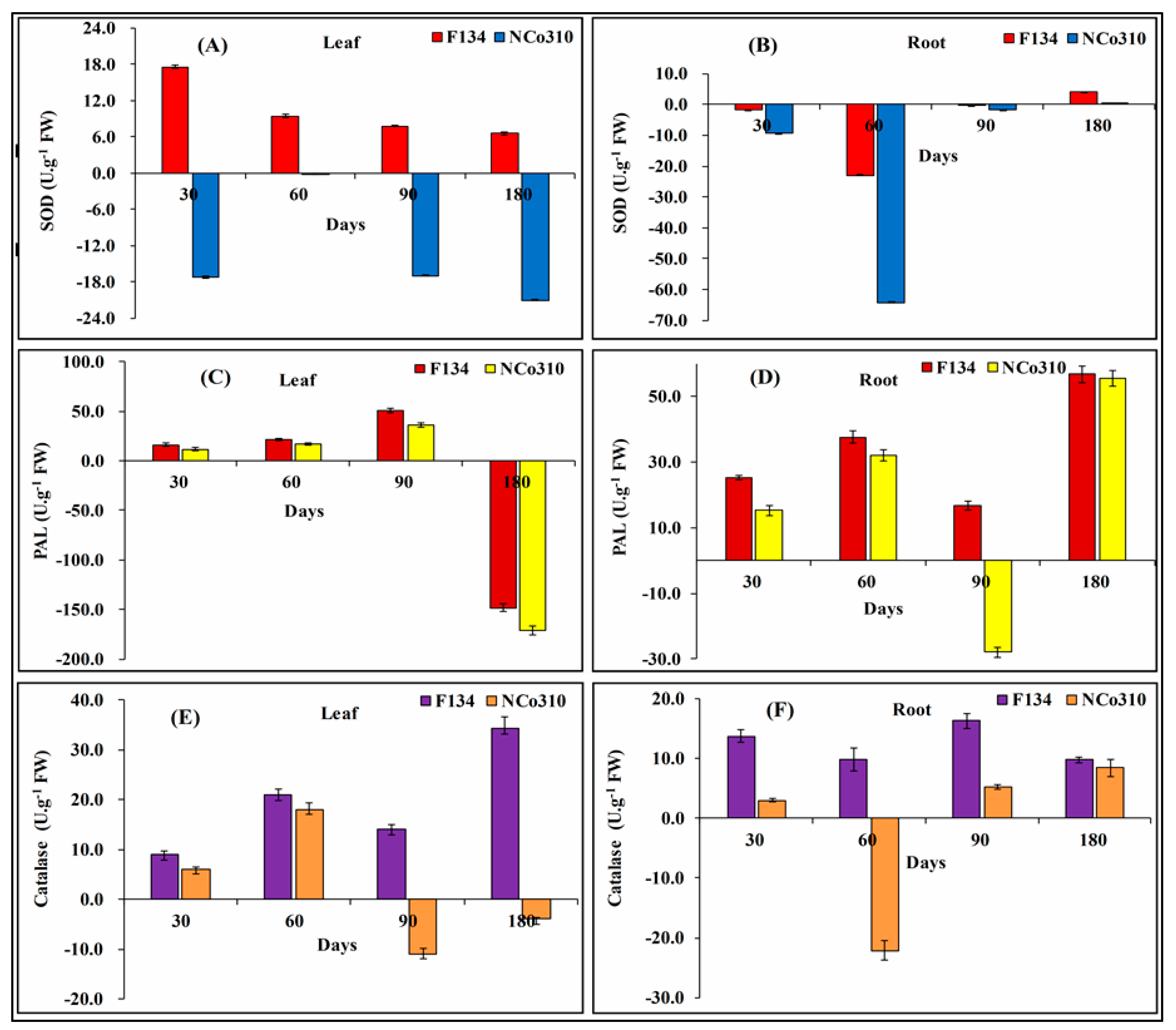
2.4. Induction of Antioxidant Defense System by S. Scitamineum Infection
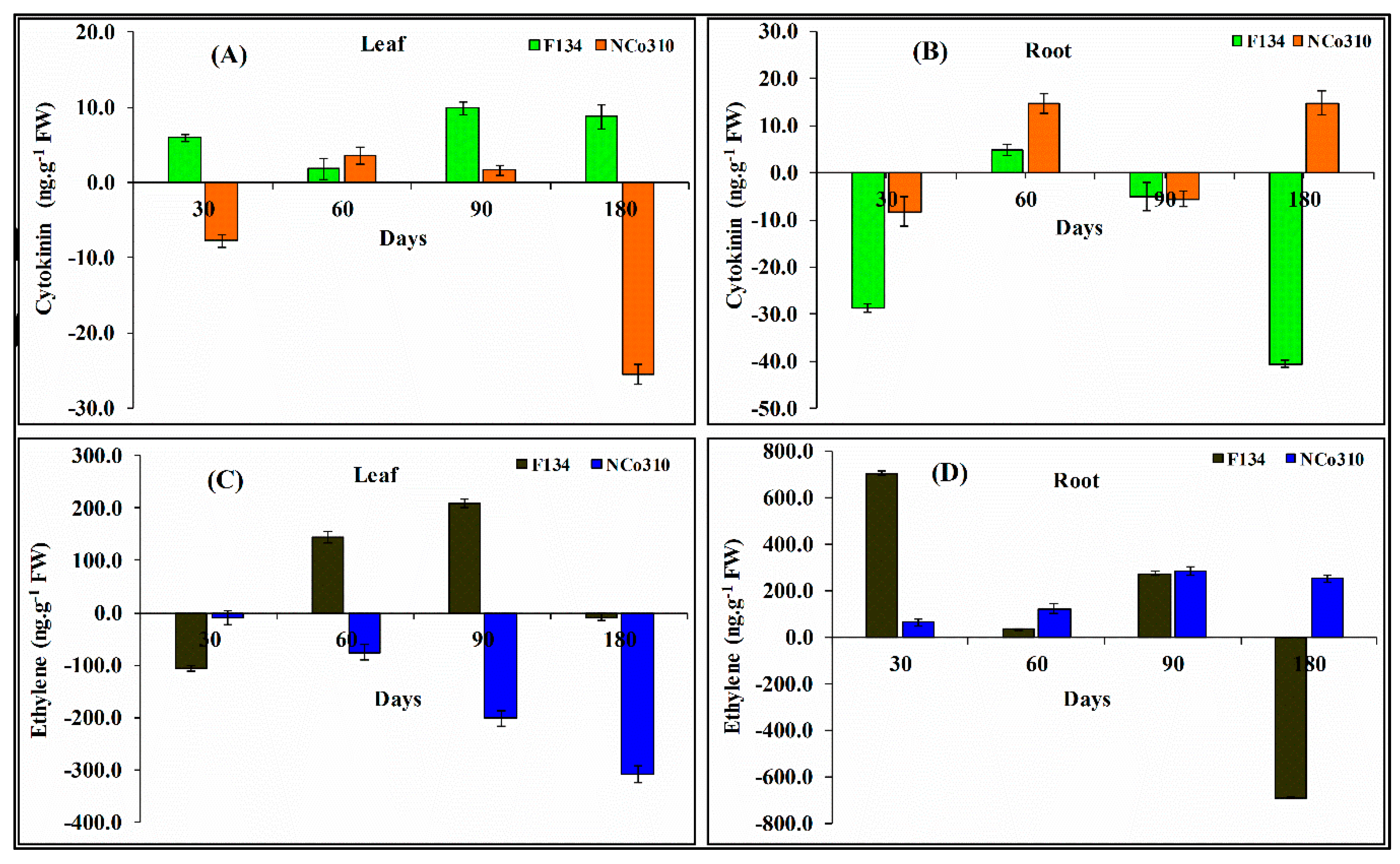
2.5. Phytohormone Levels as Affected by S. Scitamineum Infection
3. Discussion
4. Materials and Methods
4.1. Plant Material, Source of Inoculum, and Inoculation
4.2. Protein Extraction and Quantification
4.3. 2-DE, Image Acquisition, and Analysis
4.4. Mass Spectrometry and Data Analysis
4.5. RNA Extraction, cDNA Synthesis, and qRT-PCR
4.6. Determination of Biochemical Changes in Sugarcane
4.6.1. Antioxidant Enzyme Activities
4.6.2. Hormone Extraction and Assay
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Singh, P.; Song, Q.Q.; Singh, R.K.; Li, H.B.; Solanki, M.K.; Yang, L.T.; Li, Y.R. Physiological and Molecular Analysis of Sugarcane (Varieties-F134 and NCo310) During Sporisorium scitamineum Interaction. Sugar Tech. 2018. [Google Scholar] [CrossRef]
- Su, Y.; Wang, Z.; Xu, L. Early selection for smut resistance in sugarcane using pathogen proliferation and changes in physiological and biochemical indices. Front. Plant Sci. 2016, 7, 1133. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.H. Current status of sugarcane diseases and research progress in China. Sugar Crops China 2007, 3, 48–51. [Google Scholar]
- Su, Y.C.; Wang, S.S.; Guo, J.L.; Xue, B.T.; Xu, L.; Que, Y. A TaqMan real-time PCR assay for detection and quantification of Sporisorium scitamineum in sugarcane. Sci. World J. 2013, 2013, 942682. [Google Scholar] [CrossRef] [PubMed]
- Sundar, A.R.; Barnabas, E.L.; Malathi, P.; Viswanathan, R. A mini-review on smut disease of sugarcane caused by Sporisorium scitamineum. In Botany; Mworia, J., Ed.; InTech Press: Rijeka, Croatia, 2012; pp. 109–128. [Google Scholar]
- Song, X.; Huang, X.; Tian, D.; Yang, L.; Li, Y. Proteomic analysis of sugarcane seedling in response to Ustilago scitaminea infection. Life Sci. J. 2013, 10, 3026–3035. [Google Scholar]
- Croft, B.J.; Braithwaite, K.S. Management of an incursion of sugarcane smut in Australia. Austral. Plant Pathol. 2006, 35, 113–122. [Google Scholar] [CrossRef]
- Xu, L.P.; Chen, R.K.; Chen, P.H. Analysis on infection index of smut caused by Ustilago scitaminea in sugarcane segregated population. Chin. J. Trop. Crops 2004, 25, 33–36. [Google Scholar]
- Scortecci, K.C.; Creste, S.; Calsa, T.J.; Xavier, M.A.; Landell, M.G.A.; Figueira, A.; Benedito, V.A. Challenges, opportunities and recent advances in sugarcane breeding. In Plant Breeding; Abdurakhmonov, I.Y., Ed.; InTech Publisher: Rijeka, Croatia, 2012; pp. 267–296. [Google Scholar]
- Lakshmanan, P.; Geijskes, R.J.; Aitken, K.S.; Grof, C.L.P.; Bonnett, G.D.; Smith, G.R. Sugarcane biotechnology: The challenges and opportunities. In Vitro Cell. Dev. Biol. Plant. 2005, 41, 345–363. [Google Scholar] [CrossRef]
- Singh, A.; Isaac-Kirubakaran, S.; Sakthivel, N. Heterologous expression of new antifungal chitinase from wheat. Protein Exp. Purif. 2007, 56, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.P.; Hoy, J.W.; Saxton, A.M.; Martin, F.A. Heritability of resistance and repeatability of clone reactions to sugarcane smut in Louisiana. Phytopathology 1990, 80, 622–626. [Google Scholar] [CrossRef]
- Dean, J.L. The effect of wounding and high-pressure spray inoculation on the smut reactions of sugarcane clones. Phytopathology 1982, 72, 1023–1025. [Google Scholar] [CrossRef]
- Whittle, A.M.; Walker, D.I.T. Interpretation of sugarcane smut susceptibility trials. Trop. Pest Manag. 1982, 28, 228–237. [Google Scholar] [CrossRef]
- McNeil, M.D.; Bhuiyan, S.A.; Berkman, P.J.; Croft, B.J.; Aitken, K.S. Analysis of the resistance mechanisms in sugarcane during Sporisorium scitamineum infection using RNA-seq and microscopy. PLoS ONE 2018, 13, e0197840. [Google Scholar] [CrossRef] [PubMed]
- Waller, D.I.T. Sugarcane smut (Ustilago scitaminea) in Kenya. II. Infection and Resistance. Trans. Br. Mycol. Soc. 1970, 54, 405–414. [Google Scholar] [CrossRef]
- Lloyd, H.L.; Naidoo, M. Chemical assay potentially suitable for determination of smut resistance of sugarcane cultivars. Plant Dis. 1983, 67, 1103–1105. [Google Scholar] [CrossRef]
- Fontaniella, B.; Marquez, A.; Rodriguez, C.W.; Pinon, D.; Solas, M.T.; Vicente, C.; Legaz, M.E. A role for sugarcane glycoproteins in the resistance of sugarcane to Ustilago scitaminea. Plant Physiol. Biochem. 2002, 40, 881–889. [Google Scholar] [CrossRef]
- Millanes, A.M.; Fontaniella, B.; Legaz, M.E.; Vicente, C. Glycoproteins from sugarcane plants regulate cell polarity of Ustilago scitaminea teliospores. J. Plant Physiol. 2005, 162, 253–265. [Google Scholar] [CrossRef]
- Santiago, R.; Alarcon, B.; de Armas, R.; Vicente, C.; Legaz, M.E. Changes in cinnamoyl alcohol dehydrogenase activities from sugarcane cultivars inoculated with Sporisorium scitamineum sporidia. Physiol. Plant. 2012, 145, 245–259. [Google Scholar] [CrossRef]
- Legaz, M.E.; Pedrosa, M.M.; de Armas, R.; Rodrõguez, C.W.; de los, V.; Vicente, C. Separation of soluble glycoproteins from sugarcane juice by capillary electrophoresis. Anal. Chim. Acta 1998, 372, 201–208. [Google Scholar] [CrossRef]
- Borras-Hidalgo, O.; Thomma, B.P.H.J.; Carmona, E.; Borroto, C.J.; Pujol, M.; Arencibia, A.; Lopez, J. Identification of sugarcane genes induced in disease-resistant somaclones upon inoculation with Ustilago scitaminea or Bipolaris sacchari. Plant Phys. Biochem. 2005, 43, 1115–1121. [Google Scholar] [CrossRef]
- Blanch, M.; Legaz, M.E.; Milanes, A.M.; Vicente, C. Glycoproteins of sugarcane plants facilitate the infectivity of Ustilago scitaminea and Xanthomonas albilineans, two sugarcane pathogens. In Communicating Current Research and Educational Topics and Trends in Applied Microbiology; Mendez-Vilas, A., Ed.; Formatex Research Centre: Badajoz, Spain, 2007; pp. 163–169. [Google Scholar]
- Bhuiyan, S.A.; Croft, B.J.; Cox, M.C.; Bade, G. Varietal resistance of sugarcane to natural infection of smut ± preliminary results. Proc. Aust. Soc. Sug. Technol. 2010, 32, 355–365. [Google Scholar]
- Pinon, D.; de Armas, R.; Vicente, C.; Legaz, M.E. Role of polyamines in the infection of sugarcane buds by Ustilago scitaminea spores. Plant Physiol. Biochem. 1999, 37, 57–64. [Google Scholar] [CrossRef]
- Torres, M.A.; Jones, J.D.G.; Dangl, J.L. Reactive oxygen species signaling in response to pathogens. Plant Physiol. 2006, 141, 373–378. [Google Scholar] [CrossRef] [PubMed]
- De Armas, R.; Santiago, R.; Legaz, M.E.; Vicente, C. Levels of phenolic compounds and enzyme activity can be used to screen for resistance of sugarcane to smut (Ustilago scitaminea). Austral. Plant Pathol. 2007, 36, 32–38. [Google Scholar] [CrossRef]
- Doehlemann, G.; Wahl, R.; Horst, R.J.; Voll, L.M.; Usadel, B.; Poree, F.; Stitt, M.; Pons-Kühnemann, J.; Sonnewald, U.; Kahmann, R.; et al. Reprogramming a maize plant: Transcriptional and metabolic changes induced by the fungal biotroph Ustilago maydis. Plant J. 2008, 56, 181–195. [Google Scholar] [CrossRef]
- Que, Y.X.; Su, Y.C.; Guo, J.L.; Wu, Q.B.; Xu, L.P. A global view of transcriptome dynamics during Sporisorium scitamineum challenge in sugarcane by RNA-Seq. PLoS ONE 2014, 9, e106476. [Google Scholar] [CrossRef]
- Yang, L.; Lin, H.; Takahashi, Y.; Chen, F.; Andrew, W.M.; Edwin, L.C. Proteomic analysis of grapevine stem in response to Xylella fastidiosa inoculation. Physiol. Mol. Plant Pathol. 2011, 75, 90–99. [Google Scholar] [CrossRef]
- Barnabas, L.; Ramadass, A.; Amalraj, R.S.; Palaniyandi, M.; Rasappa, V. Sugarcane proteomics: An update on current status, challenges, and future prospects. Proteomics 2015, 15, 1658–1670. [Google Scholar] [CrossRef] [PubMed]
- Barnabas, L.; Ashwin, N.M.R.; Kaverinathan, K.; Trentin, A.R.; Pivato, M.; Sundar, A.R.; Malathi, P.; Viswanathan, R.; Rosana, O.B.; Neethukrishna, K.; et al. Proteomic analysis of a compatible interaction between sugarcane and Sporisorium scitamineum. Proteomics 2016, 16, 1111–1122. [Google Scholar] [CrossRef]
- Brandalise, M.; Severino, F.E.; Maluf, M.P.; Maia, I.G. The promoter of a gene encoding an isoflavone reductase-like protein in coffee (Coffea arabica) drives a stress-responsive expression in leaves. Plant Cell Rep. 2009, 28, 1699–1708. [Google Scholar] [CrossRef]
- Eilers, M.; Schatz, G. Binding of a specific ligand inhibits import of a purified precursor protein into mitochondria. Nature 1986, 322, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Shoji, T.; Winz, R.; Iwase, T.; Nakajima, K.; Yamada, Y.; Hashimoto, T. Expression patterns of two tobacco isoflavone reductase-like genes and their possible roles in secondary metabolism in tobacco. Plant Mol. Biol. 2002, 50, 427–440. [Google Scholar] [CrossRef] [PubMed]
- Que, Y.; Xu, L.; Lin, J.; Ruan, M.; Zhang, M.; Chen, R. Differential protein expression in sugarcane during sugarcane—Sporisorium scitamineum interaction revealed by 2-DE and MALDI-TOF-TOF/MS. Comp. Funct. Genom. 2011, 2011, 989016. [Google Scholar] [CrossRef] [PubMed]
- Hiraga, S.; Sasaki, K.; Ito, H.; Ohashi, Y.; Matsui, H. A large family of class III plant peroxidases. Plant Cell Physiol. 2001, 42, 462–468. [Google Scholar] [CrossRef]
- Passardi, F.; Cosio, C.; Penel, C.; Dunand, C. Peroxidases have more functions than a Swiss army knife. Plant Cell Rep. 2005, 24, 255–265. [Google Scholar] [CrossRef]
- Chen, W.J.; Zhao, G.W.; Gu, Y.H. Advance of ribulose- 1, 5-bisphosphate carboxylase/oxygenase (RubisCO). Prog. Biochem. Biophys. 1999, 26, 433–436. [Google Scholar]
- DeRocher, E.J.; Quigley, F.; Mache, R.; Bohnert, H.J. The six genes of the Rubisco small subunit multigene family from Mesembryanthemum crystallinum, a facultative CAM plant. Mol. Gen. Genet. 1993, 239, 450–462. [Google Scholar] [CrossRef]
- Carmo-Silva, A.E.; Keys, A.J.; Andralojc, P.J.; Powers, S.J.; Arrabaca, M.C.; Parry, M.A. Rubisco activities, properties, and regulation in three different C4 grasses under drought. J. Exp. Bot. 2010, 61, 2355–2366. [Google Scholar] [CrossRef]
- Moreno, J.I.; Martin, R.; Castresana, C. Arabidopsis SHMT1, a serine hydroxymethyltransferase that functions in the photorespiratory pathway influences resistance to biotic and abiotic stress. Plant J. 2005, 41, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Liu, Q.; Zhang, H.; Jia, Q.; Hong, Y.; Liu, Y. The rubisco small subunit is involved in tobamovirus movement and Tm-2(2)-mediated extreme resistance. Plant Physiol. 2013, 161, 374–383. [Google Scholar] [CrossRef]
- Qiang, X. The Molecular Mechanism of Powdery Mildew Resistance in Chestnut Rose (Rosa Roxburghii Tratt). Ph.D. Dissertation, Huangzhong Agricultural University, Wuhan, China, 2007. [Google Scholar]
- Chastain, C.J.; Fries, J.P.; Vogel, J.A.; Randklev, C.L.; Vossen, A.P.; Dittmer, S.K.; Watkins, E.E.; Fiedler, L.J.; Wacker, S.A.; Meinhover, K.C.; et al. Pyruvate, Orthophosphate Dikinase in Leaves and Chloroplasts of C3 Plants Undergoes Light-/Dark-Induced Reversible Phosphorylation. Plant Physiol. 2002, 128, 1368–1378. [Google Scholar] [CrossRef]
- Chen, H.; Cao, Y.; Li, Y.; Xia, Z.; Xie, J.; Carr John, P.; Wu, B.; Fan, Z.; Zhou, T. Identification of differentially regulated maize proteins conditioning Sugarcane mosaic virus systemic infection. New Phytol. 2017, 215, 1156–1172. [Google Scholar] [CrossRef]
- Fu, J.; Momcilovic, I.; Vara Prasad, P.V. Roles of Protein Synthesis Elongation Factor EF-Tu in Heat Tolerance in Plants Hindawi Publishing Corporation. J. Bot. 2012, 2012, 835836. [Google Scholar]
- Rodrı’guez-Lo’pez, M.; Baroja-Ferna’ndez, E.; Zandueta-Criado, A.; Pozueta-Romero, J. Adenosine diphosphate glucose pyrophosphatase: A plastidial phosphodiesterase that prevents starch biosynthesis Adenosine diphosphate glucose pyrophosphatase: A plastidial phosphodiesterase that prevents starch biosynthesis. Proc. Natl. Acad. Sci. USA 2000, 97, 8705–8710. [Google Scholar] [CrossRef] [PubMed]
- Lao, M.; Arencibia, A.D.; Carmona, E.R.; Acevedo, R.; Rodríguez, E.; León, O.; Santana, I. Differential expression analysis by cDNA-AFLP of Saccharum spp. after inoculation with the host-pathogen Sporisorium scitamineum. Plant Cell Rep. 2008, 27, 1103–1111. [Google Scholar] [CrossRef]
- Song, X.-P.; Huang, X.; Mo, F.-L.; Tian, D.-D.; Yang, L.-T.; Li, Y.-R.; Chen, B.-S. Cloning and Expression Analysis of Sugarcane Phenylalanin Ammonia-lyase (PAL) Gene. Sci. Agric. Sin. 2013, 46, 2856–2868. [Google Scholar]
- Su, Y.; Guo, J.; Ling, H.; Chen, S.; Wang, S.; Xu, L.; Allan, A.C.; Que, Y. Isolation of a Novel Peroxisomal Catalase Gene from Sugarcane, Which Is Responsive to Biotic and Abiotic Stresses. PLoS ONE 2014, 9, e84426. [Google Scholar] [CrossRef]
- Williamson, J.D.; Scandalios, J.G. Differential response of maize catalases and superoxide dismutases to the photoactivated fungal toxin cercosporin. Plant J. 1992, 2, 351–358. [Google Scholar] [PubMed]
- Jain, R.; Chandra, A.; Venugopalan, V.K.; Solomon, S. Physiological Changes and Expression of SOD and P5CS Genes in Response to Water Deficit in Sugarcane. Sugar Tech. 2015, 17, 276–282. [Google Scholar] [CrossRef]
- Bari, R.; Jones, J.D. Role of plant hormones in plant defense responses. Plant Mol. Biol. 2009, 69, 473–488. [Google Scholar] [CrossRef]
- Rivero, R.M.; Kojima, M.; Gepstein, A.; Sakakibara, H.; Mittler, R.; Gepstein, S.; Blumwald, E. Delayed leaf senescence induces extreme drought tolerance in a Flowering plant. Proc. Natl. Acad. Sci. USA 2007, 104, 19631–19636. [Google Scholar] [CrossRef] [PubMed]
- Abeles, F.B.; Morgan, P.W.; Saltveit, M.E., Jr. Ethylene in Plant Biology, 2nd ed.; Academic Press: New York, NY, USA, 1992. [Google Scholar]
- Morgan, P.W.; Drew, M.C. Ethylene and plant responses to stress. Physiol. Plant. 1997, 100, 620–630. [Google Scholar] [CrossRef]
- Su, Y.C.; Yang, Y.T.; Peng, Q.; Zhou, D.G.; Chen, Y.; Wang, Z.; Liping, X.; Youxiong, Q. Development and application of a rapid and visual loop-mediated isothermal amplification for the detection of Sporisorium scitamineum in sugarcane. Sci. Rep. 2016, 6, 23994. [Google Scholar] [CrossRef] [PubMed]
- Bhuiyan, S.A.; Croft, B.J.; Deomano, E.C.; James, R.S.; Stringer, J.K. Mechanism of resistance in Australian sugarcane parent clones to smut and the effect of hot water treatment. Crop Past. Sci. 2013, 64, 892–900. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 255–260. [Google Scholar] [CrossRef]
- Niu, J.Q.; Wang, A.Q.; Huang, J.L.; Yang, L.T.; Li, Y.R. Isolation, characterization and promoter analysis of cell wall invertase gene SoCIN1 from sugarcane (Saccharum spp.). Sugar Tech. 2015, 17, 65–76. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, S.S. Cloning and Expression Analysis of ROS Metabolism Pathway Key Genes from Sugarcane. Master’s Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2012. [Google Scholar]
- Albert, H.H.; Schenck, S. PCR amplification from a homolog of the bE mating-type gene as a sensitive assay for the presence of Ustilago scitaminea DNA. Plant Dis. 1996, 80, 1189–1192. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No. | Accession No. | Identified Protein | Species | Protein | Expression | |||
|---|---|---|---|---|---|---|---|---|
| Mw (Da) | (pI) | Score | Score C.I. (%) | |||||
| 1 | 108796050 | Pyruvate orthophosphate dikinase | Saccharum officinarum | 102,293.9 | 5.5 | 977 | 100 | Down |
| 2 | 6911551 | Heat shock protein 70 | Cucumis sativus | 71,444.1 | 5.07 | 111 | 99.994 | Down |
| 4 | 25067747 | Thioredoxin/transketolase fusion protein | synthetic construct | 86,860.6 | 5.59 | 173 | 100 | UP |
| 6 | 147843754 | Hypothetical protein | Vitis vinifera | 42,395.7 | 6.29 | 158 | 100 | UP |
| 7 | 17225494 | Translational elongation factor Tu | Oryza sativa | 50,381.8 | 6.19 | 477 | 100 | Down |
| 9 | 162463757 | Nucleic acid binding protein1 | Zea mays | 33,096.6 | 4.6 | 87 | 98.709 | UP |
| 16, 17, 19 | 56182370 | Putative thioredoxin peroxidase 1 | Saccharum officinarum | 10,780.7 | 4.9 | 317 | 100 | Down |
| 18 | 125543336 | Hypothetical protein OsI_010708 | Oryza sativa (indica cultivar group) | 22,687.1 | 5.79 | 283 | 100 | Down |
| 22 | 1568639 | Cu/Zn superoxide dismutase | Triticum aestivum | 20,310.4 | 5.35 | 310 | 100 | Down |
| 23, 24 | 3914607 | Ribulose bisphosphate carboxylase small chain, chloroplast precursor | Saccharum officinarum | 19,023.5 | 9.04 | 464 | 100 | Down |
| Spot No. | Accession No. | Identified Protein | Species | Protein | Expression | |||
|---|---|---|---|---|---|---|---|---|
| Mw (Da) | (pI) | Score | Score C.I. (%) | |||||
| 1 | 108796050 | Pyruvate orthophosphate dikinase | Saccharum officinarum | 102,293.9 | 5.5 | 977 | 100 | UP |
| 2 | 6911551 | Heat shock protein 70 | Cucumis sativus | 71,444.1 | 5.07 | 111 | 99.994 | UP |
| 4 | 25067747 | Thioredoxin/transketolase fusion protein | synthetic construct | 86,860.6 | 5.59 | 173 | 100 | UP |
| 6 | 56554972 | Heat shock protein 70 | Medicago sativa | 70,952.1 | 5.08 | 144 | 100 | UP |
| 7, 10 | 17225494 | Translational elongation factor Tu | Oryza sativa | 50,381.8 | 6.19 | 334 | 100 | UP |
| 9 | 162463757 | Nucleic acid binding protein1 | Zea mays | 33,096.6 | 4.6 | 87 | 98.709 | UP |
| 11 | 115488160 | Os12g0277500 | Oryza sativa (japonica cultivar-group) | 61,092.6 | 5.12 | 316 | 100 | Down |
| 20 | 13160411 | Adenosine diphosphate glucose pyrophosphatase | Hordeum vulgare subsp. vulgare | 21,787.1 | 5.68 | 101 | 99.945 | Down |
| 22 | 115439131 | Os01g0675100 | Oryza sativa (japonica cultivar-group) | 17,280.1 | 5.58 | 100 | 99.929 | Down |
| 23, 24 | 3914607 | Ribulose bisphosphate carboxylase small chain, chloroplast precursor | Saccharum officinarum | 19,023.5 | 9.04 | 464 | 100 | UP |
| Gene | Primer | Sequence (5′-3′) | Strategy | Reference |
|---|---|---|---|---|
| GAPDH | GAPDH-F GAPDH-R | CTCTGCCCCAAGCAAAGATG TGTTGTGCAGCTAGCATTG | RT-qPCR | [61] |
| SuPAL | PAL-F PAL-R | CTCGAGGAGAACATCAAGAC GTGATGAGCTCCTTCTCG | RT-qPCR | [50] |
| SuCAT | CAT-F CAT-R | CTTGTCTGGAGCACATACACTTGGA TTCTCCGCATAGACCTTGAACTTTG | RT-qPCR | [63] |
| SuSOD | SOD-F SOD-R | TTTGTCCAAGAGGGAGATGG CTTCTCCAGCGGTGACATTT | RT-qPCR | [53] |
| bEast mating-type | bE4-F bE8-R | CGCTCTGGTTCATCAACG TGCTGTCGATGGAAGGTGT | Genome PCR | [64] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, P.; Song, Q.-Q.; Singh, R.K.; Li, H.-B.; Solanki, M.K.; Malviya, M.K.; Verma, K.K.; Yang, L.-T.; Li, Y.-R. Proteomic Analysis of the Resistance Mechanisms in Sugarcane during Sporisorium scitamineum Infection. Int. J. Mol. Sci. 2019, 20, 569. https://doi.org/10.3390/ijms20030569
Singh P, Song Q-Q, Singh RK, Li H-B, Solanki MK, Malviya MK, Verma KK, Yang L-T, Li Y-R. Proteomic Analysis of the Resistance Mechanisms in Sugarcane during Sporisorium scitamineum Infection. International Journal of Molecular Sciences. 2019; 20(3):569. https://doi.org/10.3390/ijms20030569
Chicago/Turabian StyleSingh, Pratiksha, Qi-Qi Song, Rajesh Kumar Singh, Hai-Bi Li, Manoj Kumar Solanki, Mukesh Kumar Malviya, Krishan Kumar Verma, Li-Tao Yang, and Yang-Rui Li. 2019. "Proteomic Analysis of the Resistance Mechanisms in Sugarcane during Sporisorium scitamineum Infection" International Journal of Molecular Sciences 20, no. 3: 569. https://doi.org/10.3390/ijms20030569
APA StyleSingh, P., Song, Q.-Q., Singh, R. K., Li, H.-B., Solanki, M. K., Malviya, M. K., Verma, K. K., Yang, L.-T., & Li, Y.-R. (2019). Proteomic Analysis of the Resistance Mechanisms in Sugarcane during Sporisorium scitamineum Infection. International Journal of Molecular Sciences, 20(3), 569. https://doi.org/10.3390/ijms20030569