Windup of Nociceptive Flexion Reflex Depends on Synaptic and Intrinsic Properties of Dorsal Horn Neurons in Adult Rats


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
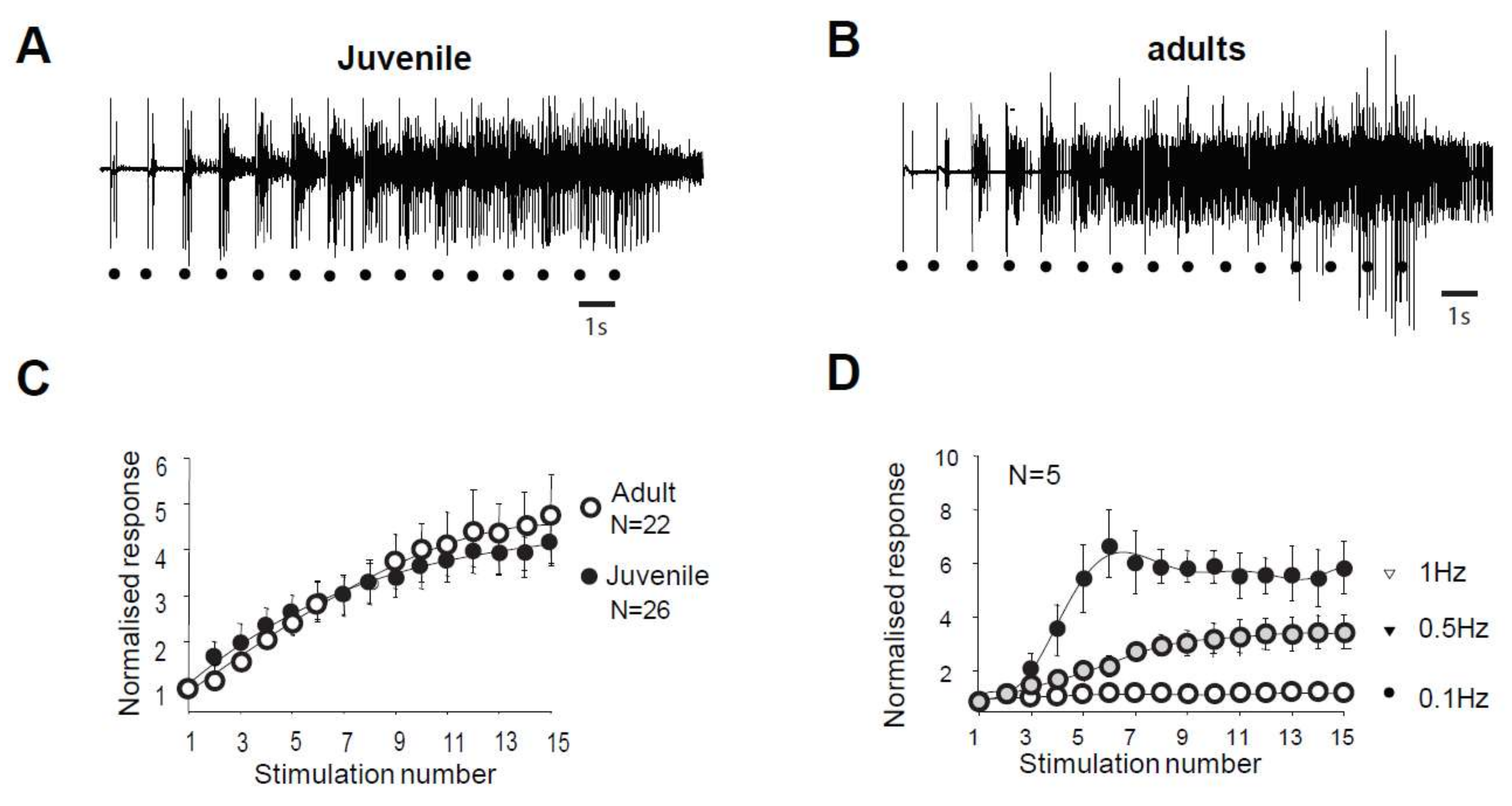
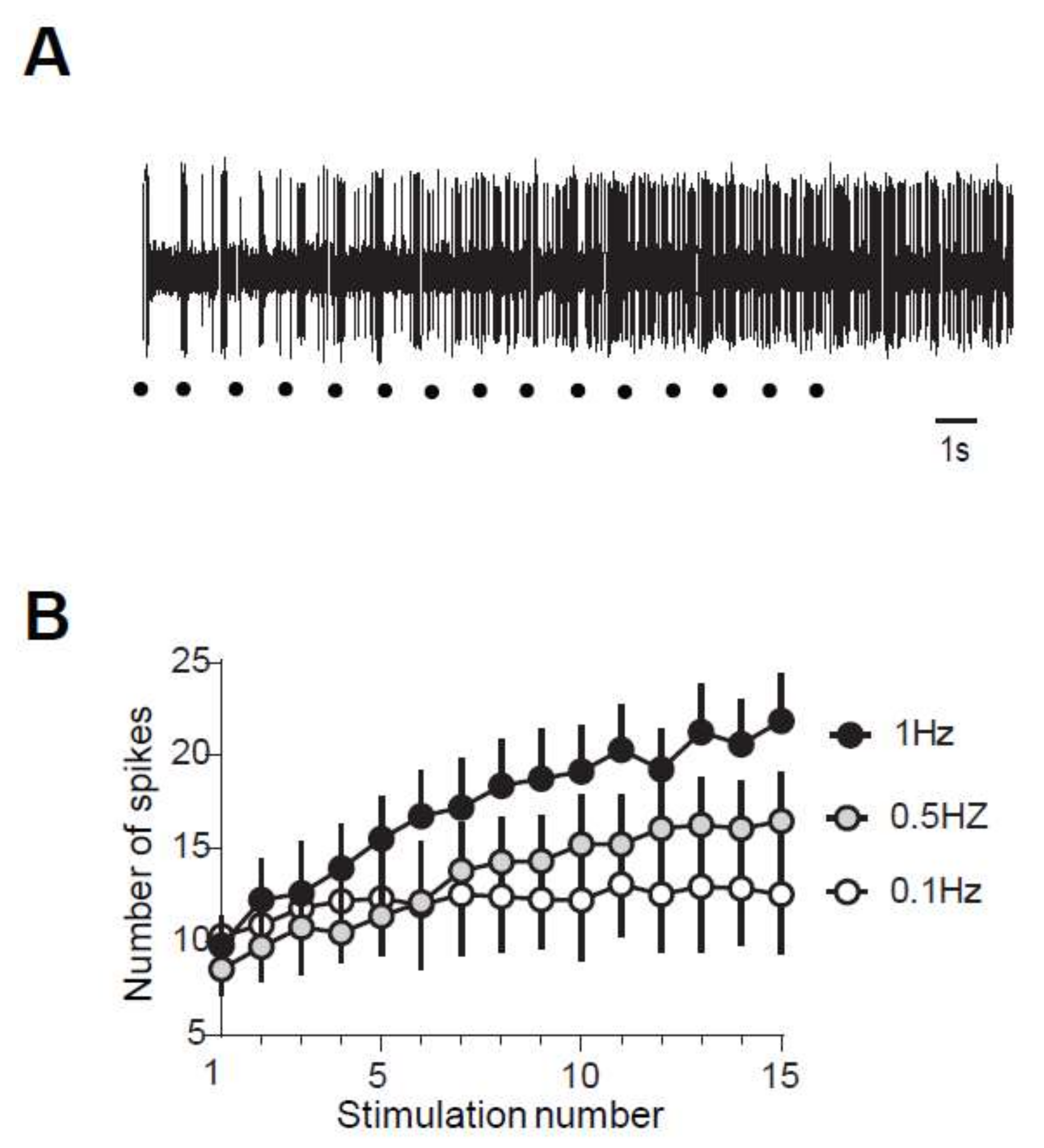
2.1. Windup of WDR Neurons and of the Flexion Reflex Share Similar Characteristics
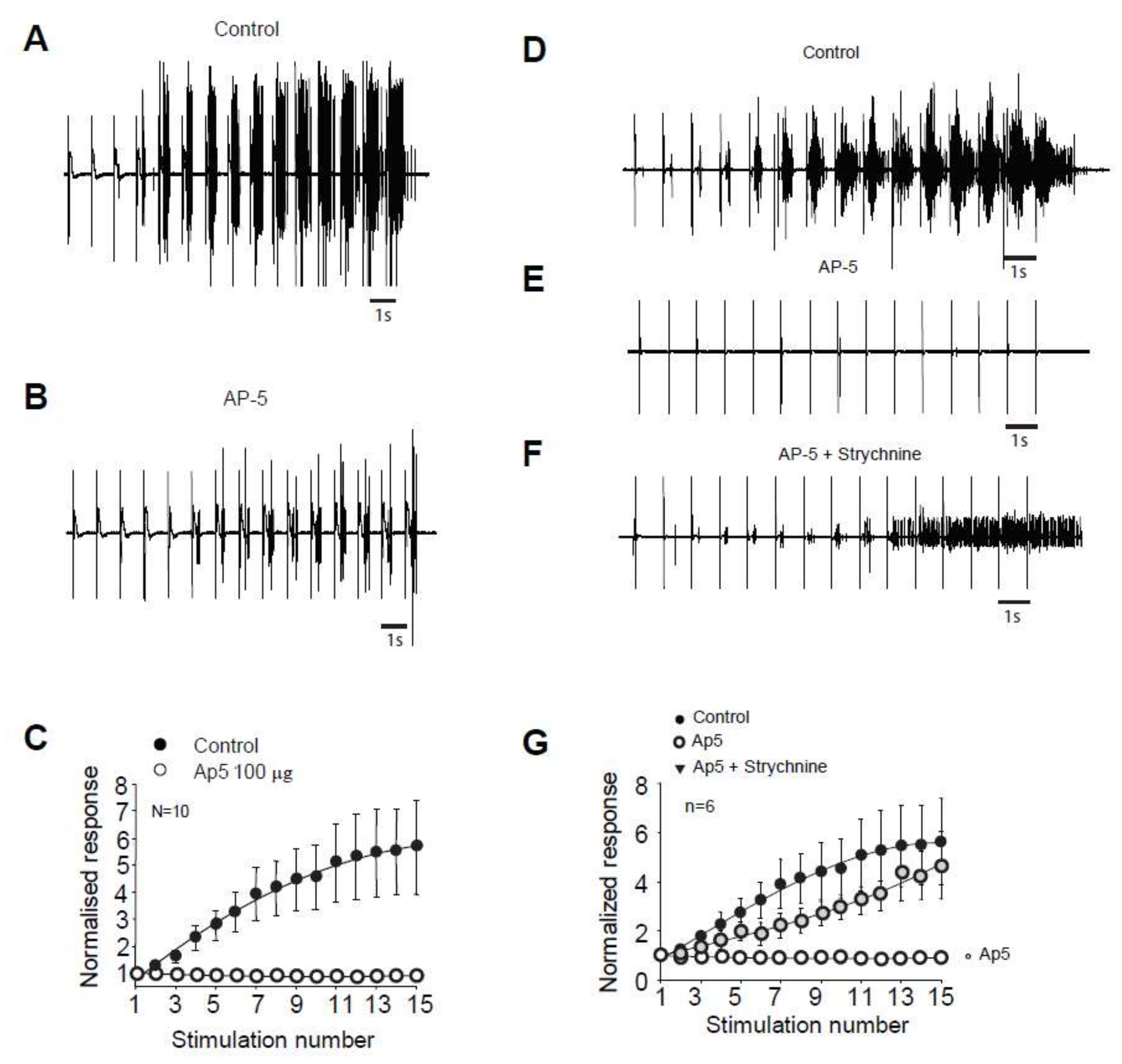
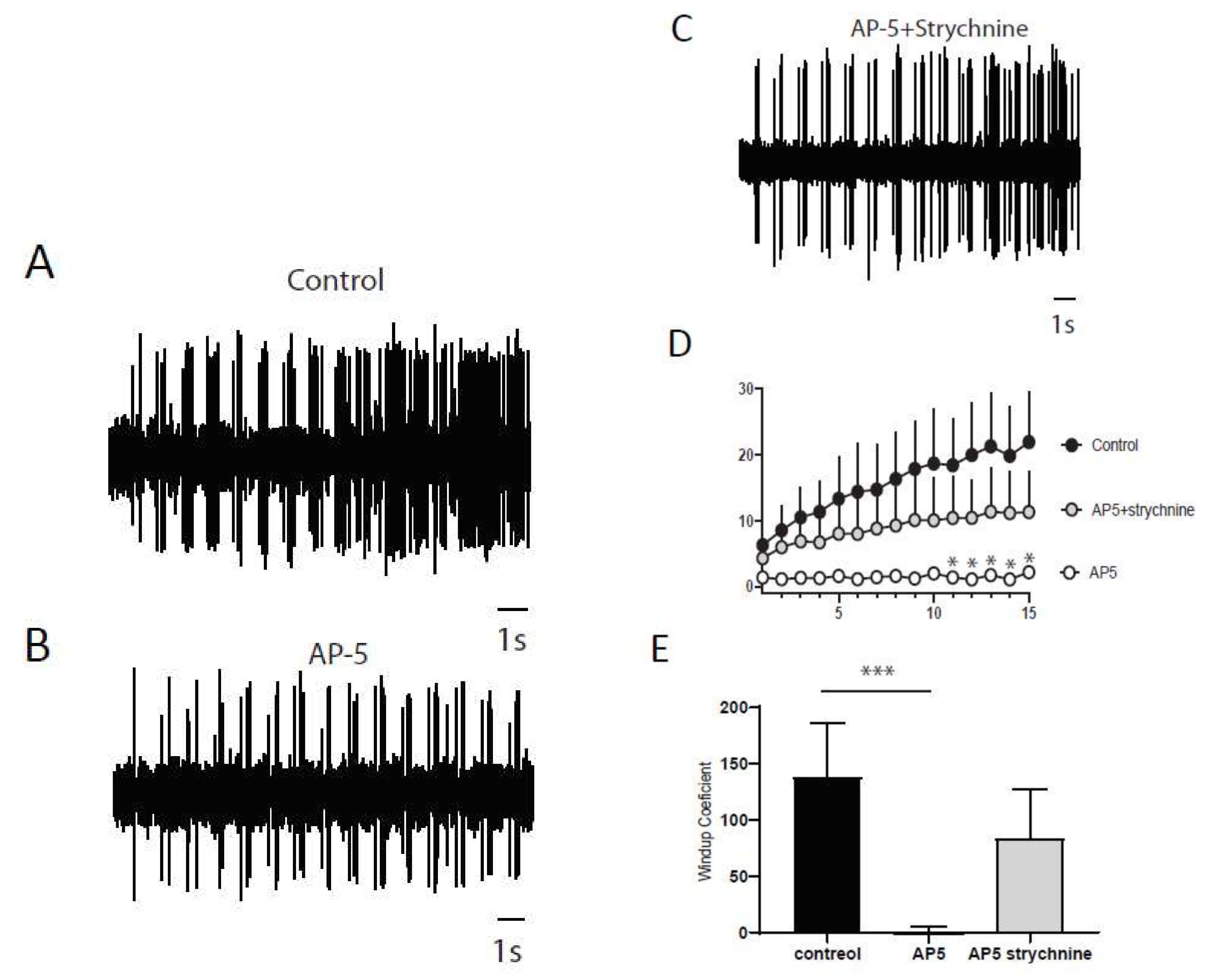
2.2. Windup of the Nociceptive Flexion Reflex in Adult Rats is Controlled by Synaptic and Intrinsic Components
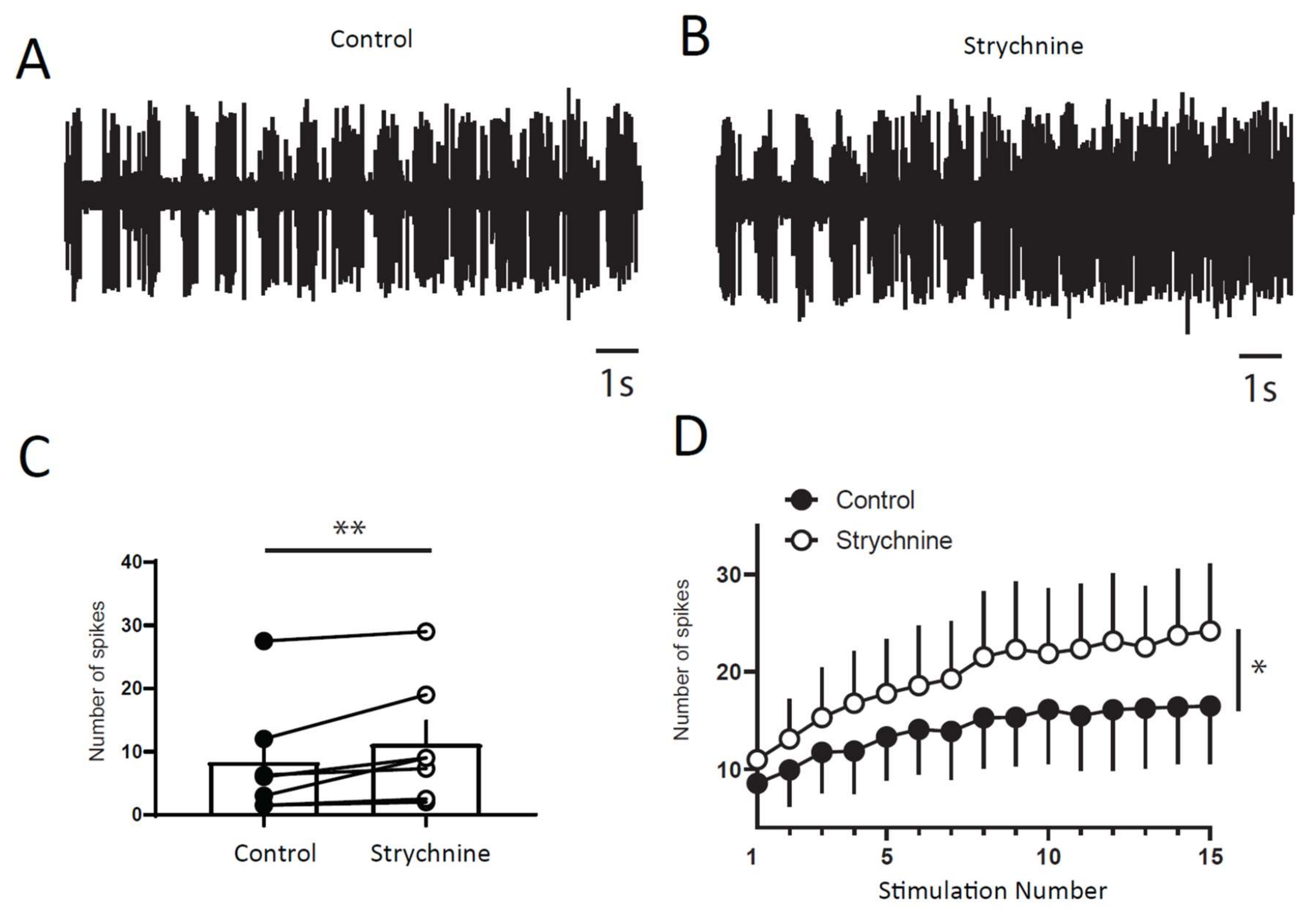
2.3. Windup of the Flexion Reflex Is Sensitive to IL Blockers in Adult Rats
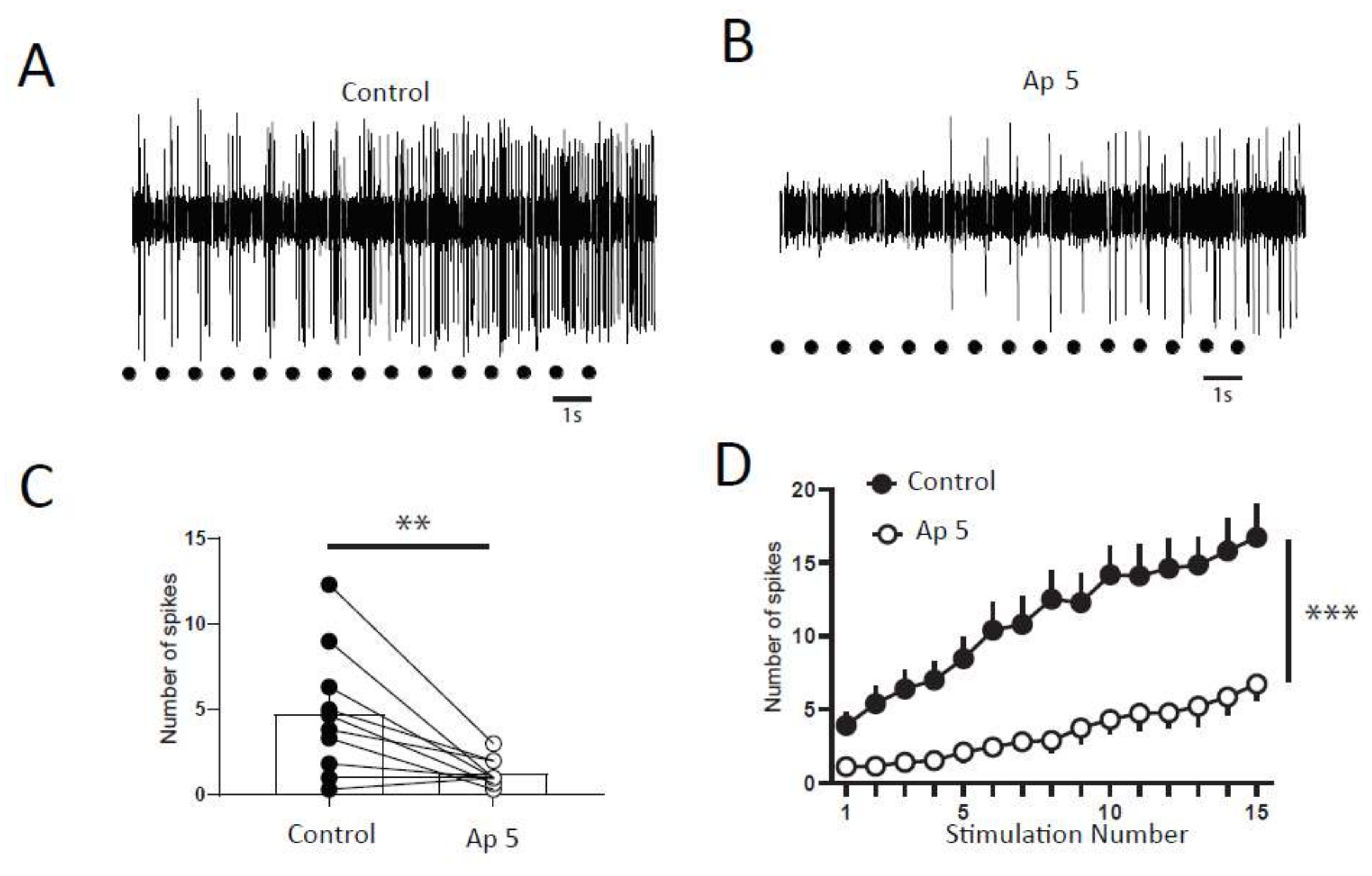
2.4. Synaptic Component of Windup in DHNs of Adult Rats
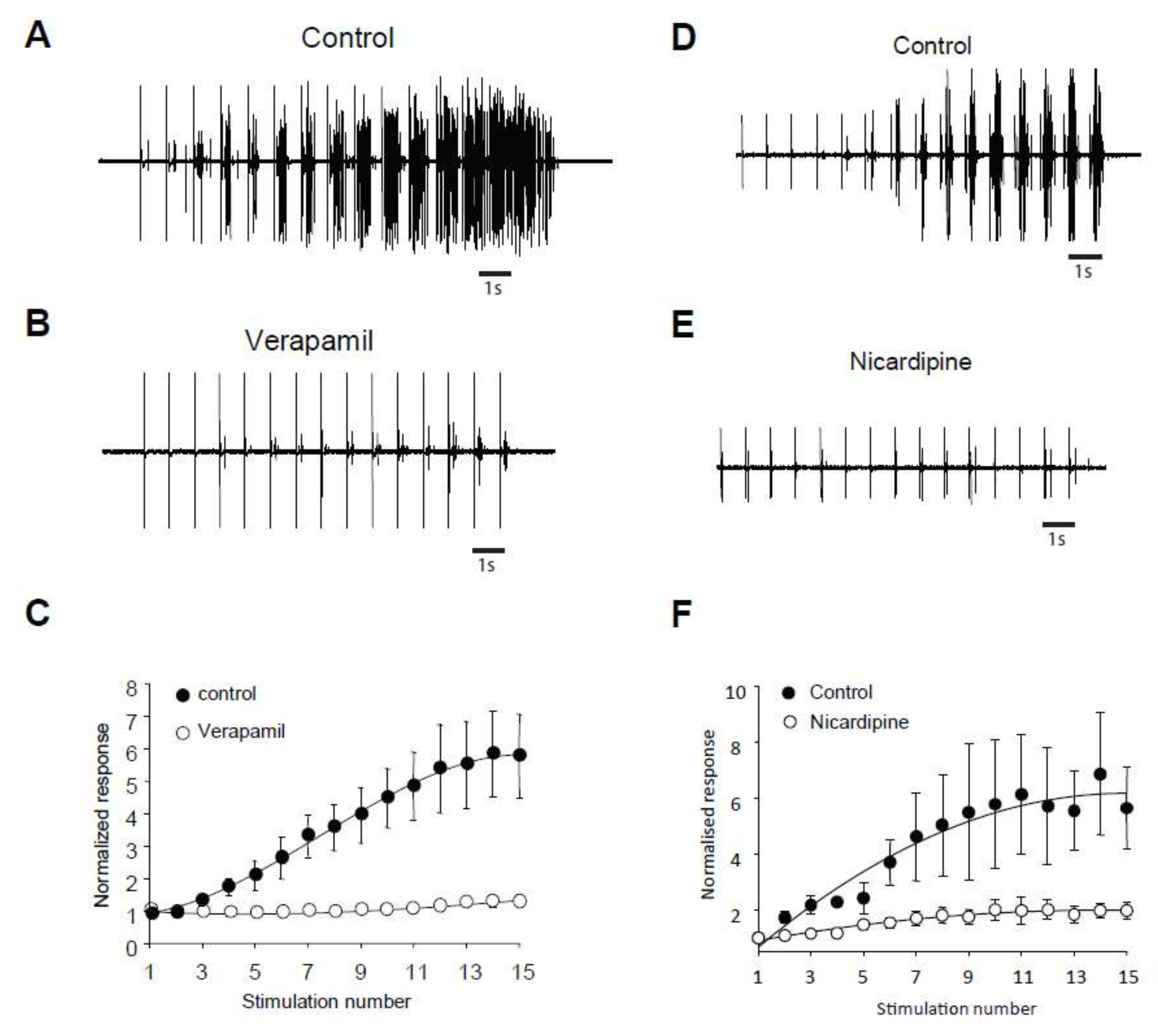
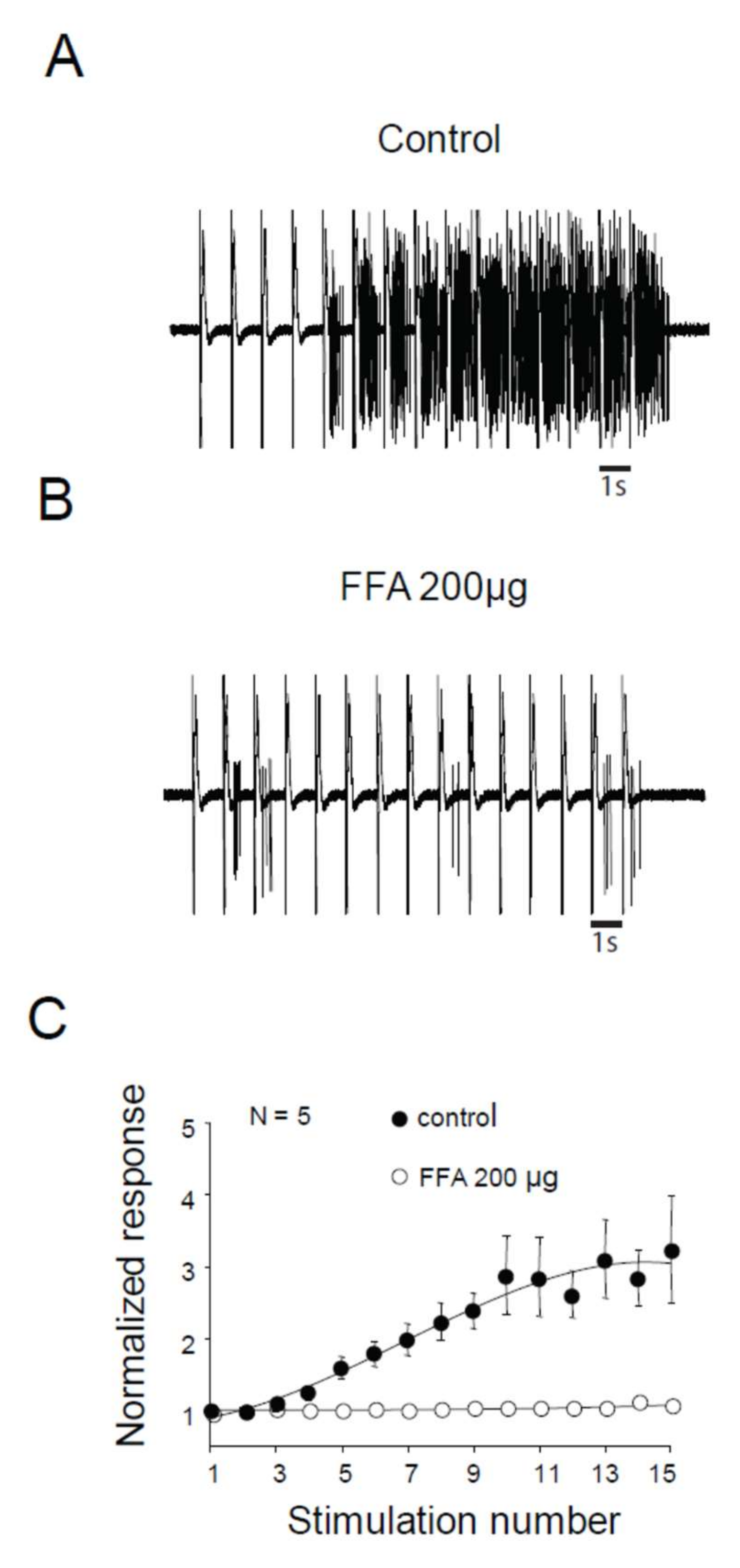
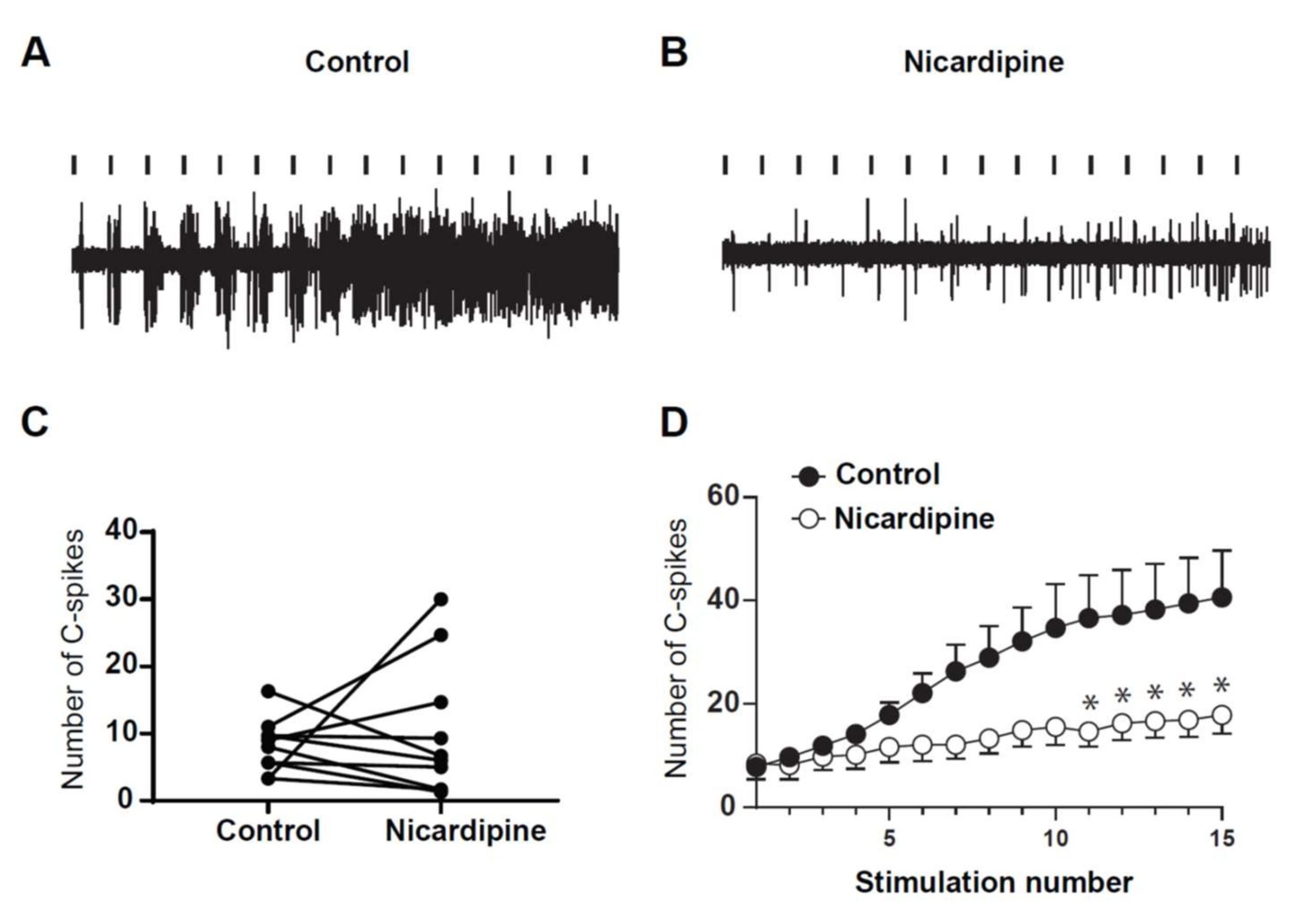
2.5. LTCs Component of Windup in DHN in Adult Rats
3. Discussion
3.1. Adult and Juvenile Windup
3.2. Importance of LTCs in Windup
3.3. Synaptic Component of Windup
3.4. Neural Substrate of Windup
3.5. Windup and Central Sensitization to Pain
4. Methods
4.1. Experimental Procedures
4.2. Drug Administration
4.3. In Vivo Electromyographic Recordings
4.4. In Vivo Extracellular Recordings
4.5. Windup Protocol
4.6. Data Analysis and Statistics
Author Contributions
Funding
Conflicts of Interest
References
- Li, J.; Simone, D.A.; Larson, A.A. Windup leads to characteristics of central sensitization. Pain 1999, 79, 75–82. [Google Scholar] [CrossRef]
- Ji, R.R.; Kohno, T.; Moore, K.A.; Woolf, C.J. Central sensitization and LTP: Do pain and memory share similar mechanisms? Trends Neurosci. 2003, 26, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Baranauskas, G.; Nistri, A. Sensitization of pain pathways in the spinal cord: Cellular mechanisms. Prog. Neurobiol. 1998, 54, 349–365. [Google Scholar] [CrossRef]
- Herrero, J.F.; Laird, J.M.; Lopez-Garcia, J.A. Wind-up of spinal cord neurones and pain sensation: Much ado about something? Prog. Neurobiol. 2000, 61, 169–203. [Google Scholar] [CrossRef]
- Price, D.D.; Staud, R.; Robinson, M.E.; Mauderli, A.P.; Cannon, R.; Vierck, C.J. Enhanced temporal summation of second pain and its central modulation in fibromyalgia patients. Pain 2002, 99, 49–59. [Google Scholar] [CrossRef]
- Staud, R.; Vierck, C.J.; Robinson, M.E.; Price, D.D. Spatial summation of heat pain within and across dermatomes in fibromyalgia patients and pain-free subjects. Pain 2004, 111, 342–350. [Google Scholar] [CrossRef]
- Gozariu, M.; Bragard, D.; Willer, J.C.; Le Bars, D. Temporal summation of C-fiber afferent inputs: Competition between facilitatory and inhibitory effects on C-fiber reflex in the rat. J. Neurophysiol. 1997, 78, 3165–3179. [Google Scholar] [CrossRef]
- Russo, R.E.; Hounsgaard, J. Short-term plasticity in turtle dorsal horn neurons mediated by L-type Ca2+ channels. Neuroscience 1994, 61, 191–197. [Google Scholar] [CrossRef]
- Baranauskas, G.; Nistri, A. NMDA receptor-independent mechanisms responsible for the rate of rise of cumulative depolarization evoked by trains of dorsal root stimuli on rat spinal motoneurones. Brain Res. 1996, 738, 329–332. [Google Scholar] [CrossRef]
- Woolf, C.J.; Thompson, S.W. The induction and maintenance of central sensitization is dependent on N-methyl-D-aspartic acid receptor activation; implications for the treatment of post-injury pain hypersensitivity states. Pain 1991, 44, 293–299. [Google Scholar] [CrossRef]
- Fossat, P.; Sibon, I.; Le Masson, G.; Landry, M.; Nagy, F. L-type calcium channels and NMDA receptors: A determinant duo for short-term nociceptive plasticity. Eur. J. Neurosci. 2007, 25, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Morisset, V.; Nagy, F. Plateau potential-dependent windup of the response to primary afferent stimuli in rat dorsal horn neurons. Eur. J. Neurosci. 2000, 12, 3087–3095. [Google Scholar] [CrossRef] [PubMed]
- Radwani, H.; Lopez-Gonzalez, M.J.; Cattaert, D.; Roca-Lapirot, O.; Dobremez, E.; Bouali-Benazzouz, R.; Eiriksdottir, E.; Langel, U.; Favereaux, A.; Errami, M.; et al. Cav1.2 and Cav1.3 L-type calcium channels independently control short- and long-term sensitization to pain. J. Physiol. 2016, 594, 6607–6626. [Google Scholar] [CrossRef] [PubMed]
- Roca-Lapirot, O.; Radwani, H.; Aby, F.; Nagy, F.; Landry, M.; Fossat, P. Calcium signalling through L-type calcium channels: Role in pathophysiology of spinal nociceptive transmission. Br. J. Pharmacol. 2018, 175, 2362–2374. [Google Scholar] [CrossRef]
- Johnson, K.P.; Tran, S.M.; Siegrist, E.A.; Paidimarri, K.B.; Elson, M.S.; Berkowitz, A. Turtle Flexion Reflex Motor Patterns Show Windup, Mediated Partly by L-type Calcium Channels. Front. Neural Circuits 2017, 11, 83. [Google Scholar] [CrossRef]
- Urch, C.E.; Rahman, W.; Dickenson, A.H. Electrophysiological studies on the role of the NMDA receptor in nociception in the developing rat spinal cord. Brain Res. Dev. Brain Res. 2001, 126, 81–89. [Google Scholar] [CrossRef]
- Ma, Q.P.; Woolf, C.J. Tachykinin NK1 receptor antagonist RP67580 attenuates progressive hypersensitivity of flexor reflex during experimental inflammation in rats. Eur. J. Pharmacol. 1997, 322, 165–171. [Google Scholar] [CrossRef]
- Nagy, I.; Miller, B.A.; Woolf, C.J. NK1 and NK2 receptors contribute to C-fibre evoked slow potentials in the spinal cord. Neuroreport 1994, 5, 2105–2108. [Google Scholar] [CrossRef]
- Morisset, V.; Nagy, F. Ionic basis for plateau potentials in deep dorsal horn neurons of the rat spinal cord. J. Neurosci. 1999, 19, 7309–7316. [Google Scholar] [CrossRef]
- Diaz, A.; Dickenson, A.H. Blockade of spinal N- and P-type, but not L-type, calcium channels inhibits the excitability of rat dorsal horn neurones produced by subcutaneous formalin inflammation. Pain 1997, 69, 93–100. [Google Scholar] [CrossRef]
- Vanegas, H.; Schaible, H. Effects of antagonists to high-threshold calcium channels upon spinal mechanisms of pain, hyperalgesia and allodynia. Pain 2000, 85, 9–18. [Google Scholar] [CrossRef]
- Fossat, P.; Dobremez, E.; Bouali-Benazzouz, R.; Favereaux, A.; Bertrand, S.S.; Kilk, K.; Leger, C.; Cazalets, J.R.; Langel, U.; Landry, M.; et al. Knockdown of L calcium channel subtypes: Differential effects in neuropathic pain. J. Neurosci. 2010, 30, 1073–1085. [Google Scholar] [CrossRef]
- Thompson, S.W.; King, A.E.; Woolf, C.J. Activity-Dependent Changes in Rat Ventral Horn Neurons in vitro; Summation of Prolonged Afferent Evoked Postsynaptic Depolarizations Produce a d-2-Amino-5-Phosphonovaleric Acid Sensitive Windup. Eur. J. Neurosci. 1990, 2, 638–649. [Google Scholar] [CrossRef] [PubMed]
- Woolf, C.J. Windup and central sensitization are not equivalent. Pain 1996, 66, 105–108. [Google Scholar] [CrossRef]
- Chapman, V.; Suzuki, R.; Dickenson, A.H. Electrophysiological characterization of spinal neuronal response properties in anaesthetized rats after ligation of spinal nerves L5-L6. J. Physiol. 1998, 507 Pt 3, 881–894. [Google Scholar] [CrossRef]
- Reali, C.; Fossat, P.; Landry, M.; Russo, R.E.; Nagy, F. Intrinsic membrane properties of spinal dorsal horn neurones modulate nociceptive information processing in vivo. J. Physiol. 2011, 589 Pt 11, 2733–2743. [Google Scholar] [CrossRef]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aby, F.; Bouali-Benazzouz, R.; Landry, M.; Fossat, P. Windup of Nociceptive Flexion Reflex Depends on Synaptic and Intrinsic Properties of Dorsal Horn Neurons in Adult Rats. Int. J. Mol. Sci. 2019, 20, 6146. https://doi.org/10.3390/ijms20246146
Aby F, Bouali-Benazzouz R, Landry M, Fossat P. Windup of Nociceptive Flexion Reflex Depends on Synaptic and Intrinsic Properties of Dorsal Horn Neurons in Adult Rats. International Journal of Molecular Sciences. 2019; 20(24):6146. https://doi.org/10.3390/ijms20246146
Chicago/Turabian StyleAby, Franck, Rabia Bouali-Benazzouz, Marc Landry, and Pascal Fossat. 2019. "Windup of Nociceptive Flexion Reflex Depends on Synaptic and Intrinsic Properties of Dorsal Horn Neurons in Adult Rats" International Journal of Molecular Sciences 20, no. 24: 6146. https://doi.org/10.3390/ijms20246146
APA StyleAby, F., Bouali-Benazzouz, R., Landry, M., & Fossat, P. (2019). Windup of Nociceptive Flexion Reflex Depends on Synaptic and Intrinsic Properties of Dorsal Horn Neurons in Adult Rats. International Journal of Molecular Sciences, 20(24), 6146. https://doi.org/10.3390/ijms20246146