PTX3 Modulates Neovascularization and Immune Inflammatory Infiltrate in a Murine Model of Fibrosarcoma

 ,
,  ,
, 
 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. In Vivo Procedures
2.3. Immunohistochemistry
2.4. Morphometric Analysis
2.5. Statistical Analysis
3. Results
4. Discussion
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| FGF-2 | fibroblast growth factor-2 |
| IHC | immunohistochemistry |
| PTX3 | Long pentraxin-3 |
| s.c. | subcutaneously |
| Tie2-hPTX3 | transgenic mice |
| WT | wild type |
References
- Folpe, A.L. Fibrosarcoma: A review and update. Histopathology 2014, 64, 12–25. [Google Scholar] [CrossRef] [PubMed]
- Augsburger, D.; Nelson, P.J.; Kalinski, T.; Udelnow, A.; Knosel, T.; Hofstetter, M.; Qin, J.W.; Wang, Y.; Gupta, A.S.; Bonifatius, S.; et al. Current diagnostics and treatment of fibrosarcoma -perspectives for future therapeutic targets and strategies. Oncotarget 2017, 8, 104638–104653. [Google Scholar] [CrossRef] [PubMed]
- Kandel, J.; Bossy-Wetzel, E.; Radvanyi, F.; Klagsbrun, M.; Folkman, J.; Hanahan, D. Neovascularization is associated with a switch to the export of bFGF in the multistep development of fibrosarcoma. Cell 1991, 66, 1095–1104. [Google Scholar] [CrossRef]
- Garlanda, C.; Bottazzi, B.; Bastone, A.; Mantovani, A. Pentraxins at the crossroads between innate immunity, inflammation, matrix deposition, and female fertility. Annu. Rev. Immunol. 2005, 23, 337–366. [Google Scholar] [CrossRef] [PubMed]
- Camozzi, M.; Rusnati, M.; Bugatti, A.; Bottazzi, B.; Mantovani, A.; Bastone, A.; Inforzato, A.; Vincenti, S.; Bracci, L.; Mastroianni, D.; et al. Identification of an antiangiogenic FGF2-binding site in the N terminus of the soluble pattern recognition receptor PTX3. J. Biol. Chem. 2006, 281, 22605–22613. [Google Scholar] [CrossRef] [PubMed]
- Leali, D.; Alessi, P.; Coltrini, D.; Rusnati, M.; Zetta, L.; Presta, M. Fibroblast growth factor-2 antagonist and antiangiogenic activity of long-pentraxin 3-derived synthetic peptides. Curr. Pharm. Des. 2009, 15, 3577–3589. [Google Scholar] [CrossRef]
- Presta, M.; Camozzi, M.; Salvatori, G.; Rusnati, M. Role of the soluble pattern recognition receptor PTX3 in vascular biology. J. Cell. Mol. Med. 2007, 11, 723–738. [Google Scholar] [CrossRef]
- Leali, D.; Alessi, P.; Coltrini, D.; Ronca, R.; Corsini, M.; Nardo, G.; Indraccolo, S.; Presta, M. Long pentraxin-3 inhibits FGF8b-dependent angiogenesis and growth of steroid hormone-regulated tumors. Mol. Cancer Ther. 2011, 10, 1600–1610. [Google Scholar] [CrossRef]
- Ronca, R.; Alessi, P.; Coltrini, D.; Di Salle, E.; Giacomini, A.; Leali, D.; Corsini, M.; Belleri, M.; Tobia, C.; Garlanda, C.; et al. Long pentraxin-3 as an epithelial-stromal fibroblast growth factor-targeting inhibitor in prostate cancer. J. Pathol. 2013, 230, 228–238. [Google Scholar] [CrossRef]
- Ronca, R.; Di Salle, E.; Giacomini, A.; Leali, D.; Alessi, P.; Coltrini, D.; Ravelli, C.; Matarazzo, S.; Ribatti, D.; Vermi, W.; et al. Long pentraxin-3 inhibits epithelial-mesenchymal transition in melanoma cells. Mol. Cancer Ther. 2013, 12, 2760–2771. [Google Scholar] [CrossRef]
- Giacomini, A.; Ghedini, G.C.; Presta, M.; Ronca, R. Long pentraxin 3: A novel multifaceted player in cancer. Biochim. Biophys. Acta Rev. Cancer 2018, 1869, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Ronca, R.; Giacomini, A.; Di Salle, E.; Coltrini, D.; Pagano, K.; Ragona, L.; Matarazzo, S.; Rezzola, S.; Maiolo, D.; Torrella, R.; et al. Long-Pentraxin 3 Derivative as a Small-Molecule FGF Trap for Cancer Therapy. Cancer Cell 2015, 28, 225–239. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, P.F.; Matarazzo, S.; Maccarinelli, F.; Foglio, E.; Giacomini, A.; Silva Nunes, J.P.; Presta, M.; Dias, A.A.M.; Ronca, R. Long Pentraxin 3-Mediated Fibroblast Growth Factor Trapping Impairs Fibrosarcoma Growth. Front. Oncol. 2018, 8, 472. [Google Scholar] [CrossRef] [PubMed]
- Ronca, R.; Tamma, R.; Coltrini, D.; Ruggieri, S.; Presta, M.; Ribatti, D. Fibroblast growth factor modulates mast cell recruitment in a murine model of prostate cancer. Oncotarget 2017, 8, 82583–82592. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D. Mast cells and macrophages exert beneficial and detrimental effects on tumor progression and angiogenesis. Immunol. Lett. 2013, 152, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Bonavita, E.; Gentile, S.; Rubino, M.; Maina, V.; Papait, R.; Kunderfranco, P.; Greco, C.; Feruglio, F.; Molgora, M.; Laface, I.; et al. PTX3 is an extrinsic oncosuppressor regulating complement-dependent inflammation in cancer. Cell 2015, 160, 700–714. [Google Scholar] [CrossRef] [PubMed]
- Ribatti, D.V.A. Overview of angiogenesis during tumor growth. In Angiogenesis. An Integrative Approach. From Science to Medicine; Figg, W.D., Folkman, J., Eds.; Springer: New York, NY, USA, 2008; pp. 161–168. [Google Scholar]
- Ronca, R.; Van Ginderachter, J.A.; Turtoi, A. Paracrine interactions of cancer-associated fibroblasts, macrophages and endothelial cells: Tumor allies and foes. Curr. Opin. Oncol. 2018, 30, 45–53. [Google Scholar] [CrossRef]
- Sawicki, B.; Kasacka, I.; Chyczewski, L. Preliminary evaluation of mast cells and angiogenesis processes in experimental fibrosarcoma. Folia Histochem. Cytobiol. 2002, 40, 197–198. [Google Scholar]
- Eisenthal, A.; Schwartz, I.; Issakov, J.; Klausner, Y.; Misonzhnik, F.; Lifschitz-Mercer, B. Immunohistochemistry Evaluation of the Effect in Vivo of Tumor Necrosis Factor (TNF)-alpha on Blood Vessel Density in Murine Fibrosarcoma. Sarcoma 2003, 7, 57–61. [Google Scholar] [CrossRef]
- Jones, R.L.; Katz, D.; Loggers, E.T.; Davidson, D.; Rodler, E.T.; Pollack, S.M. Clinical benefit of antiangiogenic therapy in advanced and metastatic chondrosarcoma. Med. Oncol. 2017, 34, 167. [Google Scholar] [CrossRef][Green Version]
- Doni, A.; Stravalaci, M.; Inforzato, A.; Magrini, E.; Mantovani, A.; Garlanda, C.; Bottazzi, B. The Long Pentraxin PTX3 as a Link Between Innate Immunity, Tissue Remodeling, and Cancer. Front. Immunol. 2019, 10, 712. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
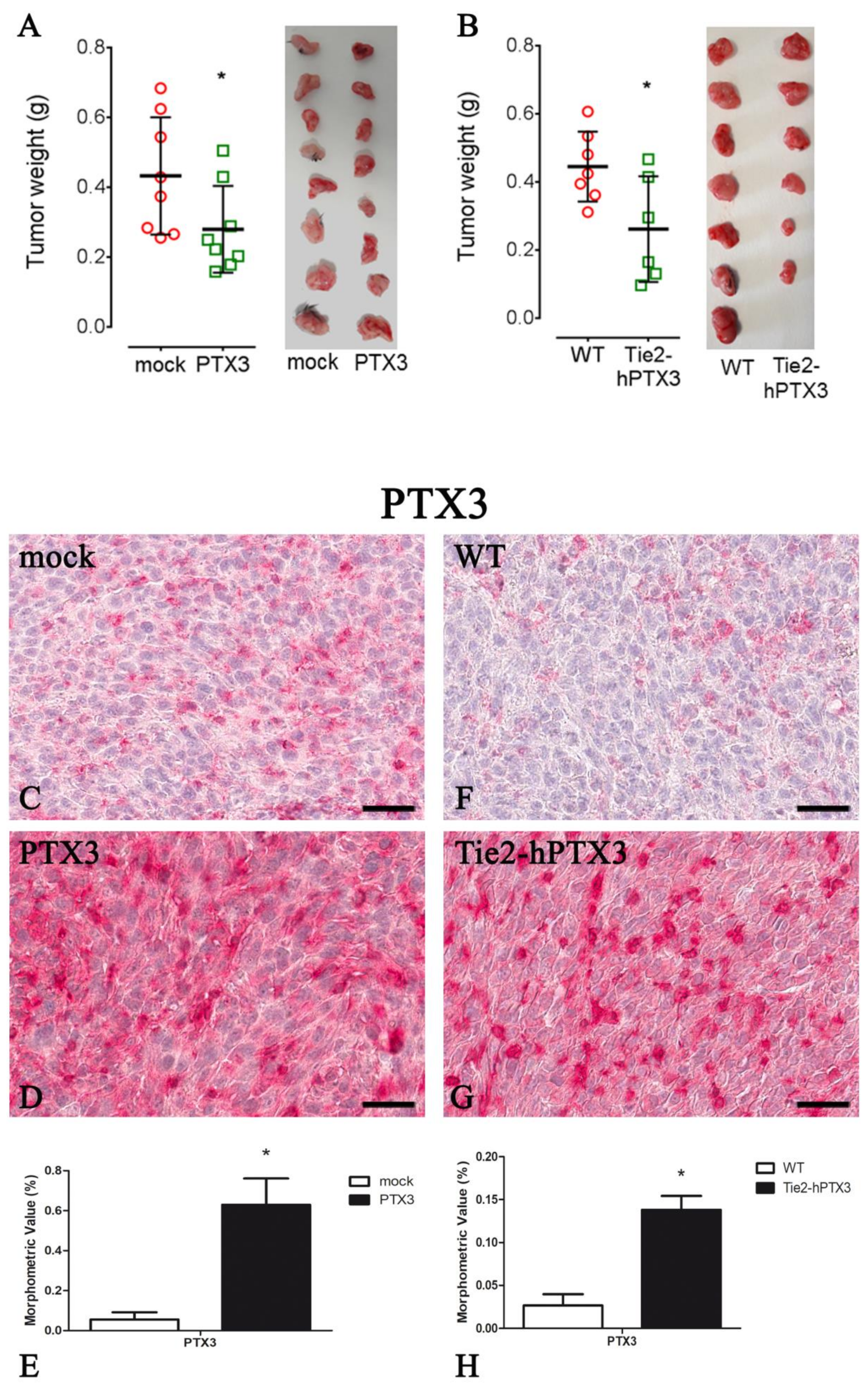
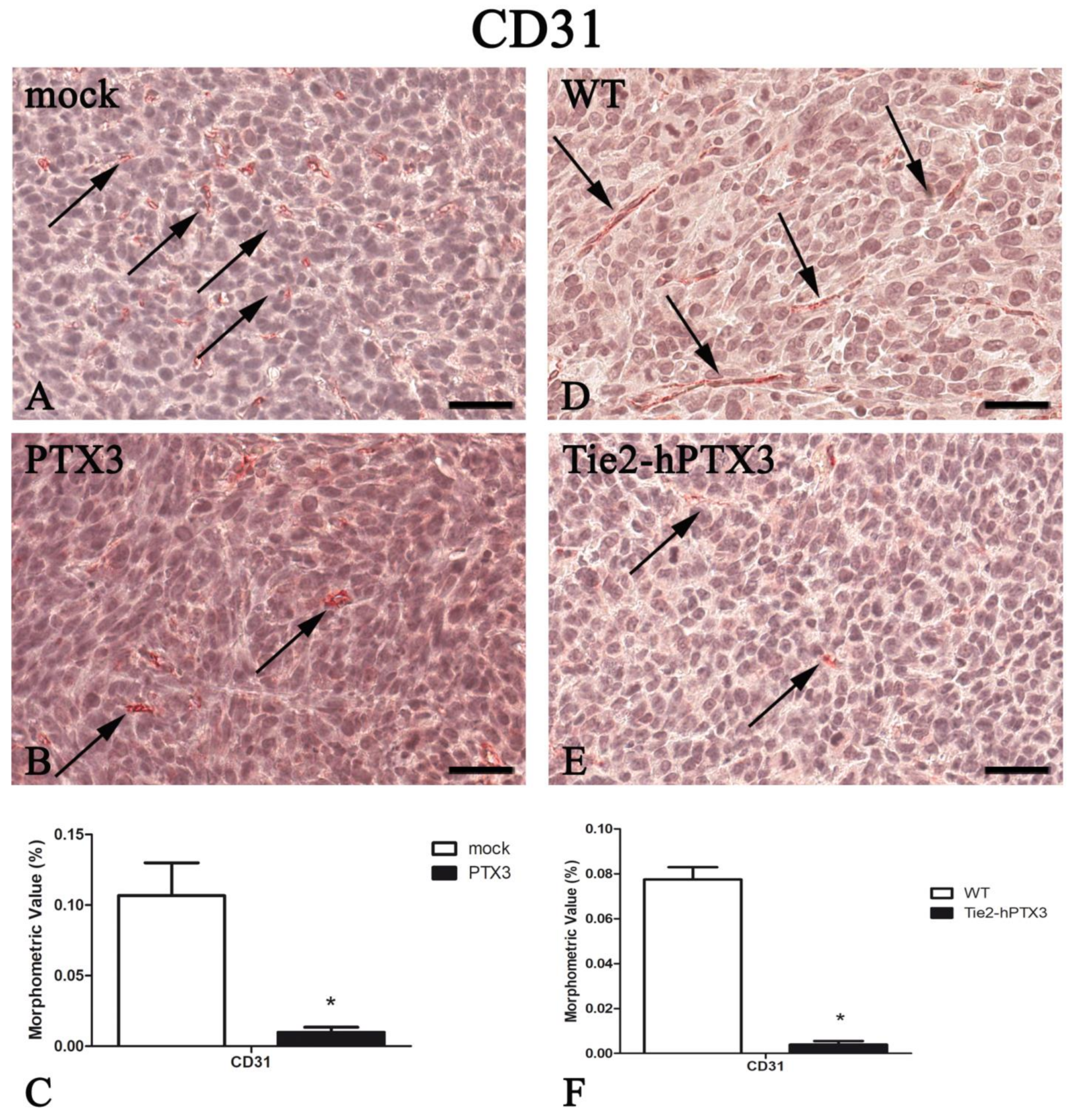
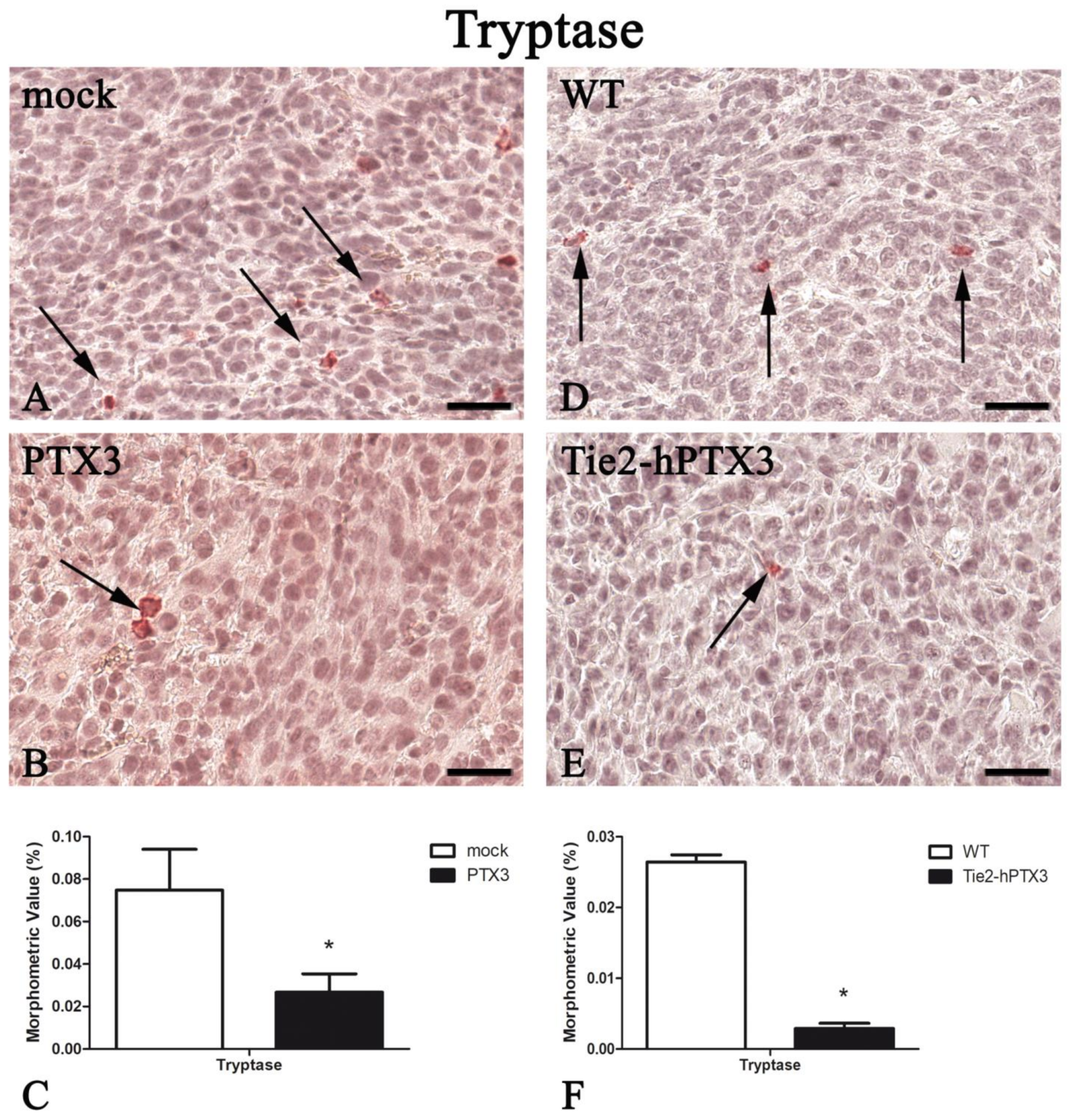
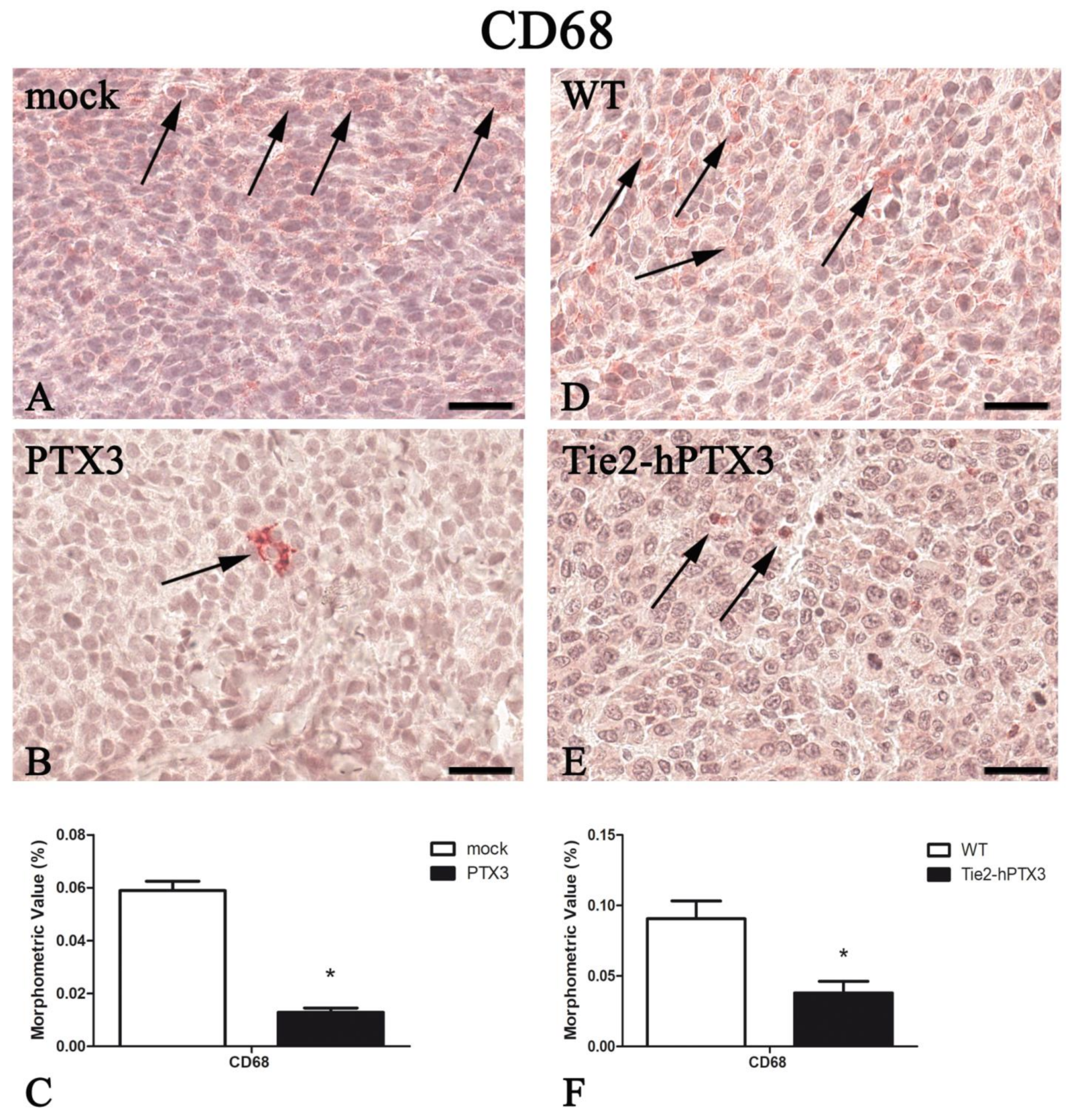
| Protein | PTX3 Overexpression by Cancer Cells | PTX3 Overexpression by Transgenic Animals | ||
|---|---|---|---|---|
| mock | PTX3 | WT | Tie2-hPTX3 | |
| PTX3 | 0.055 ± 0.036 | 0.630 ± 0.131 * | 0.027 ± 0.013 | 0.138 ± 0.016 * |
| CD31 | 0.107 ± 0.023 | 0.010 ± 0.004 * | 0.077 ± 0.005 | 0.004 ± 0.002* |
| Tryptase | 0.075 ± 0.019 | 0.027 ± 0.009 * | 0.026 ± 0.001 | 0.003 ± 0.001* |
| CD68 | 0.059 ± 0.004 | 0.013 ± 0.002 * | 0.091 ± 0.013 | 0.038 ± 0.008* |
| CD4 | 0.060 ± 0.004 | 0.005 ± 0.002 * | 0.054 ± 0.018 | 0.010 ± 0.003* |
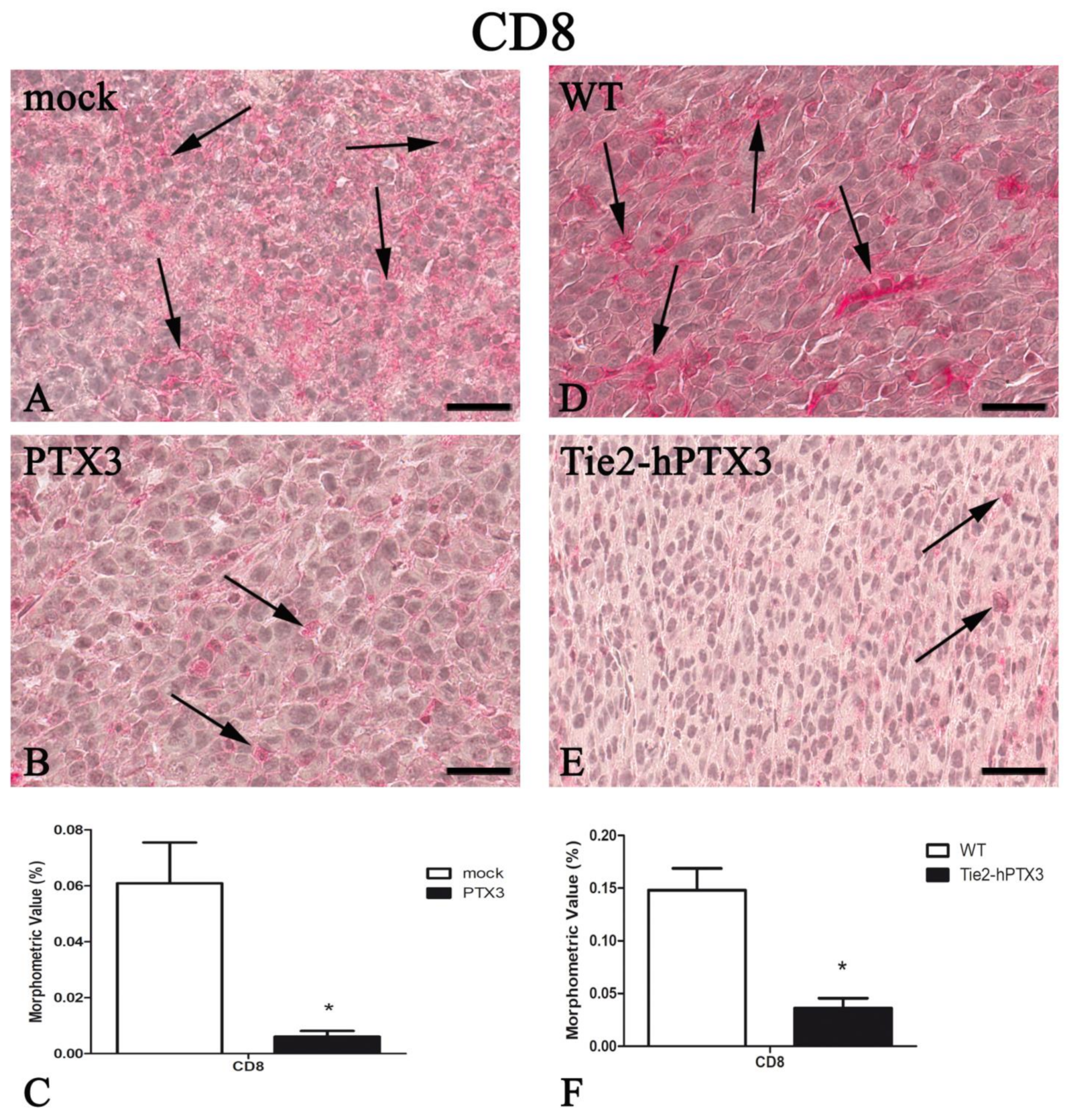
| CD8 | 0.061 ± 0.015 | 0.006 ± 0.002 * | 0.148 ± 0.021 | 0.036 ± 0.010* |
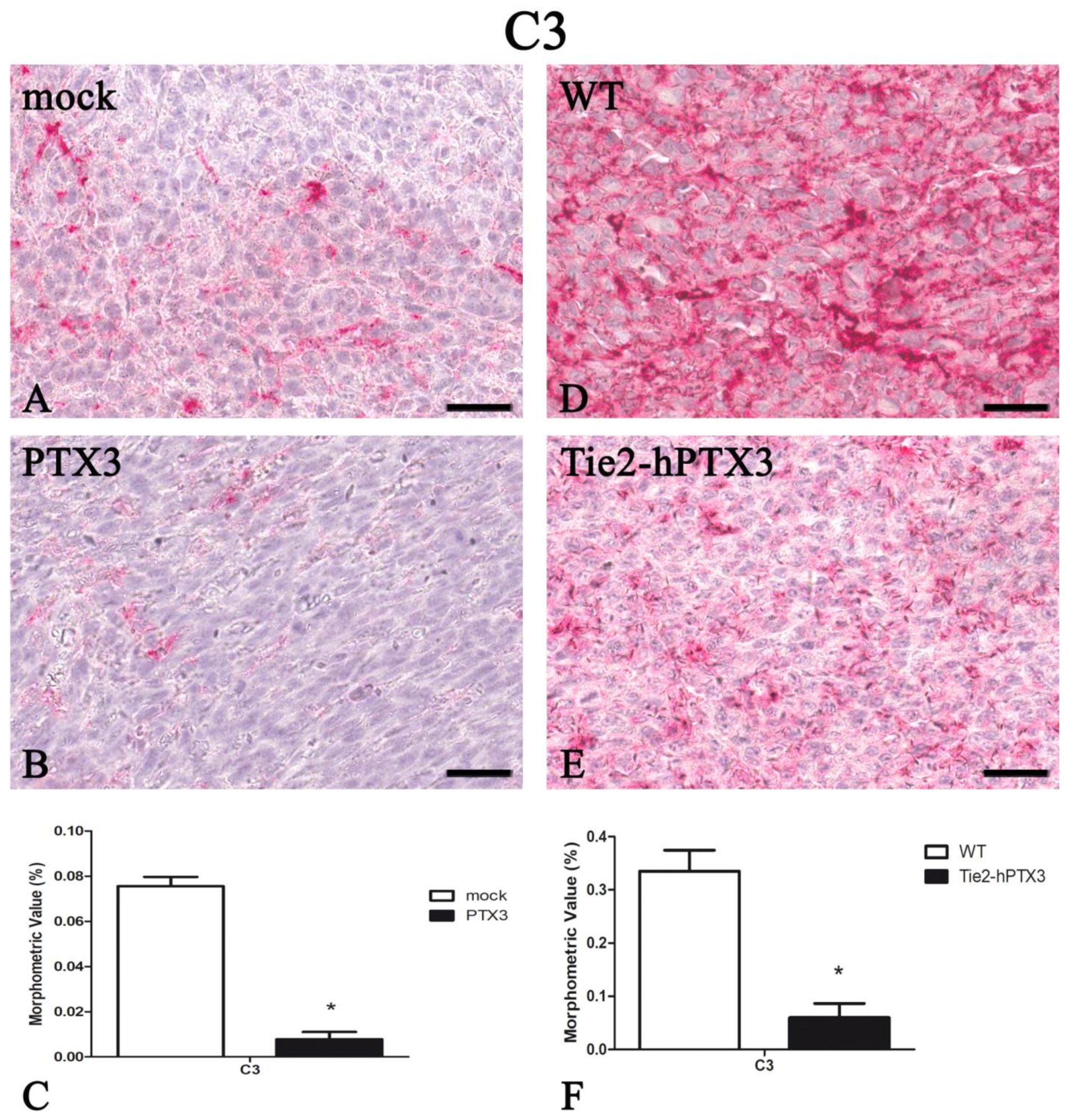
| C3 | 0.076 ± 0.004 | 0.008 ± 0.003 * | 0.335 ± 0.039 | 0.060 ± 0.027* |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Annese, T.; Ronca, R.; Tamma, R.; Giacomini, A.; Ruggieri, S.; Grillo, E.; Presta, M.; Ribatti, D. PTX3 Modulates Neovascularization and Immune Inflammatory Infiltrate in a Murine Model of Fibrosarcoma. Int. J. Mol. Sci. 2019, 20, 4599. https://doi.org/10.3390/ijms20184599
Annese T, Ronca R, Tamma R, Giacomini A, Ruggieri S, Grillo E, Presta M, Ribatti D. PTX3 Modulates Neovascularization and Immune Inflammatory Infiltrate in a Murine Model of Fibrosarcoma. International Journal of Molecular Sciences. 2019; 20(18):4599. https://doi.org/10.3390/ijms20184599
Chicago/Turabian StyleAnnese, Tiziana, Roberto Ronca, Roberto Tamma, Arianna Giacomini, Simona Ruggieri, Elisabetta Grillo, Marco Presta, and Domenico Ribatti. 2019. "PTX3 Modulates Neovascularization and Immune Inflammatory Infiltrate in a Murine Model of Fibrosarcoma" International Journal of Molecular Sciences 20, no. 18: 4599. https://doi.org/10.3390/ijms20184599
APA StyleAnnese, T., Ronca, R., Tamma, R., Giacomini, A., Ruggieri, S., Grillo, E., Presta, M., & Ribatti, D. (2019). PTX3 Modulates Neovascularization and Immune Inflammatory Infiltrate in a Murine Model of Fibrosarcoma. International Journal of Molecular Sciences, 20(18), 4599. https://doi.org/10.3390/ijms20184599