Assembly Mechanism for Aggregation of Amyloid Fibrils

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Single Molecule
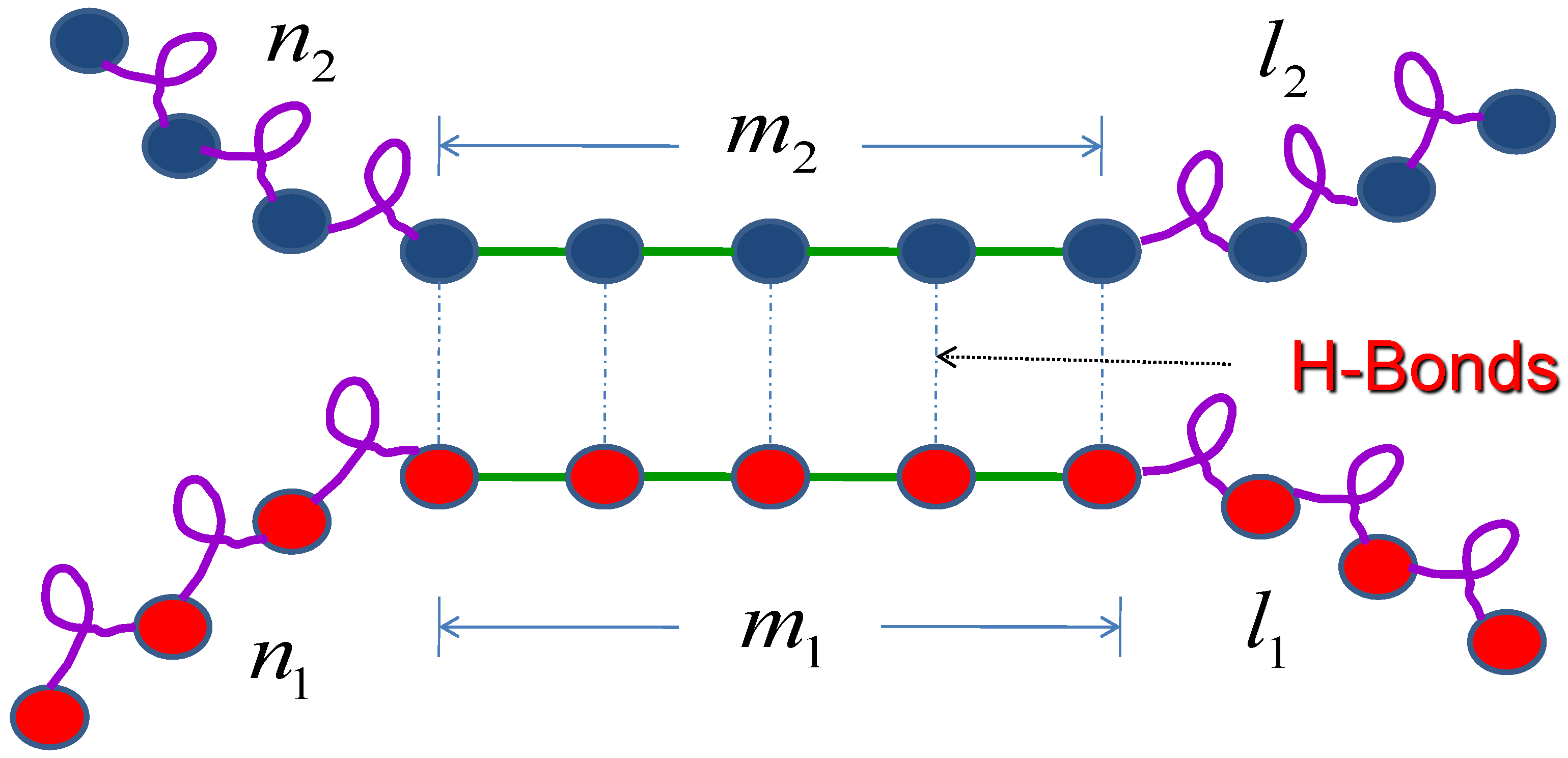
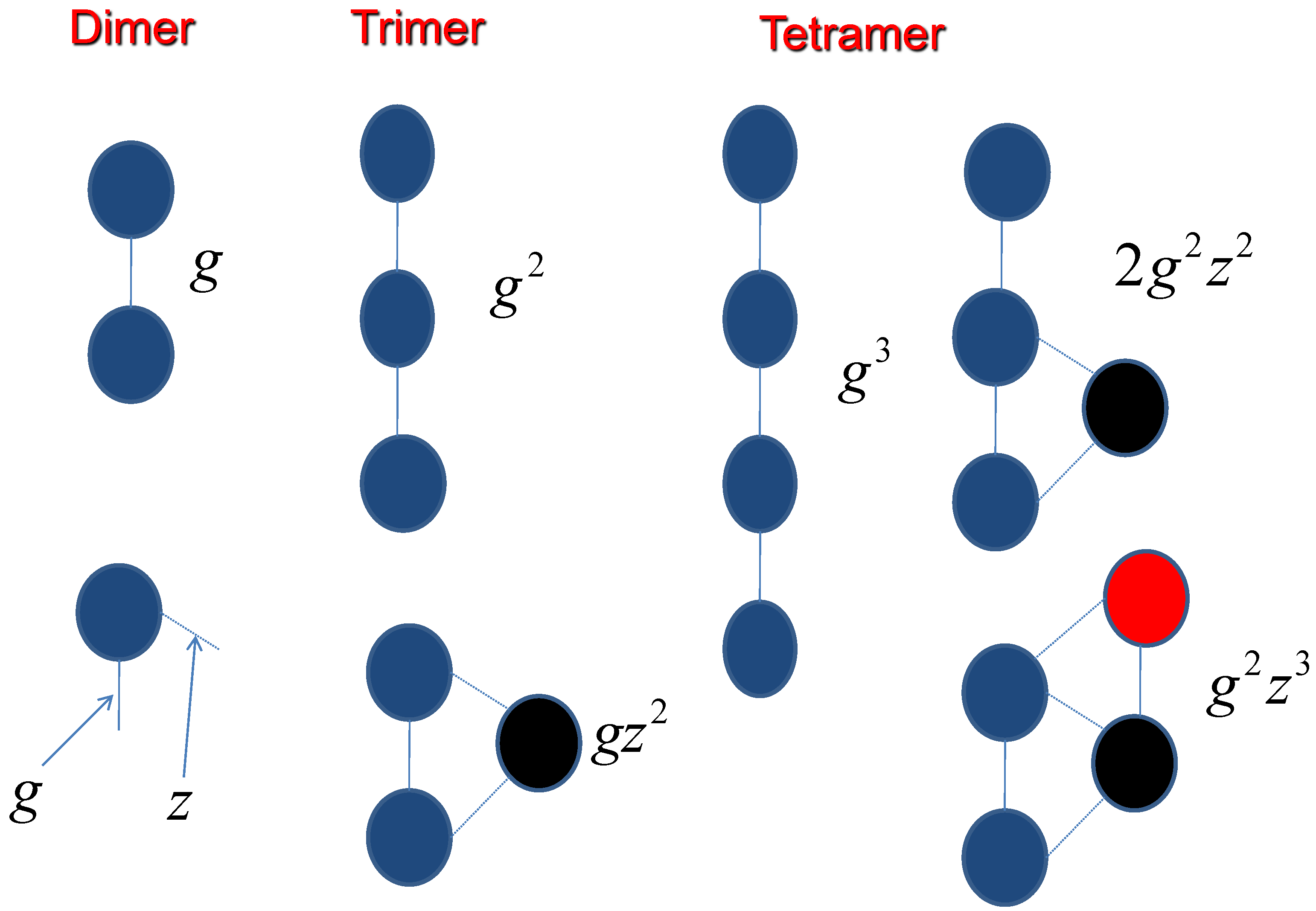
2.2. Two Molecules
2.3. Three Molecules
2.4. General Expression of n Molecules
3. Discussion
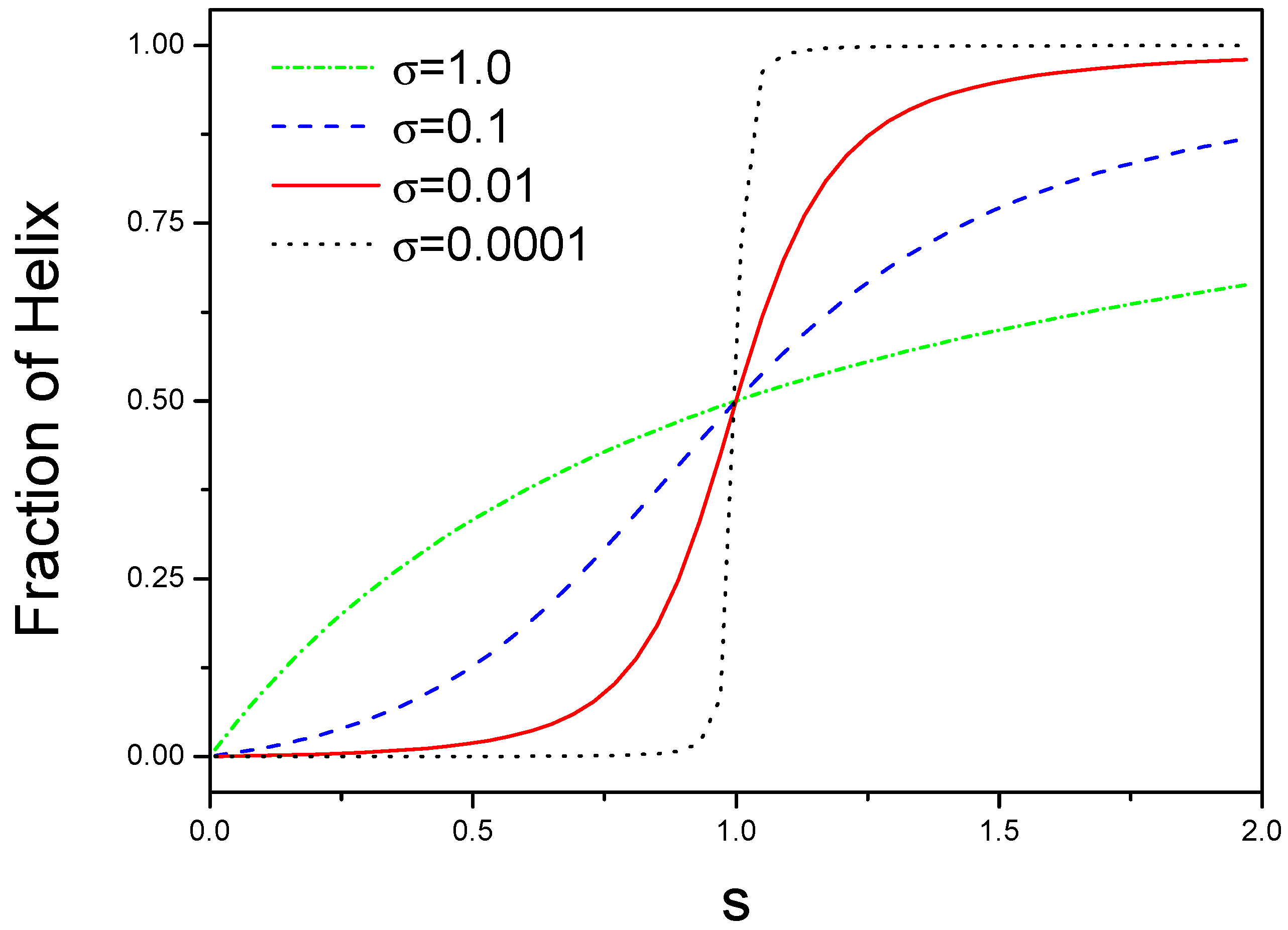
3.1. Molecular Interaction with Denaturant
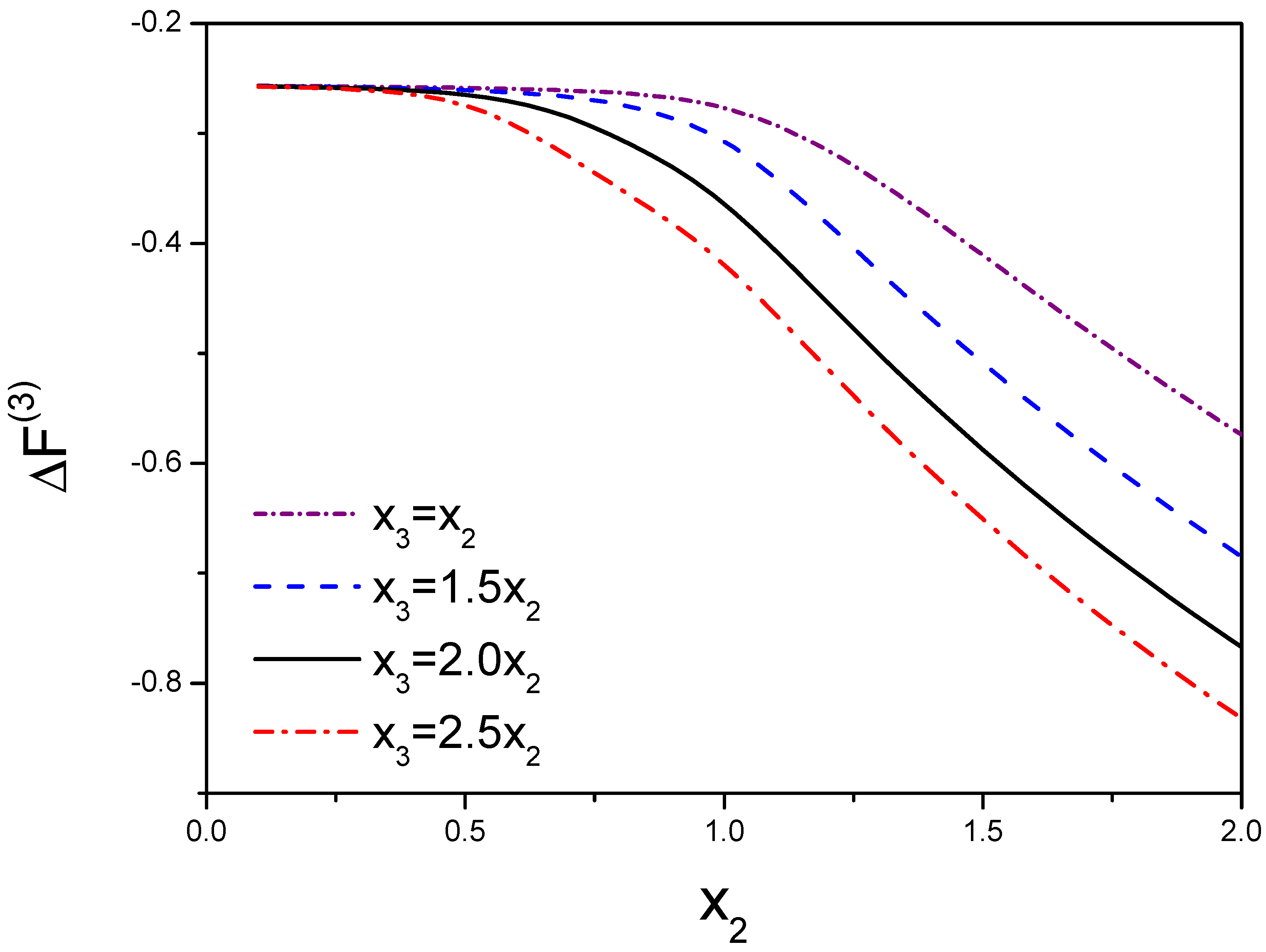
3.2. Free Energy
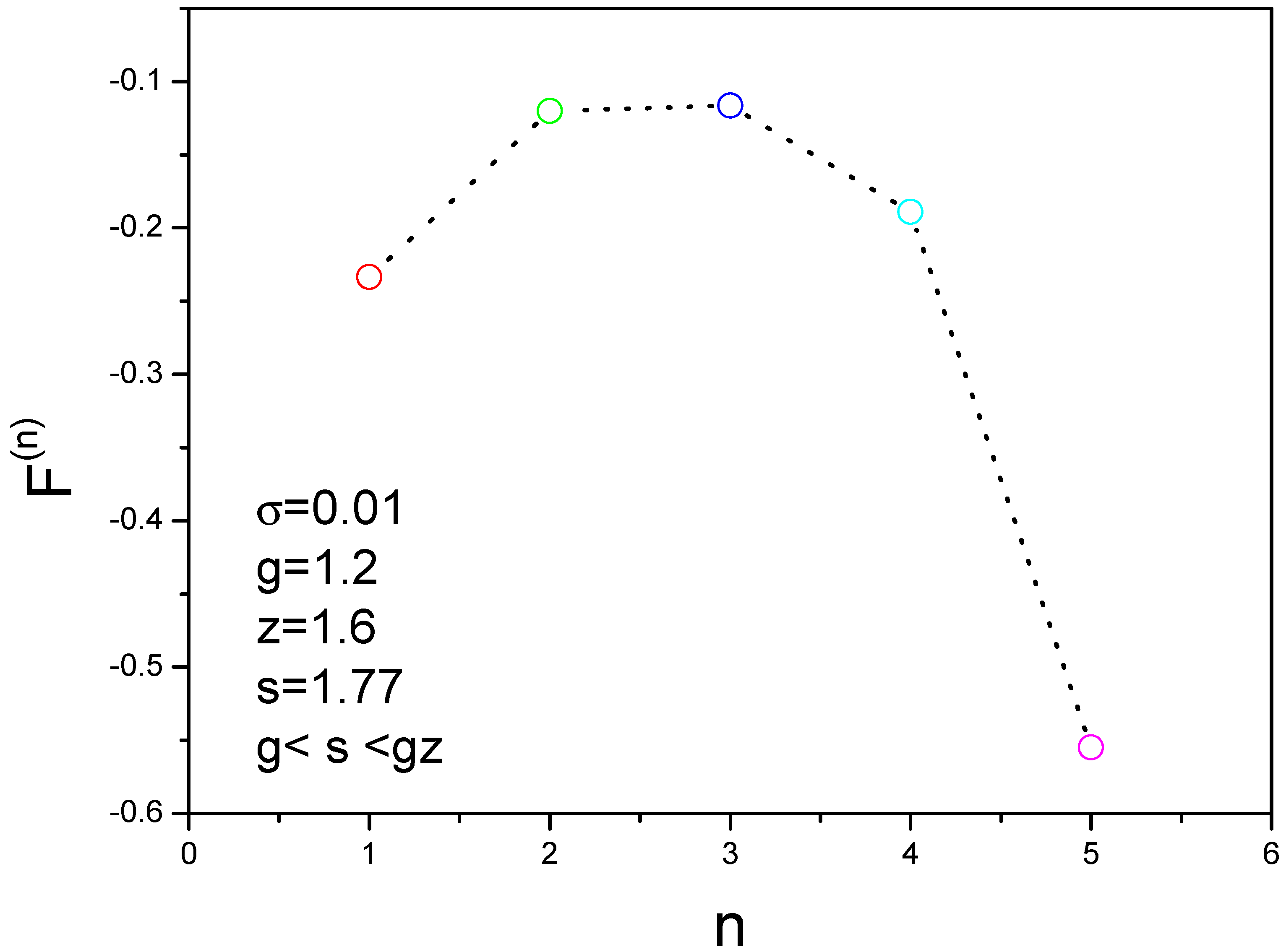
3.3. Peak Location in the 2D Nucleation Rate
3.4. Scaling Behavior of Nucleation Rate in 3D
4. Conclusions
Conflicts of Interest
References
- Nelson, R.; Sawaya, M.R.; Balbirnie, M.; Madsen, A.Ø.; Riekel, C.; Grothe, R.; Eisenberg, D. Structure of the cross-β spine of amyloid-like fibrils. Nature 2005, 435, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Knowles, T.P.J.; Waudby, C.A.; Devlin, G.L.; Cohen, S.I.A.; Aguzzi, A.; Vendruscolo, M.; Terentjev, E.M.; Welland, M.E.; Dobson, C.M. An Analytical Solution to the Kinetics of Breakable Filament Assembly. Science 2009, 326, 1533–1537. [Google Scholar] [CrossRef] [PubMed]
- Seeliger, J.; Estel, K.; Erwin, N.; Winter, R. Cosolvent effects on the fibrillation reaction of human IAPP. Phys. Chem. Chem. Phys. 2013, 15, 8902–8907. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Canchi, D.R.; Garcia, A.E. Cosolvent Effects on Protein Stability. Annu. Rev. Phys. Chem. 2013, 64, 273–293. [Google Scholar] [CrossRef] [PubMed]
- Miller, Y.; Ma, B.; Nussinov, R. Polymorphism in Alzheimer Aβ amyloid organization reflects conformational selection in a rugged energy landscape. Chem. Rev. 2010, 110, 4820–4838. [Google Scholar] [CrossRef] [PubMed]
- Dill, K.A. Theory for the folding and stability of globular proteins. Biochemistry 1985, 24, 1501–1509. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, K.; Dill, K.A. Theory for Protein Folding Cooperativity: Helix Bundles. J. Am. Chem. Soc. 2009, 131, 2306–2312. [Google Scholar] [CrossRef] [PubMed]
- Dobson, C.M. Protein folding and misfolding. Nature 2003, 426, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Carballo-Pacheo, M.; Strodel, B. Advances in the Simulation of Protein Aggregation at the Atomistic Scale. J. Phys. Chem. B 2016, 120, 2991–2999. [Google Scholar] [CrossRef] [PubMed]
- Cheon, M.; Hall, C.K.; Chang, I. Structural Conversion of A β17C42 Peptides from Disordered Oligomers to U-Shape Protofilaments via Multiple Kinetic Pathways. PLoS Comput. Biol. 2015, 11, e1004258. [Google Scholar] [CrossRef] [PubMed]
- Kashchiev, D.; Auer, S. Nucleation of amyloid fibrils. J. Chem. Phys. 2010, 132, 215101. [Google Scholar] [CrossRef] [PubMed]
- Cabriolu, R.; Auer, S. Amyloid fibrillation kinetics: insight from atomistic nucleation theory. J. Mol. Biol. 2011, 411, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Auer, S.; Meersman, F.; Dobson, C.M.; Vendruscolo, M. A Generic Mechanism of Emergence of Amyloid Protofilaments from Disordered Oligomeric Aggregates. PLoS Comput. Biol. 2008, 4, e1000222. [Google Scholar] [CrossRef] [PubMed]
- Schreck, J.S.; Yuan, J.M. Exactly solvable model for helix-coil-sheet transitions in protein systems. Phys. Rev. E 2010, 81, 061919. [Google Scholar] [CrossRef] [PubMed]
- Schmit, J.D. Kinetic theory of amyloid fibril templating. J. Chem. Phys. 2013, 138, 185102. [Google Scholar] [CrossRef] [PubMed]
- Knowles, T.P.J.; Simone, A.D.; Fitzpatrick, A.W.; Baldwin, A.; Meehan, S.; Rajah, L.; Vendruscolo, M.; Welland, M.E.; Dobson, C.M.; Terentjev, E.M. Twisting transition between crystalline and fibrillar phases of aggregated peptides. Phys. Rev. Lett. 2012, 109, 158101. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.F. Self-assembly of protein amyloids: A competition between amorphous and ordered aggregation. Phys. Rev. E 2009, 80, 031922. [Google Scholar] [CrossRef] [PubMed]
- Bellesia, G.; Shea, J.E. Diversity of kinetic pathways in amyloid fibril formation. J. Chem. Phys. 2009, 131, 111102. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.I.A.; Linse, S.; Luheshi, L.M.; Hellstrand, E.; White, D.A.; Rajah, L.; Otzen, D.E.; Vendruscolo, M.; Dobson, C.M.; Knowles, T.P.J. Proliferation of amyloid-β42 aggregates occurs through a secondary nucleation mechanism. Proc. Natl. Acad. Sci. USA 2013, 110, 9758–9763. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Muthukumar, M.J. Simulations of nucleation and elongation of amyloid fibrils. J. Chem. Phys. 2009, 130, 035102. [Google Scholar] [CrossRef] [PubMed]
- Poland, D.; Scheraga, H. Theory of Helix-Coil Transitions in Biopolymers; Academic Press: New York, NY, USA, 1970. [Google Scholar]
- Dill, K.; Bromberg, S. Molecular Driving Forces; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Smirnovas, V.; Winter, R.; Funk, T.; Dzwolak, W. Thermodynamic properties underlying the α-helix-to-β-sheet transition, aggregation, and amyloidogenesis of polylysine as probed by calorimetry, densimetry, and ultrasound velocimetry. J. Phys. Chem. B 2005, 109, 19043–19045. [Google Scholar] [CrossRef] [PubMed]
- Lifson, S.; Roig, A. On the Theory of Helix—Coil Transition in Polypeptides. J. Chem. Phys. 1961, 34, 1963–1974. [Google Scholar] [CrossRef]
- Van Gestel, J.; de Leeuw, S.W. A statistical mechanical theory of fibril formation in dilute protein solutions. Biophys. J. 2006, 90, 3134–3145. [Google Scholar] [CrossRef] [PubMed]
- Zimm, B.H.; Bragg, J.K. Theory of the Phase Transition between Helix and Random Coil in Polypeptide Chains. J. Chem. Phys. 1959, 31, 526–535. [Google Scholar] [CrossRef]
- Qian, H.; Schellman, J.A. Helix-coil studies: a comparative study for finite length polypeptides. J. Phys. Chem. 1992, 96, 3987–3994. [Google Scholar] [CrossRef]
- Bitan, G.; Kirkitadze, M.D.; Lomakin, A.; Vollers, S.S.; Benedek, G.B.; Teplow, D.B. Amyloid β-protein (Aβ) assembly: Aβ 40 and Aβ 42 oligomerize through distinct pathways. Proc. Natl. Acad. Sci. USA 2003, 100, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Barghorn, S.; Nimmrich, V.; Striebinger, A.; Krantz, C.; Keller, P.; Janson, B.; Bahr, M.; Schmidt, M.; Bitner, R.; Harlan, J.; et al. Globular amyloid β-peptide oligomer—A homogenous and stable neuropathological protein in Alzheimer’s disease. J. Neurochem. 2005, 95, 834–847. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Schmit, J. Pseudo-one-dimensional nucleation in dilute polymer solutions. Phys. Rev. E 2016, 93, 060401(R). [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Schmit, J. Theory of amyloid fibril nucleation from folded proteins. Isr. J. Chem. 2017, 57, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Ferrone, F.A. Assembly of Aβ Proceeds via Monomeric Nuclei. J. Mol. Biol. 2015, 427, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Knowles, T.P.; White, D.A.; Abate, A.R.; Agresti, J.J.; Cohen, S.I.A.; Sperling, R.A.; De Genst, E.J.; Dobson, C.M.; Weitz, D.A. Observation of spatial propagation of amyloid assembly from single nuclei. Proc. Natl. Acad. Sci. USA 2011, 108, 14746–14751. [Google Scholar] [CrossRef] [PubMed]
- Ruschak, A.M.; Miranker, A.D. Fiber-dependent amyloid formation as catalysis of an existing reaction pathway. Proc. Natl. Acad. Sci. USA 2007, 104, 12341–12346. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L. Assembly Mechanism for Aggregation of Amyloid Fibrils. Int. J. Mol. Sci. 2018, 19, 2141. https://doi.org/10.3390/ijms19072141
Zhang L. Assembly Mechanism for Aggregation of Amyloid Fibrils. International Journal of Molecular Sciences. 2018; 19(7):2141. https://doi.org/10.3390/ijms19072141
Chicago/Turabian StyleZhang, Lingyun. 2018. "Assembly Mechanism for Aggregation of Amyloid Fibrils" International Journal of Molecular Sciences 19, no. 7: 2141. https://doi.org/10.3390/ijms19072141
APA StyleZhang, L. (2018). Assembly Mechanism for Aggregation of Amyloid Fibrils. International Journal of Molecular Sciences, 19(7), 2141. https://doi.org/10.3390/ijms19072141