DEAD-Box Protein RNA-Helicase DDX6 Regulates the Expression of HER2 and FGFR2 at the Post-Transcriptional Step in Gastric Cancer Cells

 ,
,  ,
, 
Abstract
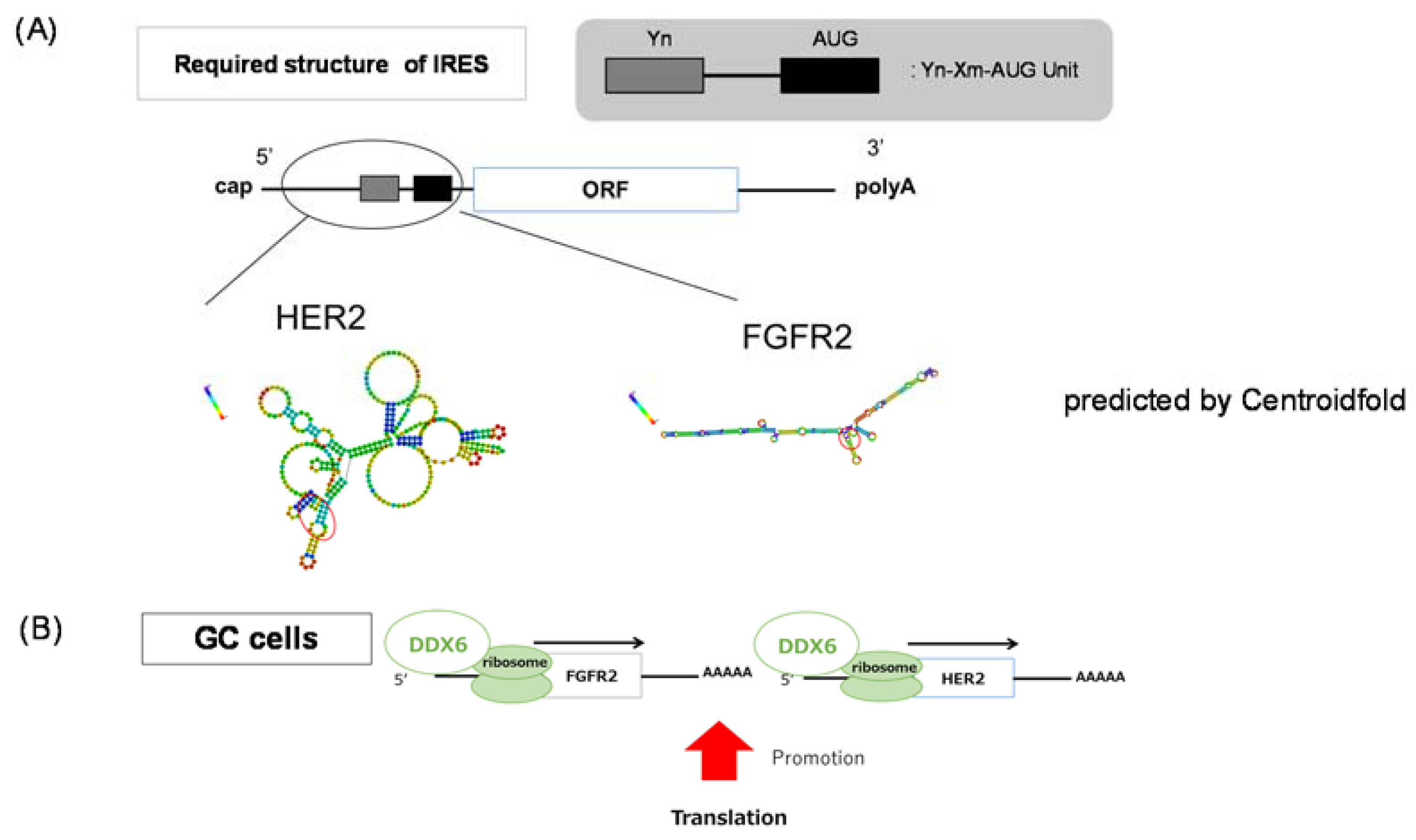
1. Introduction
2. Results
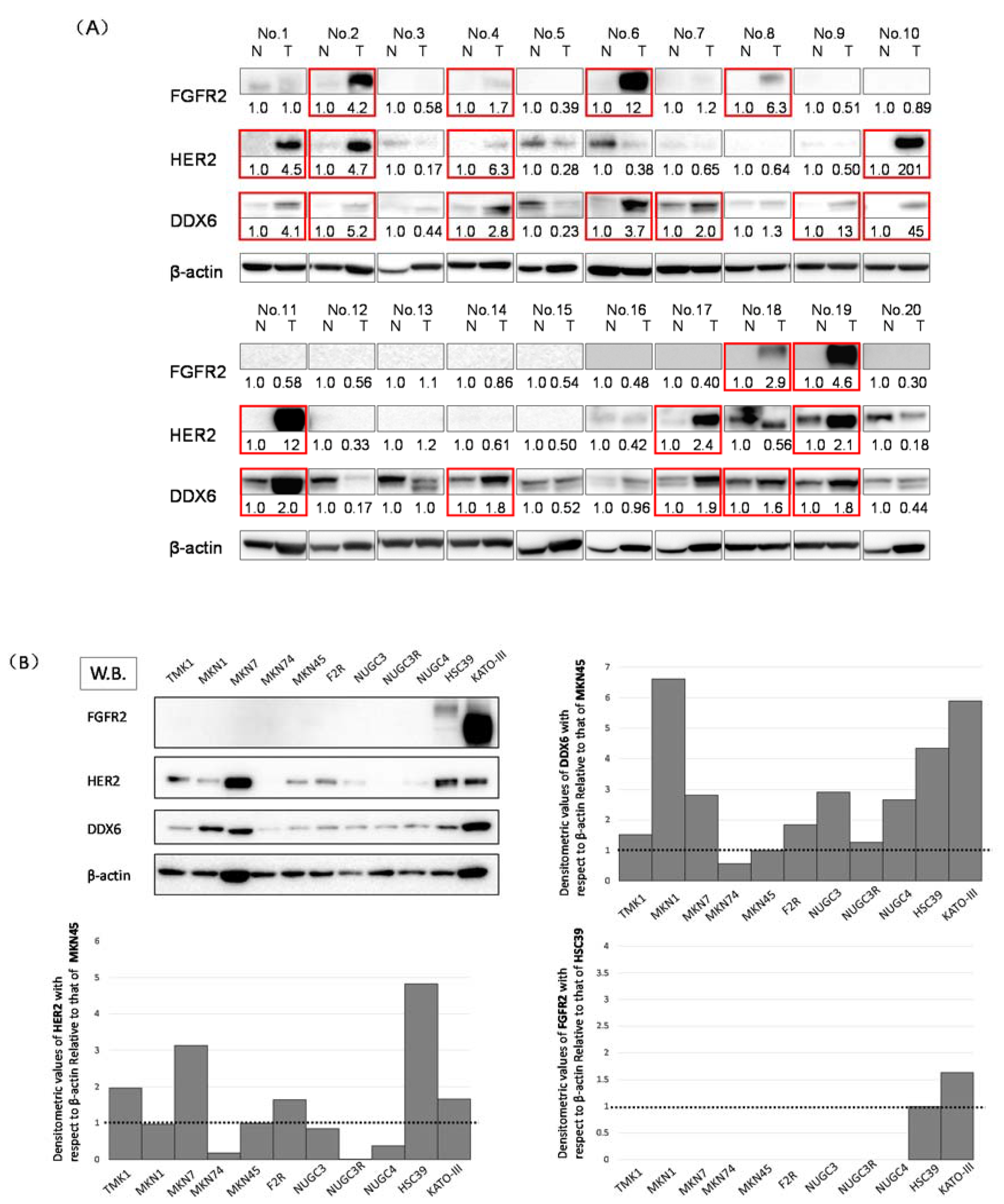
2.1. Expression Levels of DDX6, HER2, and FGFR2 in GC Clinical Tumor Samples
2.2. Expression Levels of DDX6, HER2, and FGFR2 in GC Cell Lines
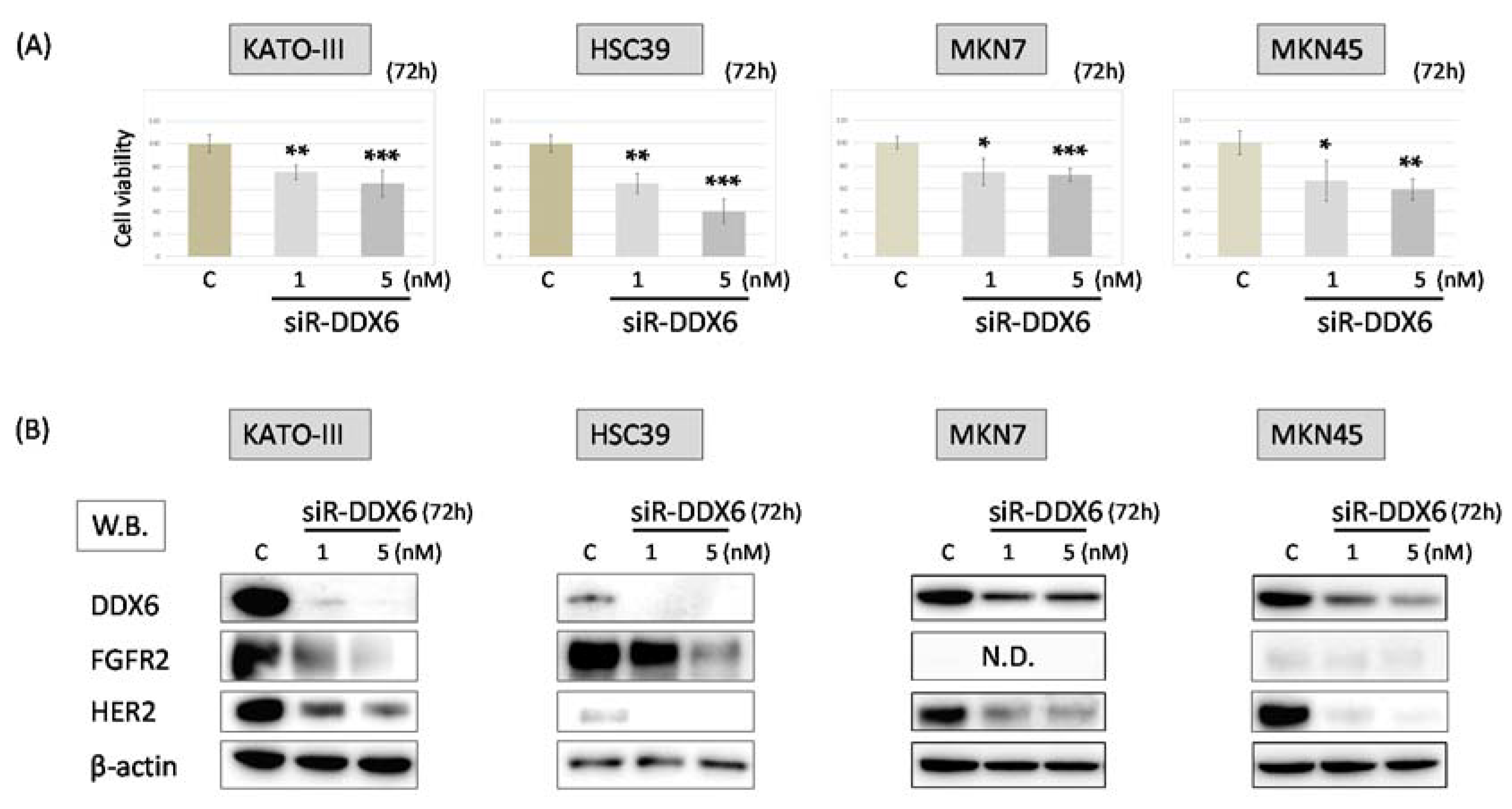
2.3. Effect of Knockdown of DDX6 on Expression of FGFR2 and HER2 in MKN7, MKN45, HSC39, and KATO-III Cells
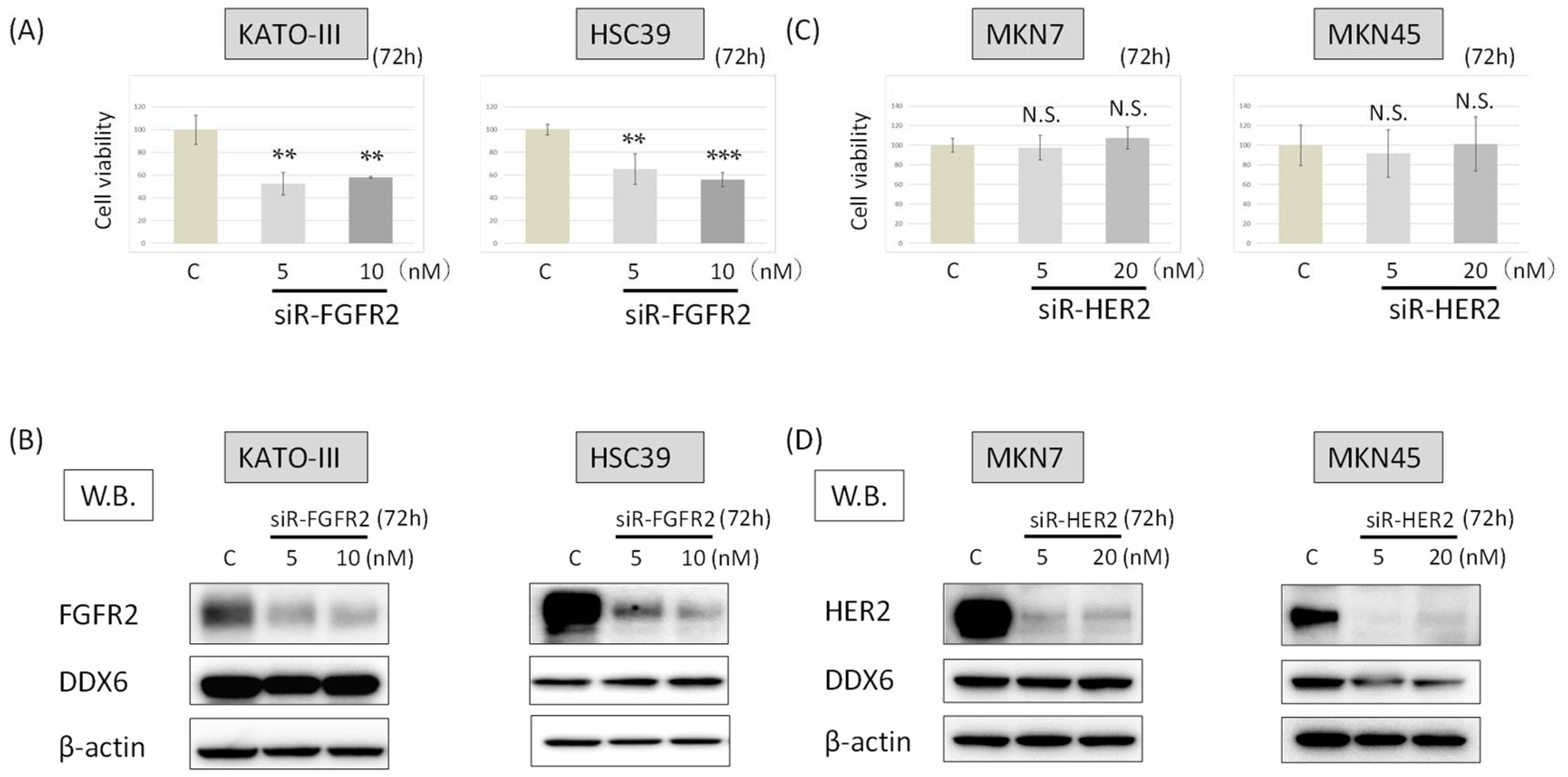
2.4. DDX6 Expression after the Knockdown of FGFR2 in HSC39 and KATO-III Cells
2.5. DDX6 Expression after the Knockdown of HER2 in MKN7 and MKN45 Cells
2.6. HER2 and FGFR2 mRNA Levels after Knockdown of DDX6 in MKN7, MKN45, HSC39, and KATO-III Cells
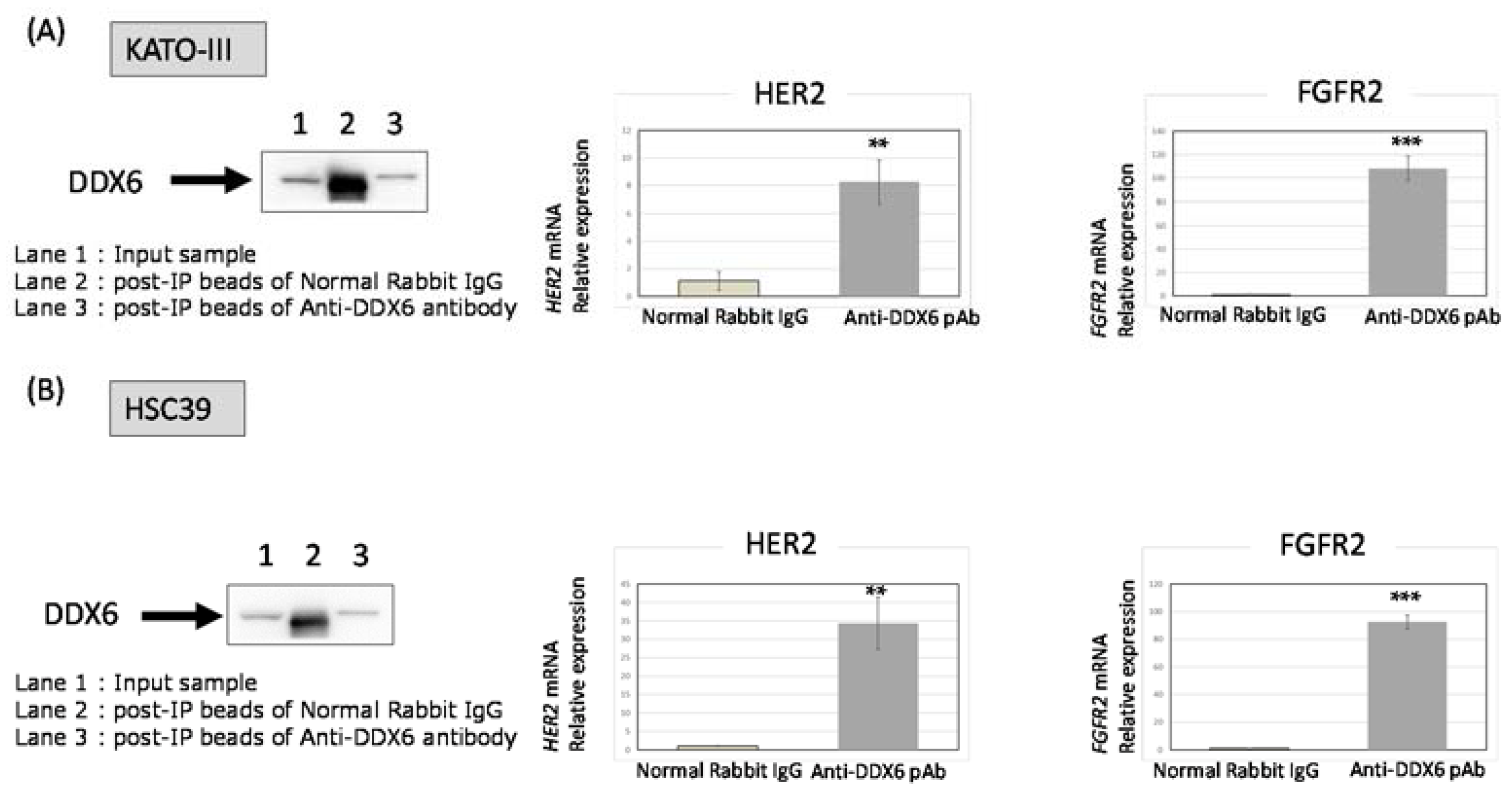
2.7. RNA Immunoprecipitation (RIP)-Assay Using GC Cells
3. Discussion
4. Materials and Methods
4.1. Clinical Samples
4.2. Cell Culture and Cell Viability
4.3. Transfection Experiments
4.4. Western Blot Analysis
4.5. Real-Time Reverse Transcription PCR
4.6. RIP-Assay
4.7. Statistics
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Karim-Kos, H.E.; de Vries, E.; Soerjomataram, I.; Lemmens, V.; Siesling, S.; Coebergh, J.W. Recent trends of cancer in Europe: A combined approach of incidence, survival and mortality for 17 cancer sites since the 1990s. Eur. J. Cancer 2008, 44, 1345–1389. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.K.; Boku, N.; Satoh, T.; Ryu, M.H.; Chao, Y.; Kato, K.; Chung, H.C.; Chen, J.S.; Muro, K.; Kang, W.K.; et al. Nivolumab in patients with advanced gastric or gastro-oesophageal junction cancer refractory to, or intolerant of, at least two previous chemotherapy regimens (ONO-4538-12, ATTRACTION-2): A randomised, double-blind, placebo-controlled, phase 3 trial. Lancet 2017, 390, 2461–2471. [Google Scholar] [CrossRef]
- Japanese Gastric Cancer Association. Japanese gastric cancer treatment guidelines 2014 (ver. 4). Gastric Cancer 2017, 20, 1–19. [Google Scholar]
- Lee, J.; Ou, S.H. Towards the goal of personalized medicine in gastric cancer—Time to move beyond HER2 inhibition. Part I: Targeting receptor tyrosine kinase gene amplification. Discov. Med. 2013, 15, 333–341. [Google Scholar] [PubMed]
- Wang, Y.; Arribas-Layton, M.; Chen, Y.; Lykke-Andersen, J.; Sen, G.L. DDX6 Orchestrates Mammalian Progenitor Function through the mRNA Degradation and Translation Pathways. Mol. Cell 2015, 60, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, Y.; Morikawa, H.; Hirata, I.; Shiozaki, M.; Matsumoto, A.; Maemura, K.; Nishikawa, T.; Niki, M.; Tanigawa, N.; Ikegami, M.; et al. Overexpression of rck/p54, a DEAD box protein, in human colorectal tumours. Br. J. Cancer 1999, 80, 914–917. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Nakagawa, Y.; Morikawa, H.; Niki, M.; Egashira, Y.; Hirata, I.; Katsu, K.; Akao, Y. Co-overexpression of DEAD box protein rck/p54 and c-myc protein in human colorectal adenomas and the relevance of their expression in cultured cell lines. Carcinogenesis 2001, 22, 1965–1970. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, K.; Iwatsuki, A.; Sugito, N.; Shinohara, H.; Kuranaga, Y.; Oshikawa, Y.; Tajirika, T.; Futamura, M.; Yoshida, K.; Uchiyama, K.; et al. Oncogene RNA helicase DDX6 promotes the process of c-Myc expression in gastric cancer cells. Mol. Carcinog. 2018, 57, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Akao, Y.; Matsumoto, K.; Ohguchi, K.; Nakagawa, Y.; Yoshida, H. Human DEAD-box/RNA unwindase rck/p54 contributes to maintenance of cell growth by affecting cell cycle in cultured cells. Int. J. Oncol. 2006, 29, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Duraes, C.; Almeida, G.M.; Seruca, R.; Oliveira, C.; Carneiro, F. Biomarkers for gastric cancer: Prognostic, predictive or targets of therapy? Virchows Arch. 2014, 464, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Normanno, N.; Bianco, C.; Strizzi, L.; Mancino, M.; Maiello, M.R.; De Luca, A.; Caponigro, F.; Salomon, D.S. The ErbB receptors and their ligands in cancer: an overview. Curr. Drug Targets 2005, 6, 243–257. [Google Scholar] [CrossRef] [PubMed]
- Gravalos, C.; Jimeno, A. HER2 in gastric cancer: A new prognostic factor and a novel therapeutic target. Ann. Oncol. 2008, 19, 1523–1529. [Google Scholar] [CrossRef] [PubMed]
- Gallardo, A.; Lerma, E.; Escuin, D.; Tibau, A.; Munoz, J.; Ojeda, B.; Barnadas, A.; Adrover, E.; Sanchez-Tejada, L.; Giner, D.; et al. Increased signalling of EGFR and IGF1R, and deregulation of PTEN/PI3K/Akt pathway are related with trastuzumab resistance in HER2 breast carcinomas. Br. J. Cancer 2012, 106, 1367–1373. [Google Scholar] [CrossRef] [PubMed]
- Dragovich, T.; McCoy, S.; Fenoglio-Preiser, C.M.; Wang, J.; Benedetti, J.K.; Baker, A.F.; Hackett, C.B.; Urba, S.G.; Zaner, K.S.; Blanke, C.D.; et al. Phase II trial of erlotinib in gastroesophageal junction and gastric adenocarcinomas: SWOG 0127. J. Clin. Oncol. 2006, 24, 4922–4927. [Google Scholar] [CrossRef] [PubMed]
- Okines, A.F.; Cunningham, D. Trastuzumab in gastric cancer. Eur. J. Cancer 2010, 46, 1949–1959. [Google Scholar] [CrossRef] [PubMed]
- Phillips, B.E.; Tubbs, R.R.; Rice, T.W.; Rybicki, L.A.; Plesec, T.; Rodriguez, C.P.; Videtic, G.M.; Saxton, J.P.; Ives, D.I.; Adelstein, D.J. Clinicopathologic features and treatment outcomes of patients with human epidermal growth factor receptor 2-positive adenocarcinoma of the esophagus and gastroesophageal junction. Dis. Esophagus 2013, 26, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Chan, D.S.; Campbell, F.; Edwards, P.; Jasani, B.; Williams, G.T.; Lewis, W.G. Relative Prognostic Value of Human Epidermal Growth Factor Receptor 2 (HER2) Expression in Operable Oesophagogastric Cancer. ISRN Surg. 2012, 2012, 804891. [Google Scholar] [CrossRef] [PubMed]
- Okines, A.F.; Thompson, L.C.; Cunningham, D.; Wotherspoon, A.; Reis-Filho, J.S.; Langley, R.E.; Waddell, T.S.; Noor, D.; Eltahir, Z.; Wong, R.; et al. Effect of HER2 on prognosis and benefit from peri-operative chemotherapy in early oesophago-gastric adenocarcinoma in the MAGIC trial. Ann. Oncol. 2013, 24, 1253–1261. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.A.; Jung, E.J.; Lee, H.S.; Lee, H.E.; Jeon, Y.K.; Yang, H.K.; Kim, W.H. Evaluation of HER-2 gene status in gastric carcinoma using immunohistochemistry, fluorescence in situ hybridization, and real-time quantitative polymerase chain reaction. Hum. Pathol. 2007, 38, 1386–1393. [Google Scholar] [CrossRef] [PubMed]
- Bilous, M.; Osamura, R.Y.; Ruschoff, J.; van de Vijver, M.; Hanna, W.; Penault-Llorca, F.; Roche, P. HER-2 amplification is highly homogenous in gastric cancer. Hum. Pathol. 2010, 41, 304–305. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.Y.; Hu, Y.; Fan, J.G.; Tao, G.Q.; Lu, Y.M.; Cai, X.; Yu, B.H.; Du, Y.Q. Clinicopathologic significance of HER-2/neu protein expression and gene amplification in gastric carcinoma. World J. Gastroenterol. 2011, 17, 1501–1506. [Google Scholar] [CrossRef] [PubMed]
- Moelans, C.B.; Milne, A.N.; Morsink, F.H.; Offerhaus, G.J.; van Diest, P.J. Low frequency of HER2 amplification and overexpression in early onset gastric cancer. Cell. Oncol. 2011, 34, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Li, N.; Jiang, W.; Hua, Z.; Xia, L.; Wei, Q.; Wang, L. Prognosis significance of HER-2/neu overexpression/amplification in Chinese patients with curatively resected gastric cancer after the ToGA clinical trial. World J. Surg. Oncol. 2012, 10, 274. [Google Scholar] [CrossRef] [PubMed]
- Tanner, M.; Hollmen, M.; Junttila, T.T.; Kapanen, A.I.; Tommola, S.; Soini, Y.; Helin, H.; Salo, J.; Joensuu, H.; Sihvo, E.; et al. Amplification of HER-2 in gastric carcinoma: Association with Topoisomerase IIalpha gene amplification, intestinal type, poor prognosis and sensitivity to trastuzumab. Ann. Oncol. 2005, 16, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Park, D.I.; Yun, J.W.; Park, J.H.; Oh, S.J.; Kim, H.J.; Cho, Y.K.; Sohn, C.I.; Jeon, W.K.; Kim, B.I.; Yoo, C.H.; et al. HER-2/neu amplification is an independent prognostic factor in gastric cancer. Dig. Dis. Sci. 2006, 51, 1371–1379. [Google Scholar] [CrossRef] [PubMed]
- Bang, Y.J.; Van Cutsem, E.; Feyereislova, A.; Chung, H.C.; Shen, L.; Sawaki, A.; Lordick, F.; Ohtsu, A.; Omuro, Y.; Satoh, T. Trastuzumab in combination with chemotherapy versus chemotherapy alone for treatment of HER2-positive advanced gastric or gastro-oesophageal junction cancer (ToGA): A phase 3, open-label, randomised controlled trial. Lancet 2010, 376, 687–697. [Google Scholar] [CrossRef]
- Katoh, Y.; Katoh, M. FGFR2-related pathogenesis and FGFR2-targeted therapeutics (Review). Int. J. Mol. Med. 2009, 23, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Itoh, H.; Uchino, S.; Hosokawa, K.; Ochiai, A.; Ino, Y.; Ishii, H.; Sakamoto, H.; Yamaguchi, N.; Yanagihara, K.; et al. Immunohistochemical detection of K-sam protein in stomach cancer. Clin. Cancer Res. 1996, 2, 1373–1381. [Google Scholar] [PubMed]
- Toyokawa, T.; Yashiro, M.; Hirakawa, K. Co-expression of keratinocyte growth factor and K-sam is an independent prognostic factor in gastric carcinoma. Oncol. Rep. 2009, 21, 875–880. [Google Scholar] [PubMed]
- Matsui, T.; Hogetsu, K.; Usukura, J.; Sato, T.; Kumasaka, T.; Akao, Y.; Tanaka, N. Structural insight of human DEAD-box protein rck/p54 into its substrate recognition with conformational changes. Genes Cells Devoted Mol. Cell. Mech. 2006, 11, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Yanagihara, K.; Seyama, T.; Tsumuraya, M.; Kamada, N.; Yokoro, K. Establishment and characterization of human signet ring cell gastric carcinoma cell lines with amplification of the c-myc oncogene. Cancer Res. 1991, 51, 381–386. [Google Scholar] [PubMed]
- Tsutani, Y.; Yoshida, K.; Sanada, Y.; Wada, Y.; Konishi, K.; Fukushima, M.; Okada, M. Decreased orotate phosphoribosyltransferase activity produces 5-fluorouracil resistance in a human gastric cancer cell line. Oncol. Rep. 2008, 20, 1545–1551. [Google Scholar] [PubMed]
- Mori, R.; Yoshida, K.; Tanahashi, T.; Yawata, K.; Kato, J.; Okumura, N.; Tsutani, Y.; Okada, M.; Oue, N.; Yasui, W. Decreased FANCJ caused by 5FU contributes to the increased sensitivity to oxaliplatin in gastric cancer cells. Gastric Cancer 2013, 16, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Akao, Y.; Nakagawa, Y.; Hirata, I.; Iio, A.; Itoh, T.; Kojima, K.; Nakashima, R.; Kitade, Y.; Naoe, T. Role of anti-oncomirs miR-143 and -145 in human colorectal tumors. Cancer Gene Ther. 2010, 17, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, S.; Iwasaki, J.; Kumazaki, M.; Mori, T.; Maruo, K.; Sakai, H.; Yamada, N.; Shimada, K.; Naoe, T.; Kitade, Y.; et al. Chemically modified synthetic microRNA-205 inhibits the growth of melanoma cells in vitro and in vivo. Mol. Ther. J. Am. Soc. Gene Ther. 2013, 21, 1204–1211. [Google Scholar] [CrossRef] [PubMed]
- Yamada, N.; Tsujimura, N.; Kumazaki, M.; Shinohara, H.; Taniguchi, K.; Nakagawa, Y.; Naoe, T.; Akao, Y. Colorectal cancer cell-derived microvesicles containing microRNA-1246 promote angiogenesis by activating Smad 1/5/8 signaling elicited by PML down-regulation in endothelial cells. Biochim. Biophys. Acta 2014, 1839, 1256–1272. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Age | Sex | Site | Form | Pathological Findings | UICC/TNM | T | N | M | Stage |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 71 | M | L | Type3 | tub2 | ⃞ | T3 | N0 | M0 | IIA |
| 2 | 91 | F | L | Type2 | tub2 | ⃞ | T4b | N2 | M0 | IIIC |
| 3 | 72 | M | L | Type3 | tub1 | ⃞ | T4a | N3a | M0 | IIIC |
| 4 | 61 | M | L | Type2 | tub2 | ⃞ | T4a | N3a | M0 | IIIC |
| 5 | 83 | M | L | Type2 | por1 | ⃞ | T3 | N0 | M0 | IIA |
| 6 | 77 | M | UM | Type3 | muc > tub2 | ⃞ | T3 | N1 | M0 | IIB |
| 7 | 66 | F | LM | Type1 | tub2 | ⃞ | T2 | N2 | M0 | IIB |
| 8 | 84 | M | MLD | Type3 | tub2 | ⃞ | T4a | N3a | M0 | IIIC |
| 9 | 85 | F | M | Type2 | por2 > tub1 | ⃞ | T3 | N0 | M0 | IIA |
| 10 | 72 | M | ML | Type1 | tub1 > pap | ⃞ | T1b | N0 | M0 | IA |
| 11 | 64 | M | U | Type1 | tub2 | ⃞ | T3 | N0 | M0 | IIA |
| 12 | 78 | M | LM | Type2 | por1 | ⃞ | T4b | N3a | M0 | IIIC |
| 13 | 64 | F | GE | Type2 | pap > muc | ⃞ | T3 | N0 | M0 | IIA |
| 14 | 73 | M | M | Type1 | tub1 (>tub2) | ⃞ | T1a | N0 | M0 | IA |
| 15 | 85 | F | M | Type3 | por1 > tub1 | ⃞ | T2 | N2 | M0 | IIB |
| 16 | 73 | M | LMU | Type4 | por2 > muc | ⃞ | T4a | N3a | M0 | IIIC |
| 17 | 80 | M | UE | Type2 | tub2 | ⃞ | T3 | N0 | M0 | IIA |
| 18 | 93 | M | ML | Type3 | tub2 | ⃞ | T4a | N0 | M0 | IIB |
| 19 | 65 | F | L | Type2 | tub2 > tub1 | ⃞ | T2 | N0 | M0 | IB |
| 20 | 74 | F | U | Type3 | por2 > sig > muc | ⃞ | T4a | N3a | M0 | IIIC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tajirika, T.; Tokumaru, Y.; Taniguchi, K.; Sugito, N.; Matsuhashi, N.; Futamura, M.; Yanagihara, K.; Akao, Y.; Yoshida, K. DEAD-Box Protein RNA-Helicase DDX6 Regulates the Expression of HER2 and FGFR2 at the Post-Transcriptional Step in Gastric Cancer Cells. Int. J. Mol. Sci. 2018, 19, 2005. https://doi.org/10.3390/ijms19072005
Tajirika T, Tokumaru Y, Taniguchi K, Sugito N, Matsuhashi N, Futamura M, Yanagihara K, Akao Y, Yoshida K. DEAD-Box Protein RNA-Helicase DDX6 Regulates the Expression of HER2 and FGFR2 at the Post-Transcriptional Step in Gastric Cancer Cells. International Journal of Molecular Sciences. 2018; 19(7):2005. https://doi.org/10.3390/ijms19072005
Chicago/Turabian StyleTajirika, Toshihiro, Yoshihisa Tokumaru, Kohei Taniguchi, Nobuhiko Sugito, Nobuhisa Matsuhashi, Manabu Futamura, Kazuyoshi Yanagihara, Yukihiro Akao, and Kazuhiro Yoshida. 2018. "DEAD-Box Protein RNA-Helicase DDX6 Regulates the Expression of HER2 and FGFR2 at the Post-Transcriptional Step in Gastric Cancer Cells" International Journal of Molecular Sciences 19, no. 7: 2005. https://doi.org/10.3390/ijms19072005
APA StyleTajirika, T., Tokumaru, Y., Taniguchi, K., Sugito, N., Matsuhashi, N., Futamura, M., Yanagihara, K., Akao, Y., & Yoshida, K. (2018). DEAD-Box Protein RNA-Helicase DDX6 Regulates the Expression of HER2 and FGFR2 at the Post-Transcriptional Step in Gastric Cancer Cells. International Journal of Molecular Sciences, 19(7), 2005. https://doi.org/10.3390/ijms19072005