Abstract
In this study, we evaluated the acute (24, 48, 72, and 96 h) and chronic (21 days) adverse effects induced by low doses (0.1, 0.5, 1, and 1.5 mg/L) of zinc chloride (ZnCl2) exposure in adult zebrafish by using behavioral endpoints like three-dimensional (3D) locomotion, passive avoidance, aggression, circadian rhythm, and predator avoidance tests. Also, brain tissues were dissected and subjected to analysis of multiple parameters related to oxidative stress, antioxidant responses, superoxide dismutase (SOD), neurotoxicity, and neurotransmitters. The results showed that ZnCl2-exposed fishes displayed decreased locomotor behavior and impaired short-term memory, which caused an Alzheimer’s Disease (AD)-like syndrome. In addition, low concentrations of ZnCl2 induced amyloid beta (amyloid β) and phosphorylated Tau (p-Tau) protein levels in brains. In addition, significant induction in oxidative stress indices (reactive oxygen species (ROS) and malondialdehyde (MDA)), reduction in antioxidant defense system (glutathione (GSH), GSH peroxidase (GSH-Px) and SOD) and changes in neurotransmitters were observed at low concentrations of ZnCl2. Neurotoxic effects of ZnCl2 were observed with significant inhibition of acetylcholine (ACh) activity when the exposure dose was higher than 1 ppm. Furthermore, we found that zinc, metallothionein (MT), and cortisol levels in brain were elevated compared to the control group. A significantly negative correlation was observed between memory and acetylcholinesterase (AChE) activity. In summary, these findings revealed that exposure to ZnCl2 affected the behavior profile of zebrafish, and induced neurotoxicity which may be associated with damaged brain areas related to memory. Moreover, our ZnCl2-induced zebrafish model may have potential for AD-associated research in the future.
1. Introduction
Currently, there are more than 50 million people living with Alzheimer’s Disease (AD) in the world; this is greater than the total population of Spain and is projected to reach nearly 150 million by 2050 [1]. Loss of memory is the first symptom reported by patients suffering from AD. It was hypothesized that the pathogenesis of AD is diverse, including free-radical-mediated processes [2], dysregulated membrane metabolism [3], trace element neurotoxicity [4], or combinations of the abovementioned factors. Another study suggested that the pyruvate transporter on the inner mitochondrial membrane impacts oxidative stress and lipid metabolism, playing a significant role in the pathogenesis of AD [5]. Among these factors, one which gained significant attention is the involvement of trace elements that caused toxicity during AD progression. Previous studies showed that imbalances of trace elements in homeostasis, such as bulk copper [6], bulk iron [7], and zinc [8,9,10], played an important role in the formation of senile plaques in the brain.
Zinc is an essential trace element, required for the functional integrity of many organ systems, as well as for development, growth, and tissue repair [11,12]. The divalent ion forms of zinc are nutritionally essential trace elements for all living organisms, and is also a co-factor for a number of enzymes [13]. However, excessive zinc is lethal and harmful in pathological and toxicological aspects. Zinc was found as an environmental hazard in at least 985 of the 1662 waste sites proposed by the Environmental Protection Agency (EPA) National Priorities list [14]. The toxicity by zinc was reported in several fish species [15,16,17]. It was well documented that zinc induced embryological damage in red sea bream Pagrus major, including alterations in fecundity, low hatching rate, and high mortality [18]. Another study revealed that zinc caused toxicity in gills, which altered oxygen consumption, ammonia excretion, osmotic pressure, and the hepatopancreas in several aquatic invertebrates [19].
It was reported that zinc is neurotoxic and causes neurodegeneration in the central nervous system (CNS) [20,21]. Increases in cytosolic Zn2+ concentration via transport through Ca2+ channels triggered several downstream effects which led to neuronal cell death [22]. Despite the very low levels of free intracellular Zn2+, exposure of 400–600 nM Zn2+ results in increased toxicity [23]. Moreover, cells with over 400 nM of Zn2+ decreased the activity of glycolytic enzymes, such as glyceraldehyde 3-phosphate dehydrogenase (GADPH) and phosphofructokinase [24,25]. Studies also reported that intracellular free Zn2+ induced intracellular proton concentration that affected the function of neurons in vitro [26]. Interestingly, a number of studies revealed the significant role of heavy metals in several neurodegenerative diseases, including Alzheimer’s disease, vascular dementia, and Parkinson’s disease [21,27,28].
Reactive oxygen species (ROSs) play an important role in physiological function and are used as a biomarker of oxidative stress in toxicity studies [29,30]. Some studies already showed that the antioxidant defense system and cellular responses to oxidative stress in zebrafish were similar to mammals [31]. In the CNS, the neurotransmitter acetylcholine (ACh), involved in the cholinergic signaling system, is associated with various physiological and behavioral processes, such as motor activity, memory, and cognition, by activating metabotropic muscarinic and ionotropic nicotinic cholinergic receptors [32]. ACh is catalyzed by two cholinesterases: acetylcholinesterase (AChE) and butyrylcholinesterse (BuChE). Zebrafish encodes only the gene for AChE, which is responsible for ACh degradation, and was already identified and functionally detected in the zebrafish brain [33]. Therefore, acetylcholinesterase (AChE) plays an active role in the nervous system [34]. Moreover, AChE is a significant biomarker for detecting environmental contaminants deposited in zebrafish [35,36]. AChE is involved in the termination of impulse exchange through rapid hydrolysis of the neurotransmitter ACh, resulting in enzyme inactivation induced by various inhibitors, and leading to disrupted neurotransmission [37]. As toxic chemicals affect non-target organisms in the aquatic biota, fish are important as bio indicators to monitor the changes in environmental toxicity compared to mammalian models.
The United States Environmental Protection Agency (USEPA) established a system as an initial screen that applied zebrafish as a model for rapid identification of chemical toxicity to evaluate the effects from myriad chemical pollutants [38]. However, the biological effects of ZnCl2 on neurobehavioral development in a vertebrate model are not well characterized in research (Table A1). Moreover, some reports found that genotoxic effects and gene expression changes in embryogenesis following ZnCl2 exposure were still largely unknown. One recent study demonstrated the toxic effects of zinc in AD in a rat model, proven using histological results [20]. In addition, it was demonstrated developmental effects of ZnCl2 during the larval stage of zebrafish [39,40]. Therefore, we sought to clarify both the short-term and long-term effects of ZnCl2 on neurotoxicity by investigating the neurological behaviors immediately after exposure and later on by assessing the molecular parameters in adult zebrafish. In addition, there is currently no study on the effect of ZnCl2 exposure on memory changes determined using a passive avoidance memory test, and its possible mechanisms in adult zebrafish.
The objective of this study was to determine if exposure to environmentally relevant ZnCl2 in adult zebrafish could result in long-term impairment of brain and muscle functions, leading to neurological defects. Therefore, we used different behavioral endpoints to analyze the toxic effects of ZnCl2, and assessed the cholinergic system by analyzing ACh and AChE activity in the zebrafish brain. Furthermore, our study was designed to assess parameters related to oxidative stress and the amino-acid neurotransmitter system after long-term exposure to ZnCl2. Considering that long-term ZnCl2 exposure promotes neurological dysfunction, and the cholinergic system, oxidative imbalance, and antioxidant activity are associated with neurological disorders related to human diseases, different biomarkers including oxidative and biochemical (ROS and malondialdehyde (MDA)), antioxidant parameters (glutathione (GSH), GSH peroxidase (GSH-PX), and superoxide dismutase (SOD)), neurotoxicity (ACh, AChE, amyloid β, and phosphorylated (p)-Tau), neurotransmitters (gamma-aminobutyric acid (GABA), dopamine, and melatonin), and metallothionein protein levels were studied for a better understanding of ZnCl2-induced toxicity in the zebrafish model.
2. Results
2.1. Mortality Curve and Median Lethal Concentration for ZnCl2 Exposure
The acute toxicity test was performed at different concentrations (2, 4, 6, 8, 10, 12, and 15 mg/L) of zinc chloride from low to high concentrations after 24, 48, 72, and 96 h of exposure (Figure 1). No deaths occurred in the untreated control group throughout the exposure experiments. Adult fish incubated at ZnCl2 concentrations of 2 mg/L or lower showed mortality similar to the control group, while fish treated with 15 mg/L had 100% mortality after 24 h of incubation. Median lethal concentration (LC50) was calculated from the percentage of fish mortality for each ZnCl2 concentration with different exposure times (Figure A1). The 96-h LC50 of ZnCl2 was estimated around 6 ppm.
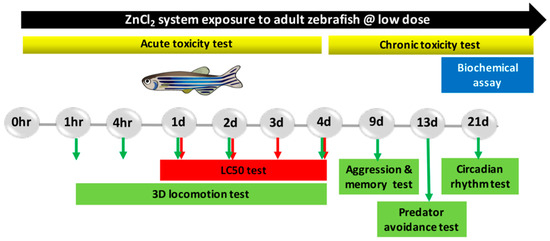
Figure 1.
Experimental design scheme of adult zebrafish exposed to ZnCl2.
2.2. ZnCl2 Exposure Impaired Zebrafish Three-Dimensional Locomotor Activity
The three-dimensional (3D) locomotion behavior of adult fish is a sensitive indicator for testing whether fish are suffering from external stimuli or pollution [39]. We analyzed fish 3D locomotion for five different time periods (1, 4, 24, 48, and 96 h) to demonstrate the acute changes caused by ZnCl2 exposure. We observed that exposure to 0.5, 1.0, and 1.5 ppm ZnCl2 resulted in a decrease in average speed (Figure 2A, p < 0.01), as well as increases in average angular velocity (Figure 2B, p < 0.01) and in meandering (Figure 2C, p < 0.001). Furthermore, treatment with 1.0 ppm ZnCl2 for 24 h increased the time for fast latency with animals showing anxiogenic effects (Figure 2D, p < 0.01). The total distances traveled by adult zebrafish treated with ZnCl2 were reduced (Figure 2E, p < 0.01), and the time spent in the upper zone of the tank by the animals decreased (Figure 2F, p < 0.0001) at all exposure concentrations. Generally, when zebrafish are exposed to a new environment, they tend to spend more time at the tank bottom and gradually move to the upper zone after a few minutes. The time spent in the upper zone indicated an anxiolytic-like behavior [40]. Therefore, our results showed that acute ZnCl2 exposure induced anxiety-like behavior.
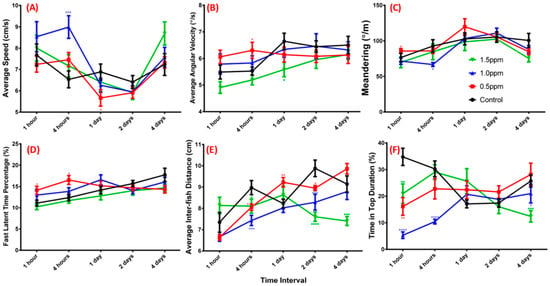
Figure 2.
Three-dimensional (3D) locomotor activity assay for ZnCl2-treated zebrafish. (A) Average speed; (B) average angular velocity; (C) meandering; (D) fast latent time; (E) total distance traveled by treated and control groups; (F) time spent at top of the tank by adult fish within five different time intervals (n = 18 for each testing concentration). Black line, control; red line, 0.5 ppm ZnCl2 exposure; blue line, 1.0 ppm ZnCl2 exposure; green line, 1.5 ppm ZnCl2 exposure. Locomotor activity was compared to control at each time point using a t-test (* p ≤ 0.05; ** p ≤ 0.01; *** p ≤ 0.001; **** p ≤ 0.0001).
2.3. ZnCl2 Exposure Reduced Zebrafish Aggression
We tested fish aggression using the mirror-biting test. Under normal conditions, zebrafish are very aggressive, biting their mirror image [41]. In the untreated control group, we found that adult zebrafish displayed a very high percentage of time visiting/interacting with their mirror images (up to 40%, Figure 3A,B). However, ZnCl2 exposure significantly influenced aggressive behavior as determined by the frequency of entries and time spent in the segment near the mirror images (Figure 3B–E). Quantitative data showed that ZnCl2-treated animals displayed dose-dependent reductions in aggressive behavior compared to control fish (Figure 3A), and spent more time in the tank bottom as ZnCl2 exposure concentration increased (Figure 3D,E; Video S1).
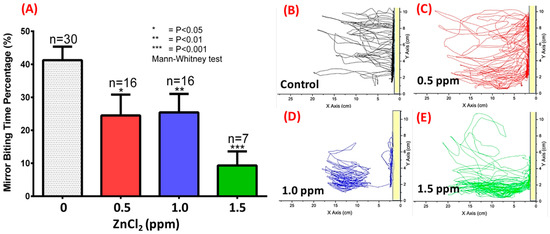
Figure 3.
Aggression test monitored by mirror-biting of ZnCl2-treated adult zebrafish after nine-day exposure. (A) Quantitative comparison of mirror-biting time percentage for zebrafish upon exposure to different concentrations of ZnCl2. Statistical analysis was performed using a non-parametric Mann–Whitney test, and compared to control (* p ≤ 0.05; ** p ≤ 0.01; *** p ≤ 0.001). Locomotion trajectories for zebrafish upon exposure to (B) 0 ppm (control), (C) 0.5 ppm, (D) 1.0 ppm, and (E) 1.5 ppm ZnCl2. The yellow bar indicates the position of the mirror.
2.4. ZnCl2 Exposure Reduced Memory in the Passive Avoidance Test
The adverse effects of zinc on memory were analyzed using the passive avoidance memory test. In the beginning, we acclimated adult zebrafish in the bright chamber after subjecting them to an experimental tank with or without ZnCl2. When the separator between the light and dark sides was removed, the untreated fish immediately swam into the dark chamber. Also, we used mild electrical shocks to punish zebrafish to build up the passive memorial connection between the dark side and pain. After being electrically shocked in the dark chamber, zebrafish could remember the pain for several days and displayed longer latency when accessing the dark chamber. We observed that short-term memory was sensitive to ZnCl2 exposure, and the reduction in memory retention was dose-dependent. The fish displayed significantly lower memory retention when exposed to very low doses of ZnCl2 (0.1 ppm) (Figure 4).
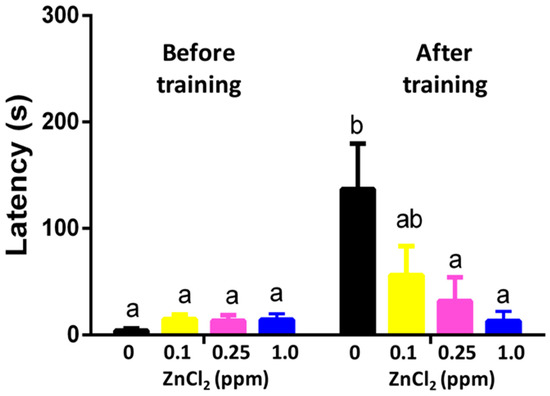
Figure 4.
Passive avoidance on training and memorial test after 96 h of exposure to ZnCl2 after nine-day exposure. Data are presented as means ± standard error of the mean (SEM) (n = 12) and analyzed individually for each treated group. Statistics analysis was performed by one-way ANOVA, followed by Tukey as the post hoc test. Different labels above columns indicate a significant difference with p < 0.01 (a, p ≤ 0.05; ab, p ≤ 0.01; b, p ≤ 0.001)).
2.5. ZnCl2 Exposure Does Not Change Predator Avoidance Behavior
Predator avoidance is an innate behavioral response; here, we used the convict cichlid (Amatitlania nigrofasciata) as a predator to induce great fear/anxiety. There was no significant change in average speed (Figure 5A), predator approaching time (Figure 5B), average distance to separator (Figure 5C), or freezing/swimming time movement ratio (Figure 5D,E) between control and ZnCl2-treated fishes. In contrast, a slightly decrease in in the rapid movement time ratio was observed in the 1 ppm ZnCl2-exposed group (Figure 5F, Video S2). This result implied that ZnCl2 exposure at our tested concentrations did not change predator avoidance behavior in zebrafish.
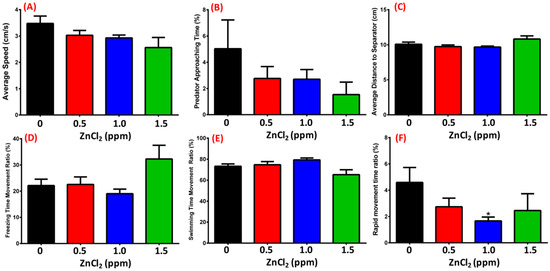
Figure 5.
Predator avoidance test after 13-day exposure. (A) Average speed; (B) predator approaching time; (C) average distance of zebrafish to separator; (D) freezing movement time ratio; (E) swimming time movement ratio; (F) rapid movement time ratio of zebrafish. Statistical analysis was performed using a non-parametric Mann–Whitney test (* p ≤ 0.05). Sample sizes were 32, 14, 16, and 7 fish for the control, 0.5 ppm, 1.0 ppm, and 1.5 ppm ZnCl2-exposed groups, respectively.
2.6. ZnCl2 Exposure Deregulated the Circadian Rhythm
The light/dark circadian test theoretically depends on the preference of staying in a dark environment, which was validated in behavioral assessment [42,43]. When tracking zebrafish locomotion every hour, we found that untreated fish displayed active locomotion (locomotion speed between 1 and 10 cm/s) during the light cycle and sleeping-like behavior during the dark cycle (locomotion speed less than 1 cm/s) (Figure 6A, black line). However, after 1.5 ppm ZnCl2 exposure, zebrafish displayed a reversed circadian rhythm pattern, with less locomotion activity during the light cycle, and higher activity during the dark cycle (Figure 6A, green line). We statistically summarized all locomotion patterns in the light and dark cycles with analysis of average speed during the light cycle (Figure 6C) or dark cycle (Figure 6D). Average angular velocity during the light cycle demonstrated a severe circadian deregulated effect when zebrafish were treated with 1.5 ppm ZnCl2 (Figure 6D). Quantitation of the difference in activity revealed that, while control fish decreased their locomotor activity, the ZnCl2-treated fish maintained almost a constant level of activity throughout the night.
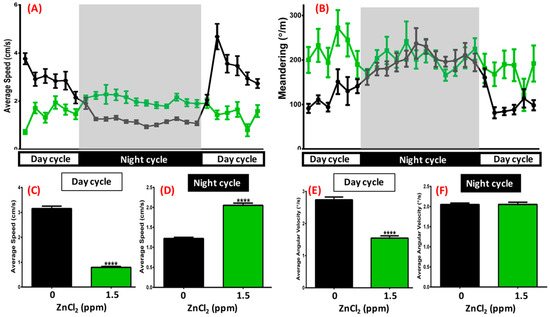
Figure 6.
Circadian rhythm test after 21-day exposure. (A) Average speed and (B) meandering of 1.5 ppm ZnCl2-treated (green line) and control fish (black line) during day/night cycle. The gray area shows the dark period and the unshaded area shows the light period. Eighteen fish were observed for both the control and ZnCl2-treated groups. Quantitative comparison of average speed during (C) day cycle and (D) night cycle. Quantitative comparison of average angular velocity during (E) day cycle and (F) night cycle. Data are presented as means ± SEM (**** p ≤ 0.0001).
2.7. ZnCl2 Exposure Increased Oxidative Stress in the Brain
Upon performing a behavioral test, we found that low doses of ZnCl2 exposure induced multiple behavioral abnormalities. Next, we attempted to elucidate possible mechanisms by performing biochemical assays. No deaths were found in the treatment group and control groups after acute exposure to ZnCl2 at low doses. However, there was a significant increase (more than 30-fold compared to the untreated control) of exogenous zinc ions in the brain after exposure to ZnCl2 higher than 1 ppm (Figure 7A). Compared to other tissues, zinc accumulation in the brain showed the highest level (Figure A2). By performing an ELISA assay, we found that the relative amounts of metallothionein (MT, a heavy-metal chelating protein) in the brain and gill tissues also increased significantly (p < 0.001) at ZnCl2 exposure higher than 1 ppm. In contrast, the MT expression level in the liver tissue was low compared to muscle tissue following 1.5 ppm zinc exposure (p < 0.01). A high level of zinc ion accumulation in the brain may induce deteriorative effects; thus, we addressed this question by measuring levels of oxidative species (H2O2 and ROS), anti-oxidative enzymes (GSH, GSH-Px, and SOD), lipid-peroxidation-related side products (MDA, thiobarbituric acid reactive substances (TBARS), and 4-hydroxynonenal (4-HNE)), DNA damage marker 8-hydroxy-2′-deoxyguanosine (8-OH-dG), and stress hormones (catecholamine and cortisol) in the brain tissue of ZnCl2-exposed fish. Results show that oxidative species levels were elevated in the brain (Figure 7C,D). Furthermore, an excessive level of active free radicals induced lipid peroxidation, which was consistent with high levels of MDA (Figure 7E), TBARS (p < 0.001, Figure 7F), and 4-HNE (Figure 7G) in the brains of ZnCl2-exposed fish. In addition, the persistent high oxidative stress impaired anti-oxidative capacity, which greatly reduced GSH (Figure 7I), GSH-Px (Figure 7J), and SOD (Figure 7K) enzyme activities. By measuring stress hormone levels, we found that brain catecholamine and cortisol levels were significantly elevated fivefold more than the untreated group, which indicated that a low dose of ZnCl2 exposure was able to induce significantly high stress in fish (Figure 7L,M). The sleeping-related hormone (melatonin) level was reduced in the brains of ZnCl2-exposed fish (Figure 7M), unlike the house-keeping protein, creatinine, which showed a constant level across tissues, providing evidence of the uniformity and consistency of protein extraction and quantification among samples (Figure 7O).
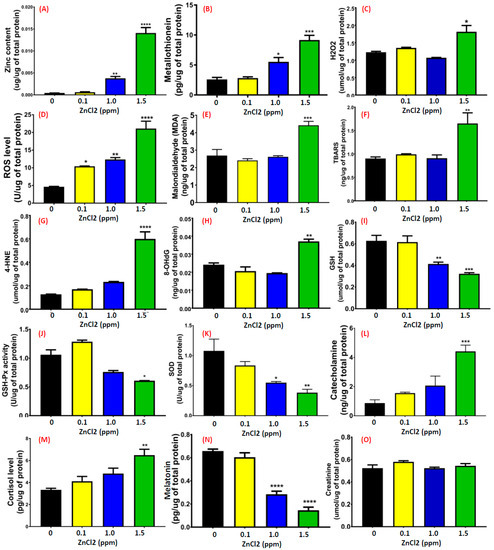
Figure 7.
Detection of biochemical parameters in the brain. Enzymatic or ELISA-based methods were applied to detect the levels of (A) zinc ions, (B) metallothionein, (C) reactive oxygen species (ROS), (D) H2O2, (E) malondialdehyde (MDA), (F) thiobarbituric acid reactive substances (TBARS), (G) 4-hydroxynonenal (4-HNE), (H) DNA damage marker 8-hydroxy-2′-deoxyguanosine (8-OH-dG), (I) glutathione (GSH), (J) GSH peroxidase (GSH-Px), (K) superoxide dismutase (SOD), (L) catecholamine, (M) cortisol, (N) melatonin, and (O) creatinine after treatment with various concentrations of ZnCl2. Data are represented as means ± SEM of four independent experiments. Statistical analysis was compared to control using a t-test (* p ≤ 0.05; ** p ≤ 0.01; *** p ≤ 0.001; **** p ≤ 0.0001).
2.8. ZnCl2 Causes the Downregulation of Neurotransmitter and AD-Related Marker Expression
Behavioral abnormalities, such as lower memory retention, lower aggressiveness, lower locomotion activity, and circadian rhythm dysregulation in low-dose ZnCl2-exposed fish led us to ask whether this phenotype was correlated with the downregulation of neurotransmitter and/or other AD-related markers expression. To address this question, we measured the relative content of several important neurotransmitters, such as AChE, ACh, dopamine, GABA, glutamate, glycine, and AD-related proteins like amyloid beta 42 (Aβ42) and p-Tau in the brain using ELISA assays. As shown in Figure 8A,B, AChE activity and ACh levels were detected at various concentrations (0.1, 0.5, 1, 1.5, and 5 ppm) of ZnCl2 exposure in zebrafish brain. A significant elevation of AChE activity was detected even at a low dose of ZnCl2 exposure at 0.1 ppm (Figure 8A). Interestingly, exposure to 1 and 1.5 ppm ZnCl2 resulted in significantly lower ACh levels compared to control, as well as compared to 0.1 and 0.5 ppm exposure (p < 0.004, Figure 8B). Carrying out ELISA with antigen-specific antibodies, we found that dopamine (Figure 8C), glutamate (Figure 8D), and GABA (Figure 8E) levels were elevated, while glycine (Figure 8F) and histamine (Figure 8G) levels were reduced. Based on multiple behavioral endpoints measured, we hypothesized that ZnCl2-exposed zebrafish display AD-like symptoms, including slow locomotion activity, high anxiety levels, circadian rhythm dysregulation, and lower memory retention. By measuring two AD-related biomarkers in the brain, we confirmed that ZnCl2 exposure could significantly elevate the relative contents of Aβ42 (Figure 8H) and p-Tau (Figure 8I).
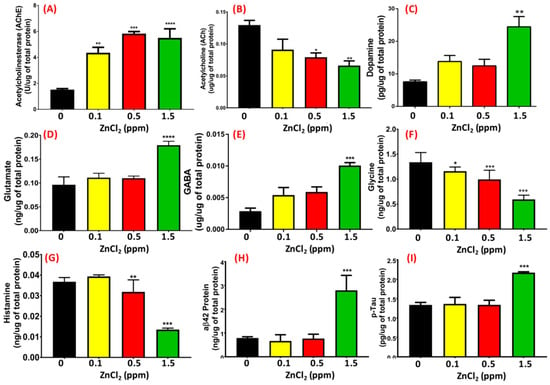
Figure 8.
Detection of neurotransmitter levels in the brain. (A) Acetylcholinesterase, (B) acetylcholine, (C) dopamine, (D) glutamate, (E) gamma-aminobutyric acid (GABA), (F) glycine, (G) histamine, (H) amyloid beta 42 (aβ42), and (I) phosphorylated (p)-Tau levels in the brain were measured using ELISA. Data are represented as means ± SEM. Statistical analysis was compared to control using a t-test (* p ≤ 0.05; ** p ≤ 0.01; *** p ≤ 0.001; **** p ≤ 0.0001).
3. Discussion
Zinc is widely considered as an essential trace element that is important for human health. In this study, we provided solid evidence showing that chronic ZnCl2 exposure can induce behavioral abnormalities and neurochemical changes in adult zebrafish (Figure 9). According to previous studies, the LC50 for 96 h of exposure to zinc was found to be 8.46 ppm in Puntius parrah and 1.35 ppm in white shrimp [44,45]. In addition, another study suggested that the LC50 value of zinc in Fundulus heteroclitus was 9.8–75.0 ppm [46]. Our results demonstrated that the 96-h LC50 in adult zebrafish was 6 ppm. We hypothesized that minor differences in LC50 might be associated with species, developmental stage, test medium, and age. Therefore, the discrepancy observed in LC50 was still reasonable.
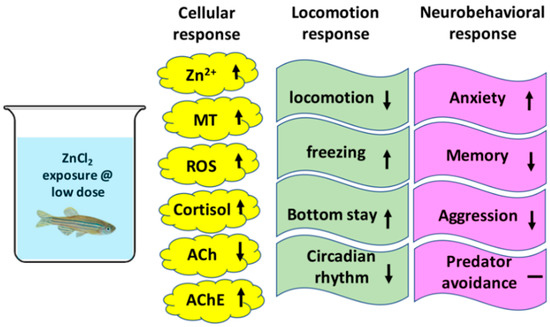
Figure 9.
Summary of the toxicological effects of ZnCl2 on cellular, locomotion, and neurobehavioral responses in adult zebrafish.
The zebrafish behavioral model is powerful, efficient, and simple for neuroscience research. Behavioral response was applied for decades as an indicator of toxicological impact within neurons [47,48,49]. Previous studies reported behavioral and locomotor activity alterations influenced by divalent zinc and/or other heavy metals in different species; however, only developmental toxicity of ZnCl2 in zebrafish embryos was addressed [50,51,52]. In contrast, our pioneer study demonstrated, for the first time, that adult zebrafish exposed to ZnCl2 had reduced locomotor behavior, average speed, entries to the upper zone, and total distance traveled. The average angular velocity and meandering of the fishes increased following 1 ppm ZnCl2 treatment compared to the control group, which was consistent with previous studies on other heavy-metal toxicity in zebrafish [53]. According to the literature research, we hypothesize that ZnCl2 exposure triggered an increase in muscle cortisol and reactive oxygen species (ROS) levels in the brains of zebrafish, which likely contributed to a stress-elicited reduction in locomotor activity [54].
Previous findings indicated that zinc was one of the potential factors causing senile plaques in the brain regions associated with AD [55]. Zinc at 300 nM showed effects of destabilizing Aβ aggregation and led to fibril formation, which caused damage in the cholinergic system. Our findings demonstrated that low concentrations (0.1 and 1 ppm) of ZnCl2 significantly impaired short-term memory in the inhibitory avoidance task, suggesting it caused significant neurotoxic effects. To investigate the mechanism related to the memory impairment induced by ZnCl2, we evaluated the activity of biomarkers, acetylcholine esterase (AChE), and acetylcholine (ACh) levels. We found that a low dose of ZnCl2 significantly elevated AChE activity and reduced ACh content in the brain tissue of zebrafish. A possible explanation is that ACh reduction may be caused by the excess hydrolysis activity of AChE, thereby altering neuromuscular activity and behavioral response. In addition, our biochemical results showed that levels of the metal-binding protein metallothionein, as well as those of ROS, glutamate, and glycine, were increased, which were associated with neuronal death and caused neurological defects [56]. Therefore, our zinc-induced zebrafish model showed potential as an AD-like model, and may provide a solid platform in neurological research.
Heavy metals are known to affect behavior, including impulsive aggression and anxiety [57,58,59]. In this study, to assess the effects of chronic ZnCl2 exposure on social behavior, we performed two behavioral paradigms, the mirror-biting and predator avoidance tests, and observed that the mirror-biting time was significantly reduced by sub-chronic exposure to ZnCl2 in adult animals (Figure 6B). Adult zebrafish responded promptly by escape and avoidance, which included freezing, erratic movements, jumping, or shoaling. Nonetheless, predator avoidance was not observed in ZnCl2-treated animals sub-chronically in our study; instead, anxiety and fear response toward the predator was expressed in zebrafish with an elevation of cortisol and catecholamine levels in the brain. On the other hand, circadian rhythms can be regulated by circadian genes in the brain or pineal gland in several vertebrates, including zebrafish [60,61]. In our study, the 1.5 ppm ZnCl2-exposed fish showed significantly different circadian patterns, with decreased locomotion during the light phase and increased locomotion during the dark phase. Our results showed that GABA expression levels increased and melatonin levels decreased in the brains of fish after exposure to ZnCl2. According to a previous study, GABA signals blocked by an antagonist increased circadian rates and rhythm in suprachiasmatic nucleus (SCN) neuron cells [62]. Moreover, disturbance of the pineal melatonin secretion played a significant role in the pathogenesis of Alzheimer’s disease, which was postulated as a circadian disorder [63]. Therefore, we hypothesized that the difference during light/dark phases might be correlated with the inhibition of melatonin in the brains of treated fish, which warrants further investigation in the future.
Previous studies demonstrated that metallothionein (MT) plays an important role in metal detoxification [64] and the homeostasis of zinc and copper [65,66]. Therefore, MTs are considered as a biomarker of environmental pollution of metals, which is consistent with the induction of MTs after chronic exposure to ZnCl2 in our study [67,68]. Furthermore, we provided a comprehensive collection of data to show that chronic ZnCl2 exposure elevated oxidative stress, as well as lipid oxidation and reactive oxygen species (ROS) levels, which resulted in MT induction.
AD is highly associated with oxidative stress and the overproduction of amyloid Aβ, while ROS levels are significantly higher than seen in healthy normal brains [69]. Over-accumulation of metal ions, inflammation, and aggregation of amyloid Aβs contribute to the high level of ROS in the brain. H2O2, a major component of ROS, is produced through the Fenton reaction of amyloid Aβ with metal ions and causes the accumulation of inflammatory cytokines such as tumor necrosis factor α (TNFα) and interleukin-1β (IL-1β), which attract active plaques [70]. To investigate the possible mechanism related to memory and other behavioral deficits induced by ZnCl2 exposure in zebrafish, we evaluated the impact of ZnCl2 on the cholinergic system in the brain. Our results showed that adult animals exposed to ZnCl2 had increased activity of neurotransmitters, such as GABA, dopamine, and serotonin, in the brain. Moreover, the inhibition of ACh content in the brain tissue caused behavioral abnormalities. ZnO and Al2O3 nanoparticles induced DNA damage in human peripheral blood lymphocytes [71]. In this study, single-stranded DNA damage was identified in the brains of ZnCl2-treated fish. Our results showed that the brain might be very sensitive to damage at the DNA level following ZnCl2 exposure, which impaired memory, decreased locomotion performance, and increased anxiogenic behavior, as well as inducing oxidative stress in brain. Therefore, we suggest that the oxidative stress in the CNS due to zinc exposure may induce changes in social behavior through irreversible DNA damage.
In summary, behavioral and neurotoxicological evaluations of ZnCl2 were assessed using multiple biomarker endpoints in our model. The underlying mechanism of ZnCl2 toxicity in adult zebrafish resulted from an increase in in oxidative stress biomarker responses (ROS and H2O2) and a decrease in in antioxidant biomarker responses (SOD, GSH, and GSH-Px). We report novel behavioral and neurotoxic effects upon exposure to divalent zinc in adult zebrafish. Our results demonstrated significant differences in neuroinflammatory effects (TNF α and IL-1β) and single-stranded DNA damage, as well as reductions in locomotion and memory, and alterations in circadian rhythm, in adult zebrafish. Moreover, our study showed that an increase in zinc could serve as a catalyst for increased ROS production, which might be important in the toxicity of amyloid Aβ and p-Tau proteins in brain pathogenesis. These deficits may result in the elevated concentration of MTs and antioxidants within brain tissue, as well as AChE expression in the cholinergic transmission system. Due to the advantage of it being relatively easy to induce an AD-like disease model in adult zebrafish using low-dose ZnCl2 exposure, our AD-like neurotoxicological model may benefit future neurodegenerative studies. In addition, it would be worthwhile to unveil the mechanism of Aβ aggregation and Tau pathology using the zinc-induced fish model and applying it to clinical research.
4. Materials and Methods
4.1. Animal Ethics
All experimental protocols and procedures involving zebrafish were approved by the Committee for Animal Experimentation of the Chung Yuan Christian University (Number: CYCU104024, issue date 21 December 2015). All experiments were performed in accordance with the guidelines for laboratory animals.
4.2. Zebrafish Rearing
Wild-type AB strain adult zebrafish (Danio rerio) were maintained in a recirculating aquatic system at 28.5 °C with a 10/14-h dark/light cycle according to standards. Circulating water in the aquarium was filtered by reverse osmosis (pH 7.0–7.5). The zebrafish were fed twice a day with lab-grown brine shrimp. For behavioral tests, we used adult zebrafish aged around 6–7 months.
4.3. Acute Toxicity Test
Zinc chloride (ZnCl2) was purchased from Sigma-Aldrich Corp. (St. Louis, MO, USA). Healthy adult zebrafish were separated into groups of 14 animals aged between six and seven months, and were housed in 3-L tanks prior to treatment. To determine the LC50, the following ZnCl2 concentrations were used: 2, 4, 6, 8, 10, 12, and 15 mg/L at 24, 48, 72 and 96 h of exposure.
4.4. Chronic Toxicity Test
The concentrations of ZnCl2 (below 5 mg/L) were chosen according to the USEPA guidelines for the disposal of ZnCl2 as an industrial effluent in the environment. Adult fish were exposed to a nominal concentration of 0 (control group), as well as concentrations of 0.1, 0.5, 1, and 1.5 ppm ZnCl2 for 24 h, 96 h, and 21 days for acute, sub-chronic, and chronic exposures, respectively. The exposure water was changed daily. The zebrafish were fasted during the entire experiment’s duration. A series of behavioral tests (3D locomotion, predator avoidance, memory, aggression, and circadian rhythm) were conducted in all groups, and performance was compared between ZnCl2-treated and control groups (procedures are summarized in Figure 1).
4.5. Adult Neurobehavioral Analysis
Five behavioral assays were used to assess neuron function in adult zebrafish upon systematical exposure to low concentrations of ZnCl2. The test battery consisted of 3D locomotor activity (for acute toxicity test), passive avoidance (on day 9 of ZnCl2 exposure), aggression (on day 9 of exposure), predator escape/avoidance (on day 13 of exposure), and circadian rhythm (on day 21 of exposure) assays which are described below.
4.6. Zebrafish 3D Locomotor Activity
Three-dimensional locomotor activity was tracked at five separate time points after exposure to ZnCl2 for 1, 4, 24, 48, and 96 h. The tracking strategy for zebrafish 3D locomotion followed the protocol described in our previous study [39]. In each test, we routinely tested the 3D locomotion activity of six fish at each time point while holding them in an acrylic tank (20 × 20 × 20 cm with a 15-cm-high water level). We tested in triplicate and had a total number of 18 fish for both the untreated and ZnCl2-treated groups.
4.7. Passive Avoidance Test (Short-Term Memory Test)
To evaluate the effects of ZnCl2 on short-term memory impairment in zebrafish, the passive avoidance test was carried out after 96 h of exposure, following a previous protocol with minor modification [72]. The test included two sessions with an interval of 24 h between sessions. In each session, fish were placed individually in an acrylic tank (30 × 20 × 20 cm) divided by a door into black and white compartments with equal size. In the training session, the fish were placed in the white compartment with the door closed for one minute for environmental acclimatization and recognition. Next, the door between the compartments was lifted and the fish were allowed to cross over to the dark side of the tank. When fish passed into the dark compartment the door was again closed, and a mild electric shock (25 V, 1 mA) was subjected to the fish with a 5-s interval. When fish passed into the white compartment, fish were pushed back into the dark compartment and shocked again until the fish returned into the white compartment. The training was repeated for a maximum of three times or until the fish passed to the white side. The fish that completed the training were stored in a small plastic container (15 × 10 × 5 cm) and kept at 28 °C. After 24 h, the fish were put into the training apparatus and tested for latency to pass into the dark area. The latency observed in the test session was used as an indicator of short-term memory retention.
4.8. Aggressiveness Test
Aggressive behavior was determined in adult zebrafish based on the mirror test as previously described with minor modification [73,74]. The test tank (28 × 5 × 15 cm) was filled with 1 L of water where a mirror was located at the side of the tank. One minute after the fish were introduced into the tank, aggressive behaviors (average speed, maximum and minimum speed, distance traveled, freezing, swimming, and rapid time movement percentage, mirror-biting time percentage, and longest duration in mirror side percentage) were recorded for a period of 5 min.
4.9. Predator Avoidance Test
To analyze fear-like and escaping behavior in zebrafish, the predator avoidance test was performed based on the mirror test described previously with minor modification [75]. The test tank (28 × 5 × 15 cm) was filled with 1 L of water where a transparent separator was in the middle of the tank. The predator convict cichlid (Amatitlania nigrofasciata) was put into the left area and zebrafish were put into the right area. After the fish were introduced into the tank, the predator avoidance behaviors (average speed, maximum and minimum speed, distance traveled, freezing, swimming, and rapid time movement percentage, top/bottom ratio of time spent and traveling distance, predator approaching time percentage, and distance to predator separator on average) were recorded for a period of 5 min.
4.10. Circadian Rhythm Test
To analyze zebrafish locomotion activity during the day and night, we performed a circadian rhythm test based on a previously published method with some modifications [76]. To prevent any external disturbance, the tested fish were moved into a temperature-controlled incubator and kept at 28 °C. One specially designed light-emitting diode (LED) light box with conventional and infrared LED arrays was used as the bottom light source. One infrared-sensitive charge-coupled device (CCD; detection window: 700–1000 nm) with a maximum resolution of 1920 × 1080 pixels and a 30-fps frame rate was used for video recording (3206_1080P module, Shenzhen, China). In this experiment, we recorded zebrafish locomotion activity (average speed, average angular velocity, and meandering) for 1 min every hour and used the idTracker software to track fish movement trajectories according to our previously published method [39].
4.11. Biochemical Parameters
4.11.1. Tissue Preparation and Total Protein Determination
After performance of the behavioral experiments, fish were euthanized with a high concentration of 200 mg/L tricaine solution (MS-222). For each independent assay, the whole brain and tissue from the caudal peduncle were extracted. A pool of three zebrafish tissues was used to prepare an independent homogenate preparation for each sample, which was homogenized on ice in 50 volumes (v/w) of phosphate-buffered saline (PBS) at pH 7.2. Samples were then centrifuged at 15,000 rpm for 15 min at 4 °C, and the supernatant was kept in microtubes at −80 °C for further assays. Total protein concentration was determined using a Pierce BCA Protein Assay Kit (23225, Thermo Fisher Scientific, Waltham, MA, USA). The color formation was analyzed at 562 nm using a microplate reader (Multiskan GO, Thermo Fisher Scientific, Waltham, MA, USA).
4.11.2. Analysis of Zinc and Metallothionein (MT) Content in Tissues
Concentrations of zinc in the brain and other tissues were determined using a zinc calorimetric quantification kit (E011, Nanjing Jiancheng Bio-Engineering Institute Co., Ltd, Nanjing, China). Tissue samples were extracted and prepared using PBS according to the method described in the above section. Calibration curves were prepared using serial dilutions of stock solutions of 100 mg/L zinc standard. The color formation was analyzed at 630 nm using a microplate reader (Multiskan GO, Thermo Fisher Scientific, Waltham, MA, USA). A fish metallothionein (MT) ELISA kit was used to measure total MT in tissues according to the manufacturer’s instructions (ZG-E1562, Zgenebio Company, Taipei, Taiwan). The color change was measured spectrophotometrically at a wavelength of 450 nm. MT content was expressed as pg/mL.
4.11.3. Determination of Acetylcholine and Acetylcholinesterase in Brain Samples
ZnCl2-treated zebrafish were anesthetized and euthanized using 200 mg/L tricaine. The whole-brain tissue was homogenized on ice in 50 volumes (v/w) of PBS at pH 7.2 using a bullet blender tissue homogenizer (Next Advance, Inc., Troy, NY, USA). For the AChE activity analysis, a pool of brain tissue from three individual fish was considered as one sample. The tests were performed in triplicate using a total of nine fish per group to ensure consistency. The activity of the homogenate was measured using an AChE calorimetric kit (A024, Nanjing Jiancheng Bio-Engineering Institute Co., Ltd., Nanjing, China) in accordance with the instructions. Briefly, 10 μL of brain homogenate was placed onto the plate. Then, 100 μL of working solution containing AChE enzyme, assay buffer, dye reagent, and enzyme mix was added into each well. After incubation for 15 min, the color formation was analyzed at 412 nm using a microplate reader (Multiskan GO, Thermo Fisher Scientific, Waltham, MA, USA) and the AChE activity was expressed in μmol. ACh levels were determined using an ACh ELISA kit (ZG-E1585, Zgenebio Company, Taipei, Taiwan) according to the specifications of the manufacturer. The absorbance was analyzed at 450 nm using a microplate reader (Multiskan GO, Thermo Fisher Scientific, Waltham, MA, USA). Data were expressed as U/mg of total protein.
4.11.4. Quantification of Hydrogen Peroxide and Reactive Oxygen Species Levels in Brain Samples
Hydrogen peroxide (H2O2) (ZG-E1602, Zgenebio Company, Taipei, Taiwan) and the reactive oxygen species (ROS) ELISA kit (ZG-E1561, Zgenebio Company, Taipei, Taiwan) were applied to detect their respective components according to the manufacturer’s instructions, and absorbance was determined at 450 nm. These assay kits are based on the sandwich ELISA method that involves a specific antibody for the detection of the chemicals of interest.
4.11.5. Determination of MDA, TBARS, 4-HNE, 8-OH-dG, Catecholamine, and Cortisol Levels in Brain Samples
Under oxidative stress, levels of biomarkers for lipid peroxidation (malondialdehyde, TBARS, and 4-HNE), DNA damage marker 8-OH-dG, and stress hormones catecholamine and cortisol were measured in the brain homogenate samples. The production of MDA, TBARS, 4-HNE, and 8-OH-dG in the samples was measured using a commercial ELISA kit (ZG-E1592, ZG-E1605, ZG-E1603 and ZG-E1564, Zgenebio Company, Taipei, Taiwan) according to the manufacturer’s protocol. The levels of catecholamine (CA) and cortisol in the brain homogenate were determined using an ELISA kit (ZG-E1590 and ZG-E1575, Zgenebio Company, Taipei, Taiwan) according to the manufacturer’s protocol. The color change was measured spectrophotometrically at a wavelength of 450 nm.
4.11.6. Determination of Antioxidant Capacity in Brain Samples
The exposure of fish to ZnCl2 promotes the production of glutathione (GSH), glutathione peroxidase (GSH-Px), and superoxide dismutase (SOD) in the fish brain, and the relative amounts of each were measured using specific ELISA kits (ZG-E1622, ZG-E1601, ZG-E1604, Zgenebio Company, Taipei, Taiwan). The color change was measured spectrophotometrically at a wavelength of 450 nm.
4.11.7. Evaluation of Neurotransmitter Levels in Brain Samples
Levels of neurotransmitters (GABA, glutamate, glycine, dopamine, 5-hydroxytryptamine (5-HT), norepinephrine, histamine and melatonin) in the fish brain were determined using commercially available ELISA kits (ZG-E1574, ZG-E1588, ZG-E1587, ZG-E1753, ZG-E1572, ZG-E1571, ZG-E1586, ZG-E1597, Zgenebio Company, Taipei, Taiwan) according to the manufacturer’s instructions. The color change was measured spectrophotometrically at a wavelength of 450 nm.
4.11.8. Estimation of Amyloid Beta (Aβ-42) and p-Tau Levels in the Brain
Aβ42 and p-Tau are two markers for Alzheimer’s Disease, and their concentrations in the brain tissues were detected using ELISA kits (ZG-E1607 and ZG-E1618, Zgenebio Company, Taipei, Taiwan) according to the manufacturer’s instructions. The absorbance was analyzed at 450 nm using a microplate reader (Multiskan GO, Thermo Fisher Scientific).
4.11.9. Statistical Analysis
For statistical analysis, the experimental values were compared between groups and data were expressed as means ± standard error of the mean (SEM). The Prism software (Graph pad Software version 7, La Jolla, CA, USA) was used to determine significant differences between treated and control groups. Data were compared using the t-test, ANOVA, or non-parametric test, depending on data normality for significance determination. A significant difference was considered to be p < 0.001.
Supplementary Materials
The following are available online at http://www.mdpi.com/1422-0067/19/10/3195/s1, Video S1: Mirror-biting assay of adult zebrafish after exposure to different concentrations of ZnCl2. Video is sped up 10×; Video S2: Predator avoidance test of adult zebrafish after exposure to different concentrations of ZnCl2. Video is sped up 10×; Video S3: Circadian rhythm test of adult zebrafish after exposure to 1.5 ppm ZnCl2. Video is sped up 5×.
Author Contributions
E.H., Y.H.L. and C.D.H. conceived and designed the study. S.S., G.A., and S.J. performed the experiments. G.A., S.J. and B.P.S. analyzed the data. S.S. and S.T.L. wrote the paper. E.H., Y.H.L. and C.D.H. reviewed and edited the manuscript. All authors read and approved the manuscript and agree to be accountable for all aspects of the research in ensuring that the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Funding
This work was funded by the Ministry of Science and Technology (grant nos. MOST105-2313-B-033-001-MY3 and MOST107-2622-B-033-001-CC2) to C.D.H.
Conflicts of Interest
There authors declare no conflicts of interest.
Appendix A

Figure A1.
LC50 test of ZnCl2 in adult zebrafish. Adult zebrafish were incubated with different concentrations of ZnCl2 (2, 4, 6, 8, 10, 12, and 15 ppm) and the fish number was recorded every day to build up the mortality curve. LC50 curves at 24, 48, 72 and 96 h are labeled in blue, red, green, and purple, respectively.

Figure A2.
Comparison of zinc levels in different tissues after chronic exposure to ZnCl2 at different concentrations for 21 days. Adult zebrafish were incubated with different concentrations of ZnCl2, and the relative amounts of zinc accumulation in (A) brain, (B) muscle, (C) gill, and (D) liver tissues were measured using enzymatic assays. Data are represented as means ± SEM. Statistical analysis was compared to control using a t-test (** p ≤ 0.01; **** p ≤ 0.0001).

Table A1.
Comparison of present study with other animal models exposed to ZnCl2.
Table A1.
Comparison of present study with other animal models exposed to ZnCl2.
| Toxicant | Model Organism | Concentration | Study | Behavior Endpoints | Toxic Effects | LC50 | References |
|---|---|---|---|---|---|---|---|
| ZnCl2 | Mice | 7.5, 10, 15 mg/kg | In vivo | No | Cytotoxicity, head abnormalities | NA | [77] |
| ZnCl2 | Neuronal cells | 0.15 mmol/L | In vitro | No | Loss of viability | NA | [78] |
| ZnCl2 + ZnO | MDKC cells | 100 μg/mL | In vitro | No | Cytotoxicity, genotoxicity | NA | [79] |
| ZnCl2 | Mice | 10 ppm | In vivo | Yes | Depressive Behavior | NA | [80] |
| Zinc | Mice | equimolar | In vitro | No | Stimulate hippocampal microglia activation | NA | [20] |
| ZnCl2 | D. magna | 0.16 ppm | In vivo | No | Toxic effects on growth and reproduction | NA | [81] |
| ZnCl2 + ZnO | C. elegans larvae | 614 μM | In vivo | No | Genotoxicity, Apoptosis | NA | [82] |
| Zinc | Red sea bream | 2.5 ppm | In vivo | No | Toxic effects on development and growth | 4.3 ppm | [18] |
| ZnCl2 | Zebrafish embryo | 0.5 ppm | In vivo | No | Skeletal anomalies | NA | [83] |
| ZnCl2 | Zebrafish embryo | 0.1–0.5 ppm | In vivo | No | Developmental deformities | NA | [84] |
| This study Low concentration of ZnCl2 | Adult zebrafish | 0.1–1.5 ppm | In vivo | Yes, we used 5 behavioralparameters | Memory impairment, locomotor deficit, and high AChE content in brain | 6 ppm | This study |
References
- Martins, R.N.; Villemagne, V.; Sohrabi, H.R.; Chatterjee, P.; Shah, T.M.; Verdile, G.; Fraser, P.; Taddei, K.; Gupta, V.B.; Rainey-Smith, S.R. Alzheimer’s Disease: A Journey from Amyloid Peptides and Oxidative Stress, to Biomarker Technologies and Disease Prevention Strategies—Gains from AIBL and DIAN Cohort Studies. J. Alzheimers Dis. 2018, 62, 965–992. [Google Scholar] [CrossRef] [PubMed]
- Markesbery, W.R. Oxidative stress hypothesis in Alzheimer’s disease. Free Radic. Biol. Med. 1997, 23, 134–147. [Google Scholar] [CrossRef]
- Nitsch, R.M.; Blusztajn, J.K.; Pittas, A.G.; Slack, B.E.; Growdon, J.H.; Wurtman, R.J. Evidence for a membrane defect in Alzheimer disease brain. Proc. Natl. Acad. Sci. USA 1992, 89, 1671–1675. [Google Scholar] [CrossRef] [PubMed]
- Lovell, M.A. Zinc and Zinc Transport and Sequestration Proteins in the Brain in the Progression of Alzheimer’s Disease. In Neurochemical Mechanisms in Disease; Springer: Berlin, Germany, 2011; pp. 669–693. [Google Scholar]
- Divakaruni, A.S.; Wiley, S.E.; Rogers, G.W.; Andreyev, A.Y.; Petrosyan, S.; Loviscach, M.; Wall, E.A.; Yadava, N.; Heuck, A.P.; Ferrick, D.A. Thiazolidinediones are acute, specific inhibitors of the mitochondrial pyruvate carrier. Proc. Natl. Acad. Sci. USA 2013, 110, 5422–5427. [Google Scholar] [CrossRef] [PubMed]
- Tandon, L.; Ni, B.-F.; Ding, X.; Ehmann, W.; Kasarskis, E.; Markesbery, W. RNAA for arsenic, cadmium, copper, and molybdenum in CNS tissues from subjects with age-related neurodegenerative diseases. J. Radioanal. Nucl. Chem. 1994, 179, 331–339. [Google Scholar] [CrossRef]
- Thompson, C.; Markesbery, W.; Ehmann, W.; Mao, Y.; Vance, D. Regional brain trace-element studies in Alzheimer’s disease. Neurotoxicology 1988, 9, 1–7. [Google Scholar] [PubMed]
- Lovell, M.; Robertson, J.; Teesdale, W.; Campbell, J.; Markesbery, W. Copper, iron and zinc in Alzheimer’s disease senile plaques. J. Neurol. Sci. 1998, 158, 47–52. [Google Scholar] [CrossRef]
- DeGrado, T.R.; Kemp, B.J.; Pandey, M.K.; Jiang, H.; Gunderson, T.M.; Linscheid, L.R.; Woodwick, A.R.; McConnell, D.M.; Fletcher, J.G.; Johnson, G.B. First PET imaging studies with 63Zn-zinc citrate in healthy human participants and patients with Alzheimer disease. Mol. Imaging 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A.; Swomley, A.M.; Sultana, R. Amyloid β-peptide (1–42)-induced oxidative stress in Alzheimer disease: Importance in disease pathogenesis and progression. Antioxid. Redox Signal. 2013, 19, 823–835. [Google Scholar] [CrossRef] [PubMed]
- Murgia, C.; Lang, C.; Truong-Tran, A.; Grosser, D.; Jayaram, L.; Ruffin, R.; Perozzi, G.; Zalewski, P. Zinc and its specific transporters as potential targets in airway disease. Curr. Drug Targets 2006, 7, 607–627. [Google Scholar] [CrossRef] [PubMed]
- Zalewski, P.D. Zinc metabolism in the airway: Basic mechanisms and drug targets. Curr. Opin. Pharmacol. 2006, 6, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Zastrow, M.L.; Pecoraro, V.L. Designing hydrolytic zinc metalloenzymes. Biochemistry 2014, 53, 957–978. [Google Scholar] [CrossRef] [PubMed]
- Fay, M. Exposure to contaminant mixtures at US hazardous waste sites. WIT Trans. Ecol. Environ. 2005, 85. [Google Scholar] [CrossRef]
- Stankevičiūtė, M.; Sauliutė, G.; Svecevičius, G.; Kazlauskienė, N.; Baršienė, J. Genotoxicity and cytotoxicity response to environmentally relevant complex metal mixture (Zn, Cu, Ni, Cr, Pb, Cd) accumulated in Atlantic salmon (Salmo salar). Part I: Importance of exposure time and tissue dependence. Ecotoxicology 2017, 26, 1051–1064. [Google Scholar] [CrossRef] [PubMed]
- Sharaf, M.S.; Stevens, D.; Kamunde, C. Zinc and calcium alter the relationship between mitochondrial respiration, ROS and membrane potential in rainbow trout (Oncorhynchus mykiss) liver mitochondria. Aquat. Toxicol. 2017, 189, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Xiao, Y.-Y.; Ji, Y.-H.; Liu, M.-Z.; Chen, Y.; Zeng, Y.-L.; Zhang, Y.-G.; Jin, L. CuInS2/ZnS QD exposure induces developmental toxicity, oxidative stress and DNA damage in rare minnow (Gobiocypris rarus) embryos and larvae. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 198, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Cao, L.; Shan, X.; Xiao, Z.; Wang, Q.; Dou, S. Toxic effects of zinc on the development, growth, and survival of red sea bream Pagrus major embryos and larvae. Arch. Environ. Contam. Toxicol. 2010, 58, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Elumalai, M.; Antunes, C.; Guilhermino, L. Enzymatic biomarkers in the crab Carcinus maenas from the Minho River estuary (NW Portugal) exposed to zinc and mercury. Chemosphere 2007, 66, 1249–1255. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-C.; Yu, W.-C.; Shih, Y.-H.; Chen, C.-Y.; Guo, Z.-H.; Huang, S.-J.; Chan, J.C.; Chen, Y.-R. Zinc ion rapidly induces toxic, off-pathway amyloid-β oligomers distinct from amyloid-β derived diffusible ligands in Alzheimer’s disease. Sci. Rep. 2018, 8, 4772. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.C.; Wu, P.c.; Chung, I.f.; Jiang, J.H.; Fann, M.J.; Kao, L.S. Cell death caused by the synergistic effects of zinc and dopamine is mediated by a stress sensor gene Gadd45b–implication in the pathogenesis of Parkinson’s disease. J. Neurochem. 2016, 139, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Cai, A.L.; Zipfel, G.J.; Sheline, C.T. Zinc neurotoxicity is dependent on intracellular NAD+ levels and the sirtuin pathway. Eur. J. Neurosci. 2006, 24, 2169–2176. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.W.; Koh, J.Y. Zinc and brain injury. Annu. Rev. Neurosci. 1998, 21, 347–375. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, T.; Kimura, K.; Morioka, S.; Tamaki, N. Inhibitory effects of Zn2+ on muscle glycolysis and their reversal by histidine. J. Nutr. Sci. Vitaminol. 1980, 26, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Krotkiewska, B.; Banas, T. Interaction of Zn2+, and Cu2+, Ions with glyceraldehyde-3-phosphate dehydrogenase from bovine heart and rabbit muscle. Int. J. Biochem. 1992, 24, 1501–1505. [Google Scholar] [CrossRef]
- Dineley, K.; Brocard, J.; Reynolds, I. Elevated intracellular zinc and altered proton homeostasis in forebrain neurons. Neuroscience 2002, 114, 439–449. [Google Scholar] [CrossRef]
- Conte-Daban, A.; Day, A.; Faller, P.; Hureau, C. How Zn can impede Cu detoxification by chelating agents in Alzheimer’s disease: A proof-of-concept study. Dalton Trans. 2016, 45, 15671–15678. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.-I.; Kawahara, M. Copper enhances zinc-induced neurotoxicity and the endoplasmic reticulum stress response in a neuronal model of vascular dementia. Front. Neurosci. 2017, 11, 58. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Zhu, L.; Dong, M.; Wang, J.; Wang, J.; Xie, H.; Liu, T.; Guo, Y. Oxidative stress and genotoxicity of the ionic liquid 1-octyl-3-methylimidazolium bromide in zebrafish (Danio rerio). Arch. Environ. Contam. Toxicol. 2014, 67, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Costa-Silva, D.; Nunes, M.M.; Wallau, G.; Martins, I.; Zemolin, A.P.; Cruz, L.; Rodrigues, N.; Lopes, A.; Posser, T.; Franco, J. Oxidative stress markers in fish (Astyanax sp. and Danio rerio) exposed to urban and agricultural effluents in the Brazilian Pampa biome. Environ. Sci. Pollut. Res. 2015, 22, 15526–15535. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, R.; Das, S.K.; Patri, M. Modulation of benzo [a]pyrene induced anxiolytic-like behavior by retinoic acid in zebrafish: Involvement of oxidative stress and antioxidant defense system. Neurotox. Res. 2017, 31, 493–504. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, J.J. Cholinergic control of GABA release: Emerging parallels between neocortex and hippocampus. Trends Neurosci. 2008, 31, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, C.; Chatonnet, A.; Takke, C.; Yan, Y.; Postlethwait, J.; Toutant, J.-P.; Cousin, X. Zebrafish acetylcholinesterase is encoded by a single gene localized on linkage group 7 gene structure and polymorphism; molecular forms and expression pattern during development. J. Biol. Chem. 2001, 276, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Senger, M.R.; Rico, E.P.; de Bem Arizi, M.; Frazzon, A.P.G.; Dias, R.D.; Bogo, M.R.; Bonan, C.D. Exposure to Hg2+ and Pb2+ changes NTPDase and ecto-5′-nucleotidase activities in central nervous system of zebrafish (Danio rerio). Toxicology 2006, 226, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Rico, E.P.; Rosemberg, D.B.; Senger, M.R.; de Bem Arizi, M.; Bernardi, G.F.; Dias, R.D.; Bogo, M.R.; Bonan, C.D. Methanol alters ecto-nucleotidases and acetylcholinesterase in zebrafish brain. Neurotoxicol. Teratol. 2006, 28, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Colovic, M.B.; Krstic, D.Z.; Lazarevic-Pasti, T.D.; Bondzic, A.M.; Vasic, V.M. Acetylcholinesterase inhibitors: Pharmacology and toxicology. Curr. Neuropharmacol. 2013, 11, 315–335. [Google Scholar] [CrossRef] [PubMed]
- Dix, D.J.; Houck, K.A.; Martin, M.T.; Richard, A.M.; Setzer, R.W.; Kavlock, R.J. The ToxCast program for prioritizing toxicity testing of environmental chemicals. Toxicol. Sci. 2006, 95, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Boran, H.; Ulutas, G. Genotoxic effects and gene expression changes in larval zebrafish after exposure to ZnCl2 and ZnO nanoparticles. Dis. Aquat. Organ. 2016, 117, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Audira, G.; Sampurna, B.P.; Juniardi, S.; Liang, S.-T.; Lai, Y.-H.; Hsiao, C.-D. A simple setup to perform 3D locomotion tracking in zebrafish by using a single camera. Inventions 2018, 3, 11. [Google Scholar] [CrossRef]
- Levin, E.D.; Bencan, Z.; Cerutti, D.T. Anxiolytic effects of nicotine in zebrafish. Physiol. Behav. 2007, 90, 54–58. [Google Scholar] [CrossRef] [PubMed]
- Pham, M.; Raymond, J.; Hester, J.; Kyzar, E.; Gaikwad, S.; Bruce, I.; Fryar, C.; Chanin, S.; Enriquez, J.; Bagawandoss, S. Assessing social behavior phenotypes in adult zebrafish: Shoaling, social preference, and mirror biting tests. In Zebrafish Protocols for Neurobehavioral Research; Springer: Berlin, Germany, 2012; pp. 231–246. [Google Scholar]
- Maximino, C.; da Silva, A.W.B.; Gouveia, A., Jr.; Herculano, A.M. Pharmacological analysis of zebrafish (Danio rerio) scototaxis. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Serra, E.; Medalha, C.; Mattioli, R. Natural preference of zebrafish (Danio rerio) for a dark environment. Braz. J. Med. Biol. Res. 1999, 32, 1551–1553. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.P.; Chen, H.-C. Effects of cadmium and zinc on oxygen consumption, ammonium excretion, and osmoregulation of white shrimp (Litopenaeus vannamei). Chemosphere 2004, 57, 1591–1598. [Google Scholar] [CrossRef] [PubMed]
- Ciji, P.; Nandan, S.B. Toxicity of copper and zinc to Puntius parrah (Day, 1865). Mar. Environ. Res. 2014, 93, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Loro, V.L.; Nogueira, L.; Nadella, S.R.; Wood, C.M. Zinc bioaccumulation and ionoregulatory impacts in Fundulus heteroclitus exposed to sublethal waterborne zinc at different salinities. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2014, 166, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, K.; Oshima, Y.; Yamaguchi, T.; Tsuruda, Y.; Kang, I.J.; Kobayashi, M.; Imada, N.; Honjo, T. Fertilization success and sexual behavior in male medaka, Oryzias latipes, exposed to tributyltin. Chemosphere 2004, 55, 1331–1337. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-S.; Chung, N.-I.; Choi, K.-H.; Cha, E.Y.; Lee, S.-K.; Chon, T.-S. Computational characterization of behavioral response of medaka (Oryzias latipes) treated with diazinon. Aquat. Toxicol. 2005, 71, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Azizullah, A.; Richter, P.; Häder, D.-P. Comparative toxicity of the pesticides carbofuran and malathion to the freshwater flagellate Euglena gracilis. Ecotoxicology 2011, 20, 1442–1454. [Google Scholar] [CrossRef] [PubMed]
- Garaventa, F.; Gambardella, C.; Di Fino, A.; Pittore, M.; Faimali, M. Swimming speed alteration of Artemia sp. and Brachionus plicatilis as a sub-lethal behavioural end-point for ecotoxicological surveys. Ecotoxicology 2010, 19, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Makaras, T.; Svecevičius, G.; Kazlauskienė, N.; Montvydienė, D. Rapid Detection of Sublethal Toxicity Using Locomotor Activity of Rainbow Trout Juveniles. Bull. Environ. Contam. Toxicol. 2018, 100, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Tu, H.; Fan, C.; Chen, X.; Liu, J.; Wang, B.; Huang, Z.; Zhang, Y.; Meng, X.; Zou, F. Effects of cadmium, manganese, and lead on locomotor activity and neurexin 2a expression in zebrafish. Environ. Toxicol. Chem. 2017, 36, 2147–2154. [Google Scholar] [CrossRef] [PubMed]
- Abreu, M.S.; Giacomini, A.C.; Rodriguez, R.; Kalueff, A.V.; Barcellos, L.J. Effects of ZnSO4-induced peripheral anosmia on zebrafish behavior and physiology. Behav. Brain Res. 2017, 320, 275–281. [Google Scholar] [CrossRef] [PubMed]
- Mocelin, R.; Herrmann, A.P.; Marcon, M.; Rambo, C.L.; Rohden, A.; Bevilaqua, F.; de Abreu, M.S.; Zanatta, L.; Elisabetsky, E.; Barcellos, L.J. N-acetylcysteine prevents stress-induced anxiety behavior in zebrafish. Pharmacol. Biochem. Behav. 2015, 139, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Deibel, M.; Ehmann, W.; Markesbery, W. Copper, iron, and zinc imbalances in severely degenerated brain regions in Alzheimer’s disease: Possible relation to oxidative stress. J. Neurol. Sci. 1996, 143, 137–142. [Google Scholar] [CrossRef]
- Tekkök, S.B.; Ye, Z.; Ransom, B.R. Excitotoxic mechanisms of ischemic injury in myelinated white matter. J. Cereb. Blood Flow Metab. 2007, 27, 1540–1552. [Google Scholar] [CrossRef] [PubMed]
- De Matos Mansur, B.; Cavalcante, C.N.S.; dos Santos, B.R.; Gouveia, A. Effects of mercury chloride (HgCl2) on Betta splendens aggressive display. Span. J. Psychol. 2012, 15, 442–450. [Google Scholar] [CrossRef]
- Weber, D.N.; Ghorai, J.K. Experimental design affects social behavior outcomes in adult zebrafish developmentally exposed to lead. Zebrafish 2013, 10, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Scott, G.R.; Sloman, K.A. The effects of environmental pollutants on complex fish behaviour: Integrating behavioural and physiological indicators of toxicity. Aquat. Toxicol. 2004, 68, 369–392. [Google Scholar] [CrossRef] [PubMed]
- Bégay, V.R.; Falcón, J.; Cahill, G.M.; Klein, D.C.; Coon, S.L. Transcripts encoding two melatonin synthesis enzymes in the teleost pineal organ: Circadian regulation in pike and zebrafish, but not in trout. Endocrinology 1998, 139, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Noche, R.R.; Lu, P.-N.; Goldstein-Kral, L.; Glasgow, E.; Liang, J.O. Circadian rhythms in the pineal organ persist in zebrafish larvae that lack ventral brain. BMC Neurosci. 2011, 12, 7. [Google Scholar] [CrossRef] [PubMed]
- Aton, S.J.; Huettner, J.E.; Straume, M.; Herzog, E.D. GABA and Gi/o differentially control circadian rhythms and synchrony in clock neurons. Proc. Natl. Acad. Sci. USA 2006, 103, 19188–19193. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, D.P.; Furio, A.M.; Brusco, L.I. Clinical aspects of melatonin intervention in Alzheimer’s disease progression. Curr. Neuropharmacol. 2010, 8, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Morris, S.; Dinman, J.; Pine, R.; Smith, B. Role of metallothionein in detoxification and tolerance to transition metals. In Metallothionein II; Springer: Berlin, Germany, 1987; pp. 439–446. [Google Scholar]
- De Lisle, R.C.; Sarras, M.P., Jr.; Hidalgo, J.; Andrews, G.K. Metallothionein is a component of exocrine pancreas secretion: Implications for zinc homeostasis. Am. J. Physiol. Cell Physiol. 1996, 271, C1103–C1110. [Google Scholar] [CrossRef] [PubMed]
- Pearce, L.L.; Wasserloos, K.; Croix, C.M.S.; Gandley, R.; Levitan, E.S.; Pitt, B.R. Metallothionein, nitric oxide and zinc homeostasis in vascular endothelial cells. J. Nutr. 2000, 130, 1467S–1470S. [Google Scholar] [CrossRef] [PubMed]
- Morris, C.A.; Stürzenbaum, S.; Nicolaus, B.; Morgan, A.J.; Harwood, J.L.; Kille, P. Identification and characterisation of metallothioneins from environmental indicator species as potential biomonitors. In Metallothionein IV; Springer: Berlin, Germany, 1999; pp. 621–627. [Google Scholar]
- Knapen, D.; Reynders, H.; Bervoets, L.; Verheyen, E.; Blust, R. Metallothionein gene and protein expression as a biomarker for metal pollution in natural gudgeon populations. Aquat. Toxicol. 2007, 82, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Praticò, D. Oxidative stress hypothesis in Alzheimer’s disease: A reappraisal. Trends Pharmacol. Sci. 2008, 29, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Condello, C.; Yuan, P.; Schain, A.; Grutzendler, J. Microglia constitute a barrier that prevents neurotoxic protofibrillar Aβ42 hotspots around plaques. Nat. Commun. 2015, 6, 6176. [Google Scholar] [CrossRef] [PubMed]
- Sliwinska, A.; Kwiatkowski, D.; Czarny, P.; Milczarek, J.; Toma, M.; Korycinska, A.; Szemraj, J.; Sliwinski, T. Genotoxicity and cytotoxicity of ZnO and Al2O3 nanoparticles. Toxicol. Mech. Methods 2015, 25, 176–183. [Google Scholar] [CrossRef] [PubMed]
- Blank, M.; Guerim, L.D.; Cordeiro, R.F.; Vianna, M.R. A one-trial inhibitory avoidance task to zebrafish: Rapid acquisition of an NMDA-dependent long-term memory. Neurobiol. Learn. Mem. 2009, 92, 529–534. [Google Scholar] [CrossRef] [PubMed]
- Gerlai, R.; Lahav, M.; Guo, S.; Rosenthal, A. Drinks like a fish: Zebra fish (Danio rerio) as a behavior genetic model to study alcohol effects. Pharmacol. Biochem. Behav. 2000, 67, 773–782. [Google Scholar] [CrossRef]
- Moretz, J.A.; Martins, E.P.; Robison, B.D. The effects of early and adult social environment on zebrafish (Danio rerio) behavior. Environ. Biol. Fishes 2007, 80, 91–101. [Google Scholar] [CrossRef]
- Bass, S.L.; Gerlai, R. Zebrafish (Danio rerio) responds differentially to stimulus fish: The effects of sympatric and allopatric predators and harmless fish. Behav. Brain Res. 2008, 186, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Steenbergen, P.J.; Richardson, M.K.; Champagne, D.L. Patterns of avoidance behaviours in the light/dark preference test in young juvenile zebrafish: A pharmacological study. Behav. Brain Res. 2011, 222, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Gupta, T.; Talukder, G.; Sharma, A. Cytotoxicity of zinc chloride in mice in vivo. Biol. Trace Elem. Res. 1991, 30, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Zyśk, M.; Bielarczyk, H.; Gul-Hinc, S.; Dyś, A.; Gapys, B.; Ronowska, A.; Sakowicz-Burkiewicz, M.; Szutowicz, A. Phenotype-dependent interactions between N-acetyl-l-aspartate and acetyl-coa in septal sn56 cholinergic cells exposed to an excess of zinc. J. Alzheimer’s Dis. 2017, 56, 1145–1158. [Google Scholar] [CrossRef] [PubMed]
- Kononenko, V.; Repar, N.; Marušič, N.; Drašler, B.; Romih, T.; Hočevar, S.; Drobne, D. Comparative in vitro genotoxicity study of zno nanoparticles, zno macroparticles and ZnCl2 to mdck kidney cells: Size matters. Toxicol. In Vitro 2017, 40, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Manosso, L.M.; Moretti, M.; Colla, A.R.; Ribeiro, C.M.; Dal-Cim, T.; Tasca, C.I.; Rodrigues, A.L.S. Involvement of glutamatergic neurotransmission in the antidepressant-like effect of zinc in the chronic unpredictable stress model of depression. J. Neural Transm. 2016, 123, 339–352. [Google Scholar] [CrossRef] [PubMed]
- Adam, N.; Schmitt, C.; Galceran, J.; Companys, E.; Vakurov, A.; Wallace, R.; Knapen, D.; Blust, R. The chronic toxicity of zno nanoparticles and ZnCl2 to daphnia magna and the use of different methods to assess nanoparticle aggregation and dissolution. Nanotoxicology 2014, 8, 709–717. [Google Scholar] [PubMed]
- O’donnell, B.; Huo, L.; Polli, J.R.; Qiu, L.; Collier, D.N.; Zhang, B.; Pan, X. From the cover: Zno nanoparticles enhanced germ cell apoptosis in Caenorhabditis elegans, in comparison with ZnCl2. Toxicol. Sci. 2016, 156, 336–343. [Google Scholar]
- Salvaggio, A.; Marino, F.; Albano, M.; Pecoraro, R.; Camiolo, G.; Tibullo, D.; Bramanti, V.; Lombardo, B.M.; Saccone, S.; Mazzei, V. Toxic effects of zinc chloride on the bone development in Danio rerio (hamilton, 1822). Front. Physiol. 2016, 7, 153. [Google Scholar] [CrossRef] [PubMed]
- Küçükoğlu, M.; Binokay, U.S.; Pekmezekmek, A.B. The effects of zinc chloride during early embryonic development in zebrafish (Brachydanio rerio). Turkish J. Biol. 2013, 37, 158–164. [Google Scholar]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).