
Molecular Characterization, mRNA Expression and Alternative Splicing of Ryanodine Receptor Gene in the Brown Citrus Aphid, Toxoptera citricida (Kirkaldy)


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Cloning the Coding Sequence of Ryanodine Receptor (RyR) in Toxoptera citricida (TciRyR)
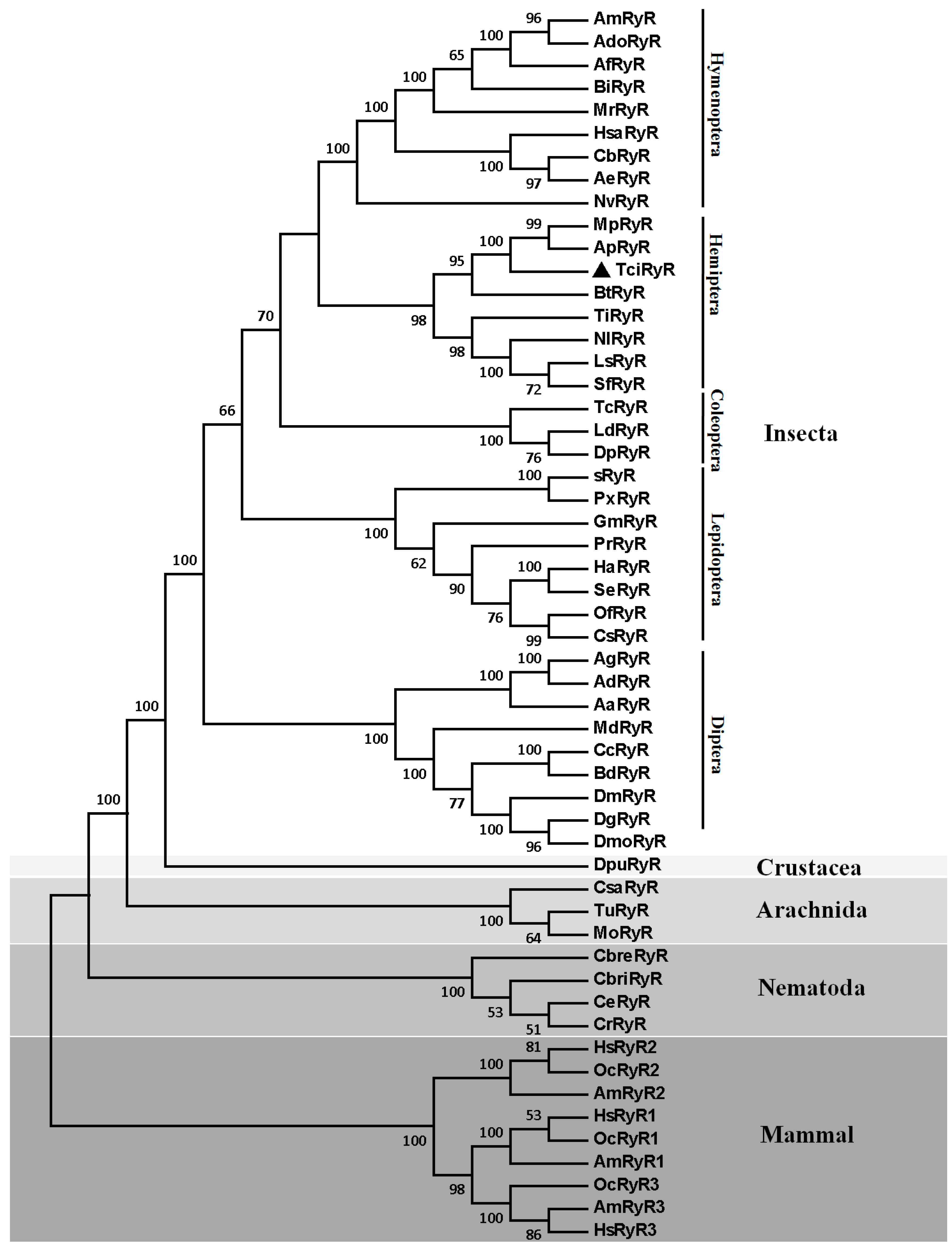
2.2. RyR Orthologs and Their Phylogeny
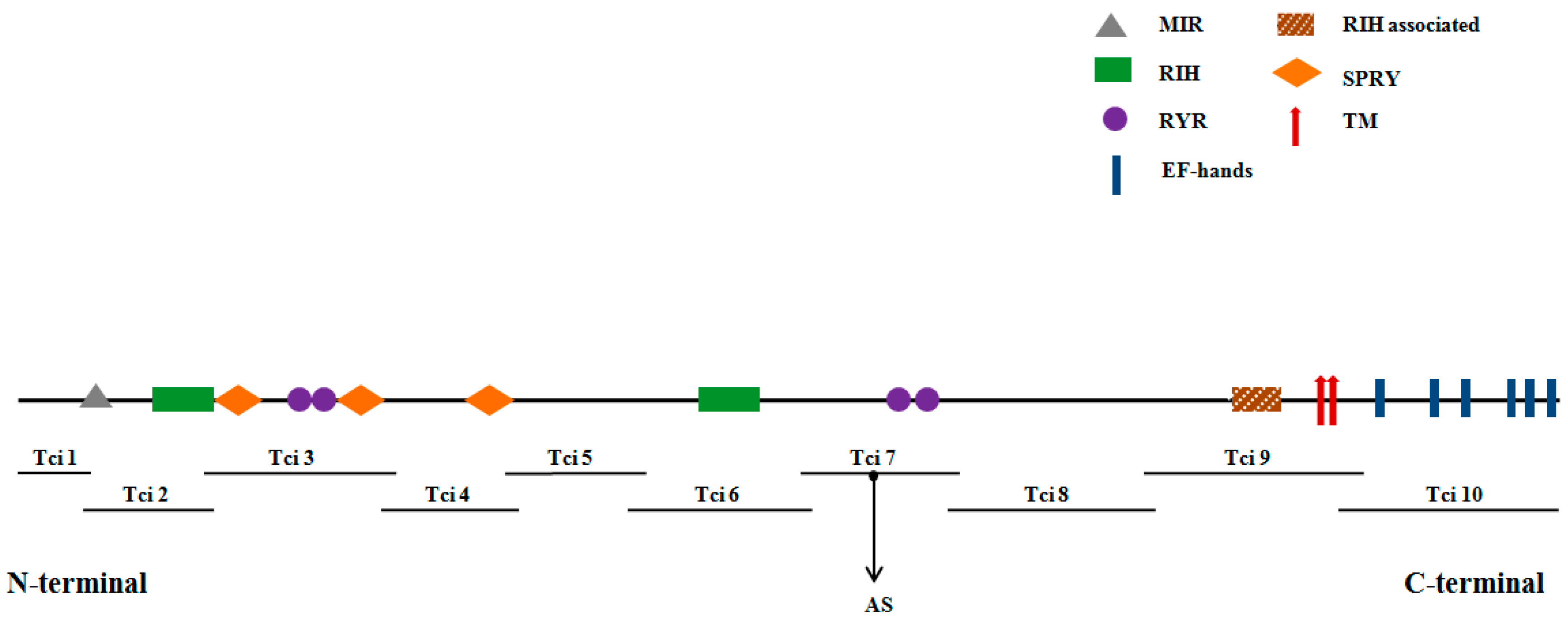
2.3. Conserved Structural Domains in TciRyR

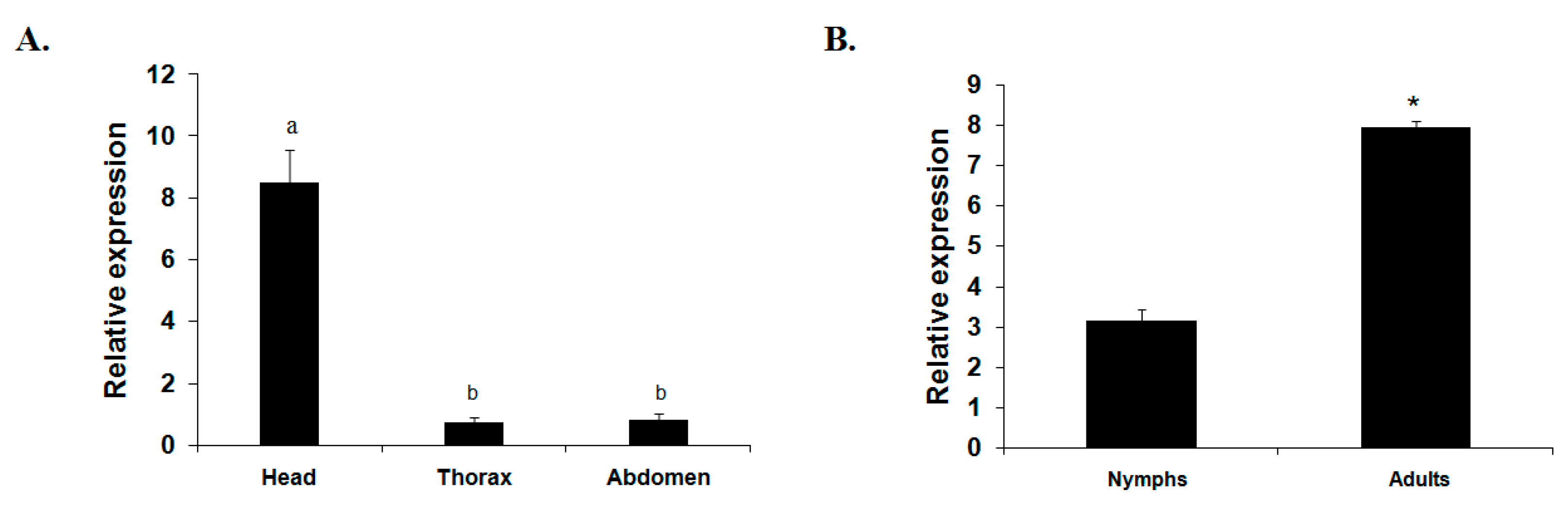
2.4. mRNA Expression Profiles of TciRyR
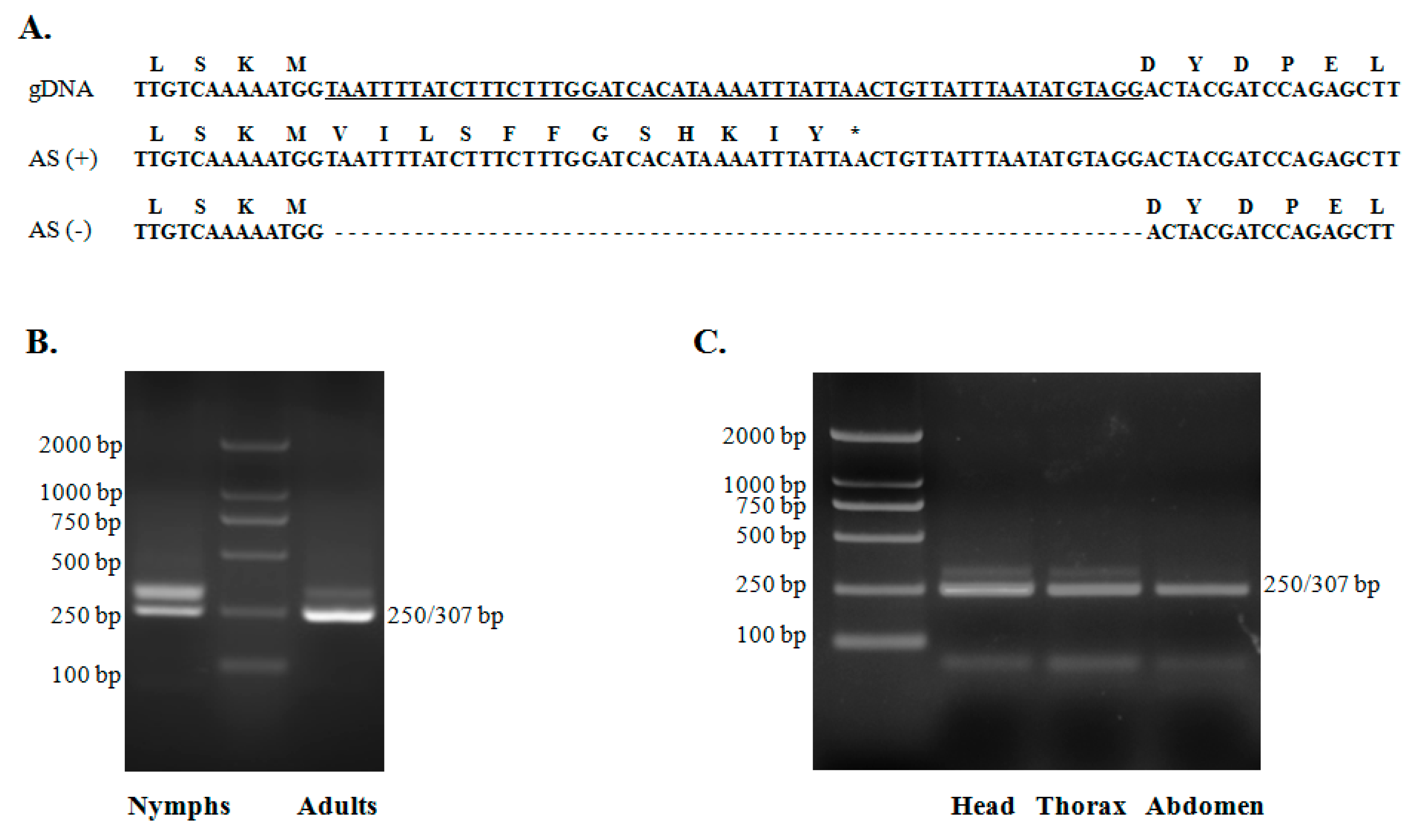
2.5. Alternative Splicing of TciRyR
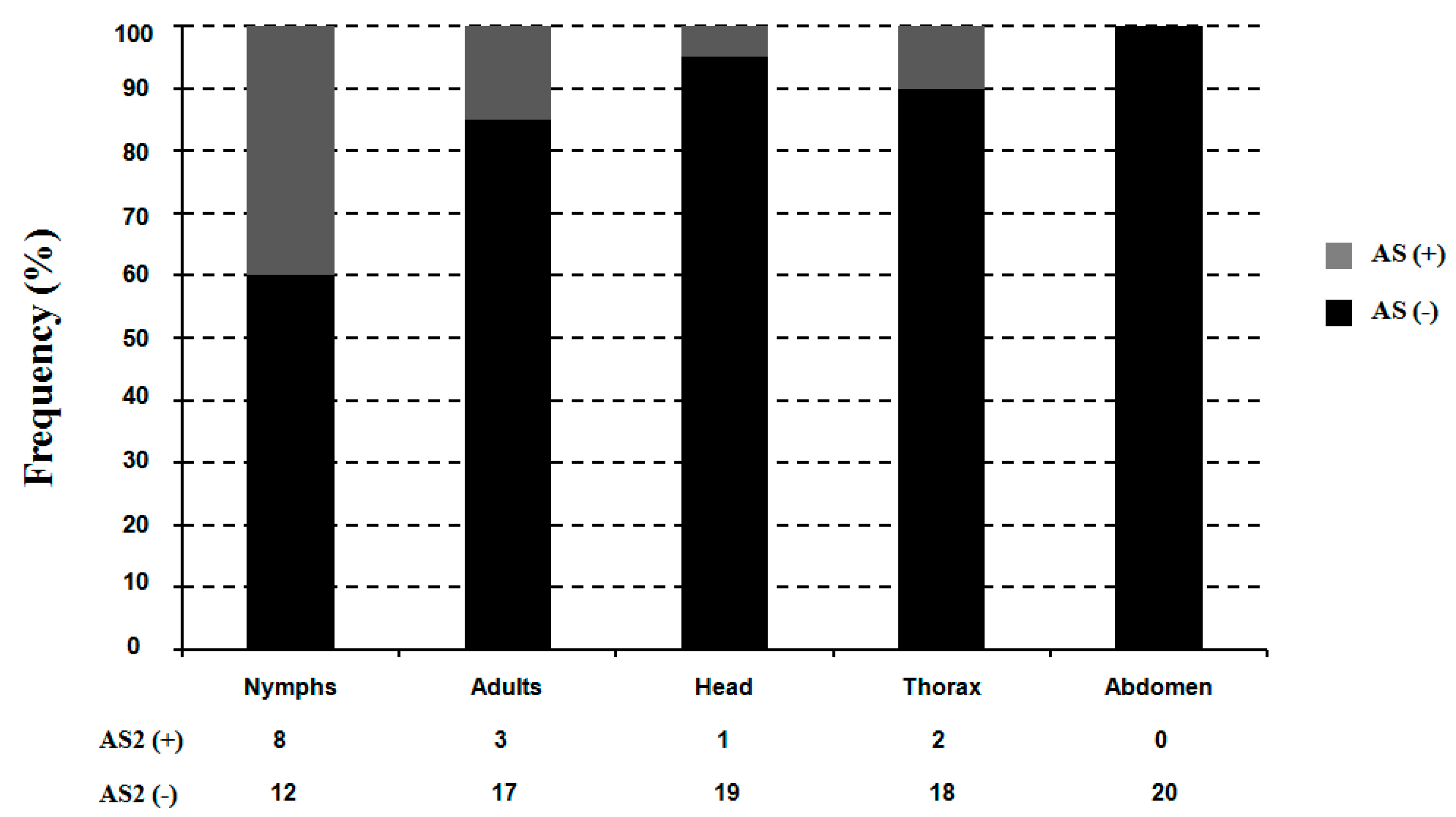
3. Discussion
4. Experimental Section
4.1. Insects
4.2. RNA Extraction and cDNA Synthesis
4.3. Cloning of RyR Genes from T. citricida
4.4. Sequence Analysis and Phylogenetic Tree Construction
4.5. Quantitative Real-Time PCR
4.6. Diagnostic PCR Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fill, M.; Copello, J.A. Ryanodine receptor calcium release channels. Physiol. Rev. 2002, 82, 893–922. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.J. Inositol trisphosphate and calcium signaling. Nature 1993, 361, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E. Calcium signaling. Cell 1995, 80, 259–268. [Google Scholar] [CrossRef]
- Takeshima, H. Primary structure and expression from cDNAs of the ryanodine receptor. Ann. N. Y. Acad. Sci. 1993, 707, 165–177. [Google Scholar] [CrossRef] [PubMed]
- Wagenknecht, T.; Grassucci, R.; Frank, J.; Saito, A.; Inui, M.; Fleischer, S. Three dimensional architecture of the calcium channel/foot structure of sarcoplasmic reticulum. Nature 1989, 338, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Bhat, M.B.; Zhao, J.; Takeshima, H.; Ma, J. Functional calcium release channel formed by the carboxyl-terminal portion of ryanodine receptor. Biophys. J. 1997, 73, 1329–1336. [Google Scholar] [CrossRef]
- Otsu, K.; Willard, H.F.; Khanna, V.K.; Zorzato, F.; Green, N.M.; MacLennan, D.H. Molecular cloning of cDNA encoding the Ca2+ release channel (ryanodine receptor) of rabbit cardiac muscle sarcoplasmic reticulum. J. Biol. Chem. 1990, 265, 13472–13483. [Google Scholar] [PubMed]
- Ogawa, Y.; Murayama, T.; Kurebayashi, N. Comparison of properties of Ca2+ release channels between rabbit and frog skeletal muscles. Mol. Cell. Biochem. 1999, 190, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Ottini, L.; Marziali, G.; Conti, A.; Charlesworth, A.; Sorrentino, V. Alpha and beta isoforms of ryanodine receptor from chicken skeletal muscle are the homologues of mammalian RyR1 and RyR3. Biochem. J. 1996, 315, 207–216. [Google Scholar] [PubMed]
- Takeshima, H.; Nishi, M.; Iwabe, N.; Miyata, T.; Hosoya, T.; Masai, I.; Hotta, Y. Isolation and characterization of a gene for a ryanodine receptor/calcium release channel in Drosophila melanogaster. FEBS Lett. 1994, 337, 81–87. [Google Scholar] [CrossRef]
- Xu, X.; Bhat, M.B.; Nishi, M.; Takeshima, H.; Ma, J. Molecular cloning of cDNA encoding a Drosophila ryanodine receptor and functional studies of the carboxyl-terminal calcium release channel. Biophys. J. 2000, 78, 1270–1281. [Google Scholar] [CrossRef]
- Tohnishi, M.; Nakao, H.; Furuya, T.; Seo, A.; Kodama, H. Flubendiamide, a novel insecticide highly active against Lepidopterous insect pests. J. Pestic. Sci. 2005, 30, 354–360. [Google Scholar] [CrossRef]
- Lahm, G.P.; Selby, T.P.; Freudenberger, J.H.; Stevenson, T.M.; Myers, B.J. Insecticidal anthranilic diamides: A new class of potent ryanodine receptor activators. Bioorg. Med. Chem. Lett. 2005, 15, 4898–4906. [Google Scholar] [CrossRef] [PubMed]
- Pepper, B.P.; Carruth, L.A. A new plant insecticide for control of the European corn borer. J. Econ. Entomol. 1945, 38, 59–66. [Google Scholar] [CrossRef]
- Li, X.C.; Degain, B.A.; Harpold, V.S.; Marcon, P.G.; Nichols, R.; Fournier, A.J.; Naranjo, S.E.; Palumbo, J.C.; Ellsworth, P.C. Baseline susceptibilities of B- and Q-biotype Bemisia tabaci to anthranilic diamides in Arizona. Pest Manag. Sci. 2012, 68, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.M.; Jiang, E.B.; He, S.Y.; Chen, J.H. Lethal and sublethal effects of cyantraniliproleon Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Pest Manag. Sci. 2015, 71, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, Y.; Han, Z.; Zhu, Y.; Xie, Z. Molecular characterization of a ryanodine receptor gene in the rice leaffolder, Cnaphalocrocis medinalis (Guenee). PLoS ONE 2012, 7, e36623. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xie, Z.; Gao, J.; Liu, Y.; Wang, W. Molecular cloning and characterization of a ryanodine receptor gene in brown plant hopper (BPH), Nilaparvata lugens (Stal). Pest Manag. Sci. 2014, 70, 790–797. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.R.; Shi, W.Z.; Yang, W.J. Molecular characteristics, mRNA expression, and alternative splicing of a ryanodine receptor gene in the Oriental Fruit Fly, Bactrocera dorsalis (Hendel). PLoS ONE 2014, 9, e95199. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wu, S.; Yang, Y.; Wu, Y. Molecular cloning, characterization and mRNA expression of a ryanodine receptor gene from diamondback moth, Plutella xylostella. Pestic. Biochem. Physiol. 2012, 102, 204–212. [Google Scholar] [CrossRef]
- Cui, L.; Yang, D.; Yan, X.; Rui, C.; Wang, Z. Molecular cloning, characterization and expression profiling of a ryanodine receptor gene in Asian corn borer, Ostrinia furnacalis (Guenee). PLoS ONE 2013, 8, e75825. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Y.; Gao, J.; Xie, Z.; Huang, L. Molecular cloning and mRNA expression of a ryanodine receptor gene in the cotton bollworm, Helicoverpa armigera. Pestic. Biochem. Physiol. 2013, 107, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Wang, F.; Huang, J.; Fang, Q.; Shen, Z. Molecular and cellular analyses of a ryanodine receptor from hemocytes of Pieris rapae. Dev. Comp. Immunol. 2013, 41, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Troczka, B.J.; Williams, A.J.; Bass, C. Molecular cloning, characterisation and mRNA expression of the ryanodine receptor from the peach-potato aphid, Myzus persicae. Gene 2015, 556, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Wan, P.J.; Guo, W.Y.; Yang, Y. RNAi suppression of the ryanodine receptor gene results in decreased susceptibility to chlorantraniliprole in Colorado potato beetle Leptinotarsa decemlineata. J. Insect Physiol. 2014, 63, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wan, P.J.; Hu, X.X. RNAi mediated knockdown of the ryanodine receptor gene decreases chlorantraniliprole susceptibility in Sogatella furcifera. Pestic. Biochem. Physiol. 2014, 108, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.P.; Li, C.J.; Gao, J.K.; Wang, W.L.; Li, H.; Guo, X.Z.; Li, B.; Wang, J.J. Comparative characterization of two intracellular Ca2+-release channels from the red flour beetle, Tribolium castaneum. Sci. Rep. 2014, 4, 6702. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Shahzad, M.F.; Zhang, L.; Li, F.; Lin, K.J. Amplifying long transcripts of ryanodine receptors of five agricultural pests by transcriptome analysis and gap filling. Genome 2013, 56, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Sattelle, D.B.; Cordova, D.; Cheek, T.R. Insect ryanodine receptors: molecular targets for novel pest control chemicals. Invertebr. Neurosci. 2008, 8, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Troczka, B.; Zimmer, C.T.; Elias, J.; Schorn, C.; Bass, C.; Davies, T.G. Resistance to diamide insecticides in diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae) is associated with a mutation in the membrane-spanning domain of the ryanodine receptor. Insect Biochem. Mol. Biol. 2012, 42, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Wang, Y.; Zhou, X. Functional analysis of a point mutation in the ryanodine receptor of Plutella xylostella (L.) associated with resistance to chlorantraniliprole. Pest Manag. Sci. 2014, 70, 1083–1089. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Pei, L.; Zhou, X.G.; Gao, X.W. Novel mutations and mutation combinations of ryanodine receptor in a chlorantraniliprole resistant population of Plutella xylostella (L.). Sci. Rep. 2014, 4, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Bai, X.C.; Yan, C.Y.; Wu, J.P.; Li, Z.Q.; Xie, T.; Peng, W.; Yin, C.C.; Li, X.M.; Scheres, H.W.; et al. Structure of the rabbit ryanodine receptor RyR1 at near-atomic resolution. Nature 2015, 517, 50–66. [Google Scholar] [CrossRef] [PubMed]
- Ikenoue, T.; Inoki, K.; Yang, Q. Essential function of TORC2 in PKC and Akt turn motif phosphorylation, maturation and signaling. EMBO J. 2008, 27, 1919–1931. [Google Scholar] [CrossRef] [PubMed]
- Ponting, C.P. Novel repeats in ryanodine and IP3 receptors and protein O-mannosyltransferases. Trends Biochem. Sci. 2000, 25, 47–50. [Google Scholar] [CrossRef]
- Cui, Y.; Tae, H.S.; Norris, N.C.; Karunasekara, Y.; Pouliquin, P.; Board, P.G.; Dulhunty, A.F.; Casarotto, M.G. A dihydropyridine receptor α1s loop region critical for skeletal muscle contraction is intrinsically unstructured and binds to a SPRY domain of the type 1 ryanodine receptor. Int. J. Biochem. Cell Biol. 2009, 41, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, V.; Barone, V.; Rossi, D. Intracellular Ca2+ release channels in evolution. Curr. Opin. Genet. Dev. 2000, 10, 662–667. [Google Scholar] [CrossRef]
- Zhao, M.; Li, P.; Li, X. Molecular identification of the ryanodine receptor pore-forming segment. J. Biol. Chem. 1999, 274, 25971–25974. [Google Scholar] [CrossRef] [PubMed]
- Xiao, B.; Jiang, M.T.; Zhao, M. Characterization of a novel PKA phosphorylation site, serine-2030, reveals no PKA hyperphosphorylation of the cardiac ryanodine receptor in canine heart failure. Circ. Res. 2005, 96, 847–855. [Google Scholar] [CrossRef] [PubMed]
- George, C.H.; Rogers, S.A.; Bertrand, B.M.A.; Tunwell, R.E.A.; Thomas, N.L. Alternative splicing of ryanodine receptors modulates cardiomyocyte Ca2+ signaling and susceptibility to apoptosis. Circ. Res. 2007, 100, 874–883. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Lueck, J.D.; Harvey, P.J.; Pace, S.M.; Ikemoto, N. Alternative splicing of RyR1 alters the efficacy of skeletal EC coupling. Cell Calcium 2009, 45, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Takasawa, S.; Kuroki, M.; Nata, K.; Noguchi, N.; Ikeda, T. A Novel ryanodine receptor expressed in pancreatic islets by alternative splicing from type 2 ryanodine receptor gene. Biochem. Biophys. Res. Commun. 2010, 397, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Southwest University, Chongqing, China. Unpublished work. 2015.
- Larkin, M.; Blackshields, G.; Brown, N.; Chenna, R.; McGettigan, P.A.; McWilliam, H. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein identification and analysis tools on the ExPASy server. Proteomics Protoc. Handb. 2005, 571–607. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; von Heijne, G.; Sonnhammer, E.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Marchler-Bauer, A.; Derbyshire, M.K.; Gonzales, N.R.; Lu, S.; Chitsaz, F.; Geer, L.Y.; Geer, R.C.; He, J.; Gwadz, M.; Hurwitz, D.I.; et al. CDD: NCBI’s conserved domain database. Nucleic Acids Res. 2015, 43, D222–D226. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.D.; Lee, T.Y.; Tseng, S.W. KinasePhos: A web tool for identifying protein kinase-specific phosphorylation sites. Nucleic Acids Res. 2005, 33, 226–229. [Google Scholar] [CrossRef] [PubMed]
- Blom, N.; Sicheritz-Pontén, T.; Gupta, R. Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence. Proteomics 2004, 4, 1633–1649. [Google Scholar] [CrossRef] [PubMed]
- Benson, D.A.; Clark, K.; Karsch-Mizrachi, I.; Lipman, D.J.; Ostell, J.; Sayers, E.W. GenBank. Nucleic Acids Res. 2015, 43, D30–D35. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Untergrasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer 3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef] [PubMed]
- Kenneth, L.J.; Thomas, S.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods 2001, 25, 402–408. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, K.-Y.; Jiang, X.-Z.; Yuan, G.-R.; Shang, F.; Wang, J.-J. Molecular Characterization, mRNA Expression and Alternative Splicing of Ryanodine Receptor Gene in the Brown Citrus Aphid, Toxoptera citricida (Kirkaldy). Int. J. Mol. Sci. 2015, 16, 15220-15234. https://doi.org/10.3390/ijms160715220
Wang K-Y, Jiang X-Z, Yuan G-R, Shang F, Wang J-J. Molecular Characterization, mRNA Expression and Alternative Splicing of Ryanodine Receptor Gene in the Brown Citrus Aphid, Toxoptera citricida (Kirkaldy). International Journal of Molecular Sciences. 2015; 16(7):15220-15234. https://doi.org/10.3390/ijms160715220
Chicago/Turabian StyleWang, Ke-Yi, Xuan-Zhao Jiang, Guo-Rui Yuan, Feng Shang, and Jin-Jun Wang. 2015. "Molecular Characterization, mRNA Expression and Alternative Splicing of Ryanodine Receptor Gene in the Brown Citrus Aphid, Toxoptera citricida (Kirkaldy)" International Journal of Molecular Sciences 16, no. 7: 15220-15234. https://doi.org/10.3390/ijms160715220
APA StyleWang, K.-Y., Jiang, X.-Z., Yuan, G.-R., Shang, F., & Wang, J.-J. (2015). Molecular Characterization, mRNA Expression and Alternative Splicing of Ryanodine Receptor Gene in the Brown Citrus Aphid, Toxoptera citricida (Kirkaldy). International Journal of Molecular Sciences, 16(7), 15220-15234. https://doi.org/10.3390/ijms160715220