
Metabolite Profiling and Antimicrobial Activities of Brassica rapa ssp. narinosa (Tatsoi), Brassica rapa var. narinosa × chinensis (Dacheongchae), and Brassica rapa ssp. chinensis (Pakchoi)

 ,
,  ,
,  ,
,  , and
, and
Abstract
1. Introduction
2. Results
2.1. Carotenoid Analysis of Tatsoi, Dacheongchae, and Pakchoi
2.2. Glucosinolate Analysis of Tatsoi, Dacheongchae, and Pakchoi
2.3. Phenolic Content Analysis of Tatsoi, Dacheongchae, and Pakchoi
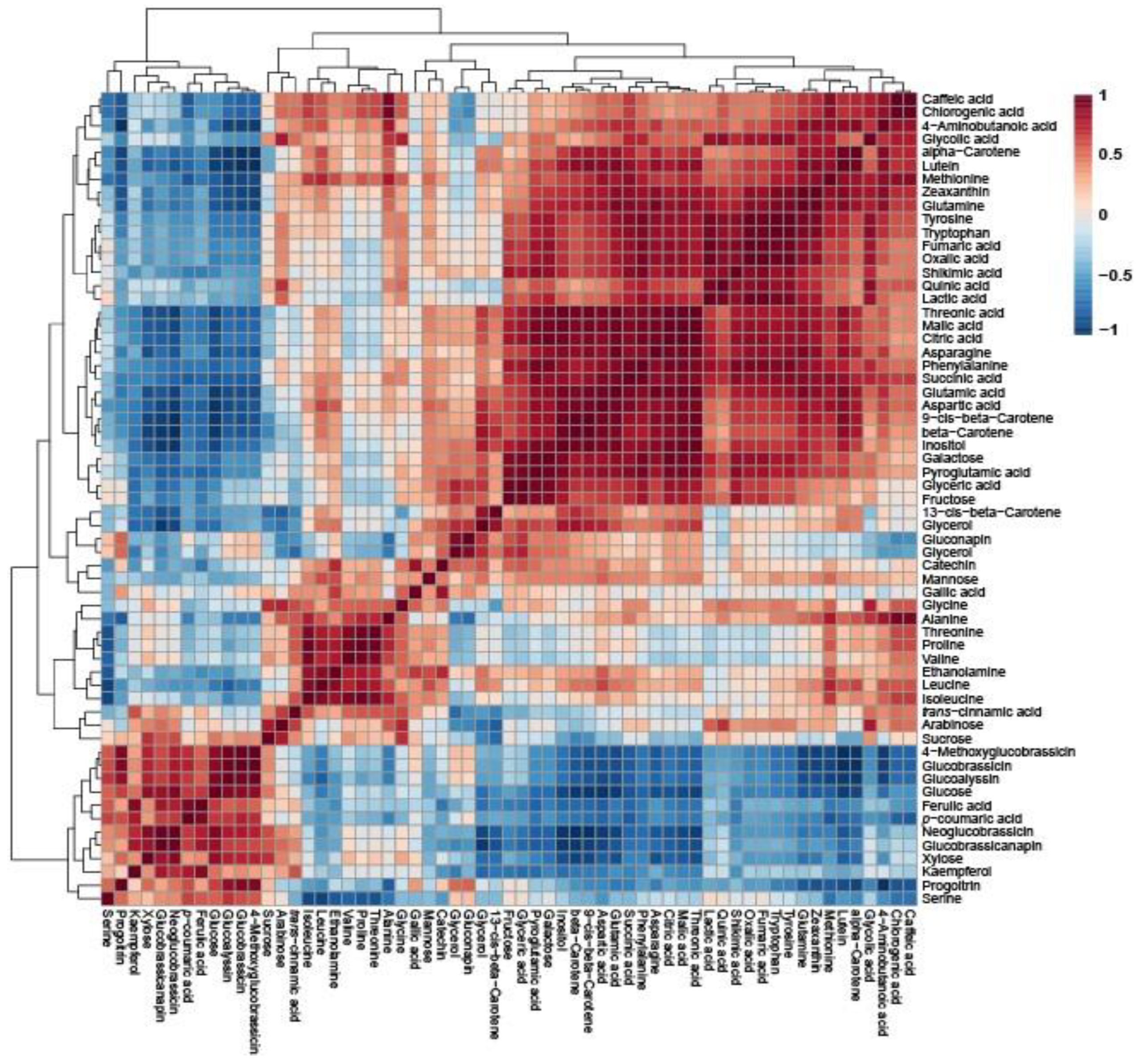
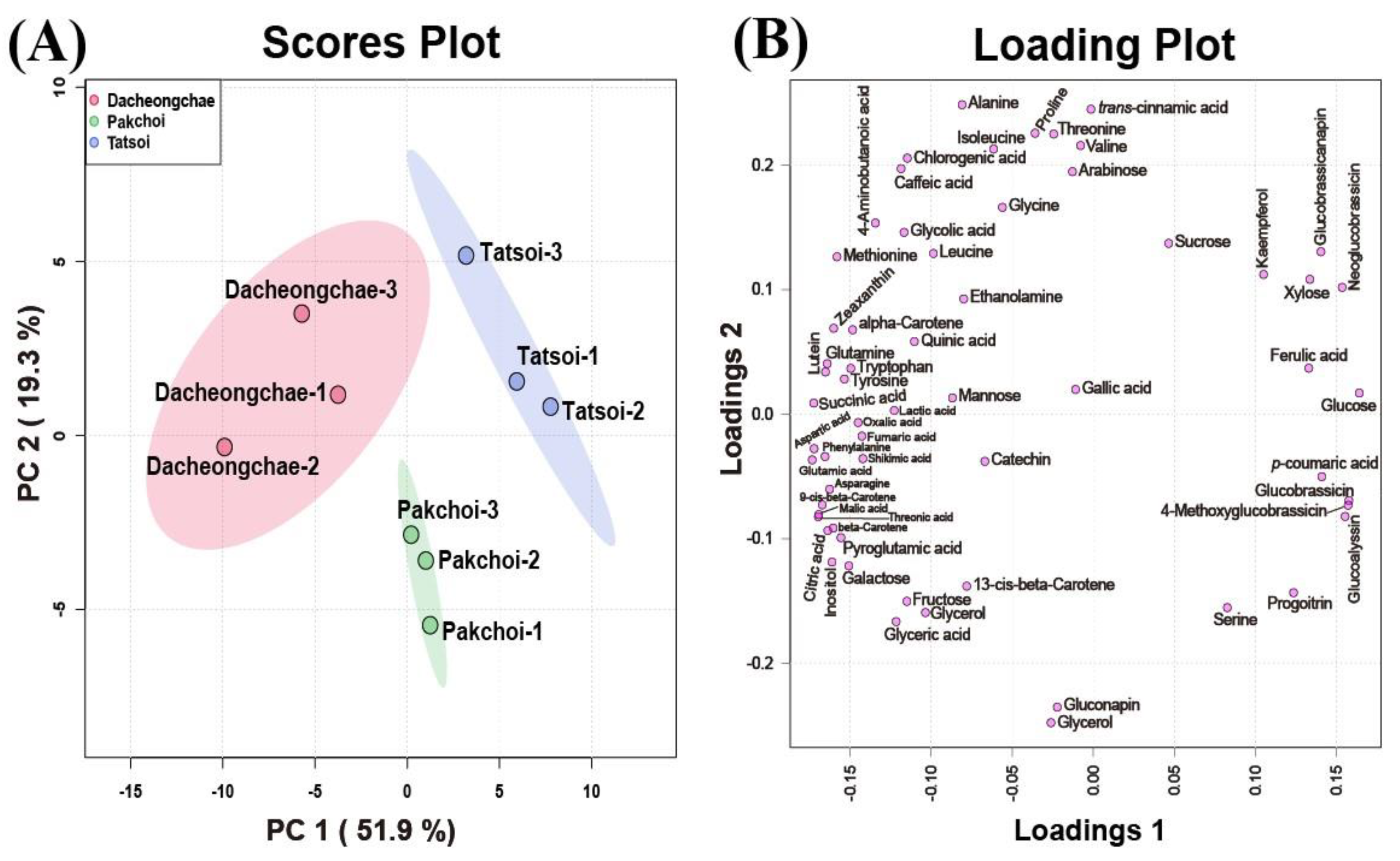
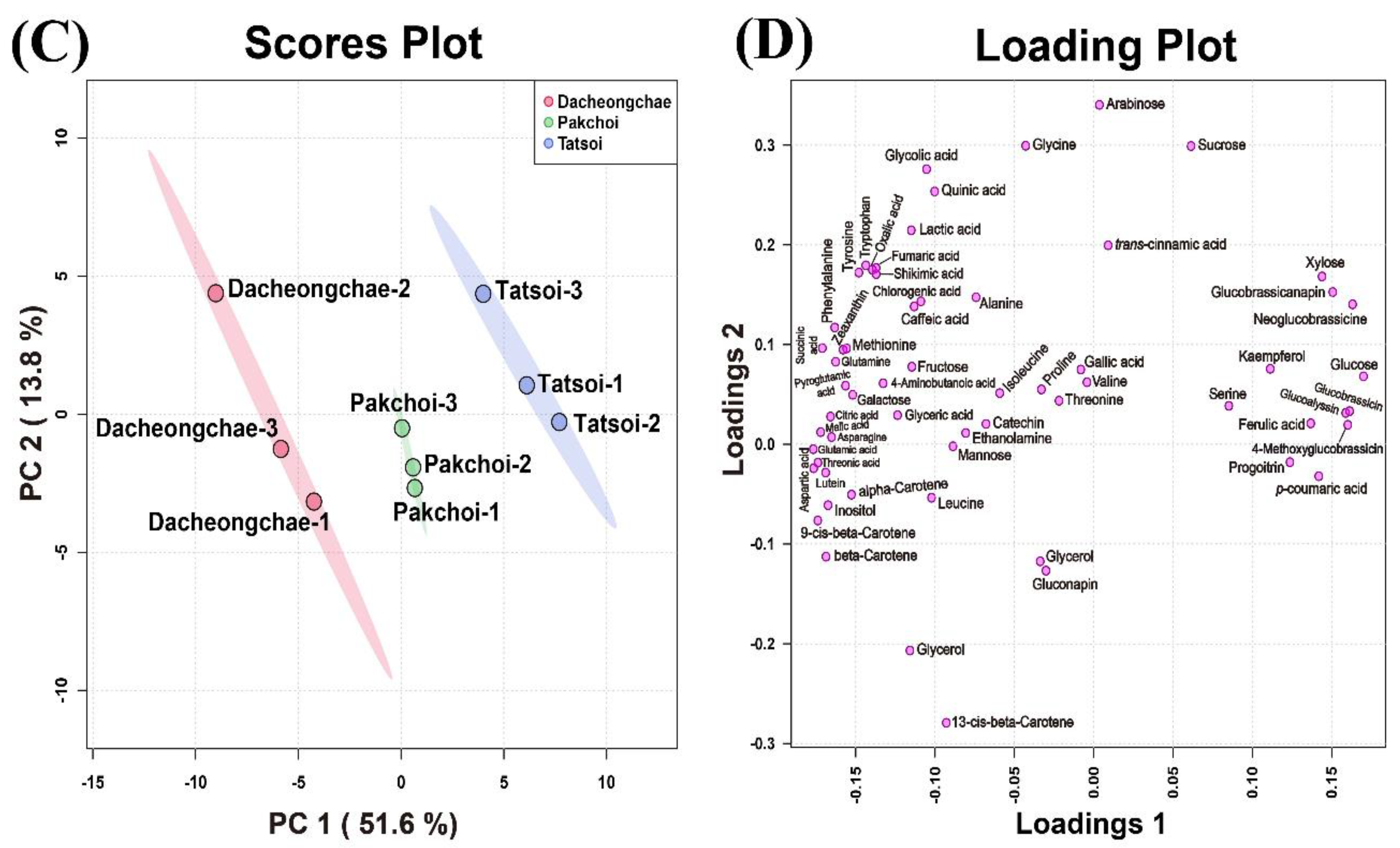
2.4. Metabolic Profiling of Tatsoi, Dacheongchae, and Pakchoi
2.5. In Vitro Antimicrobial Activities of Tatsoi, Dacheongchae, and Pakchoi
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Sample Preparation for HPLC Chemical Analysis
4.3. HPLC Analysis of Phenolics, Carotenoids, and Glucosinolates
4.4. Gas Chromatography-Time-of-Flight Mass Spectrometry (GC-TOFMS) Analysis
4.5. In Vitro Antimicrobial Properties Assay
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Artemyeva, A.; Solovyeva, A.; Vavilov, N. Quality evaluation of some cultivar types of leafy Brassica rapa. Acta Hortic. 2006, 706, 121. [Google Scholar] [CrossRef]
- Samuolienė, G.; Brazaitytė, A.; Jankauskienė, J.; Viršilė, A.; Sirtautas, R.; Novičkovas, A.; Sakalauskienė, S.; Sakalauskaitė, J.; Duchovskis, P. LED irradiance level affects growth and nutritional quality of Brassica microgreens. Cent. Eur. J. Biol. 2013, 8, 1241–1249. [Google Scholar] [CrossRef]
- Brazaitytė, A.; Sakalauskienė, S.; Samuolienė, G.; Jankauskienė, J.; Viršilė, A.; Novičkovas, A.; Sirtautas, R.; Miliauskienė, J.; Vaštakaitė, V.; Dabašinskas, L. The effects of LED illumination spectra and intensity on carotenoid content in Brassicaceae microgreens. Food Chem. 2015, 173, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Kyriacou, M.C.; El-Nakhel, C.; Graziani, G.; Pannico, A.; Soteriou, G.A.; Giordano, M.; Ritieni, A.; De Pascale, S.; Rou-phael, Y. Functional quality in novel food sources: Genotypic variation in the nutritive and phytochemical composition of thirteen microgreens species. Food Chem. 2019, 277, 107–118. [Google Scholar] [CrossRef]
- Kim, Y.B.; Li, X.; Kim, S.-J.; Kim, H.H.; Lee, J.; Kim, H.; Park, S.U. MYB transcription factors regulate glucosinolate biosynthesis in different organs of Chinese cabbage (Brassica rapa ssp. pekinensis). Molecules 2013, 18, 8682–8695. [Google Scholar] [CrossRef]
- Bischoff, K. Glucosinolates and organosulfur compounds. In Nutraceuticals in Veterinary Medicine; Springer International Publishing: Cham, Switzerland, 2019; pp. 113–119. [Google Scholar]
- Connolly, E.L.; Sim, M.; Travica, N.; Marx, W.; Beasy, G.; Lynch, G.S.; Bondonno, C.P.; Lewis, J.R.; Hodgson, J.M.; Blekkenhorst, L.C. Glucosinolates from cruciferous vegetables and their potential role in chronic disease: Investigating the preclinical and clinical evidence. Front. Pharmacol. 2021, 12, 767975. [Google Scholar] [CrossRef]
- Martínez-Cámara, S.; Ibañez, A.; Rubio, S.; Barreiro, C.; Barredo, J.-L. Main carotenoids produced by microorganisms. Encyclopedia 2021, 1, 1223–1245. [Google Scholar] [CrossRef]
- Park, C.H.; Park, S.-Y.; Park, Y.J.; Kim, J.K.; Park, S.U. Metabolite profiling and comparative analysis of secondary metabolites in Chinese cabbage, radish, and hybrid xBrassicoraphanus. J. Agric. Food Chem. 2020, 68, 13711–13719. [Google Scholar] [CrossRef]
- Björkman, M.; Klingen, I.; Birch, A.N.; Bones, A.M.; Bruce, T.J.; Johansen, T.J.; Meadow, R.; Mølmann, J.; Seljåsen, R.; Smart, L.E. Phytochemicals of Brassicaceae in plant protection and human health–Influences of climate, environment and agronomic practice. Phytochemistry 2011, 72, 538–556. [Google Scholar] [CrossRef]
- Gan, R.-Y.; Chan, C.-L.; Yang, Q.-Q.; Li, H.-B.; Zhang, D.; Ge, Y.-Y.; Gunaratne, A.; Ge, J.; Corke, H. Bioactive compounds and beneficial functions of sprouted grains. In Sprouted Grains; Elsevier: Amsterdam, The Netherlands, 2019; pp. 191–246. [Google Scholar]
- Hill, C.B.; Roessner, U. Metabolic profiling of plants by GC–MS. In The Handbook of Plant Metabolomics; Wiley: Hoboken, NJ, USA, 2013; pp. 1–23. [Google Scholar]
- Harrigan, G.G.; Goodacre, R. Metabolic Profiling: Its Role in Biomarker Discovery and Gene Function Analysis: Its Role in Biomarker Discovery and Gene Function Analysis; Springer Science & Business Media: Boston, NY, USA, 2003. [Google Scholar]
- Park, C.H.; Yeo, H.J.; Park, S.-Y.; Kim, J.K.; Park, S.U. Comparative phytochemical analyses and metabolic profiling of different phenotypes of Chinese cabbage (Brassica rapa ssp. pekinensis). Foods 2019, 8, 587. [Google Scholar] [CrossRef]
- Baek, S.-A.; Ahn, S.K.; Kim, K.W.; Choi, J.; Kim, J.; Ahn, J.; Ha, S.-H.; Park, S.U.; Kim, J.K. Metabolic profiling reveals glucose and fructose accumulation in gcr1 knock-out mutant of Arabidopsis. Appl. Biol. Chem. 2019, 62, 23. [Google Scholar] [CrossRef]
- Du, Z.; Lin, W.; Zhu, J.; Li, J. Amino acids profiling and transcriptomic data integration demonstrates the dynamic regulation of amino acids synthesis in the leaves of Cyclocarya paliurus. PeerJ 2022, 10, e13689. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Lee, H.W.; Li, Z.; Lu, Y.; Zou, L.; Ong, C.N. Simultaneous quantification of 22 glucosinolates in 12 Brassicaceae vegetables by hydrophilic interaction chromatography–tandem mass spectrometry. ACS Omega 2018, 3, 15546–15553. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.; Lim, C.J.; Kim, J.K.; Park, S.U. Comparative metabolic profiling of green and purple pakchoi (Brassica Rapa subsp. Chinensis). Molecules 2018, 23, 1613. [Google Scholar] [CrossRef]
- Yeo, H.J.; Baek, S.-A.; Sathasivam, R.; Kim, J.K.; Park, S.U. Metabolomic analysis reveals the interaction of primary and secondary metabolism in white, pale green, and green pak choi (Brassica rapa subsp. chinensis). Appl. Biol. Chem. 2021, 64, 3. [Google Scholar] [CrossRef]
- Pannico, A.; El-Nakhel, C.; Graziani, G.; Kyriacou, M.C.; Giordano, M.; Soteriou, G.A.; Zarrelli, A.; Ritieni, A.; De Pascale, S.; Rouphael, Y. Selenium biofortification impacts the nutritive value, polyphenolic content, and bioactive constitution of variable microgreens genotypes. Antioxidants 2020, 9, 272. [Google Scholar] [CrossRef]
- Saini, R.K.; Shang, X.M.; Ko, E.Y.; Choi, J.H.; Kim, D.; Keum, Y.-S. Characterization of nutritionally important phytoconstituents in minimally processed ready-to-eat baby-leaf vegetables using HPLC–DAD and GC–MS. J. Food Meas. Charact. 2016, 10, 341–349. [Google Scholar] [CrossRef]
- Sung, J.; Baek, S.; Kim, J.; Kim, Y.; Lee, Y.; Lee, S.; Lee, D.; Jung, H. Responses of primary metabolites and glucosinolates in sulfur deficient-cabbage (Brassica rapa L. ssp. pekinensis). J. Plant Biochem. Physiol. 2018, 6, 223. [Google Scholar] [CrossRef]
- Kirschning, A. The coenzyme/protein pair and the molecular evolution of life. Nat. Prod. Rep. 2021, 38, 993–1010. [Google Scholar] [CrossRef]
- Okumoto, S.; Funck, D.; Trovato, M.; Forlani, G. Amino acids of the glutamate family: Functions beyond primary metabolism. Front. Plant Sci. 2016, 7, 318. [Google Scholar] [CrossRef]
- Tohge, T.; Watanabe, M.; Hoefgen, R.; Fernie, A.R. Shikimate and phenylalanine biosynthesis in the green lineage. Front. Plant Sci. 2013, 4, 62. [Google Scholar] [CrossRef] [PubMed]
- Mayes, P.A.; Bender, D.A. The citric acid cycle: The catabolism of AcetylCoA. In Harper’s illustrated Biochemistry; Murray, R.K., Granner, D.K., Mayes, P.A., Rodwell, V., Eds.; McGraw-Hill: New York, NY, USA, 2003; pp. 130–135. [Google Scholar]
- Qu, Q.; Jin, L. Application of nuclear magnetic resonance in food analysis. Food Sci. Technol. 2022, 42, e43622. [Google Scholar] [CrossRef]
- Abdel-Farid, I.B.; Kim, H.K.; Choi, Y.H.; Verpoorte, R. Metabolic characterization of Brassica rapa leaves by NMR spectroscopy. J. Agric. Food Chem. 2007, 55, 7936–7943. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.S.; Choi, Y.H.; Kim, H.K.; Linthorst, H.J.; Verpoorte, R. Metabolomic analysis of methyl jasmonate treated Brassica rapa leaves by 2-dimensional NMR spectroscopy. Phytochemistry 2006, 67, 2503–2511. [Google Scholar] [CrossRef]
- Kim, J.; Jung, Y.; Song, B.; Bong, Y.S.; Lee, K.S.; Hwang, G.S. Discrimination of cabbage (Brassica rapa ssp. pekinensis) cultivars grown in different geographical areas using 1H NMR-based metabolomics. Food Chem. 2013, 137, 68–75. [Google Scholar] [CrossRef]
- Rubab, M.; Chellia, R.; Saravanakumar, K.; Mandava, S.; Khan, I.; Tango, C.N.; Hussain, M.S.; Daliri, E.B.-M.; Kim, S.-H.; Ramakrishnan, S.R. Preservative effect of Chinese cabbage (Brassica rapa subsp. pekinensis) extract on their molecular docking, antioxidant and antimicrobial properties. PLoS ONE 2018, 13, e0203306. [Google Scholar] [CrossRef]
- Yeo, H.J.; Ki, W.Y.; Lee, S.; Kim, C.Y.; Kim, J.K.; Park, S.U.; Park, C.H. Metabolite profiles and biological activities of different phenotypes of Chinese cabbage (Brassica rapa ssp. Pekinensis). Food Res. Int. 2023, 174, 113619. [Google Scholar] [CrossRef]
- Carlo Tenore, G.; Troisi, J.; Di Fiore, R.; Basile, A.; Novellino, E. Chemical composition, antioxidant and antimicrobial properties of Rapa Catozza Napoletana (Brassica rapa L. var. rapa DC.) seed meal, a promising protein source of Campania region (southern Italy) horticultural germplasm. J. Sci. Food Agric. 2012, 92, 1716–1724. [Google Scholar] [CrossRef]
- Varghese, P. Red cabbage methanol extract inhibits the growth of vancomycin-resistant Enterococcus faecalis during kirby-bauer disk diffusion susceptibility test. J. Stud. Res. 2015, 4, 87–89. [Google Scholar] [CrossRef]
- Pacheco-Cano, R.; Salcedo-Hernández, R.; López-Meza, J.; Bideshi, D.; Barboza-Corona, J. Antimicrobial activity of broccoli (Brassica oleracea var. italica) cultivar Avenger against pathogenic bacteria, phytopathogenic filamentous fungi and yeast. J. Appl. Microbiol. 2018, 124, 126–135. [Google Scholar] [CrossRef]
- Arrais, A.; Testori, F.; Calligari, R.; Gianotti, V.; Roncoli, M.; Caramaschi, A.; Todeschini, V.; Massa, N.; Bona, E. Extracts from Cabbage Leaves: Preliminary Results towards a “Universal” Highly-Performant Antibacterial and Antifungal Natural Mixture. Biology 2022, 11, 1080. [Google Scholar] [CrossRef] [PubMed]
- Andrés, C.M.C.; Pérez de la Lastra, J.M.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. Polyphenols as antioxidant/pro-oxidant compounds and donors of reducing species: Relationship with human antioxidant metabolism. Processes 2023, 11, 2771. [Google Scholar] [CrossRef]
- Abou Elmaaty, T.; Sayed-Ahmed, K.; Elsisi, H.; Magdi, M. Optimization of extraction of natural antimicrobial pigments using supercritical fluids: A review. Processes 2022, 10, 2111. [Google Scholar] [CrossRef]
- Gantait, A.; Masih, S.A.; Addesso, R.; Maxton, A.; Sofo, A. Glucosinolates Mediated Regulation of Enzymatic Activity in Response to Oxidative Stress in Brassica spp. Plants 2024, 13, 3422. [Google Scholar] [CrossRef] [PubMed]
- Lobiuc, A.; Pavăl, N.E.; Mangalagiu, I.I.; Gheorghiță, R.; Teliban, G.C.; Amăriucăi-Mantu, D.; Stoller, V. Future antimicrobials: Natural and functionalized phenolics. Molecules 2023, 28, 1114. [Google Scholar] [CrossRef]
- Hoch, C.C.; Shoykhet, M.; Weiser, T.; Griesbaum, L.; Petry, J.; Hachani, K.; Multhoff, G.; Dezfouli, A.B.; Wollenberg, B. Isothiocyanates in medicine: A comprehensive review on phenylethyl-, allyl-, and benzyl-isothiocyanates. Pharmacol. Res. 2024, 201, 107107. [Google Scholar] [CrossRef]
- Karpiński, T.M.; Ożarowski, M.; Alam, R.; Łochyńska, M.; Stasiewicz, M. What do we know about antimicrobial activity of astaxanthin and fucoxanthin? Mar. Drugs 2021, 20, 36. [Google Scholar] [CrossRef]
- Lee, S.; Park, C.H.; Kim, J.K.; Ahn, K.; Kwon, H.; Kim, J.K.; Park, S.U.; Yeo, H.J. LED lights influenced phytochemical contents and biological activities in kale (Brassica oleracea L. var. acephala) microgreens. Antioxidants 2023, 12, 1686. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds (μg g−1 dw) | Tatsoi | Dacheongchae | Pakchoi |
|---|---|---|---|
| Lutein | 259.02 ± 25.06 c | 668.71 ± 18.28 a | 352.77 ± 38.37 b1 |
| Zeaxantin | N.D. | 4.29 ± 1.54 a | data N.D.2 |
| 13Z-β-Carotene | 32.78 ± 4.65 b | 61.53 ± 17.32 a | 64.29 ± 9.37 a |
| α-Carotene | 7.13 ± 0.28 b | 13.98 ± 0.92 a | 7.95 ± 1.44 b |
| β-Carotene | 298.14 ± 8.75 c | 683.42 ± 19.08 a | 554.71 ± 20.00 b |
| 9Ζ-β-Carotene | 34.77 ± 3.94 c | 104.61 ± 8.33 a | 76.76 ± 2.18 b |
| Total | 631.85 ± 30.80 c | 1536.54 ± 31.18 a | 1056.48 ± 58.78 b |
| Compounds (μmol g−1 dw) | Tatsoi | Dacheongchae | Pakchoi |
|---|---|---|---|
| Progoitrin | 0.47 ± 0.04 c | 0.77 ± 0.02 b | 0.88 ± 0.07 a1 |
| Glucoalyssin | 0.34 ± 0.03 b | 0.58 ± 0.03 a | 0.54 ± 0.04 a |
| Gluconapin | 3.87 ± 0.36 b | 3.59 ± 0.22 b | 4.82 ± 0.29 a |
| Glucobrassicanapin | 1.18 ± 0.06 b | 2.32 ± 0.12 a | 1.26 ± 0.05 b |
| Glucobrassicin | 0.08 ± 0.02 b | 0.42 ± 0.03 a | 0.37 ± 0.03 a |
| 4-Methoxyglucobrassicin | 0.05 ± 0.01 c | 0.84 ± 0.03 a | 0.77 ± 0.03 b |
| Neoglucobrassicin | 0.05 ± 0.01 b | 0.39 ± 0.01 a | 0.14 ± 0.01 b |
| Total | 6.04 ± 0.43 b | 8.91 ± 0.40 a | 8.79 ± 0.44 a |
| Compounds (μg g−1 dw) | Tatsoi | Dacheongchae | Pakchoi |
|---|---|---|---|
| Gallic acid | 16.93 ± 0.21 a | 16.91 ± 0.22 a | 17.07 ± 0.39 a1 |
| Catechin | 661.76 ± 41.62 a | 709.72 ± 32.31 a | 738.99 ± 41.67 a |
| Chlorogenic acid | 116.49 ± 0.78 b | 199.51 ± 2.09 a | 107.56 ± 1.13 c |
| Caffeic acid | 36.02 ± 0.41 b | 105.05 ± 1.82 a | 33.92 ± 0.19 b |
| p-coumaric acid | 7.11 ± 0.18 a | 3.14 ± 0.19 c | 4.78 ± 0.52 b |
| Ferulic acid | 7.24 ± 0.53 a | 1.89 ± 0.32 b | 2.50 ± 0.15 b |
| trans-cinnamic acid | 1.46 ± 0.13 a | 1.37 ± 0.09 ab | 1.18 ± 0.15 b |
| Kaempferol | 79.38 ± 0.55 a | 77.90 ± 0.10 b | 77.72 ± 0.36 b |
| Total | 882.33 ± 43.89 c | 1115.49 ± 37.14 a | 983.72 ± 44.56 b |
| Group | Bacterial Strains | Zone of Inhibition (mm) | ||
|---|---|---|---|---|
| Tatsoi | Dacheongchae | Pakchoi | ||
| Pathogens | Vibrio parahaemolyticus (KCTC 2471) | — | — | — 1 |
| Streptococcus mutans (KCTC 3065) | — | — | — | |
| Staphylococcus aureus (KCTC 3881) | — | — | — | |
| Micrococcus luteus (KCTC 3063) | — | — | — | |
| Bacillus cereus (KCTC 3624) | 11.5–12 | 11.5–12 | 10.5–11 | |
| Escherichia coli (KCTC 1682) | 9.5–10 | 12–12.5 | 12.5–13 | |
| Proteus mirabilis (KCTC 2510) | 13–14 | 11–12 | 10–10.5 | |
| Salmonella paratyphi C (KCCM 41577) | — | 11–12 | — | |
| Pseudomonas aeruginosa (KCCM 11803) | 11.5–12 | 15–16 | 12–13 | |
| Pathogenic yeast | Candida albicans (ATCC 28367) | 11.5–12 | 11.5–12.5 | 11–11.5 |
| Multidrug-resistant pathogens | Pseudomonas aeruginosa (0225) | 11–11.5 | 11–11.5 | 9.5–10 |
| Pseudomonas aeruginosa (0254) | 12–12.5 | 14–15 | 11.5–12 | |
| Pseudomonas aeruginosa (0826) | 10–10.5 | 9–10 | 10.5–11 | |
| Pseudomonas aeruginosa (1113) | 12–13 | 12–13 | 10–11 | |
| Pseudomonas aeruginosa (1378) | 11–12 | 12–13 | 11–11.5 | |
| Pseudomonas aeruginosa (1731) | 10.5–11 | 12–13 | 10–10.5 | |
| Pseudomonas aeruginosa (P01827) | 11–12 | 11–12 | 9–10 | |
| Pseudomonas aeruginosa (P01828) | 9–9.5 | 13–14 | 9.5–10 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, C.-H.; Yeo, H.-J.; Park, Y.-J.; Kwon, H.; Cho, J.; Chung, S.-O.; Lee, G.-J.; Kim, J.-K.; Park, S.-U. Metabolite Profiling and Antimicrobial Activities of Brassica rapa ssp. narinosa (Tatsoi), Brassica rapa var. narinosa × chinensis (Dacheongchae), and Brassica rapa ssp. chinensis (Pakchoi). Molecules 2025, 30, 1693. https://doi.org/10.3390/molecules30081693
Park C-H, Yeo H-J, Park Y-J, Kwon H, Cho J, Chung S-O, Lee G-J, Kim J-K, Park S-U. Metabolite Profiling and Antimicrobial Activities of Brassica rapa ssp. narinosa (Tatsoi), Brassica rapa var. narinosa × chinensis (Dacheongchae), and Brassica rapa ssp. chinensis (Pakchoi). Molecules. 2025; 30(8):1693. https://doi.org/10.3390/molecules30081693
Chicago/Turabian StylePark, Chang-Ha, Hyeon-Ji Yeo, Young-Jin Park, Haejin Kwon, Jongki Cho, Sun-Ok Chung, Geung-Joo Lee, Jae-Kwang Kim, and Sang-Un Park. 2025. "Metabolite Profiling and Antimicrobial Activities of Brassica rapa ssp. narinosa (Tatsoi), Brassica rapa var. narinosa × chinensis (Dacheongchae), and Brassica rapa ssp. chinensis (Pakchoi)" Molecules 30, no. 8: 1693. https://doi.org/10.3390/molecules30081693
APA StylePark, C.-H., Yeo, H.-J., Park, Y.-J., Kwon, H., Cho, J., Chung, S.-O., Lee, G.-J., Kim, J.-K., & Park, S.-U. (2025). Metabolite Profiling and Antimicrobial Activities of Brassica rapa ssp. narinosa (Tatsoi), Brassica rapa var. narinosa × chinensis (Dacheongchae), and Brassica rapa ssp. chinensis (Pakchoi). Molecules, 30(8), 1693. https://doi.org/10.3390/molecules30081693