
Profiling and Bioactivity of Polyphenols from the Omani Medicinal Plant Terminalia dhofarica (syn. Anogeissus dhofarica)


Abstract

1. Introduction
2. Results and Discussion
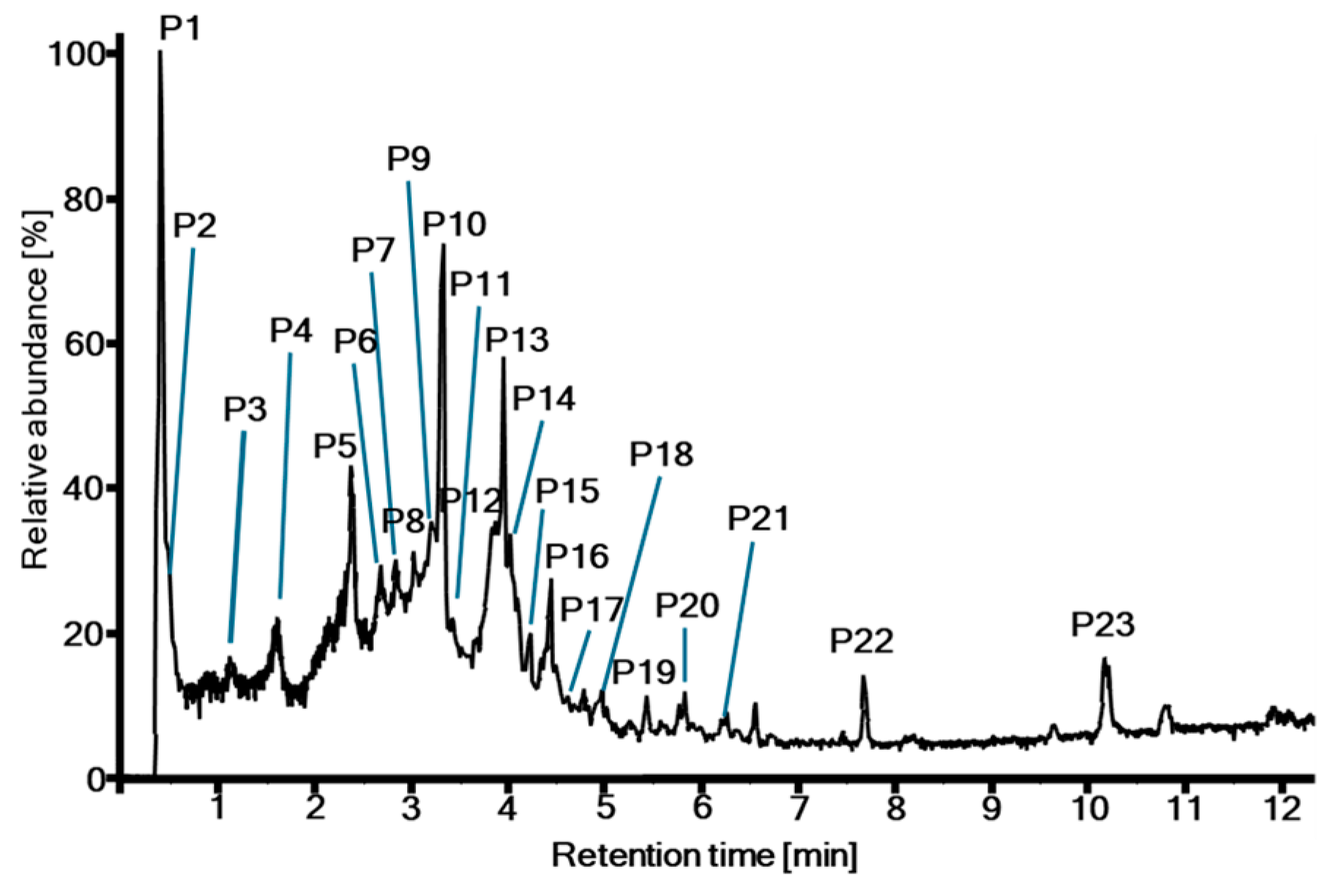
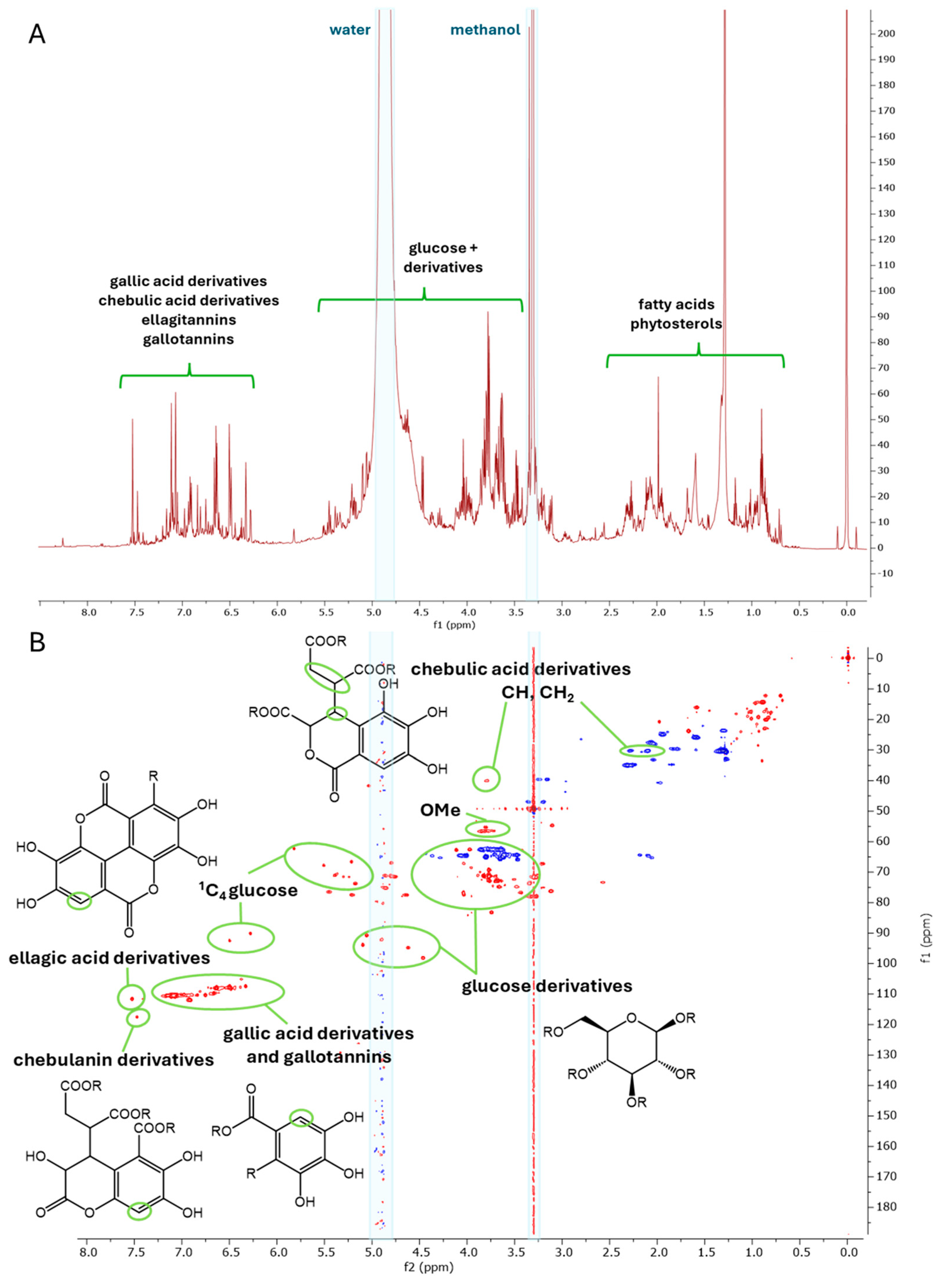
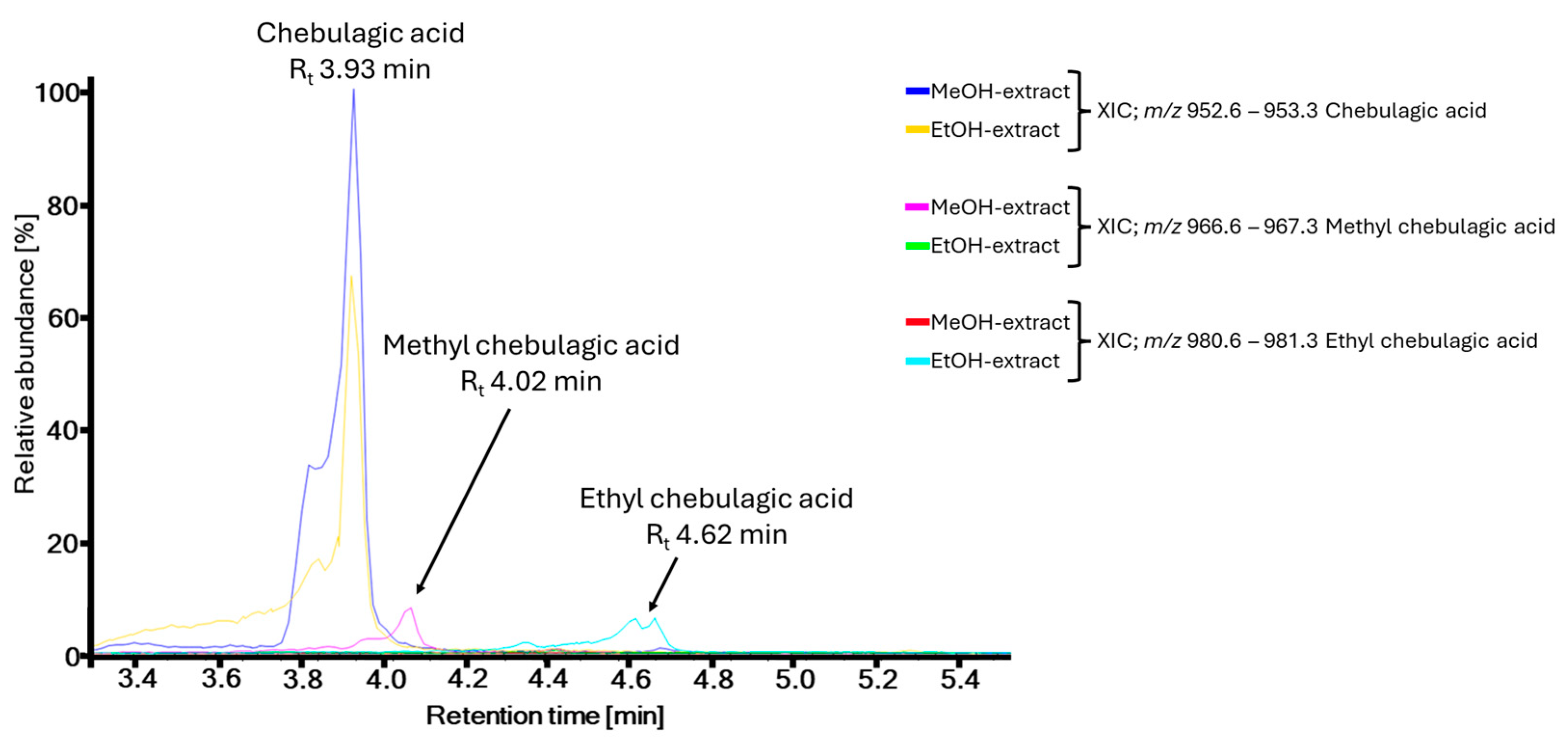
2.1. Metabolite Profiling
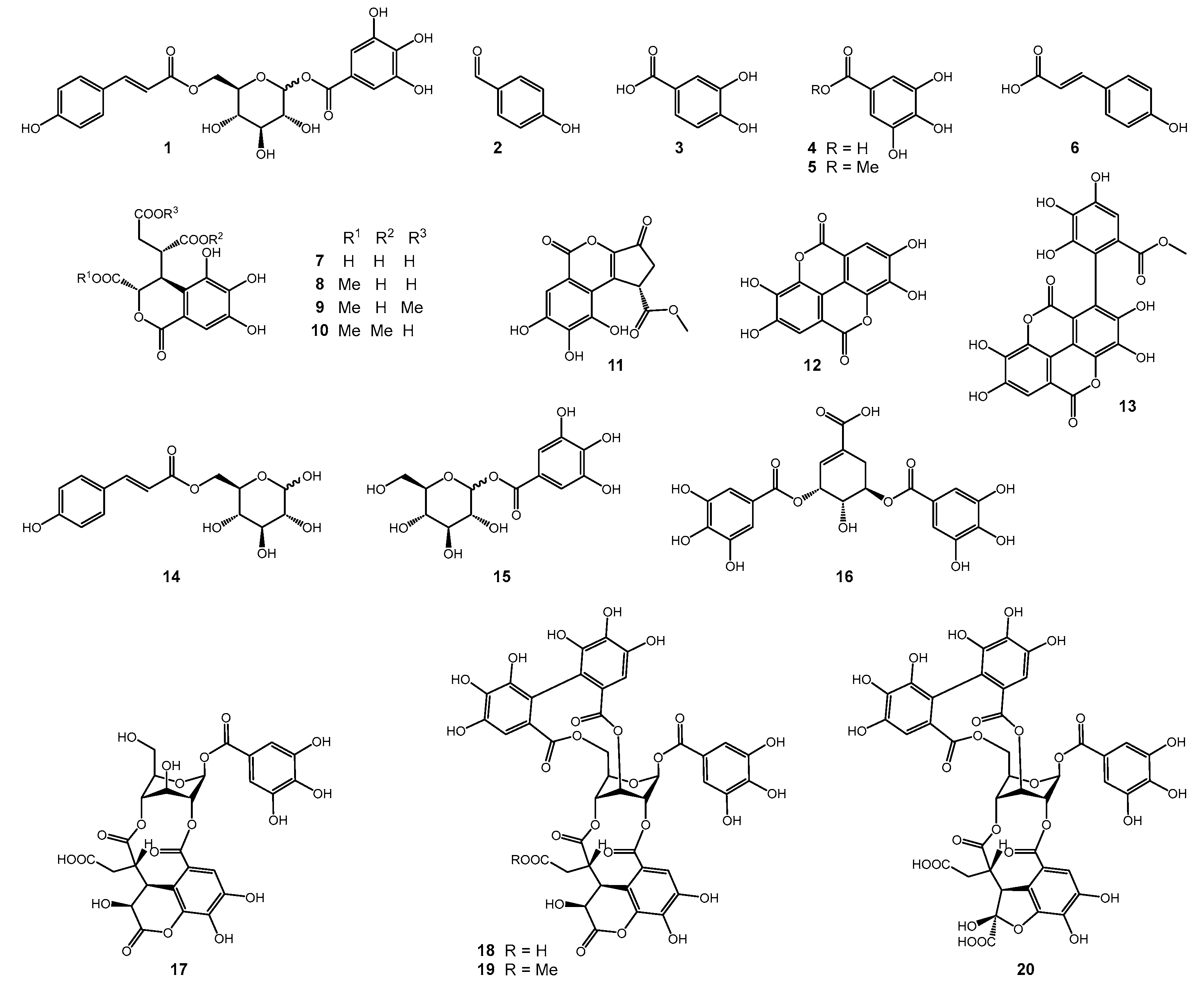
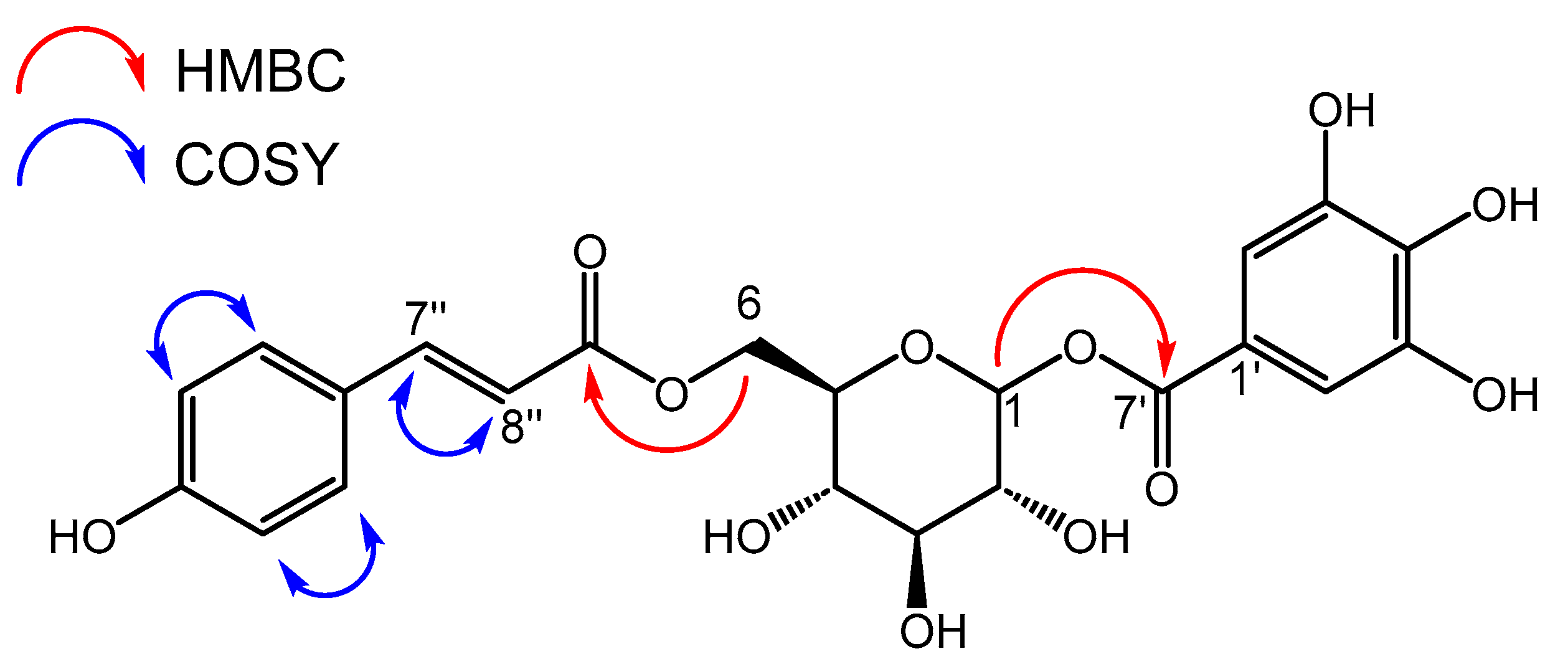
2.2. Isolation and Structure Elucidation
2.3. Evaluation of Artifacts
2.4. Evaluation of Bioactivity
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Analytical Instruments and General Procedures
3.3. Plant Material
3.4. Isolation
3.5. Biological Assays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kürschner, H.; Hein, P.; Kilian, N.; Hubaishan, M.A. The Hybantho durae-Anogeissetum dhofaricae ass. Nova-phytosociology, structure and ecology of an endemic South Arabian forest community. Phytocoenologia 2004, 34, 569–612. [Google Scholar] [CrossRef]
- Maurin, O.; Gere, J.; van der Bank, M.; Boatwright, J.S. The inclusion of Anogeissus, Buchenavia and Pteleopsis in Terminalia (Combretaceae: Terminaliinae). Bot. J. Linn. 2017, 184, 312–325. [Google Scholar] [CrossRef]
- IPNI. Anogeissus dhofarica. Available online: https://www.ipni.org/n/169727-1 (accessed on 8 November 2024).
- IPNI. Terminalia dhofarica. Available online: https://www.ipni.org/n/77164562-1 (accessed on 8 November 2024).
- Scott, A.J. A revision of Anogeissus (Combretaceae). Kew Bull. 1979, 33, 555. [Google Scholar] [CrossRef]
- Jain, A.; Katewa, S.S.; Galav, P.K.; Sharma, P. Medicinal plant diversity of Sitamata wildlife sanctuary, Rajasthan, India. J. Ethnopharmacol. 2005, 102, 143–157. [Google Scholar] [CrossRef]
- Meena, K.L.; Yadav, B.L. Studies on ethnomedicinal plants conserved by Garasia tribes of Sirohi district, Rajasthan, India. Indian J. Nat. Prod. Resour. 2010, 1, 500–506. [Google Scholar]
- Marwah, R.G.; Fatope, M.O.; Mahrooqi, R.A.; Varma, G.B.; Abadi, H.A.; Al-Burtamani, S.K.S. Antioxidant capacity of some edible and wound healing plants in Oman. Food Chem. 2007, 101, 465–470. [Google Scholar] [CrossRef]
- Manosroi, J.; Moses, Z.Z.; Manosroi, W.; Manosroi, A. Hypoglycemic activity of Thai medicinal plants selected from the Thai/Lanna Medicinal Recipe Database MANOSROI II. J. Ethnopharmacol. 2011, 138, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.; Baghel, U.S.; Gautam, A.; Baghel, D.S.; Yadav, D.; Malik, J.; Yadav, R. The genus Anogeissus: A review on ethnopharmacology, phytochemistry and pharmacology. J. Ethnopharmacol. 2016, 194, 30–56. [Google Scholar] [CrossRef] [PubMed]
- Barthélemy, A.; Latifou, L.; Dodehe, Y.; Cyril, A.; Catherine, V.-S. In vitro antiplasmodial and antileishmanial activities of flavonoids from Anogeissus leiocarpus (Combretaceae). Int. J. Pharm. Sci. Rev. Res. 2011, 11, 1–6. [Google Scholar]
- Miller, A.G.; Morris, M. Plants of Dhofar: The Southern Region of Oman; Traditional, Economic and Medicinal Uses; The Office of The Adviser for Conservation of The Environment: Muscat, Oman, 1988; ISBN 0715708082. [Google Scholar]
- Al-Noumani, A.J.; Al-Qasmi, M.Z.J.; Al-Shabibi, A.S.A.; Al-Mashani, S.A.I.; Said, S.A. Antimicrobial, antioxidant and cytotoxic activities of Anogeissus dhofarica from Oman. Int. J. Recent Adv. Pharm. Res. 2013, 3, 35–38. [Google Scholar]
- Maqsood, R.; Khan, F.; Ullah, S.; Khan, A.; Al-Jahdhami, H.; Hussain, J.; Weli, M.; Maqsood, D.; Rahman, S.M.; Hussain, A.; et al. Evaluation of antiproliferative, antimicrobial, antioxidant, antidiabetic and phytochemical analysis of Anogeissus dhofarica A. J. Scott. Antibiotics 2023, 12, 354. [Google Scholar] [CrossRef]
- Abuarqoub, D.; Aburayyan, W.; Rashan, L.; Dayyih, W.A.; Al-Matubsi, H.Y. Phytochemical analysis and in vitro investigation of wound healing, cytotoxicity, and inflammatory response in potentially active extracts of Anogeissus dhofarica. J. Appl. Pharm. Sci. 2024, 14, 152–162. [Google Scholar] [CrossRef]
- Mei, Y.; Hu, Y.; Tao, X.; Shang, J.; Qian, M.; Suo, F.; Li, J.; Cao, L.; Wang, Z.; Xiao, W. Chemical profiling of Shen-Wu-Yi-Shen tablets using UPLC-Q-TOF-MS/MS and its quality evaluation based on UPLC-DAD combined with multivariate statistical analysis. J. Chromatogr. Sci. 2024, 62, 534–553. [Google Scholar] [CrossRef] [PubMed]
- Kuivila, H.G.; Nahabedian, K.V. Electrophilic displacement reactions. X. General acid catalysis in the protodeboronation of areneboronic acids 1–3. J. Am. Chem. Soc. 1961, 83, 2159–2163. [Google Scholar] [CrossRef]
- Yan, P.; Zeng, R.; Bao, B.; Yang, X.-M.; Zhu, L.; Pan, B.; Niu, S.-L.; Qi, X.-W.; Li, Y.-L.; Ouyang, Q. Red light-induced highly efficient aerobic oxidation of organoboron compounds using spinach as a photocatalyst. Green Chem. 2022, 24, 9263–9268. [Google Scholar] [CrossRef]
- Meng, Q.; Li, G.; Luo, B.; Wang, L.; Lu, Y.; Liu, W. Screening and isolation of natural antioxidants from Ziziphora clinopodioides Lam. with high performance liquid chromatography coupled to a post-column Ce(IV) reduction capacity assay. RSC Adv. 2016, 6, 62378–62384. [Google Scholar] [CrossRef]
- Zaher, A.M.; Anwar, W.S.; Makboul, M.A.; Abdel-Rahman, I.A.M. Potent anticancer activity of (Z)-3-hexenyl-β-D-glucopyranoside in pancreatic cancer cells. Naunyn Schmiedebergs Arch. Pharmacol. 2024, 397, 2311–2320. [Google Scholar] [CrossRef]
- Xu, K.-Z.; Xiang, S.-L.; Wang, Y.-J.; Wang, B.; Jia, A.-Q. Methyl gallate isolated from partridge tea (Mallotus oblongifolius (Miq.) Müll.Arg.) inhibits the biofilms and virulence factors of Burkholderia thailandensis. J. Ethnopharmacol. 2024, 320, 117422. [Google Scholar] [CrossRef] [PubMed]
- Terfassi, S.; Dauvergne, X.; Cérantola, S.; Lemoine, C.; Bensouici, C.; Fadila, B.; Christian, M.; Marchioni, E.; Benayache, S. First report on phytochemical investigation, antioxidant and antidiabetic activities of Helianthemum getulum. Nat. Prod. Res. 2022, 36, 2806–2813. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.-R.; Kuo, Y.-H.; Ho, Y.-L.; Wang, C.-Y.; Yang, C.-S.; Lin, C.-W.; Chang, Y.-S. Studies on cytotoxic constituents from the leaves of Elaeagnus oldhamii Maxim. in non-small cell lung cancer A549 cells. Molecules 2014, 19, 9515–9534. [Google Scholar] [CrossRef]
- Lee, H.-S.; Jung, S.-H.; Yun, B.-S.; Lee, K.-W. Isolation of chebulic acid from Terminalia chebula Retz. and its antioxidant effect in isolated rat hepatocytes. Arch. Toxicol. 2007, 81, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Kim, H.W.; Yang, H.; Sung, S.H. Hydrolyzable tannins from the fruits of Terminalia chebula Retz and their α-glucosidase inhibitory activities. Phytochemistry 2017, 137, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.-Y.; Lan, Q.; He, S.; Su, B.-J.; Wang, Y.-Q.; Liao, H.-B.; Wang, H.-S.; Liang, D. Chebulic acid derivatives from Balakata baccata and their antineuroinflammatory and antioxidant activities. Bioorg. Chem. 2021, 116, 105332. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Yang, L.; Zhai, L.; Huang, Y.; Chen, F.; Duan, W.; Yang, J. Methyl brevifolincarboxylate, a novel influenza virus PB2 inhibitor from Canarium album (Lour.) Raeusch. Chem. Biol. Drug Des. 2020, 96, 1280–1291. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-N.; Su, B.-J.; Wang, Y.-Q.; Liao, H.-B.; Chen, Z.-F.; Liang, D. Isolation, absolute configuration, and biological activities of chebulic acid and brevifolincarboxylic acid derivatives from Euphorbia hirta. J. Nat. Prod. 2020, 83, 985–995. [Google Scholar] [CrossRef]
- Navarro, F.; Hamri, S.; Reches, R.; Viñas, M.; Jahani, D.; Ginard, J.; Vilardell, J.; Abián, O.; Pujol, M.D. Convenient synthesis of ellagic acid from methyl gallate and SARS-CoV-2 3CLpro antiviral activity. Synthesis 2023, 55, 657–662. [Google Scholar] [CrossRef]
- Srivastava, S.; Mishra, D.; Bisht, R.; Savita, K.; Singh, K.; Rani, P.; Chanda, D.; Dev, K. Psiguanol, a novel α-pyrone derivative from Psidium guajava leaves and vasorelaxant activity in rat aorta cells through intracellular cGMP-dependent opening of calcium-activated potassium channels. Nat. Prod. Res. 2024, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Marzouk, M.S.A.; El-Toumy, S.A.A.; Moharram, F.A.; Shalaby, N.M.M.; Ahmed, A.A.E. Pharmacologically active ellagitannins from Terminalia myriocarpa. Planta Med. 2002, 68, 523–527. [Google Scholar] [CrossRef]
- Shimomura, H.; Sashida, Y.; Adachi, T. Phenylpropanoid glucose esters from Prunus buergeriana. Phytochemistry 1988, 27, 641–644. [Google Scholar] [CrossRef]
- Ni, J.-C.; Shi, J.-T.; Tan, Q.-W.; Chen, Q.-J. Phenylpropionamides, piperidine, and phenolic derivatives from the fruit of Ailanthus altissima. Molecules 2017, 22, 2107. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.-J.; Wu, M.-D.; Khamthong, N.; Tseng, M. Polar metabolites from the actinobacterium Isoptericola chiayiensis isolated from mangrove soil. Chem. Nat. Compd. 2021, 57, 1134–1136. [Google Scholar] [CrossRef]
- Pfundstein, B.; El Desouky, S.K.; Hull, W.E.; Haubner, R.; Erben, G.; Owen, R.W. Polyphenolic compounds in the fruits of Egyptian medicinal plants (Terminalia bellerica, Terminalia chebula and Terminalia horrida): Characterization, quantitation and determination of antioxidant capacities. Phytochemistry 2010, 71, 1132–1148. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Abe, T.; Tanaka, T.; Yang, C.R.; Kouno, I. Phyllanemblinins A–F, new ellagitannins from Phyllanthus emblica. J. Nat. Prod. 2001, 64, 1527–1532. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, Y.-N.; Guo, L.-B.; He, J.; Liu, P.-H.; Tian, H.-Y.; Zhang, W.-K.; Xu, J.-K. Diverse gallotannins with α-glucosidase and α-amylase inhibitory activity from the roots of Euphorbia fischeriana Steud. Phytochemistry 2022, 202, 113304. [Google Scholar] [CrossRef]
- Lee, D.Y.; Yang, H.; Kim, H.W.; Sung, S.H. New polyhydroxytriterpenoid derivatives from fruits of Terminalia chebula Retz. and their α-glucosidase and α-amylase inhibitory activity. Bioorg. Med. Chem. Lett. 2017, 27, 34–39. [Google Scholar] [CrossRef]
- Kappen, J.; Manurung, J.; Fuchs, T.; Vemulapalli, S.P.B.; Schmitz, L.M.; Frolov, A.; Agusta, A.; Muellner-Riehl, A.N.; Griesinger, C.; Franke, K.; et al. Challenging structure elucidation of lumnitzeralactone, an ellagic acid derivative from the mangrove Lumnitzera racemosa. Mar. Drugs 2023, 21, 242. [Google Scholar] [CrossRef] [PubMed]
- Ware, I.; Franke, K.; Dube, M.; El Enshasy, H.A.; Wessjohann, L.A. Characterization and bioactive potential of secondary metabolites isolated from Piper sarmentosum Roxb. Int. J. Mol. Sci. 2023, 24, 1328. [Google Scholar] [CrossRef]
- Utpal, B.K.; Sutradhar, B.; Zehravi, M.; Sweilam, S.H.; Panigraphy, U.P.; Urs, D.; Fatima, A.F.; Nallasivan, P.K.; Chabra, G.S.; Sayeed, M.; et al. Polyphenols in wound healing: Unlocking prospects with clinical applications. Naunyn Schmiedeberg’s Arch. Pharmacol. 2024, 1–27. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak No. * | tR (min) | m/z [M − H]− Measured | Error (ppm) | Molecular Formula | MS2 Product Ions m/z (rel. Intensity [%]) | Annotation |
|---|---|---|---|---|---|---|
| P1 | 0.38 | 169.0151 | 5.1 | C7H6O5 | 169 (28), 125 (100), 81 (6), 79 (22), 69 (6) | Gallic acid (4) |
| 191.0562 | 0.5 | C7H12O6 | 191 (100), 93 (5), 85 (12) | Quinic acid | ||
| P2 | 0.52 | 355.0310 | 0.9 | C14H12O11 | 355 (31), 337 (100), 293 (6), 249 (29), 205 (50), 193 (40), 187 (8), 179 (22), 163 (26), 149 (18), 135 (5) | Chebulic acid (7) |
| 331.0675 | 1.3 | C13H16O10 | 331 (100), 271 (24), 211 (18), 169 (86), 151 (11), 125 (26), 123 (20), 107 (10) | Galloylglucoside (e.g., 15) | ||
| P3 | 1.04 | 153.0191 | −1.5 | C7H6O4 | 153 (2), 109 (100), 91 (10), 81 (7), 65 (9) | Protocatechuic acid (3) |
| 783.0649 | −4.8 | C34H24O22 | 783 (100), 631 (3), 481 (4), 451 (16), 301 (53), 275 (14), 257 (3) | Terflavin B | ||
| 933.0596 | −4.7 | C41H26O26 | 933 (100), 781 (10), 721 (8), 601 (13), 575 (4), 451 (4), 299 (4) | O-Galloyl punicalin | ||
| P4 | 1.58 | 1083.0570 | −2.1 | C48H28O30 | 1083 (100), 781 (9), 721 (3), 601 (15), 575 (5), 301 (3) 273 (2) | Punicalagin isomer I |
| P5 | 2.36 | 1083.0553 | −3.7 | C48H28O30 | 1083 (100), 781 (10), 721 (2), 601 (18), 575 (5), 301 (2) 273 (2) | Punicalagin isomer II |
| P6 | 2.63 | 291.0146 | −0.1 | C13H8O8 | 291 (6), 247 (100), 219 (8), 191 (19), 173 (10), 145 (17) | Brevifolincarboxylate |
| 121.0294 | −0.9 | C7H6O2 | 121 (86), 92 (100), 65 (3) | Hydroxybenzaldehyde (2) | ||
| P7 | 2.82 | 935.0763 | −3.5 | C41H28O26 | 935 (100), 633 (9), 301 (5), 275 (17) | O-Galloyl bis-HHDP-glc |
| 305.0693 | 8.6 | C15H14O7 | 305 (60), 225 (34), 97 (100), 80 (16) | Gallocatechin | ||
| P8 | 3.01 | 651.0823 | −2.5 | C27H24O19 | 651 (98), 633 (9), 481 (44), 275 (26), 247 (12), 231 (18), 205 (14), 203 (15), 169 (100), 125 (29) | Chebulanin (17) |
| 469.0039 | −2.1 | C21H10O13 | 425 (27), 301 (100), 299 (27), 282 (9), 271 (14), 244 (9), 228 (8), 216 (8), 200 (6), 172 (12), 144 (6) | Flavogallonic acid | ||
| P9 | 3.17 | 633.0677 | −8.9 | C27H22O18 | 633 (100), 463 (6), 301 (66), 275 (7) | Corilagin |
| P10 | 3.31 | 1235.0712 | 0.8 | C55H32O34 | 1235 (19), 617 (100) | O-Galloyl punicalagin |
| P11 | 3.45 | 163.0401 | 0.2 | C9H8O3 | 163 (3), 119 (100), 117 (8), 93 (62) | p-Coumaric acid (6) |
| 600.9896 | −0.01 | C28H10O16 | 601 (100), 583 (3), 301 (18), 298 (20), 271 (22), 243 (5), 214 (2) | Terminalin | ||
| P12 | 3.84 | 433.0397 | −3.6 | C19H14O12 | 433 (78), 301 (77), 300 (100), 272 (4), 244 (7), 216 (10), 200 (5), 172 (6), 132 (4) | Ellagic acid pentoside |
| P13 | 3.93 | 953.0899 | −0.3 | C41H30O27 | 953 (100), 463 (2), 301 (61), 275 (5), 205 (4), 169 (2) | Chebulagic acid (18) |
| P14 | 4.02 | 300.9978 | −4.0 | C14H6O8 | 301 (100), 284 (7), 257 (2), 245 (3), 229 (4), 201 (3), 185 (3), 173 (3), 161 (1), 145 (5) | Ellagic acid (12) |
| P15 | 4.20 | 431.0969 | −3.4 | C21H20O10 | 431 (48), 341 (25), 323 (6), 311 (100), 283 (54), 269 (6), 161 (6), 117 (8) | Flavon-C-glucoside (Vitexin) |
| P16 | 4.42 | 955.1002 | −5.9 | C41H32O27 | 955 (100), 937 (6), 785 (4), 617 (3), 465 (4), 337 (3), 319 (4), 275 (8), 231 (11), 205 (6), 169 (4) | Chebulinic acid |
| P17 | 4.53 | 477.1006 | −6.8 | C22H22O12 | 477 (100), 313 (6), 265 (26), 235 (7), 211 (5), 205 (7), 169 (37), 163 (6) | Galloyl-coumaroyl-glucose (1) |
| P18 | 4.95 | 315.0126 | −6.5 | C15H8O8 | 315 (11), 300 (100), 271 (7), 216 (12), 200 (6), 160 (7), 132 (7) | O-Methyl ellagic acid |
| 583.1075 | −3.1 | C28H24O14 | 583 (100), 431 (22), 341 (31), 323 (8), 311 (75), 283 (40), 271 (14), 241 (10), 211 (9), 169 (18), 125 (8) | Galloylvitexin isomer I | ||
| P19 | 5.42 | 583.1085 | −1.4 | C28H24O14 | 583 (84), 431 (37), 341 (55), 311 (100), 323 (17), 283 (53), 271 (27), 211 (14), 169 (23), 125 (9) | Galloylvitexin isomer II |
| P20 | 5.79 | 735.1184 | −2.6 | C35H28O18 | 735 (100), 583 (90), 565 (45), 431 (24), 341 (25), 311 (39), 293 (42), 271 (24), 211 (20), 169 (38), 125 (8) | Digalloylvitexin isomer |
| 545.2010 | -3.4 | C28H34O11 | 545 (100), 307 (6), 265 (37), 235 (15), 219 (18), 205 (34), 201 (9), 177 (11), 163 (43), 145 (69), 119 (16) | e.g., Cinnamrutinose B | ||
| P21 | 6.24 | 329.0293 | −3.0 | C16H10O8 | 329 (8), 314 (55), 298 (85), 271 (100), 243 (48), 214 (29), 187 (21), 159 (24), 131 (12), 103 (7), 75 (6) | Di-O-methyl ellagic acid |
| P22 | 7.80 | 343.0443 | −4.8 | C17H12O8 | 343 (9), 328 (45), 313 (100), 297 (87), 285 (22), 269 (77), 241 (16), 213 (30), 185 (32), 157 (19), 130 (12) | Tri-O-methyl ellagic acid |
| Moiety | No. | δC, Type | δH (Multiplicity, J) | HMBC | COSY |
|---|---|---|---|---|---|
| Glucose | 1α | 95.9, CH | 5.66 (d, 3.3, 0.5 H) | 3, 4, 7′ | 2 |
| 1β | 5.66 (d, 7.7, 0.5 H) | 3, 4, 7′ | 2 | ||
| 2 | 74.1, CH | 3.46–3.52 (m, 2 H) | 1, 3, 4 | 1, 3 | |
| 3 | 78.1, CH | 3.46–3.52 (m, 2 H) | 2, 3, 4 | 2, 4 | |
| 4 | 71.3, CH | 3.41–3.46 (m, 1 H) | 3, 5 | 3, 5 | |
| 5 | 76.3, CH | 3.65–3.70 (m, 1 H) | 1, 3, 4 | 4, 6a, 6b | |
| 6a | 64.4, CH2 | 4.50 (dd, 12.2, 2.8 Hz, 1 H) | 4, 5, 9″ | 5, 6b | |
| 6b | 4.31 (dd, 12.2, 5.6 Hz, 1 H) | 4, 5, 9″ | 5, 6a | ||
| Galloyl | 1′ | 120.5, C | - | - | - |
| 2′/6′ | 110.5, CH | 7.13 (s, 2 H) | 1′, 2′, 3′, 4′, 5′, 6′, 7′ | - | |
| 3′/5′ | 146.6, C | - | - | ||
| 4′ | 140.7, C | - | - | ||
| 7′ | 166.9, C | - | - | ||
| Coumaroyl | 1″ | 127.2, C | - | - | - |
| 2″/6″ | 131.2, CH | 7.45 (d, 8.5 Hz, 2 H) | 2″, 4″, 6″, 7″ | 3″, 5″ | |
| 3″/5″ | 116.9, CH | 6.80 (d, 8.5 Hz, 2 H) | 1″, 3″, 4″, 5″ | 2″, 6″ | |
| 4″ | 161.3, C | - | - | - | |
| 7″ | 146.9, CH | 7.63 (d, 15.7 Hz, 1 H) | 1″, 2″, 6″, 8″, 9″ | 8″ | |
| 8″ | 114.9, CH | 6.36 (d, 15.7 Hz, 1 H) | 1′, 9′ | 7″ | |
| 9″ | 169.1, C | - | - | - |
| Growth Inhibition [%] * | ||||
|---|---|---|---|---|
| Antifungal Assays | Antibacterial Assay | |||
| S. tritici | B. cinerea | P. infestans | A. fischeri | |
| Crude extract | 100 µg/mL | 100 µg/mL | 100 µg/mL | 500 µg/mL |
| 79 ± 2 | 29 ± 9 | 82 ± 5 | 100 ± 1 | |
| Compound | 100 µM | 100 µM | 100 µM | 100 µM |
| 1 | 46 ± 1 | 1 ± 6 | 79 ± 7 | −28 ± 6 |
| 2 | 41 ± 1 | −13 ± 7 | 61 ± 10 | −51 ± 10 |
| 3 | 4 ± 5 | 11 ± 3 | 13 ± 2 | −29 ± 3 |
| 4 | 34 ± 3 | −22 ± 15 | 31 ± 1 | −5 ± 3 |
| 5 | −21 ± 14 | −25 ± 14 | 6 ± 1 | −31 ± 5 |
| 6 | 92 ± 1 | −11 ± 28 | 10 ± 5 | −6 ± 5 |
| 7 | 35 ± 4 | −4 ± 8 | 30 ± 8 | −114 ± 8 |
| 8 | 44 ± 1 | 12 ± 2 | 73 ± 4 | −76± 11 |
| 9 | 51 ± 1 | 6 ± 8 | 81 ± 1 | −77 ± 7 |
| 10 | 54 ± 3 | 14 ± 3 | 79 ± 1 | −51 ± 3 |
| 11 | 14 ± 1 | 15 ± 8 | 32 ± 11 | −22 ± 5 |
| 12 | −2 ± 8 | 23 ± 4 | 63 ± 4 | −389 ± 26 |
| 13 | 19 ± 7 | 4 ± 7 | 93 ± 1 | −384 ± 27 |
| 14 | 36 ± 1 | −6 ± 7 | 2 ± 4 | −17 ± 7 |
| 15 | 38 ± 3 | −35 ± 34 | 39 ± 4 | −172 ± 11 |
| 16 | −119 ± 26 | 20 ± 2 | 0 ± 2 | −17 ± 5 |
| 17 | 46 ± 2 | −3 ± 4 | 82 ± 1 | −350 ± 111 |
| 18 | 13 ± 4 | 2 ± 4 | 19 ± 2 | −23 ± 4 |
| 19 | 61 ± 3 | −17 ± 5 | 94 ± 1 | −643 ± 10 |
| 20 | −4 ± 5 | 8 ± 4 | 94 ± 1 | −564 ± 45 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kappen, J.; Rashan, L.; Franke, K.; Wessjohann, L.A. Profiling and Bioactivity of Polyphenols from the Omani Medicinal Plant Terminalia dhofarica (syn. Anogeissus dhofarica). Molecules 2025, 30, 952. https://doi.org/10.3390/molecules30040952
Kappen J, Rashan L, Franke K, Wessjohann LA. Profiling and Bioactivity of Polyphenols from the Omani Medicinal Plant Terminalia dhofarica (syn. Anogeissus dhofarica). Molecules. 2025; 30(4):952. https://doi.org/10.3390/molecules30040952
Chicago/Turabian StyleKappen, Jonas, Luay Rashan, Katrin Franke, and Ludger A. Wessjohann. 2025. "Profiling and Bioactivity of Polyphenols from the Omani Medicinal Plant Terminalia dhofarica (syn. Anogeissus dhofarica)" Molecules 30, no. 4: 952. https://doi.org/10.3390/molecules30040952
APA StyleKappen, J., Rashan, L., Franke, K., & Wessjohann, L. A. (2025). Profiling and Bioactivity of Polyphenols from the Omani Medicinal Plant Terminalia dhofarica (syn. Anogeissus dhofarica). Molecules, 30(4), 952. https://doi.org/10.3390/molecules30040952