
Physalin F, a Potent Inhibitor of Lymphocyte Function, Is a Calcineurin Inhibitor and Has Synergistic Effect with Dexamethasone

 ,
,
Abstract
1. Introduction
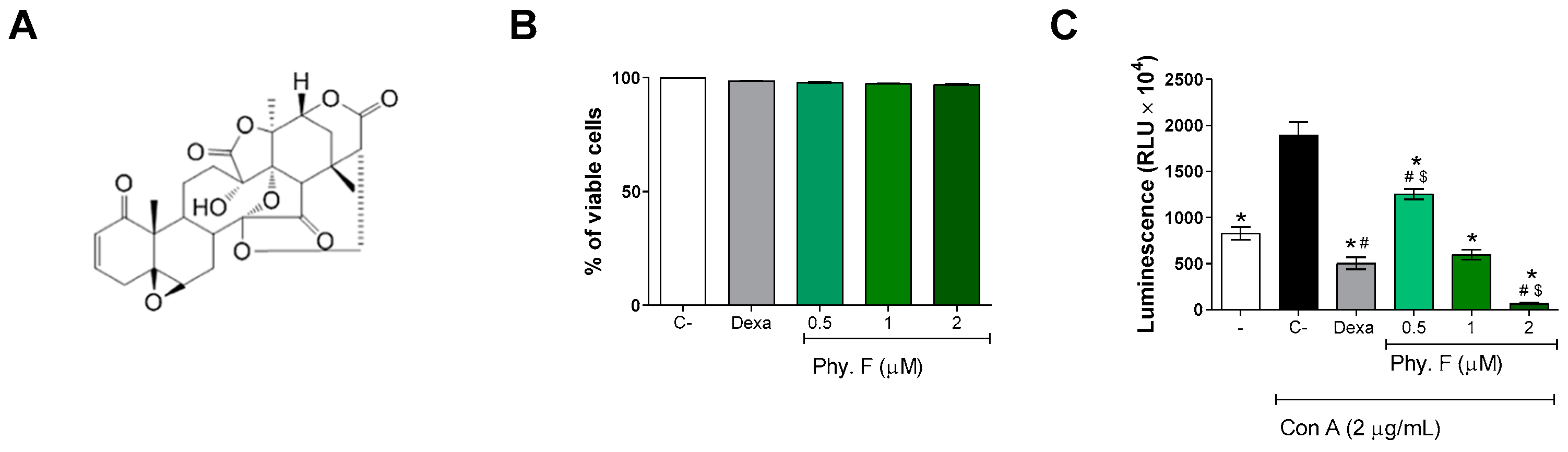
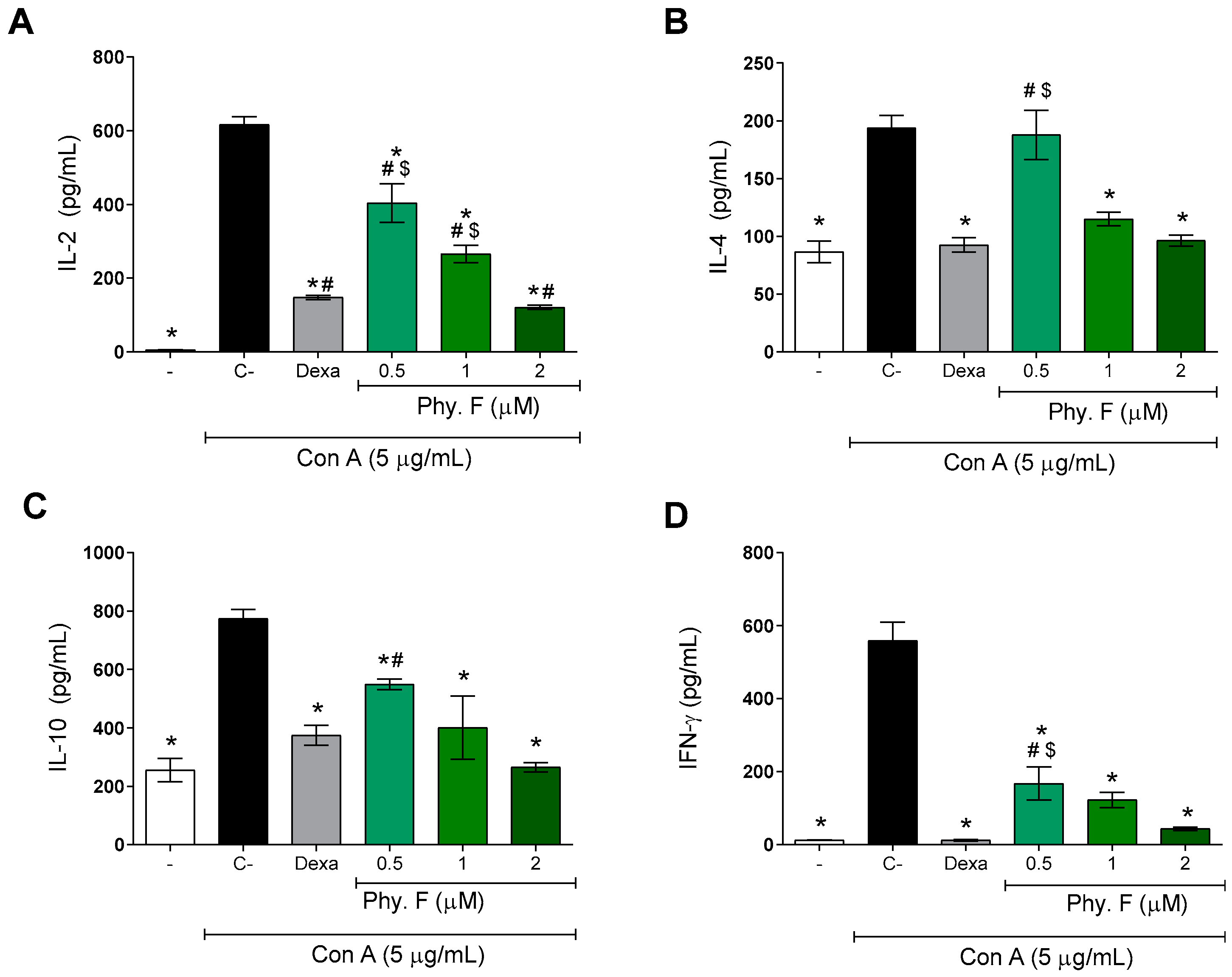
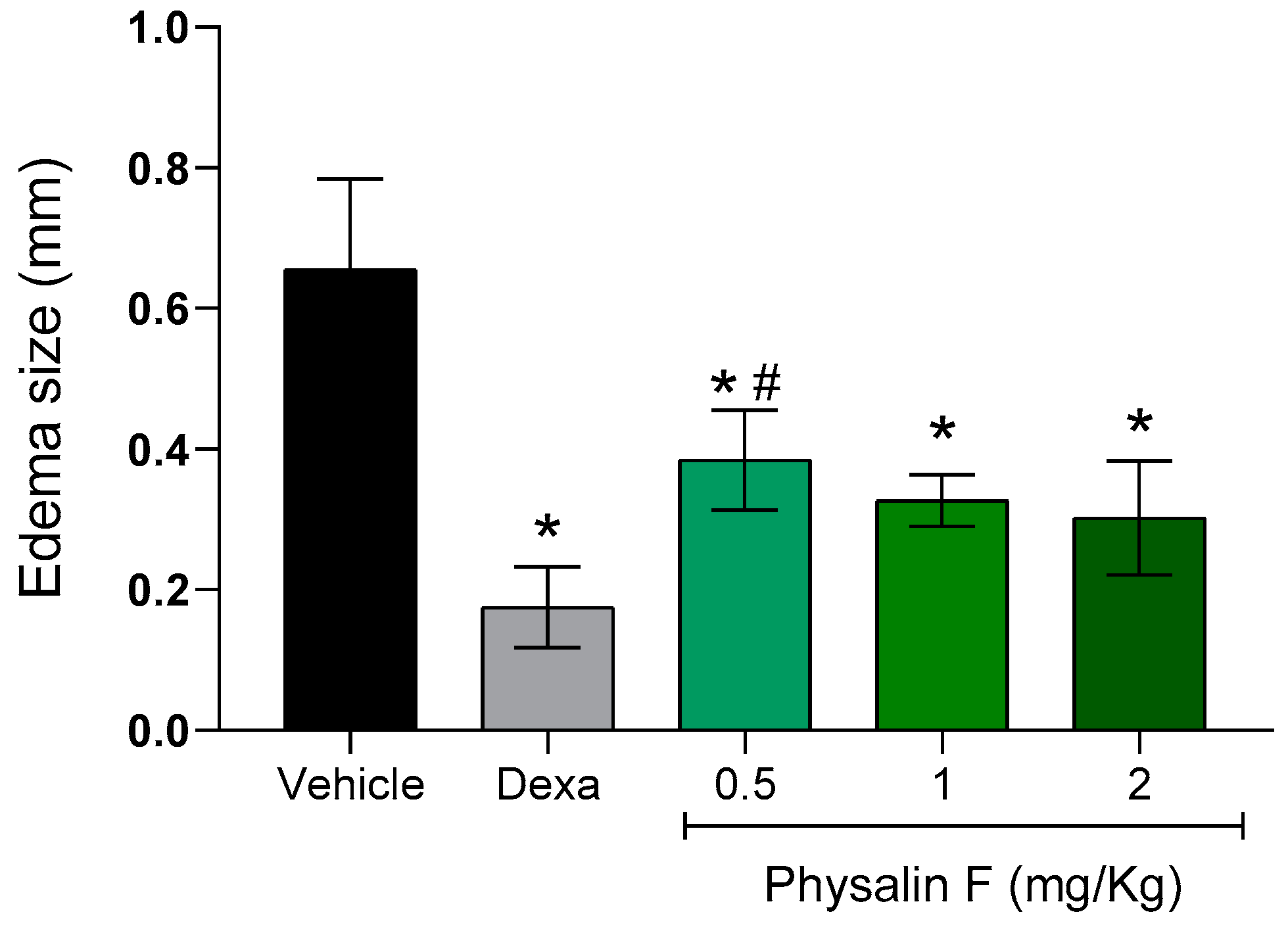
2. Results and Discussion
3. Materials and Methods
3.1. Compounds
3.2. Animals
3.3. Cytotoxicity to Mammalian Cells
3.4. Lymphoproliferation Assay
3.5. Assessment of Cytokine Production
3.6. Calcineurin Activity
3.7. Combination Therapy
3.8. Delayed-Type Hypersensitivity Assay
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Hanaoka, B.Y.; Shimizu, K.; Doke, T.; Koide, H. Chronic Inflammation in RA: Mediator of Skeletal Muscle Pathology and Physical Impairment. Arthritis Care Res. 2019, 71, 173. [Google Scholar] [CrossRef] [PubMed]
- Schenk, M.; Bierwirth, T.; Berkowitz, S.A. TREM-1–Expressing Intestinal Macrophages Crucially Amplify Chronic Inflammation in Experimental Colitis. J. Clin. Investig. 2007, 117, 3097–3106. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.L.; Su, Y.W.; Chen, C.J.; Chang, S.H.; Huang, W.C.; Chien, H.W.; Wang, S.C.; Lee, W.S.; Lin, C.W.; Chen, W.H. Corylin Protects LPS-Induced Sepsis and Attenuates LPS-Induced Inflammatory Response. Sci. Rep. 2017, 7, 46299. [Google Scholar] [CrossRef] [PubMed]
- Perez-Sepulveda, A.; Torres, M.J.; Khan, R.N.; Oliver, C. Innate Immune System and Preeclampsia. Front. Immunol. 2014, 5, 244. [Google Scholar] [CrossRef]
- Ingawale, D.K.; Mandlik, S.K.; Patel, S.S. An Emphasis on Molecular Mechanisms of Anti-Inflammatory Effects and Glucocorticoid Resistance. J. Complement. Integr. Med. 2015, 12, 1–13. [Google Scholar] [CrossRef]
- Rosenblatt, M.I. Anti-Infective and Anti-Inflammatory Pharmacotherapies. In Pharmacologic Therapy of Ocular Disease; Springer: Cham, Switzerland, 2017; pp. 119–135. [Google Scholar]
- Schäcke, H.; Döcke, W.D.; Asadullah, K. Mechanisms Involved in the Side Effects of Glucocorticoids. Pharmacol. Ther. 2002, 96, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Dias, D.A.; Urban, S.; Roessner, U. A Historical Overview of Natural Products in Drug Discovery. Metabolites 2012, 2, 303–336. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs Over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Li, A.L.; Chen, B.J.; Li, G.H.; Zhou, M.X.; Li, Y.R.; Ren, D.M.; Lou, H.X.; Wang, X.N.; Shen, T. Physalis alkekengi L. var. franchetii (Mast.) Makino: An Ethnomedical, Phytochemical and Pharmacological Review. J. Ethnopharmacol. 2018, 210, 260–274. [Google Scholar] [CrossRef] [PubMed]
- Tomassini, T.C.B.; Silva, M.T.; Pinheiro, M.L.B.; Fernandes, H.B. Gênero Physalis: Uma Revisão Sobre Vitaesteróides. Quím. Nova 2000, 23, 47–57. [Google Scholar] [CrossRef]
- Zhang, Q.; Lenardo, M.J.; Baltimore, D. 30 Years of NF-κB: A Blossoming of Relevance to Human Pathobiology. Cell 2017, 168, 37–57. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.P.; Wang, Y.; Zhou, W.; Zhang, L. Unprecedented 22,26-Seco Physalins from Physalis angulata and Their Anti-Inflammatory Potential. Org. Biomol. Chem. 2017, 15, 8700–8704. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.B.; Bellintani, M.C.; Ribeiro, I.M.; Tomassini, T.C.; dos Santos, R.R. Inhibition of Macrophage Activation and Lipopolysaccharide-Induced Death by Seco-Steroids Purified from Physalis angulata L. Eur. J. Pharmacol. 2003, 459, 107–112. [Google Scholar] [CrossRef]
- Meira, C.S.; Santos, R.R.; da Silva, A.J.; Siqueira-Neto, J.L.; Soares, M.B.P. Physalins B and F, Seco-Steroids Isolated from Physalis angulata L., Strongly Inhibit Proliferation, Ultrastructure, and Infectivity of Trypanosoma cruzi. Parasitology 2013, 140, 1811–1821. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.S.; Meira, C.S.; Soares, M.B.P.; Tomassini, T.C.B.; dos Santos, R.R. Antinociceptive Properties of Physalins from Physalis angulata. J. Nat. Prod. 2014, 77, 2397–2403. [Google Scholar] [CrossRef]
- Wang, A.; Sun, C.; Zhang, Q.; Li, J. Physalin B Induces Cell Cycle Arrest and Triggers Apoptosis in Breast Cancer Cells through Modulating p53-Dependent Apoptotic Pathway. Biomed. Pharmacother. 2018, 101, 334–341. [Google Scholar] [CrossRef]
- Meira, C.S.; Santos, R.R.; Soares, M.B.P. Therapeutic Applications of Physalins: Powerful Natural Weapons. Front. Pharmacol. 2022, 13, 864714. [Google Scholar] [CrossRef] [PubMed]
- Pinto, L.A.; Soares, M.B.P.; Meira, C.S.; Santos, R.R. Physalin F, a Seco-Steroid from Physalis angulata L., Has Immunosuppressive Activity in Peripheral Blood Mononuclear Cells from Patients with HTLV1-Associated Myelopathy. Biomed. Pharmacother. 2016, 79, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Sá, M.S.; de Menezes, M.N.; Krettli, A.U.; Ribeiro, I.M.; Tomassini, T.C.; Ribeiro dos Santos, R.; de Azevedo, W.F., Jr.; Soares, M.B. Antimalarial Activity of Physalins B, D, F, and G. J. Nat. Prod. 2011, 74, 2269–2272. [Google Scholar] [CrossRef] [PubMed]
- Soares, M.B.P.; Brustolim, D.; Santos, L.A.; Bellintani, M.C.; Paiva, F.P.; Ribeiro, Y.M.; Tomassini, T.C.; Ribeiro dos Santos, R. Physalins B, F, and G, Seco-Steroids Purified from Physalis angulata L., Inhibit Lymphocyte Function and Allogeneic Transplant Rejection. Int. Immunopharmacol. 2006, 6, 408–414. [Google Scholar] [CrossRef]
- Yu, Y.; Sun, L.; Ma, L.; Li, J.; Hu, L.; Liu, J. Investigation of the Immunosuppressive Activity of Physalin H on T Lymphocytes. Int. Immunopharmacol. 2010, 10, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Abbas, A.K.; Trotta, E.; Simeonov, D.R.; Marson, A.; Bluestone, J.A. Revisiting IL-2: Biology and Therapeutic Prospects. Sci. Immunol. 2018, 3, eaat1482. [Google Scholar] [CrossRef]
- Kak, G.; Raza, M.; Tiwari, B.K. Interferon-Gamma (IFN-γ): Exploring Its Implications in Infectious Diseases. Biomol. Concepts 2018, 9, 64–79. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, M.; Vieira, P.; O’Garra, A. Biology and Therapeutic Potential of Interleukin-10. J. Exp. Med. 2020, 217, e20190418. [Google Scholar] [CrossRef] [PubMed]
- Mikhalkevich, N.; Kurohara, M.L.; Altman, A. Responsiveness of Naive CD4 T Cells to Polarizing Cytokine Determines the Ratio of Th1 and Th2 Cell Differentiation. J. Immunol. 2006, 176, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Luckheeram, R.V.; Zhao, B.; Shi, Z.; Zhou, Z. CD4+ T Cells: Differentiation and Functions. Clin. Dev. Immunol. 2012, 2012, 925135. [Google Scholar] [CrossRef] [PubMed]
- Kaminuma, O.; Ishii, K.; Mori, A.; Miyasaka, T.; Kumazawa, T.; Kitamura, N. T-box 21 Transcription Factor Is Responsible for Distorted TH2 Differentiation in Human Peripheral CD4+ T Cells. J. Allergy Clin. Immunol. 2009, 123, 813–823. [Google Scholar] [CrossRef]
- Jacobo-Herrera, N.J.; Andrade-Cetto, A.; Heinrich, M.; León-Rivera, I.; Rivera, D. Physalins from Witheringia solanacea as Modulators of the NF-κB Cascade. J. Nat. Prod. 2006, 69, 328–331. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, D.E.; Kirschner, K.; Demirci, H.; Himmerkus, N.; Bachmann, S.; Mutig, K. Immunosuppressive Calcineurin Inhibitor Cyclosporine A Induces Proapoptotic Endoplasmic Reticulum Stress in Renal Tubular Cells. J. Biol. Chem. 2022, 298, 101589. [Google Scholar] [CrossRef]
- Safarini, O.A.; Patel, J. Calcineurin Inhibitors. Drugs Today 2020, 56, 1–15. [Google Scholar]
- Colombel, J.F.; Sandborn, W.J.; Reinisch, W.; Mantzaris, G.J.; Kornbluth, A.; Rachmilewitz, D.; Lichtiger, S.; d’Haens, G.; Diamond, R.H.; Broussard, D.L.; et al. Infliximab and Azathioprine for the Treatment of Crohn’s Disease. N. Engl. J. Med. 2010, 362, 1383–1395. [Google Scholar] [CrossRef]
- Lus, G.; Romano, F.; Scuotto, A.; Accardo, C.; Cotrufo, R. Azathioprine and interferon beta(1a) in relapsing-remitting multiple sclerosis patients: Increasing efficacy of combined treatment. Eur. Neurol. 2004, 51, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.C.; Ortigosa, L.C.M.; Benard, G. Anti-TNF-α agents in the treatment of immune-mediated inflammatory diseases: Mechanisms of action and pitfalls. Immunotherapy 2010, 2, 817–833. [Google Scholar] [CrossRef] [PubMed]
- Meira, C.S.; Guimarães, E.T.; Dos Santos, J.A.; Moreira, D.R.; Nogueira, R.C.; Tomassini, T.C.; Ribeiro, I.M.; de Souza, C.V.; Dos Santos, R.R.; Soares, M.B. In vitro and in vivo Antiparasitic Activity of Physalis angulata L. Concentrated Ethanolic Extract Against Trypanosoma cruzi. Phytomedicine 2015, 22, 969–974. [Google Scholar] [CrossRef] [PubMed]
- Gholijani, N.; Amirghofran, Z. Effects of Thymol and Carvacrol on T-Helper Cell Subset Cytokines and Their Main Transcription Factors in Ovalbumin-Immunized Mice. J. Immunotoxicol. 2016, 13, 729–737. [Google Scholar] [CrossRef]
- Brustolim, D.; Santos, L.A.; Ribeiro, Y.M.; Tomassini, T.C.; Ribeiro dos Santos, R.; Soares, M.B.P. Activity of Physalin F in a Collagen-Induced Arthritis Model. J. Nat. Prod. 2010, 73, 1323–1326. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C.; Martin, N. CompuSyn for Drug Combinations: PC Software and User’s Guide: A Computer Program for Quantitation of Synergism and Antagonism in Drug Combinations, and the Determination of IC50 and ED50 and LD50 Values; ComboSyn: Paramus, NJ, USA, 2005. [Google Scholar]
- Fivelman, Q.L.; Adagu, I.S.; Warhurst, D.C. Modified Fixed-Ratio Isobologram Method for Studying in vitro Interactions Between Atovaquone and Proguanil or Dihydroartemisinin Against Drug-Resistant Strains of Plasmodium falciparum. Antimicrob. Agents Chemother. 2004, 48, 4097–4102. [Google Scholar] [CrossRef] [PubMed]
- Titus, R.G.; Chiller, J.M. A simple and effective method to assess murine delayed type hypersensitivity to proteins. J. Immunol. Methods 1981, 45, 65–78. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | EC50 ± S.D (μM) a | ||
|---|---|---|---|
| Drug Alone | Combination | CI b | |
| Physalin F | 0.65 ± 0.1 | 0.006 ± 0.007 | 0.17 ± 0.06 |
| Dexamethasone | 0.055 ± 0.01 | 0.004 ± 0.003 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, D.K.C.; Novo, L.B.d.C.; Ribeiro, I.M.; Barreto, B.C.; Opretzka, L.C.F.; Meira, C.S.; Soares, M.B.P. Physalin F, a Potent Inhibitor of Lymphocyte Function, Is a Calcineurin Inhibitor and Has Synergistic Effect with Dexamethasone. Molecules 2025, 30, 916. https://doi.org/10.3390/molecules30040916
Silva DKC, Novo LBdC, Ribeiro IM, Barreto BC, Opretzka LCF, Meira CS, Soares MBP. Physalin F, a Potent Inhibitor of Lymphocyte Function, Is a Calcineurin Inhibitor and Has Synergistic Effect with Dexamethasone. Molecules. 2025; 30(4):916. https://doi.org/10.3390/molecules30040916
Chicago/Turabian StyleSilva, Dahara Keyse Carvalho, Laura Beatriz da Cruz Novo, Ivone Maria Ribeiro, Breno Cardim Barreto, Luiza Carolina França Opretzka, Cássio Santana Meira, and Milena Botelho Pereira Soares. 2025. "Physalin F, a Potent Inhibitor of Lymphocyte Function, Is a Calcineurin Inhibitor and Has Synergistic Effect with Dexamethasone" Molecules 30, no. 4: 916. https://doi.org/10.3390/molecules30040916
APA StyleSilva, D. K. C., Novo, L. B. d. C., Ribeiro, I. M., Barreto, B. C., Opretzka, L. C. F., Meira, C. S., & Soares, M. B. P. (2025). Physalin F, a Potent Inhibitor of Lymphocyte Function, Is a Calcineurin Inhibitor and Has Synergistic Effect with Dexamethasone. Molecules, 30(4), 916. https://doi.org/10.3390/molecules30040916