
Immunometabolic Effects of Ginger (Zingiber officinale Roscoe) Supplementation in Obesity: A Comprehensive Review

 , , and
, , and 
Abstract

1. Introduction
2. Metabolic and Immune Alterations in Obesity
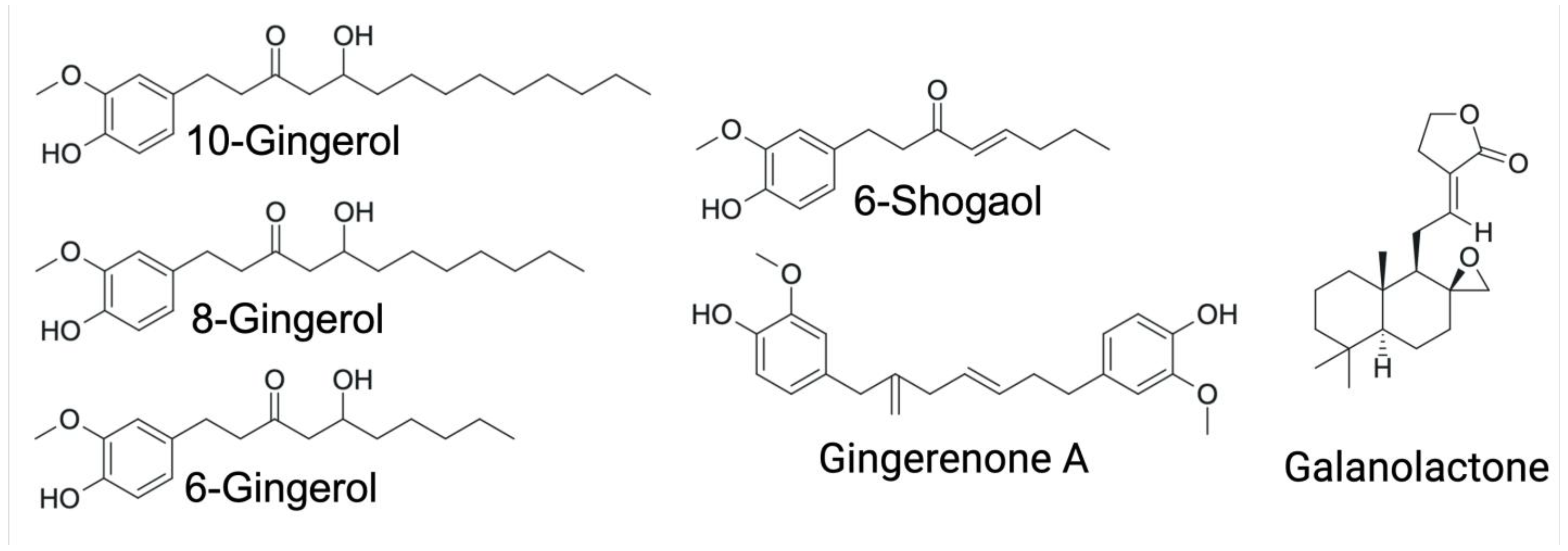
3. Ginger and Its Bioactive Compounds
4. Immunometabolic Effects of Ginger Bioactive Compounds in Obesity
4.1. In Vitro Studies
4.2. In Vivo Studies
4.3. Clinical Trials

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Trials | |||||
| Author, Year | Subjects | Intervention | Dose | Duration | Results |
| Kamari N et al. (2023) [61] | 160 children with obesity | Ginger rhizome powder | 1000 mg/day | 12 weeks | Serum fasting blood sugar and high-sensitivity C-reactive protein levels were significantly decreased. Significant reduction in BMI, waist circumference, waist-to-height ratio, ALT, total cholesterol, and LDL-C. |
| Ebrahimzadeh et al. (2016) [63] | 80 obese women (18–45 years) | Ginger powder in tea | 2 g/day | 12 weeks | BMI, serum insulin, HOMA-IR significantly decreased; QUICKI increased; serum leptin, resistin, and glucose also decreased. |
| Shidfar F et al. (2015) [65] | 45 type 2 diabetic patients | Ginger powder (capsules) | 3 g/day | 3 months | Improved glucose, HbA1c, insulin, insulin resistance, CRP, antioxidant capacity, PON-1. |
| Karimi N et al. (2015) [71] | 40 women with breast neoplasms | Ginger rhizome powder | 4 × 750 mg/day | 6 weeks | Reduced IL-10, hs-CRP, insulin; increased HDL-C and HDL/LDL ratio. |
| Arablou T et al. (2014) [62] | 70 type 2 diabetic patients | Ginger rhizome powder | 1600 mg/day | 12 weeks | Reduced TG, total cholesterol, fasting glucose, HbA1C, insulin, HOMA, CRP, PGE2; no change in HDL or LDL. |
| Mozaffari-Khosravi H et al. (2014) [64] | 88 type 2 diabetic patients | Ginger powder | 3 g/day | 8 weeks | Reduced fasting blood sugar and HbA1c; improved QUICKI. |
| Mansour MS et al. (2012) [73] | 10 overweight men | Ginger powder (hot water) | 2 g | Single dose | Enhanced thermogenesis and reduced hunger and food intake compared to placebo. |
| Atashak S et al. (2011) [60] | 32 obese men | Ginger powder | 1 g/day | 10 weeks | No effect on lipids or insulin resistance; CRP decreased. |
| Alizadeh-Navaei R et al. (2008) [68] | 85 hyperlipidemic patients | Ginger capsules | 3 g/day | 45 days | Reduced TG, cholesterol, LDL. |
| Mohammadzadeh Honarvar et al. (2019) [74] | 48 Type 2 diabetic patients | Oral ginger capsules | 2 g/day | 10 weeks | Trend towards reduced NF-κB; reduced systemic inflammation; decreased hip circumference. |
| In vivo studies | |||||
| Author, Year | Animal Model | Intervention | Dose/Groups | Duration | Results |
| Hong KH et al. (2023) [45] | Male C57BL/6J mice | 6-gingerol | ND, HFD, HFD + 0.05% 6G | 8 weeks | Reduced weight gain, WAT mass, adipocyte size, serum insulin/leptin/TG; increased adiponectin; reduced TNF-α, MCP-1. |
| Cheng Z et al. (2022) [46] | Male C57BL/6J mice | 6-gingerol | ND, HFD, HFD + 25 mg/kg | 4 weeks | Reduced body weight, TC, and TG levels. |
| Seo SH et al. (2021) [75] | Male C57BL/6 mice | Ginger powder | -LF: low-fat diet (16% of calories from fat) -HFD -HFD + 5% ginger powder | 7 weeks | Reduced body weight, glucose, total cholesterol, and hepatic lipids were observed, along with a reduction in adipocyte size. Additionally, supplementation upregulated the expression of FGF21, ACOX1, CPT1, and antioxidant enzymes SOD1/2, NRF1/2, GPX. |
| Sayed et al. (2020) [49] | Adult male Wistar rats | Ginger water | Control, 25%, 50% (v/v) | 4 weeks | Ginger water administration significantly reduced serum total cholesterol and triglyceride levels compared to the control group. Treatment with 25% and 50% ginger water induced a 50% and 60% downregulation, respectively, in SREBP-1c mRNA expression, with no changes observed in HSL mRNA levels. In white adipose tissue (WAT), leptin and resistin mRNA expression levels were significantly decreased, whereas adiponectin mRNA expression was upregulated. |
| Wang J et al. (2019) [48] | Male C57BL/6J mice | Ginger powder | -NCD: normal control diet (10% of calories from fat) -NCD-G: normal control diet supplemented with ginger. -HFD -HFD + G: high-fat diet with ginger supplementation (500 mg/kg, w/w) | 16 weeks | Ginger supplementation alleviated the HFD-induced increases in body weight, fat accumulation, and serum levels of glucose, triglycerides, and cholesterol. It also corrected the HFD-induced alterations in the concentrations of intermediates involved in glycolysis and the TCA cycle. |
| Kim S et al. (2018) [26] | Sprague–Dawley rats | Ginger Extract | HFD, HFD + WEG/HPG (8 g/kg) | 10 weeks | A reduction in body weight, serum lipids (TG, TC, LDL-C), and inflammatory cytokines (TNF-α, IL-6) was observed, along with decreased hepatic levels of total lipids, TG, and TC compared to the HFD group. Additionally, HDL-C levels were significantly higher in the WEG and HPG groups. These effects were associated with AMPK activation and modulation of anti-inflammatory microRNAs, including miR-21 and miR-132, in white adipose tissue (WAT). |
| Suk et al. (2017) [36] | Male C57BL/6J mice | Gingerol A (GA) | ND, HFD, HFD + GA (10/50 mg/kg) | 15 weeks | A reduction in body fat, body weight, adipocyte size, and inflammation was observed, accompanied by increased levels of ATGL and phosphorylated HSL and decreased expression of SREBP-1 and FAS. In addition, macrophage infiltration and the expression of pro-inflammatory markers such as TNF-α and MCP-1 in adipose tissue were reduced. GA also upregulated the expression of genes involved in mitochondrial biogenesis, NRF1 and TFAM, in WAT. |
| Brahma Naidu et al. (2016) [44] | HFD-induced obese rats | [6]-gingerol | HFD, HFD + 75 mg/kg | 30 days | A reduction in body weight, glucose, insulin, and fat was observed. The gingerol-treated group exhibited decreased activity of lipogenic proteins, including HMG-CoA reductase, FAS, PPARγ, and SREBP1c, as well as lower expression of inflammatory markers TNF-α and IL-6 compared to the control group. |
| Misawa K et al. (2015) [76] | Male C57BL/6J mice | Ginger extract | LF, HFD, HFD + 0.3% GE | 18 weeks | Reduced adipocyte size, leptin, cholesterol, insulin, HOMA-R. |
| Saravanan et al. (2014) [47] | HFD-induced obese rats | [6]-gingerol | HFD + 25/50/75 mg/kg | 10 weeks | Supplemented groups exhibited significantly lower body weight and reduced adipose tissue mass, as well as decreased levels of leptin, glucose, insulin, and serum lipid profile parameters. |
| Oliveira et al. (2018) [51] | In vivo: BALB/c mice on high refined carbohydrate diet | Ginger extract (5% gingerols) | 200, 600, 1800 mg/kg | 4 weeks | Reduced adiposity, decreased TNF-α and IL-6 (non-significant), and IL-13 in adipose tissue, increased serum adiponectin, and decreased leukocyte infiltration, improving immunometabolic dysfunction. |
| Kim et al. (2018) [52] | In vivo: Colitis model | Ginger extract | 100–500 mg/kg | 21 days | Reduced IL-6, TNF-α, and IL-1β in the colon; increased tight junction proteins (ZO-1, occludin), improved epithelial integrity and reduced systemic inflammation. |
| In vitro studies | |||||
| Author, Year | Cell Line | Treatment | Dose | Duration | Results |
| Tzeng et al. (2013) [33] | 3T3-L1 preadipocytes | [6]-gingerol | 5, 10, 15 µg/mL | 8 days | Inhibited adipogenesis and lipid accumulation; reduced PPARγ, C/EBPα, FAS, aP2; decreased p-GSK3β, Akt. |
| Li et al. (2015) [34] | 3T3-L1 preadipocytes | [6]-gingerol | 5, 10, 15 µg/mL | 7 days | Adipogenic differentiation was inhibited in a dose-dependent manner, accompanied by reduced mRNA levels and protein expression of PPARγ, C/EBPα, FAS, and ACC. Increased mRNA levels and expression of β-catenin, CCND1, LRP6, and DVL2 were also observed. |
| Suk et al. (2016) [37] | 3T3-L1 preadipocytes | [6]-gingerol, [6]-shogaol | 10, 20, 40 µM | 6 days | 6-Shogaol inhibits adipogenesis and more potently decreases the expression of adipogenic (PPARγ, C/EBPα) and lipogenic (FAS) proteins compared to 6-gingerol. |
| Rani et al. (2012) [30] | 3T3-L1 preadipocytes | Ethyl-acetate ginger extract | 0.1–50 µg/mL | 10 days | Inhibited differentiation; 50 µg/mL reduced lipids by 43.5%. |
| Suk et al. (2017) [36] | 3T3-L1 preadipocytes | Multiple gingerols and shogaol | 40 µM | 6 days | Gingerol A showed the strongest anti-adipogenic/lipogenic effect; 10-gingerol was the second most effective. |
| Ahn et al. (2012) [35] | 3T3-L1 preadipocytes | Galanolactone | 25, 50, 100 µM | 8 days | Dose-dependent inhibition of differentiation and lipid accumulation; decreased the levels of mRNA and proteins PPARγ, C/EBPα, aP2, and resistin. |
| Lee et al. (2009) [40] | In vitro: RAW 264.7 macrophages stimulated with LPS | 6-Gingerol | 20, 40, 80 µM | 4–12 h | Inhibited iNOS and TNF-α, reduced ROS and intracellular Ca2+, blocked PKC-α and NF-κB, showing strong anti-inflammatory and antioxidant effects relevant for obesity. |
| Ho S. C et al. (2018) [42] | In vitro: Human THP-1 macrophages stimulated with LPS | 6-, 8-, 10-Gingerols and Shogaols | 5–20 µM | 24 h | 6-Shogaol inhibited IL-1β secretion by blocking NLRP3 inflammasome activation and reduced TNF-α levels, demonstrating a relevant anti-inflammatory effect in the context of obesity. |
5. Modulation of Immunometabolic Alterations by Ginger Bioactive Compounds in Obesity
5.1. Ginger Compounds Suppress Adipogenesis and Lipogenesis
5.2. Ginger Compounds Promote Lipid Catabolism
5.3. AMPK: A Key Molecule in the Immunometabolic Effects of Ginger
5.4. Ginger Exhibits Epigenetic Mechanisms to Modulate Inflammation
5.5. Ginger Compounds Reduce Chronic Inflammation
5.6. Ginger Modulates Microbiota Composition and Function
5.7. Ginger Improves Hepatic and Serum Lipid Profiles
5.8. Ginger Improves Glycemic Control and Enhances Insulin Sensitivity
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sørensen, T.I.A.; Martinez, A.R.; Jørgensen, T.S.H. Epidemiology of Obesity. In From Obesity to Diabetes; Eckel, J., Clément, K., Eds.; Springer International Publishing: Cham, Switzerland, 2022; pp. 3–27. ISBN 978-3-030-99995-7. [Google Scholar]
- Blüher, M. Obesity: Global Epidemiology and Pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 26 May 2025).
- Bray, G.A.; Kim, K.K.; Wilding, J.P.H.; World Obesity Federation. Obesity: A Chronic Relapsing Progressive Disease Process. A Position Statement of the World Obesity Federation. Obes. Rev. 2017, 18, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Brondeel, K.C.; Lakatta, A.C.; Torres, G.B.; Hurley, J.J.; Kunik, I.L.; Haney, K.F.; Cornett, E.M.; Kaye, A.D. Physiologic and Pharmacologic Considerations in Morbid Obesity and Bariatric Anesthesia. Saudi J. Anaesth. 2022, 16, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimzadeh Attari, V.; Malek Mahdavi, A.; Javadivala, Z.; Mahluji, S.; Zununi Vahed, S.; Ostadrahimi, A. A Systematic Review of the Anti-Obesity and Weight Lowering Effect of Ginger (Zingiber officinale Roscoe) and Its Mechanisms of Action. Phytother. Res. 2018, 32, 577–585. [Google Scholar] [CrossRef]
- Anh, N.H.; Kim, S.J.; Long, N.P.; Min, J.E.; Yoon, Y.C.; Lee, E.G.; Kim, M.; Kim, T.J.; Yang, Y.Y.; Son, E.Y.; et al. Ginger on Human Health: A Comprehensive Systematic Review of 109 Randomized Controlled Trials. Nutrients 2020, 12, 157. [Google Scholar] [CrossRef]
- Sampath, S.J.P.; Birineni, S.; Perugu, S.; Kotikalapudi, N.; Venkatesan, V. Therapeutic Efficacy of 6-Gingerol and 6-Shogaol in Promoting Browning of White Adipocytes Vis-à-Vis Enhanced Thermogenesis Portrayed in High Fat Milieu. Food Biosci. 2021, 42, 101211. [Google Scholar] [CrossRef]
- Mao, Q.-Q.; Xu, X.-Y.; Cao, S.-Y.; Gan, R.-Y.; Corke, H.; Beta, T.; Li, H.-B. Bioactive Compounds and Bioactivities of Ginger (Zingiber officinale Roscoe). Foods 2019, 8, 185. [Google Scholar] [CrossRef]
- Unuofin, J.O.; Masuku, N.P.; Paimo, O.K.; Lebelo, S.L. Ginger from Farmyard to Town: Nutritional and Pharmacological Applications. Front. Pharmacol. 2021, 12, 779352. [Google Scholar] [CrossRef]
- Andersen, C.J.; Murphy, K.E.; Fernandez, M.L. Impact of Obesity and Metabolic Syndrome on Immunity. Adv. Nutr. 2016, 7, 66–75. [Google Scholar] [CrossRef]
- Parra-Peralbo, E.; Talamillo, A.; Barrio, R. Origin and Development of the Adipose Tissue, a Key Organ in Physiology and Disease. Front. Cell Dev. Biol. 2021, 9, 786129. [Google Scholar] [CrossRef]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and Metabolic Health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef]
- González-Muniesa, P.; Mártinez-González, M.-A.; Hu, F.B.; Després, J.-P.; Matsuzawa, Y.; Loos, R.J.F.; Moreno, L.A.; Bray, G.A.; Martinez, J.A. Obesity. Nat. Rev. Dis. Primers 2017, 3, 17034. [Google Scholar] [CrossRef] [PubMed]
- Zatterale, F.; Longo, M.; Naderi, J.; Raciti, G.A.; Desiderio, A.; Miele, C.; Beguinot, F. Chronic Adipose Tissue Inflammation Linking Obesity to Insulin Resistance and Type 2 Diabetes. Front. Physiol. 2020, 10, 1607. [Google Scholar] [CrossRef] [PubMed]
- Pchelin, I.Y.; Lukianets, K.Y.; Mironova, E.S.; Kosova, A.D.; Belova, Y.I.; Ayrapetyan, S.A. Inflammaging and metabolic syndrome: Molecular mechanisms. Adv. Gerontol. 2025, 38, 118–125. [Google Scholar] [PubMed]
- Giralt, M.; Cereijo, R.; Villarroya, F. Adipokines and the Endocrine Role of Adipose Tissues. In Metabolic Control; Herzig, S., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 265–282. ISBN 978-3-319-29806-1. [Google Scholar]
- Lago, F.; Gómez, R.; Gómez-Reino, J.J.; Dieguez, C.; Gualillo, O. Adipokines as Novel Modulators of Lipid Metabolism. Trends Biochem. Sci. 2009, 34, 500–510. [Google Scholar] [CrossRef]
- Scheja, L.; Heeren, J. The Endocrine Function of Adipose Tissues in Health and Cardiometabolic Disease. Nat. Rev. Endocrinol. 2019, 15, 507–524. [Google Scholar] [CrossRef]
- Luo, L.; Liu, M. Adipose Tissue in Control of Metabolism. J. Endocrinol. 2016, 231, R77–R99. [Google Scholar] [CrossRef]
- Makki, K.; Froguel, P.; Wolowczuk, I. Adipose Tissue in Obesity-Related Inflammation and Insulin Resistance: Cells, Cytokines, and Chemokines. Int. Sch. Res. Not. 2013, 2013, 139239. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, J.; Zhang, Y. Research Progress on Chemical Constituents of Zingiber officinale Roscoe. BioMed Res. Int. 2019, 2019, 5370823. [Google Scholar] [CrossRef]
- Zhang, M.; Zhao, R.; Wang, D.; Wang, L.; Zhang, Q.; Wei, S.; Lu, F.; Peng, W.; Wu, C. Ginger (Zingiber officinale Rosc.) and Its Bioactive Components Are Potential Resources for Health Beneficial Agents. Phytother. Res. 2021, 35, 711–742. [Google Scholar] [CrossRef]
- Ho, S.-C.; Chang, K.-S.; Lin, C.-C. Anti-Neuroinflammatory Capacity of Fresh Ginger Is Attributed Mainly to 10-Gingerol. Food Chem. 2013, 141, 3183–3191. [Google Scholar] [CrossRef] [PubMed]
- Kiyama, R. Nutritional Implications of Ginger: Chemistry, Biological Activities and Signaling Pathways. J. Nutr. Biochem. 2020, 86, 108486. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, M.-S.; Jung, S.; Son, H.-Y.; Park, S.; Kang, B.; Kim, S.-Y.; Kim, I.-H.; Kim, C.-T.; Kim, Y. Ginger Extract Ameliorates Obesity and Inflammation via Regulating MicroRNA-21/132 Expression and AMPK Activation in White Adipose Tissue. Nutrients 2018, 10, 1567. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Lv, Y.; Cao, B.; Chen, Z.; Xu, K. Physiological and Molecular Mechanism of Ginger (Zingiber officinale Roscoe) Seedling Response to Salt Stress. Front. Plant Sci. 2023, 14, 1073434. [Google Scholar] [CrossRef]
- Chi, H. Immunometabolism at the Intersection of Metabolic Signaling, Cell Fate, and Systems Immunology. Cell Mol. Immunol. 2022, 19, 299–302. [Google Scholar] [CrossRef]
- Wu, J. Adipogenesis: One Process with Two Stages. Chin. Sci. Bull. 2011, 56, 1327–1334. [Google Scholar] [CrossRef]
- Rani, M.P.; Krishna, M.S.; Padmakumari, K.P.; Raghu, K.G.; Sundaresan, A. Zingiber officinale Extract Exhibits Antidiabetic Potential via Modulating Glucose Uptake, Protein Glycation and Inhibiting Adipocyte Differentiation: An In Vitro Study. J. Sci. Food Agric. 2012, 92, 1948–1955. [Google Scholar] [CrossRef]
- Lee, G.-H.; Peng, C.; Jeong, S.-Y.; Park, S.-A.; Lee, H.-Y.; Hoang, T.-H.; Kim, J.; Chae, H.-J. Ginger Extract Controls mTOR-SREBP1-ER Stress-Mitochondria Dysfunction through AMPK Activation in Obesity Model. J. Funct. Foods 2021, 87, 104628. [Google Scholar] [CrossRef]
- Gembe-Olivarez, G.; Preciado-Ortiz, M.E.; Campos-Perez, W.; Rodríguez-Reyes, S.C.; Martínez-López, E.; Rivera-Valdés, J.J. A Mix of Ginger Phenols Exhibits Anti-adipogenic and Lipolytic Effects in Mature Adipocytes Derived from 3T3-L1 Cells. Exp. Ther. Med. 2023, 26, 336. [Google Scholar] [CrossRef]
- Tzeng, T.-F.; Liu, I.-M. 6-Gingerol Prevents Adipogenesis and the Accumulation of Cytoplasmic Lipid Droplets in 3T3-L1 Cells. Phytomedicine 2013, 20, 481–487. [Google Scholar] [CrossRef]
- Li, C.; Zhou, L. Inhibitory Effect 6-Gingerol on Adipogenesis through Activation of the Wnt/β-Catenin Signaling Pathway in 3T3-L1 Adipocytes. Toxicol. In Vitro 2015, 30, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Ahn, E.-K.; Oh, J.S. Inhibitory Effect of Galanolactone Isolated from Zingiber officinale Roscoe Extract on Adipogenesis in 3T3-L1 Cells. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 63–68. [Google Scholar] [CrossRef]
- Suk, S.; Kwon, G.T.; Lee, E.; Jang, W.J.; Yang, H.; Kim, J.H.; Thimmegowda, N.R.; Chung, M.-Y.; Kwon, J.Y.; Yang, S.; et al. Gingerenone A, a Polyphenol Present in Ginger, Suppresses Obesity and Adipose Tissue Inflammation in High-Fat Diet-Fed Mice. Mol. Nutr. Food Res. 2017, 61, 1700139. [Google Scholar] [CrossRef] [PubMed]
- Suk, S.; Seo, S.G.; Yu, J.G.; Yang, H.; Jeong, E.; Jang, Y.J.; Yaghmoor, S.S.; Ahmed, Y.; Yousef, J.M.; Abualnaja, K.O.; et al. A Bioactive Constituent of Ginger, 6-Shogaol, Prevents Adipogenesis and Stimulates Lipolysis in 3T3-L1 Adipocytes. J. Food Biochem. 2016, 40, 84–90. [Google Scholar] [CrossRef]
- Preciado-Ortiz, M.E.; Martínez-López, E.; García-Iglesias, T.; Gembe-Olivarez, G.; Torres-Castillo, N.; Llamas-Covarrubias, I.M.; Rivera-Valdés, J.J. 10-Gingerol Reduces Cytoplasmic Lipid Droplets and Induces Lipolysis in 3T3-L1 Adipocytes. Adipocyte 2024, 13, 2411453. [Google Scholar] [CrossRef]
- Duffen, J.; Zhang, M.; Masek-Hammerman, K.; Nunez, A.; Brennan, A.; Jones, J.E.C.; Morin, J.; Nocka, K.; Kasaian, M. Modulation of the IL-33/IL-13 Axis in Obesity by IL-13Rα2. J. Immunol. 2018, 200, 1347–1359. [Google Scholar] [CrossRef]
- Lee, T.-Y.; Lee, K.-C.; Chen, S.-Y.; Chang, H.-H. 6-Gingerol Inhibits ROS and iNOS through the Suppression of PKC-Alpha and NF-kappaB Pathways in Lipopolysaccharide-Stimulated Mouse Macrophages. Biochem. Biophys. Res. Commun. 2009, 382, 134–139. [Google Scholar] [CrossRef]
- Dorrington, M.G.; Fraser, I.D.C. NF-κB Signaling in Macrophages: Dynamics, Crosstalk, and Signal Integration. Front. Immunol. 2019, 10, 705. [Google Scholar] [CrossRef]
- Ho, S.-C.; Chang, Y.-H. Comparison of Inhibitory Capacities of 6-, 8- and 10-Gingerols/Shogaols on the Canonical NLRP3 Inflammasome-Mediated IL-1β Secretion. Molecules 2018, 23, 466. [Google Scholar] [CrossRef]
- Lantz, R.C.; Chen, G.J.; Sarihan, M.; Sólyom, A.M.; Jolad, S.D.; Timmermann, B.N. The Effect of Extracts from Ginger Rhizome on Inflammatory Mediator Production. Phytomedicine 2007, 14, 123–128. [Google Scholar] [CrossRef]
- Brahma Naidu, P.; Uddandrao, V.V.S.; Ravindar Naik, R.; Suresh, P.; Meriga, B.; Begum, M.S.; Pandiyan, R.; Saravanan, G. Ameliorative Potential of Gingerol: Promising Modulation of Inflammatory Factors and Lipid Marker Enzymes Expressions in HFD Induced Obesity in Rats. Mol. Cell Endocrinol. 2016, 419, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Hong, K.H.; Um, M.Y.; Ahn, J.; Ha, T.Y. 6-Gingerol Ameliorates Adiposity and Inflammation in Adipose Tissue in High Fat Diet-Induced Obese Mice: Association with Regulating of Adipokines. Nutrients 2023, 15, 3457. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Xiong, X.; Zhou, Y.; Wu, F.; Shao, Q.; Dong, R.; Liu, Q.; Li, L.; Chen, G. 6-Gingerol Ameliorates Metabolic Disorders by Inhibiting Hypertrophy and Hyperplasia of Adipocytes in High-Fat-Diet Induced Obese Mice. Biomed. Pharmacother. 2022, 146, 112491. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, G.; Ponmurugan, P.; Deepa, M.A.; Senthilkumar, B. Anti-Obesity Action of Gingerol: Effect on Lipid Profile, Insulin, Leptin, Amylase and Lipase in Male Obese Rats Induced by a High-Fat Diet. J. Sci. Food Agric. 2014, 94, 2972–2977. [Google Scholar] [CrossRef]
- Wang, J.; Li, D.; Wang, P.; Hu, X.; Chen, F. Ginger Prevents Obesity through Regulation of Energy Metabolism and Activation of Browning in High-Fat Diet-Induced Obese Mice. J. Nutr. Biochem. 2019, 70, 105–115. [Google Scholar] [CrossRef]
- Sayed, S.; Ahmed, M.; El-Shehawi, A.; Alkafafy, M.; Al-Otaibi, S.; El-Sawy, H.; Farouk, S.; El-Shazly, S. Ginger Water Reduces Body Weight Gain and Improves Energy Expenditure in Rats. Foods 2020, 9, 38. [Google Scholar] [CrossRef]
- Oh, S.; Lee, M.-S.; Jung, S.; Kim, S.; Park, H.; Park, S.; Kim, S.-Y.; Kim, C.-T.; Jo, Y.-H.; Kim, I.-H.; et al. Ginger Extract Increases Muscle Mitochondrial Biogenesis and Serum HDL-Cholesterol Level in High-Fat Diet-Fed Rats. J. Funct. Foods 2017, 29, 193–200. [Google Scholar] [CrossRef]
- Oliveira, C.T.; Lacerda, D.R.; Zicker, M.C.; Martins, L.B.; Teixeira, M.M.; de Araujo, R.L.B.; Ferreira, A.V.M. Ginger (Zingiber officinale Rosc.) Ameliorated Metabolic and Inflammatory Dysfunction Induced by High-Refined Carbohydrate-Containing Diet in Mice. J. Med. Food 2019, 22, 38–45. [Google Scholar] [CrossRef]
- Kim, H.-J.; Kim, B.; Mun, E.-G.; Jeong, S.-Y.; Cha, Y.-S. The Antioxidant Activity of Steamed Ginger and Its Protective Effects on Obesity Induced by High-Fat Diet in C57BL/6J Mice. Nutr. Res. Pract. 2018, 12, 503–511. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Jaafar, H.Z.E.; Rahmat, A. Antioxidant Activities, Total Phenolics and Flavonoids Content in Two Varieties of Malaysia Young Ginger (Zingiber officinale Roscoe). Molecules 2010, 15, 4324–4333. [Google Scholar] [CrossRef]
- Zagórska, J.; Czernicka-Boś, L.; Kukula-Koch, W.; Iłowiecka, K.; Koch, W. Impact of Thermal Processing on the Selected Biological Activities of Ginger Rhizome—A Review. Molecules 2023, 28, 412. [Google Scholar] [CrossRef] [PubMed]
- Ginger: Usefulness and Safety. Available online: https://www.nccih.nih.gov/health/ginger (accessed on 2 July 2025).
- Vaes, L.P.; Chyka, P.A. Interactions of Warfarin with Garlic, Ginger, Ginkgo, or Ginseng: Nature of the Evidence. Ann. Pharmacother. 2000, 34, 1478–1482. [Google Scholar] [CrossRef] [PubMed]
- Puteri, M.U.; Afifah, N.; Mathriul, A.Q.; Wicaksono, F.M.; Sugiarti, M.T.; Izzatinisa, R.; Kato, M.; Saputri, F.C. Antiplatelet Effects of a Combination of Sappan Wood (Caesalpinia sappan L.) and Red Ginger (Zingiber officinale var. Rubrum) Extracts in a High-Fat Diet-Induced Rat Model. Adv. Pharmacol. Pharm. Sci. 2024, 2024, 5543717. [Google Scholar] [CrossRef] [PubMed]
- Husain, I.; Dale, O.R.; Idrisi, M.; Gurley, B.J.; Avula, B.; Katragunta, K.; Ali, Z.; Chittiboyina, A.; Noonan, G.; Khan, I.A.; et al. Evaluation of the Herb-Drug Interaction (HDI) Potential of Zingiber officinale and Its Major Phytoconstituents. J. Agric. Food Chem. 2023, 71, 7521–7534. [Google Scholar] [CrossRef]
- Talasaz, A.H.; McGonagle, B.; HajiQasemi, M.; Ghelichkhan, Z.A.; Sadeghipour, P.; Rashedi, S.; Cuker, A.; Lech, T.; Goldhaber, S.Z.; Jennings, D.L.; et al. Pharmacokinetic and Pharmacodynamic Interactions between Food or Herbal Products and Oral Anticoagulants: Evidence Review, Practical Recommendations, and Knowledge Gaps. Semin. Thromb. Hemost. 2025, 51, 560–571. [Google Scholar] [CrossRef]
- Atashak, S.; Peeri, M.; Azarbayjani, M.A.; Stannard, S.R.; Haghighi, M.M. Obesity-Related Cardiovascular Risk Factors after Long- Term Resistance Training and Ginger Supplementation. J. Sports Sci. Med. 2011, 10, 685–691. [Google Scholar]
- Kamari, N.; Moradinazar, M.; Qasemi, M.; Khosravy, T.; Samadi, M.; Abdolahzad, H. Combination of the Effect of Ginger and Anti-Inflammatory Diet on Children with Obesity with Nonalcoholic Fatty Liver Disease: A Randomized Clinical Trial. Food Sci. Nutr. 2023, 11, 1846–1859. [Google Scholar] [CrossRef]
- Arablou, T.; Aryaeian, N.; Valizadeh, M.; Sharifi, F.; Hosseini, A.; Djalali, M. The Effect of Ginger Consumption on Glycemic Status, Lipid Profile and Some Inflammatory Markers in Patients with Type 2 Diabetes Mellitus. Int. J. Food Sci. Nutr. 2014, 65, 515–520. [Google Scholar] [CrossRef]
- Ebrahimzadeh Attari, V.; Ostadrahimi, A.; Asghari Jafarabadi, M.; Mehralizadeh, S.; Mahluji, S. Changes of Serum Adipocytokines and Body Weight Following Zingiber officinale Supplementation in Obese Women: A RCT. Eur. J. Nutr. 2016, 55, 2129–2136. [Google Scholar] [CrossRef]
- Mozaffari-Khosravi, H.; Talaei, B.; Jalali, B.-A.; Najarzadeh, A.; Mozayan, M.R. The Effect of Ginger Powder Supplementation on Insulin Resistance and Glycemic Indices in Patients with Type 2 Diabetes: A Randomized, Double-Blind, Placebo-Controlled Trial. Complement. Ther. Med. 2014, 22, 9–16. [Google Scholar] [CrossRef]
- Shidfar, F.; Rajab, A.; Rahideh, T.; Khandouzi, N.; Hosseini, S.; Shidfar, S. The Effect of Ginger (Zingiber officinale) on Glycemic Markers in Patients with Type 2 Diabetes. J. Complement. Integr. Med. 2015, 12, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Engin, A. The Definition and Prevalence of Obesity and Metabolic Syndrome. Adv. Exp. Med. Biol. 2017, 960, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Rahimlou, M.; Yari, Z.; Rayyani, E.; Keshavarz, S.A.; Hosseini, S.; Morshedzadeh, N.; Hekmatdoost, A. Effects of Ginger Supplementation on Anthropometric, Glycemic and Metabolic Parameters in Subjects with Metabolic Syndrome: A Randomized, Double-Blind, Placebo-Controlled Study. J. Diabetes Metab. Disord. 2019, 18, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh-Navaei, R.; Roozbeh, F.; Saravi, M.; Pouramir, M.; Jalali, F.; Moghadamnia, A.A. Investigation of the Effect of Ginger on the Lipid Levels. A Double Blind Controlled Clinical Trial. Saudi Med. J. 2008, 29, 1280–1284. [Google Scholar]
- Prasoppokakorn, T. Applicability of Statins in Metabolic Dysfunction-Associated Steatotic Liver Disease (MASLD). Livers 2025, 5, 4. [Google Scholar] [CrossRef]
- Katsiki, N.; Perez-Martinez, P.; Anagnostis, P.; Mikhailidis, D.P.; Karagiannis, A. Is Nonalcoholic Fatty Liver Disease Indeed the Hepatic Manifestation of Metabolic Syndrome? Curr. Vasc. Pharmacol. 2018, 16, 219–227. [Google Scholar] [CrossRef]
- Karimi, N.; Roshan, V.D.; Bayatiyani, Z.F. Individually and Combined Water-Based Exercise With Ginger Supplement, on Systemic Inflammation and Metabolic Syndrome Indices, Among the Obese Women With Breast Neoplasms. Iran. J. Cancer Prev. 2015, 8, e3856. [Google Scholar] [CrossRef]
- Ayaz, A.; Roshan, V.D. Effects of 6-Weeks Water-Based Intermittent Exercise with and without Zingiber officinale on pro-Inflammatory Markers and Blood Lipids in Overweight Women with Breast Cancer. J. Appl. Pharm. Sci. 2012, 2, 218–224. [Google Scholar] [CrossRef]
- Mansour, M.S.; Ni, Y.M.; Roberts, A.L.; Kelleman, M.; Roychoudhury, A.; St-Onge, M.P. Ginger consumption enhances the thermic effect of food and promotes feelings of satiety without affecting metabolic and hormonal parameters in overweight men: A pilot study. Metabolism 2012, 61, 1347–1352. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mohammadzadeh Honarvar, N.; Zarezadeh, M.; Khorshidi, M.; Makhdoomi Arzati, M.; Yekaninejad, M.S.; Abdollahi, M.; Effatpanah, M.; Hashemi, R.; Saedisomeolia, A. The Effect of an Oral Ginger Supplementation on NF-κB Concentration in Peripheral Blood Mononuclear Cells and Anthropomorphic Data of Patients with Type 2 Diabetes: A Randomized Double-Blind, Placebo-Controlled Clinical Trial. Complement. Ther. Med. 2019, 42, 7–11. [Google Scholar] [CrossRef]
- Seo, S.H.; Fang, F.; Kang, I. Ginger (Zingiber officinale) Attenuates Obesity and Adipose Tissue Remodeling in High-Fat Diet-Fed C57BL/6 Mice. Int. J. Environ. Res. Public Health 2021, 18, 631. [Google Scholar] [CrossRef] [PubMed]
- Misawa, K.; Hashizume, K.; Yamamoto, M.; Minegishi, Y.; Hase, T.; Shimotoyodome, A. Ginger Extract Prevents High-Fat Diet-Induced Obesity in Mice via Activation of the Peroxisome Proliferator-Activated Receptor δ Pathway. J. Nutr. Biochem. 2015, 26, 1058–1067. [Google Scholar] [CrossRef] [PubMed]
- Madsen, M.S.; Siersbæk, R.; Boergesen, M.; Nielsen, R.; Mandrup, S. Peroxisome Proliferator-Activated Receptor γ and C/EBPα Synergistically Activate Key Metabolic Adipocyte Genes by Assisted Loading. Mol. Cell. Biol. 2014, 34, 939–954. [Google Scholar] [CrossRef] [PubMed]
- Kuri-Harcuch, W.; Velez-delValle, C.; Vazquez-Sandoval, A.; Hernández-Mosqueira, C.; Fernandez-Sanchez, V. A Cellular Perspective of Adipogenesis Transcriptional Regulation. J. Cell. Physiol. 2019, 234, 1111–1129. [Google Scholar] [CrossRef]
- Ferré, P.; Foufelle, F. SREBP-1c Transcription Factor and Lipid Homeostasis: Clinical Perspective. Horm. Res. 2007, 68, 72–82. [Google Scholar] [CrossRef]
- Chirala, S.S.; Wakil, S.J. Structure and Function of Animal Fatty Acid Synthase. Lipids 2004, 39, 1045–1053. [Google Scholar] [CrossRef]
- Kim, B.; Kim, H.-J.; Cha, Y.-S. The Protective Effects of Steamed Ginger on Adipogenesis in 3T3-L1 Cells and Adiposity in Diet-Induced Obese Mice. Nutr. Res. Pract. 2021, 15, 279–293. [Google Scholar] [CrossRef]
- Preciado-Ortiz, M.E.; Martinez-Lopez, E.; Rodriguez-Echevarría, R.; Perez-Robles, M.; Gembe-Olivarez, G.; Rivera-Valdés, J.J. 10-Gingerol, a Novel Ginger Compound, Exhibits Antiadipogenic Effects without Compromising Cell Viability in 3T3-L1 Cells. Biomed. Rep. 2023, 19, 105. [Google Scholar] [CrossRef]
- Liu, G.Y.; Sabatini, D.M. mTOR at the Nexus of Nutrition, Growth, Ageing and Disease. Nat. Rev. Mol. Cell Biol. 2020, 21, 183–203. [Google Scholar] [CrossRef]
- Choi, J.; Kim, K.-J.; Kim, B.-H.; Koh, E.-J.; Seo, M.-J.; Lee, B.-Y. 6-Gingerol Suppresses Adipocyte-Derived Mediators of Inflammation In Vitro and in High-Fat Diet-Induced Obese Zebra Fish. Planta Med. 2017, 83, 245–253. [Google Scholar] [CrossRef]
- Ducharme, N.A.; Bickel, P.E. Lipid Droplets in Lipogenesis and Lipolysis. Endocrinology 2008, 149, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Zechner, R.; Zimmermann, R.; Eichmann, T.O.; Kohlwein, S.D.; Haemmerle, G.; Lass, A.; Madeo, F. FAT SIGNALS—Lipases and Lipolysis in Lipid Metabolism and Signaling. Cell Metab. 2012, 15, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Duncan, R.E.; Ahmadian, M.; Jaworski, K.; Sarkadi-Nagy, E.; Sul, H.S. Regulation of Lipolysis in Adipocytes. Annu. Rev. Nutr. 2007, 27, 79–101. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Park, A.; Oh, K.-J.; Lee, S.C.; Kim, W.K.; Bae, K.-H. The Role of Adipose Tissue Mitochondria: Regulation of Mitochondrial Function for the Treatment of Metabolic Diseases. Int. J. Mol. Sci. 2019, 20, 4924. [Google Scholar] [CrossRef]
- Schlaepfer, I.R.; Joshi, M. CPT1A-Mediated Fat Oxidation, Mechanisms, and Therapeutic Potential. Endocrinology 2020, 161, bqz046. [Google Scholar] [CrossRef]
- Carrière, A.; Casteilla, L. Chapter 7—Role of Mitochondria in Adipose Tissues Metabolism and Plasticity. In Mitochondria in Obesity and Type 2 Diabetes; Morio, B., Pénicaud, L., Rigoulet, M., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 173–194. ISBN 978-0-12-811752-1. [Google Scholar]
- Garcia, D.; Shaw, R.J. AMPK: Mechanisms of Cellular Energy Sensing and Restoration of Metabolic Balance. Mol. Cell 2017, 66, 789–800. [Google Scholar] [CrossRef]
- Martin, S.S.; Qasim, A.; Reilly, M.P. Leptin Resistance: A Possible Interface of Inflammation and Metabolism in Obesity-Related Cardiovascular Disease. J. Am. Coll. Cardiol. 2008, 52, 1201–1210. [Google Scholar] [CrossRef]
- Preciado-Ortiz, M.E.; Martínez-López, E.; Pedraza-Chaverri, J.; Medina-Campos, O.N.; Rodríguez-Echevarría, R.; Reyes-Pérez, S.D.; Rivera-Valdés, J.J. 10-Gingerol Increases Antioxidant Enzymes and Attenuates Lipopolysaccharide-Induced Inflammation by Modulating Adipokines in 3T3-L1 Adipocytes. Antioxidants 2024, 13, 1093. [Google Scholar] [CrossRef]
- Bradley, J. TNF-Mediated Inflammatory Disease. J. Pathol. 2008, 214, 149–160. [Google Scholar] [CrossRef]
- Arndt, L.; Lindhorst, A.; Neugebauer, J.; Hoffmann, A.; Hobusch, C.; Alexaki, V.-I.; Ghosh, A.; Blüher, M.; Wolfrum, C.; Glaß, M.; et al. The Role of IL-13 and IL-4 in Adipose Tissue Fibrosis. Int. J. Mol. Sci. 2023, 24, 5672. [Google Scholar] [CrossRef]
- Shimobayashi, M.; Albert, V.; Woelnerhanssen, B.; Frei, I.C.; Weissenberger, D.; Meyer-Gerspach, A.C.; Clement, N.; Moes, S.; Colombi, M.; Meier, J.A.; et al. Insulin Resistance Causes Inflammation in Adipose Tissue. J. Clin. Investig. 2018, 128, 1538–1550. [Google Scholar] [CrossRef] [PubMed]
- Sartipy, P.; Loskutoff, D.J. Monocyte Chemoattractant Protein 1 in Obesity and Insulin Resistance. Proc. Natl. Acad. Sci. USA 2003, 100, 7265–7270. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Hwang, S.J.; Bae, Y.C.; Jung, J.S. MiR-21 Regulates Adipogenic Differentiation through the Modulation of TGF-Beta Signaling in Mesenchymal Stem Cells Derived from Human Adipose Tissue. Stem Cells 2009, 27, 3093–3102. [Google Scholar] [CrossRef]
- Talotta, F.; Cimmino, A.; Matarazzo, M.R.; Casalino, L.; De Vita, G.; D’Esposito, M.; Di Lauro, R.; Verde, P. An Autoregulatory Loop Mediated by miR-21 and PDCD4 Controls the AP-1 Activity in RAS Transformation. Oncogene 2009, 28, 73–84. [Google Scholar] [CrossRef]
- Krichevsky, A.M.; Gabriely, G. miR-21: A Small Multi-Faceted RNA. J. Cell Mol. Med. 2009, 13, 39–53. [Google Scholar] [CrossRef]
- Strum, J.C.; Johnson, J.H.; Ward, J.; Xie, H.; Feild, J.; Hester, A.; Alford, A.; Waters, K.M. MicroRNA 132 Regulates Nutritional Stress-Induced Chemokine Production through Repression of SirT1. Mol. Endocrinol. 2009, 23, 1876–1884. [Google Scholar] [CrossRef]
- Wang, J.; Wang, P.; Li, D.; Hu, X.; Chen, F. Beneficial Effects of Ginger on Prevention of Obesity through Modulation of Gut Microbiota in Mice. Eur. J. Nutr. 2020, 59, 699–718. [Google Scholar] [CrossRef]
- Alhamoud, Y.; Ijaz Ahmad, M.; Abudumijiti, T.; Wu, J.; Zhao, M.; Feng, F.; Wang, J. 6-Gingerol, an Active Ingredient of Ginger, Reshapes Gut Microbiota and Serum Metabolites in HFD-Induced Obese Mice. J. Funct. Foods 2023, 109, 105783. [Google Scholar] [CrossRef]
- Hall, A.B.; Yassour, M.; Sauk, J.; Garner, A.; Jiang, X.; Arthur, T.; Lagoudas, G.K.; Vatanen, T.; Fornelos, N.; Wilson, R.; et al. A Novel Ruminococcus gnavus Clade Enriched in Inflammatory Bowel Disease Patients. Genome Med. 2017, 9, 103. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, D.; Jiang, H.; Zhang, S.; Pang, X.; Gao, S.; Zhang, H.; Zhang, S.; Xiao, Q.; Chen, L.; et al. Gut Microbiota Variation With Short-Term Intake of Ginger Juice on Human Health. Front. Microbiol. 2021, 11, 576061. [Google Scholar] [CrossRef] [PubMed]
- Crichton, M.; Marshall, S.; Marx, W.; Isenring, E.; Vázquez-Campos, X.; Dawson, S.L.; Lohning, A. Effect of Ginger Root Powder on Gastrointestinal Bacteria Composition, Gastrointestinal Symptoms, Mental Health, Fatigue, and Quality of Life: A Double-Blind Placebo-Controlled Trial. J. Nutr. 2023, 153, 3193–3206. [Google Scholar] [CrossRef] [PubMed]
- Shirdel, Z.; Mirbadalzadeh, H.; Madani, H. Anti-Diabetic and Anti Lipidemic Properties of Ginger in Comparison Glibenclamide in Alloxan-Diabetes Rat. Iran. J. Diabetes Lipid Disord. 2009, 9, 7–15. [Google Scholar]
- Sharma, I.; Gusain, D.; Dixit, V.P. Hypolipidaemic and Antiatherosclerotic Effects of Plumbagin in Rabbits. Indian J. Physiol. Pharmacol. 1991, 35, 10–14. [Google Scholar]
- Nammi, S.; Sreemantula, S.; Roufogalis, B.D. Protective Effects of Ethanolic Extract of Zingiber officinale Rhizome on the Development of Metabolic Syndrome in High-Fat Diet-Fed Rats. Basic Clin. Pharmacol. Toxicol. 2009, 104, 366–373. [Google Scholar] [CrossRef]
- Gao, H.; Guan, T.; Li, C.; Zuo, G.; Yamahara, J.; Wang, J.; Li, Y. Treatment with Ginger Ameliorates Fructose-Induced Fatty Liver and Hypertriglyceridemia in Rats: Modulation of the Hepatic Carbohydrate Response Element-Binding Protein-Mediated Pathway. Evid.-Based Complement. Altern. Med. 2012, 2012, 570948. [Google Scholar] [CrossRef]
- Li, Y.; Tran, V.H.; Duke, C.C.; Roufogalis, B.D. Preventive and Protective Properties of Zingiber officinale (Ginger) in Diabetes Mellitus, Diabetic Complications, and Associated Lipid and Other Metabolic Disorders: A Brief Review. Evid.-Based Complement. Altern. Med. 2012, 2012, 516870. [Google Scholar] [CrossRef]
- Xu, X.; So, J.-S.; Park, J.-G.; Lee, A.-H. Transcriptional Control of Hepatic Lipid Metabolism by SREBP and ChREBP. Semin. Liver Dis. 2013, 33, 301–311. [Google Scholar] [CrossRef]
- Freeman, A.M.; Acevedo, L.A.; Pennings, N. Insulin Resistance. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2025. [Google Scholar]
- Isa, Y.; Miyakawa, Y.; Yanagisawa, M.; Goto, T.; Kang, M.-S.; Kawada, T.; Morimitsu, Y.; Kubota, K.; Tsuda, T. 6-Shogaol and 6-Gingerol, the Pungent of Ginger, Inhibit TNF-Alpha Mediated Downregulation of Adiponectin Expression via Different Mechanisms in 3T3-L1 Adipocytes. Biochem. Biophys. Res. Commun. 2008, 373, 429–434. [Google Scholar] [CrossRef]
- World Health Organization. Use of Glycated Haemoglobin (HbA1c) in the Diagnosis of Diabetes Mellitus: Abbreviated Report of a WHO Consultation; WHO Guidelines Approved by the Guidelines Review Committee; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Huang, F.-Y.; Deng, T.; Meng, L.-X.; Ma, X.-L. Dietary Ginger as a Traditional Therapy for Blood Sugar Control in Patients with Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis. Medicine 2019, 98, e15054. [Google Scholar] [CrossRef]
- Purohit, V.; Wagner, A.; Yosef, N.; Kuchroo, V.K. Systems-based approaches to study immunometabolism. Cell. Mol. Immunol. 2022, 19, 409–420. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Preciado-Ortiz, M.E.; Gembe-Olivarez, G.; Martínez-López, E.; Rivera-Valdés, J.J. Immunometabolic Effects of Ginger (Zingiber officinale Roscoe) Supplementation in Obesity: A Comprehensive Review. Molecules 2025, 30, 2933. https://doi.org/10.3390/molecules30142933
Preciado-Ortiz ME, Gembe-Olivarez G, Martínez-López E, Rivera-Valdés JJ. Immunometabolic Effects of Ginger (Zingiber officinale Roscoe) Supplementation in Obesity: A Comprehensive Review. Molecules. 2025; 30(14):2933. https://doi.org/10.3390/molecules30142933
Chicago/Turabian StylePreciado-Ortiz, María Elizabeth, Gildardo Gembe-Olivarez, Erika Martínez-López, and Juan José Rivera-Valdés. 2025. "Immunometabolic Effects of Ginger (Zingiber officinale Roscoe) Supplementation in Obesity: A Comprehensive Review" Molecules 30, no. 14: 2933. https://doi.org/10.3390/molecules30142933
APA StylePreciado-Ortiz, M. E., Gembe-Olivarez, G., Martínez-López, E., & Rivera-Valdés, J. J. (2025). Immunometabolic Effects of Ginger (Zingiber officinale Roscoe) Supplementation in Obesity: A Comprehensive Review. Molecules, 30(14), 2933. https://doi.org/10.3390/molecules30142933