

Hemp Seed Oil Inhibits the Adipogenicity of the Differentiation-Induced Human Mesenchymal Stem Cells through Suppressing the Cannabinoid Type 1 (CB1)


 , ,
, ,
Abstract
1. Introduction
2. Results
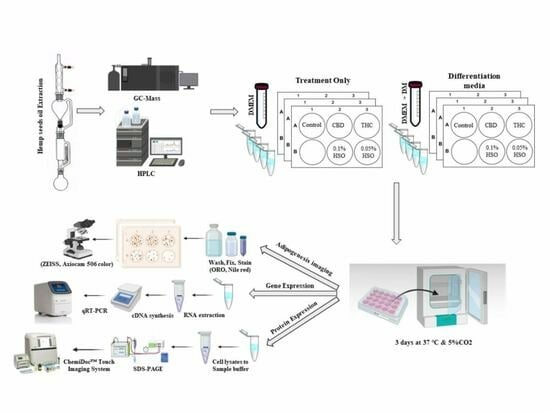
2.1. Soxhlet Extraction Method Produces More Oil and Cold-Pressed Hemp Seed Oil Has the Optimal Ratios to Carry out the Research
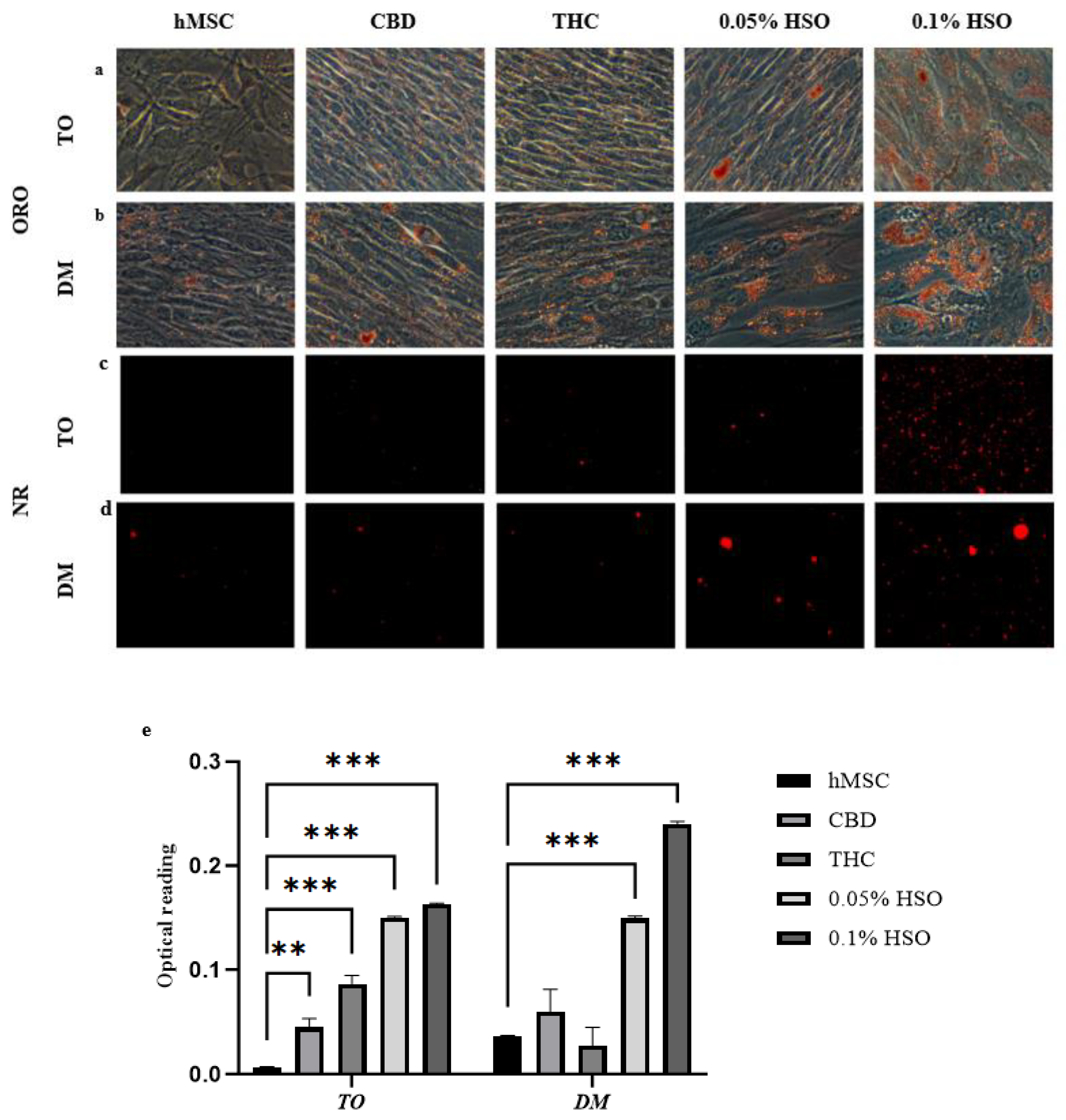
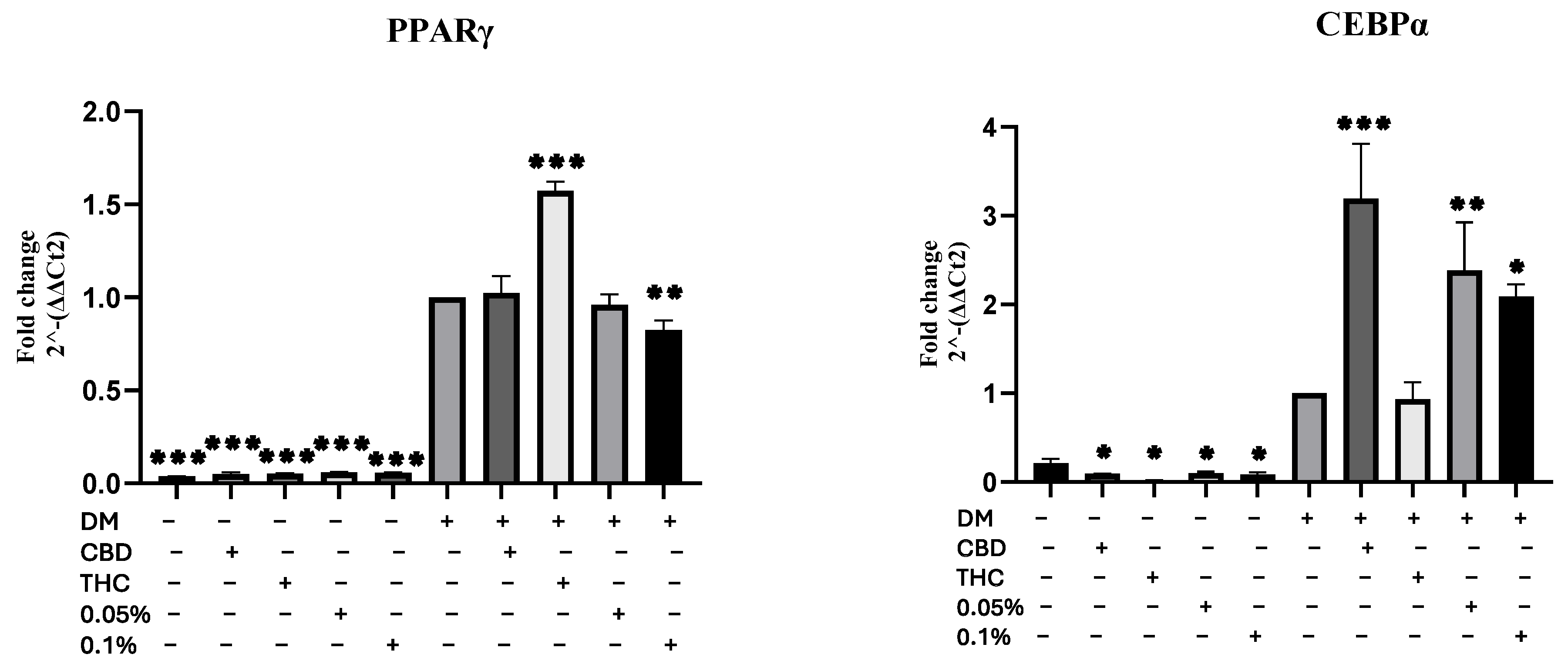
2.2. HSO, CBD, and THC Differentiate hMSC into Adipocyte
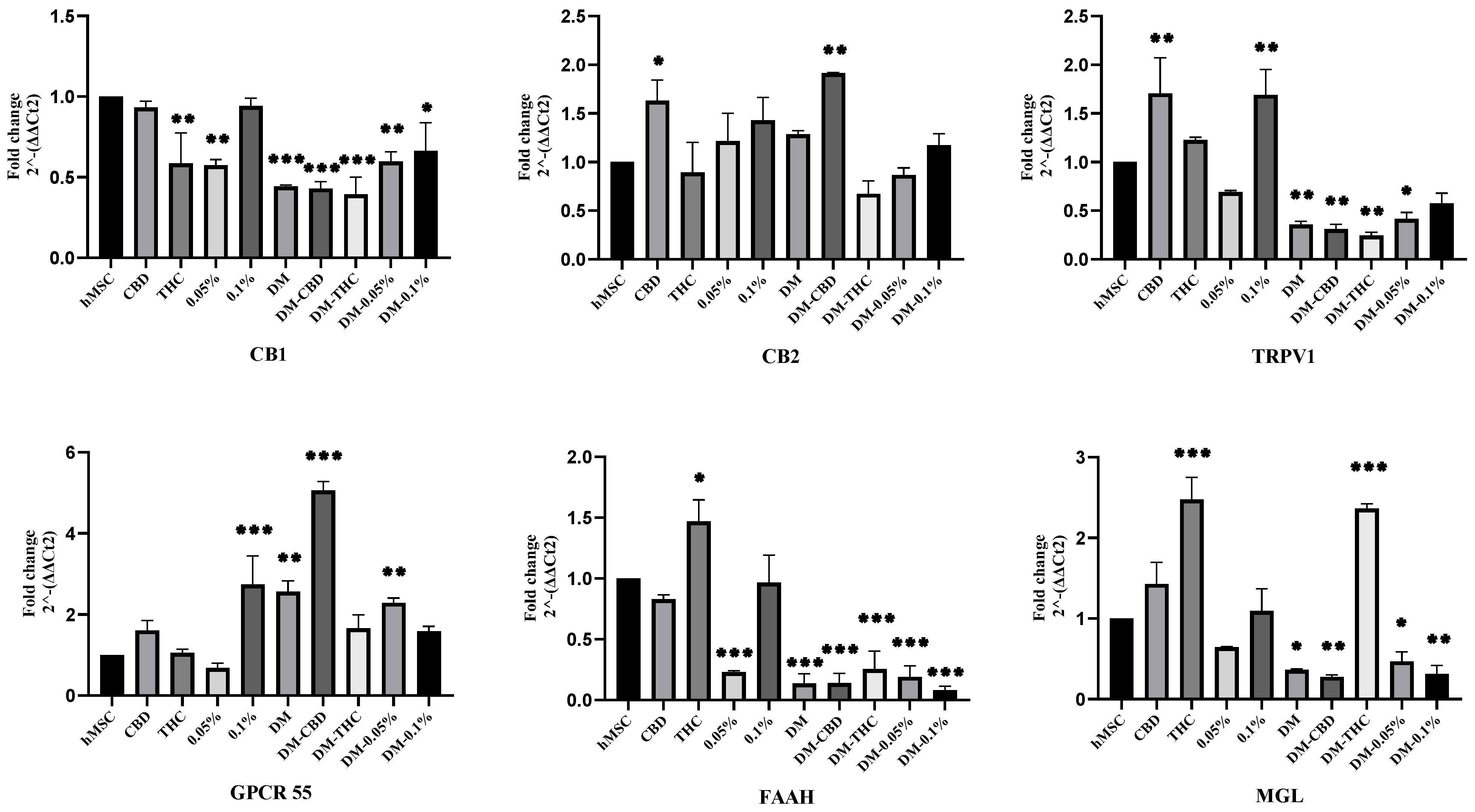
2.3. HSO, CBD, and THC Treatments Regulated the ECS
3. Discussion
4. Materials and Methods
4.1. Plant Products and Chemicals
4.2. Instrument
4.3. Extraction Method
4.4. Cell Culture
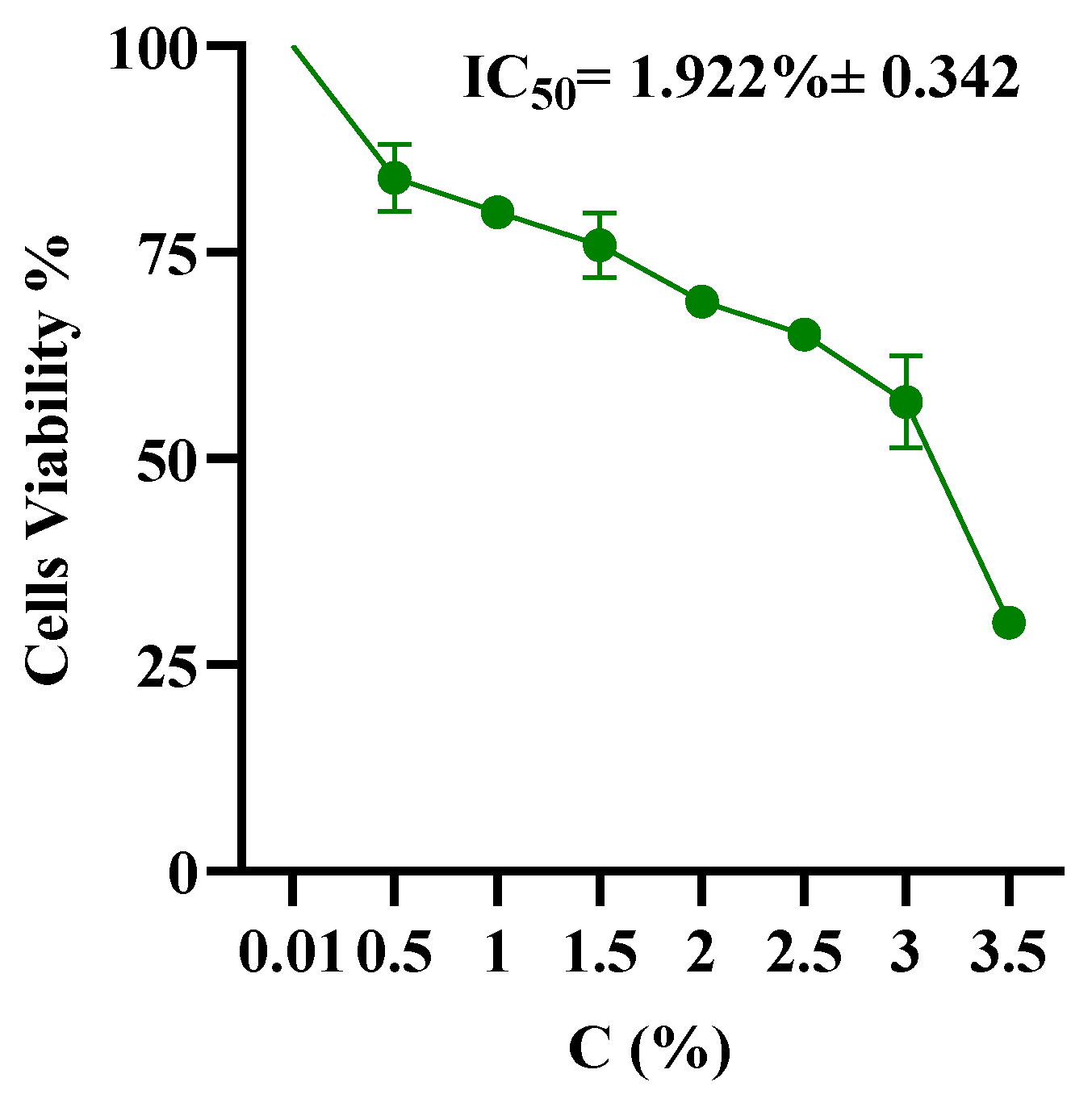
4.5. Cell Viability Assay
4.6. Oil Red O and Nile Red Staining
4.7. RNA Extraction and Real-Time Polymerase Chain Reaction (RT-PCR)
4.8. Immunoblot Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Matias, I.; Belluomo, I.; Cota, D. The fat side of the endocannabinoid system: Role of endocannabinoids in the adipocyte. Cannabis Cannabinoid Res. 2016, 1, 176–185. [Google Scholar] [CrossRef]
- Bermudez-Silva, F.J.; Viveros, M.P.; McPartland, J.M.; Rodriguez de Fonseca, F. The endocannabinoid system, eating behavior and energy homeostasis: The end or a new beginning? Pharmacol. Biochem. Behav. 2010, 95, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Silvestri, C.; Di Marzo, V. The endocannabinoid system in energy homeostasis and the etiopathology of metabolic disorders. Cell Metab. 2013, 17, 475–490. [Google Scholar] [CrossRef] [PubMed]
- Tudurí, E.; Imbernon, M.; Hernández-Bautista, R.J.; Tojo, M.; Fernø, J.; Diéguez, C.; Nogueiras, R. GPR55: A new promising target for metabolism? J. Mol. Endocrinol. 2017, 58, R191–R202. [Google Scholar] [CrossRef]
- Bennetzen, M.F.; Wellner, N.; Ahmed, S.S.; Ahmed, S.M.; Diep, T.A.; Hansen, H.S.; Richelsen, B.; Pedersen, S.B. Investigations of the human endocannabinoid system in two subcutaneous adipose tissue depots in lean subjects and in obese subjects before and after weight loss. Int. J. Obes. 2011, 35, 1377–1384. [Google Scholar] [CrossRef]
- De Petrocellis, L.; Ligresti, A.; Moriello, A.S.; Allarà, M.; Bisogno, T.; Petrosino, S.; Stott, C.G.; Di Marzo, V. Effects of cannabinoids and cannabinoid-enriched Cannabis extracts on TRP channels and endocannabinoid metabolic enzymes. Br. J. Pharmacol. 2011, 163, 1479–1494. [Google Scholar] [CrossRef] [PubMed]
- Batetta, B.; Griinari, M.; Carta, G.; Murru, E.; Ligresti, A.; Cordeddu, L.; Giordano, E.; Sanna, F.; Bisogno, T.; Uda, S.; et al. Endocannabinoids may mediate the ability of (n-3) fatty acids to reduce ectopic fat and inflammatory mediators in obese Zucker rats. J. Nutr. 2009, 139, 1495–1501. [Google Scholar] [CrossRef] [PubMed]
- Callaway, J.C. Hempseed as a nutritional resource: An overview. Euphytica 2004, 140, 65–72. [Google Scholar] [CrossRef]
- NHA. Hemp Resources—Hemp Facts and Statistics. The National Hemp Association. Available online: https://nationalhempassociation.org/hemp-resources/#Hemp%20Facts%20and%20Statistics (accessed on 8 February 2024).
- Subash-Babu, P.; Alshatwi, A.A. Aloe-emodin inhibits adipocyte differentiation and maturation during in vitro human mesenchymal stem cell adipogenesis. J. Biochem. Mol. Toxicol. 2012, 26, 291–300. [Google Scholar] [CrossRef]
- Mead, A. The legal status of cannabis (marijuana) and cannabidiol (CBD) under U.S. law. Epilepsy Behav. 2017, 70, 288–291. [Google Scholar] [CrossRef]
- Greenbaum, D.; Colangelo, C.; Williams, K.; Gerstein, M. Comparing protein abundance and mRNA expression levels on a genomic scale. Genome Biol. 2003, 4, 117. [Google Scholar] [CrossRef] [PubMed]
- Villarroel-Vicente, C.; Gutiérrez-Palomo, S.; Ferri, J.; Cortes, D.; Cabedo, N. Natural products and analogs as preventive agents for metabolic syndrome via peroxisome proliferator-activated receptors: An overview. Eur. J. Med. Chem. 2021, 221, 113535. [Google Scholar] [CrossRef] [PubMed]
- Palomares, B.; Ruiz-Pino, F.; Garrido-Rodriguez, M.; Eugenia Prados, M.; Sánchez-Garrido, M.A.; Velasco, I.; Vazquez, M.J.; Nadal, X.; Ferreiro-Vera, C.; Morrugares, R.; et al. Tetrahydrocannabinolic acid A (THCA-A) reduces adiposity and prevents metabolic disease caused by diet-induced obesity. Biochem. Pharmacol. 2020, 171, 113693. [Google Scholar] [CrossRef] [PubMed]
- Cardenia, V.; Toschi, T.G.; Scappini, S.; Rubino, R.C.; Rodriguez-Estrada, M.T. Development and validation of a Fast gas chromatography/mass spectrometry method for the determination of cannabinoids in Cannabis sativa L. J. Food Drug Anal. 2018, 26, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, M.; Marchei, E.; Pacifici, R.; Pichini, S. A rapid and simple procedure for the determination of cannabinoids in hemp food products by gas chromatography-mass spectrometry. J. Pharm. Biomed. Anal. 2005, 36, 939–946. [Google Scholar] [CrossRef]
- Oosterveer, M.H.; Koolman, A.H.; de Boer, P.T.; Bos, T.; Bleeker, A.; van Dijk, T.H.; Bloks, V.W.; Kuipers, F.; Sauer, P.J.; van Dijk, G. Resistance to diet-induced adiposity in cannabinoid receptor-1 deficient mice is not due to impaired adipocyte function. Nutr. Metab. 2011, 8, 93. [Google Scholar] [CrossRef]
- Chang, R.C.; Thangavelu, C.S.; Joloya, E.M.; Kuo, A.; Li, Z.; Blumberg, B. Cannabidiol promotes adipogenesis of human and mouse mesenchymal stem cells via PPARγ by inducing lipogenesis but not lipolysis. Biochem. Pharmacol. 2022, 197, 114910. [Google Scholar] [CrossRef]
- Roche, R.; Hoareau, L.; Bes-Houtmann, S.; Gonthier, M.-P.; Laborde, C.; Baron, J.-F.; Haffaf, Y.; Cesari, M.; Festy, F. Presence of the cannabinoid receptors, CB1 and CB2, in human omental and subcutaneous adipocytes. Histochem. Cell Biol. 2006, 126, 177–187. [Google Scholar] [CrossRef]
- Tedesco, L.; Valerio, A.; Cervino, C.; Cardile, A.; Pagano, C.; Vettor, R.; Pasquali, R.; Carruba, M.O.; Marsicano, G.; Lutz, B.; et al. Cannabinoid type 1 receptor blockade promotes mitochondrial biogenesis through endothelial nitric oxide synthase expression in white adipocytes. Diabetes 2008, 57, 2028–2036. [Google Scholar] [CrossRef]
- Bajzer, M.; Olivieri, M.; Haas, M.K.; Pfluger, P.T.; Magrisso, I.J.; Foster, M.T.; Tschöp, M.H.; Krawczewski-Carhuatanta, K.A.; Cota, D.; Obici, S. Cannabinoid receptor 1 (CB1) antagonism enhances glucose utilisation and activates brown adipose tissue in diet-induced obese mice. Diabetologia 2011, 54, 3121–3131. [Google Scholar] [CrossRef]
- Rakotoarivelo, V.; Sihag, J.; Flamand, N. Role of the endocannabinoid system in the adipose tissue with focus on energy metabolism. Cells 2021, 10, 1279. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Julius, D. The vanilloid receptor: A molecular gateway to the pain pathway. Annu. Rev. Neurosci. 2001, 24, 487–517. [Google Scholar] [CrossRef] [PubMed]
- Premkumar, L.S.; Sikand, P. TRPV1: A target for next generation analgesics. Curr. Neuropharmacol. 2008, 6, 151–163. [Google Scholar] [CrossRef]
- Christie, S.; Wittert, G.A.; Li, H.; Page, A.J. Involvement of TRPV1 channels in energy homeostasis. Front. Endocrinol. 2018, 9, 420. [Google Scholar] [CrossRef] [PubMed]
- Hermann, H.; De Petrocellis, L.; Bisogno, T.; Schiano Moriello, A.; Lutz, B.; Di Marzo, V. Dual effect of cannabinoid CB 1 receptor stimulation on a vanilloid VR1 receptor-mediated response. Cell. Mol. Life Sci. 2003, 60, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, L.A.; Lolait, S.J.; Brownstein, M.J.; Young, A.C.; Bonner, T.I. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature 1990, 346, 561–564. [Google Scholar] [CrossRef] [PubMed]
- Premkumar, L.S.; Ahern, G.P. Induction of vanilloid receptor channel activity by protein kinase C. Nature 2000, 408, 985–990. [Google Scholar] [CrossRef] [PubMed]
- Vellani, V.; Mapplebeck, S.; Moriondo, A.; Davis, J.B.; McNaughton, P.A. Protein kinase C activation potentiates gating of the vanilloid receptor VR1 by capsaicin, protons, heat and anandamide. J. Physiol. 2001, 534, 813–825. [Google Scholar] [CrossRef]
- Mahmud, A.; Santha, P.; Paule, C.C.; Nagy, I. Cannabinoid 1 receptor activation inhibits transient receptor potential vanilloid type 1 receptor-mediated cationic influx into rat cultured primary sensory neurons. Neuroscience 2009, 162, 1202–1211. [Google Scholar] [CrossRef]
- Sántha, P.; Jenes, Á.; Somogyi, C.; Nagy, I. The endogenous cannabinoid anandamide inhibits transient receptor potential vanilloid type 1 receptor-mediated currents in rat cultured primary sensory neurons. Acta Physiol. Hung. 2010, 97, 149–158. [Google Scholar] [CrossRef]
- Della Pietra, A.; Giniatullin, R.; Savinainen, J.R. Distinct activity of endocannabinoid-hydrolyzing enzymes MAGL and FAAH in key regions of peripheral and central nervous system implicated in migraine. Int. J. Mol. Sci. 2021, 22, 1204. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V.; Maccarrone, M. FAAH and anandamide: Is 2-AG really the odd one out? Trends Pharmacol. Sci. 2008, 29, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Matias, I.; Carta, G.; Murru, E.; Petrosino, S.; Banni, S.; Marzo, V.D. Effect of polyunsaturated fatty acids on endocannabinoid and N-acyl-ethanolamine levels in mouse adipocytes. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2008, 1781, 52–60. [Google Scholar] [CrossRef]
- Koolman, A.H. The Role of the Endocannabinoid System in the Regulation of Energy Balance; University of Groningen: Groningen, The Netherlands, 2010. [Google Scholar]
- Moreno-Navarrete, J.M.; Catalán, V.; Whyte, L.; Díaz-Arteaga, A.; Vázquez-Martínez, R.; Rotellar, F.; Guzmán, R.; Gómez-Ambrosi, J.; Pulido, M.R.; Russell, W.R.; et al. The L-α-lysophosphatidylinositol/GPR55 system and its potential role in human obesity. Diabetes 2012, 61, 281–291. [Google Scholar] [CrossRef]
- Choi, J.W.; Lee, C.-W.; Chun, J. Biological roles of lysophospholipid receptors revealed by genetic null mice: An update. Biochim. Biophys. Acta 2008, 1781, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Brighenti, V.; Pellati, F.; Steinbach, M.; Maran, D.; Benvenuti, S. Development of a new extraction technique and HPLC method for the analysis of non-psychoactive cannabinoids in fibre-type Cannabis sativa L. (hemp). J. Pharm. Biomed. Anal. 2017, 143, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Devi, V.; Khanam, S. Comparative study of different extraction processes for hemp (Cannabis sativa) seed oil considering physical, chemical and industrial-scale economic aspects. J. Clean. Prod. 2019, 207, 645–657. [Google Scholar] [CrossRef]
- Mandrioli, M.; Tura, M.; Scotti, S.; Gallina Toschi, T. Fast detection of 10 cannabinoids by RP-HPLC-UV method in Cannabis sativa L. Molecules 2019, 24, 2113. [Google Scholar] [CrossRef]
- PromoCell. Preadipocyte/Adipocyte Media Instruction Manual. PromoCell. Available online: https://promocell.com/wp-content/uploads/product-information/manual/C-27436.pdf (accessed on 8 February 2024).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Seed | Extraction Method | Average (mL) | Average (mL) by Extraction Method |
|---|---|---|---|
| S | DyM | 2.4 ± 0.39 | DyM |
| Sox | 3.3 ± 0.50 | 3 mL | |
| CEM | 2.5 ± 0.25 | Sox | |
| NS | DyM | 3.6 ± 0.52 d | 3.8 mL |
| Sox | 4.3 ± 0.19 a,b,c | CEM | |
| CEM | 3 ± 0.12 | 2.75 mL |
| Oil Content | DyM | Sox | CEM | HSO | |||
|---|---|---|---|---|---|---|---|
| S | NS | S | NS | S | NS | Cold-Pressed | |
| ω3 | 30.65% | 14.46% | 28.78% | 34.08% * | 1.24% | 0.69% | 13.36% |
| ω6 | 41.98% | 20.4% | 64.74% * | 55.02% | 54.06% | 60.71% | 25.09% |
| Vitamin E | 0 | 0 | 0 | 0 | 0 | 2.65% * | 0 |
| Palmitic acid | 0 | 0 | 4.13% | 6.78% | 7.68% * | 5.71% | 5.02% |
| Stearic acid | 1.79% | 0 | 0.89% | 2.20% * | 0 | 0 | 1.87% |
| Pentadecanoic acid | 0 | 0 | 0 | 0 | 0 | 0 | 3.80% * |
| DyM | Sox | CEM | HSO | |||
|---|---|---|---|---|---|---|
| S | NS | S | NS | S | NS | Cold-Pressed |
| 0.131 | 0.141 | 0.162 | 0.144 | 0.101 | 0.109 | 0.327 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almousa, A.S.; Subash-Babu, P.; Alanazi, I.O.; Alshatwi, A.A.; Alkhalaf, H.; Bahattab, E.; Alsiyah, A.; Alzahrani, M. Hemp Seed Oil Inhibits the Adipogenicity of the Differentiation-Induced Human Mesenchymal Stem Cells through Suppressing the Cannabinoid Type 1 (CB1). Molecules 2024, 29, 1568. https://doi.org/10.3390/molecules29071568
Almousa AS, Subash-Babu P, Alanazi IO, Alshatwi AA, Alkhalaf H, Bahattab E, Alsiyah A, Alzahrani M. Hemp Seed Oil Inhibits the Adipogenicity of the Differentiation-Induced Human Mesenchymal Stem Cells through Suppressing the Cannabinoid Type 1 (CB1). Molecules. 2024; 29(7):1568. https://doi.org/10.3390/molecules29071568
Chicago/Turabian StyleAlmousa, Albatul S., Pandurangan Subash-Babu, Ibrahim O. Alanazi, Ali A. Alshatwi, Huda Alkhalaf, Eman Bahattab, Atheer Alsiyah, and Mohammad Alzahrani. 2024. "Hemp Seed Oil Inhibits the Adipogenicity of the Differentiation-Induced Human Mesenchymal Stem Cells through Suppressing the Cannabinoid Type 1 (CB1)" Molecules 29, no. 7: 1568. https://doi.org/10.3390/molecules29071568
APA StyleAlmousa, A. S., Subash-Babu, P., Alanazi, I. O., Alshatwi, A. A., Alkhalaf, H., Bahattab, E., Alsiyah, A., & Alzahrani, M. (2024). Hemp Seed Oil Inhibits the Adipogenicity of the Differentiation-Induced Human Mesenchymal Stem Cells through Suppressing the Cannabinoid Type 1 (CB1). Molecules, 29(7), 1568. https://doi.org/10.3390/molecules29071568