Abstract
Panax quinquefolius (PQ) has been widely used in traditional Chinese medicine and functional food. Ginsenosides are the important functional components of PQ. The ginsenosides’ diversity is deeply affected by the processing conditions. The ginsenosides in the steamed PQ have been not well-characterized yet because of the complexity of their structure. In the study, the comprehensive investigation of ginsenosides was performed on the steamed PQ with different steaming times and temperatures by UPLC-Q-TOF-MS. Based on the molecular weight, retention time and characterized fragment ions, 175 ginsenosides were unambiguously identified or tentatively characterized, including 45 protopanaxatriol type, 49 protopanaxadiol type, 19 octillol type, 6 oleanolic acid type ginsenosides, and 56 other ginsenosides. Ten new ginsenosides and three new aglycones were discovered in the steamed PQ samples through searching the database of CAS SciFindern. Principal component analysis showed the significant influence on the chemical components of PQ through different processing conditions. The steaming temperature was found to promote the transformation of ginsenosides more than the steaming time. The protoginsenosides were found to transform into the rare ginsenosides by elimination reactions. The malonyl ginsenosides were degraded into acetyl ginsenosides, and then degraded into neutral ginsenosides. The sugar chain experienced degradation, with position changes and configuration inversions. Furthermore, 20 (S/R)-ginsenoside Rh1, Rh2, Rg2, and Rh12 were found to transform from the S-configuration to the R-configuration significantly. This study could present a comprehensive ginsenosides profile of PQ with different steaming conditions, and provide technical support for the development and utilization of PQ.
1. Introduction
Panax quinquefolius (PQ) belongs to the genus Panax in the family Araliaceae. It has been widely used in traditional Chinese medicine, dietary supplements, and functional food [1,2]. It is native to the south of Canada and the northern USA, and has been widely planted in China. In vivo and in vitro studies have shown that PQ has many biological activities, including antioxidative, antidiabetic, anti-inflammatory, anti-cancer, etc., [3,4]. It has been demonstrated to have a positive impact on the treatment of various diseases such as in the central nervous system, endocrine system, cardiovascular system, as well as cancer [5]. In particular, PQ has unique advantages in treating chronic diseases because of its better compatibility with the human body and fewer side effects [3]. It was found that steamed PQ has been reported to exhibit enhanced antiproliferative activity and antioxidant capacity [6,7]. PQ contains many chemical components, including ginsenosides, polyacetylenes, polyphenolic compounds, etc. Ginsenosides are the most important active ingredients [8,9]. More than 500 ginsenosides have been discovered from Panax species [10], but only a few have been characterized in the steamed PQ samples. A comprehensive investigation of ginsenosides is essential for the further development and utilization of the steamed PQ.
Based on the structure difference of the aglycone, ginsenosides are divided into protopanaxadiol (PPD-type), protopanaxatriol (PPT-type), octillol (OT-type), oleanolic (OA-type), and other types [11]. OT-type ginsenosides are characteristic in the PQ, differing from other Panax species [12]. The ginsenosides Rb1, Rb2, Rc, Rd, Re, and Rg1 are generally considered as the main protoginsenosides. Protoginsenosides have a larger molecular structure that is difficult to be absorbed by the human body, whereas less-polar ginsenosides are easily absorbed by the intestinal microflora [13,14]. These ginsenosides are called rare ginsenosides, and they have been found to have special pharmacological activities [15]. Compared to ginsenoside Rb1, ginsenoside Rg5 has been demonstrated to have stronger antiproliferative activity against breast cancer [16,17]. Ginsenoside Rh4 has been reported to have anti-esophageal cancer effects through inhibiting aerobic glycolysis [18,19]. Rare ginsenosides can be prepared by physical, chemical, and biological methods [20]. The chemical method has usually the disadvantage of a long reaction time and the production of byproducts. The biotransformation of rare ginsenosides needs to solve the problems of low specific activities, unidentified enzymes, and uncovered catalytic mechanisms [21]. The physical method commonly refers to the steaming method. It is a green and efficient method to regulate the transformation of ginsenosides [22]. Ginsenosides undergo chemical modifications and generate rare ginsenosides during the steaming process [23]. There have been several reports on steamed PQ. Twelve ginsenosides were determined in the roots of steamed PQ, and the anticancer activities of the extract from roots steamed for 2 h were greater than 1 h [24]. Twenty-nine major ginsenosides have been studied in the multi-steamed PQ samples and a possible chemical conversion was deduced [25]. Fifty-nine ginsenosides of PPT, PPD, OA, and OT types have been analyzed in PQ with different steaming processes [26]. However, a few known ginsenosides cannot meet the needs of holistic studies or the discovery of chemical markers in the PQ steaming process [27]. Further studies of the steam processing mechanism will be of great significance for the rational utilization of PQ.
The development of analytical technologies has made it possible for an holistic study. LC-MS has been widely used in secondary metabolite analysis for its high sensitivity and high throughput, such as for flavonoids, phenolic acids, and ginsenosides [28,29]. In the study, ginsenosides profiling has been established for the steamed PQ samples based on LC-MS. The in-depth identification of PQ was performed, the aglycone and the sugar chains were annotated, and new ginsenosides were discovered. The structure and content changes of the ginsenosides were analyzed with the different steaming processes, and the transformation mechanisms of ginsenosides were further investigated. The study could present the comprehensive ginsenoside profiling of steamed PQ with different times and temperatures.
2. Results and Discussion
2.1. Identification of Ginsenosides in PQ Samples
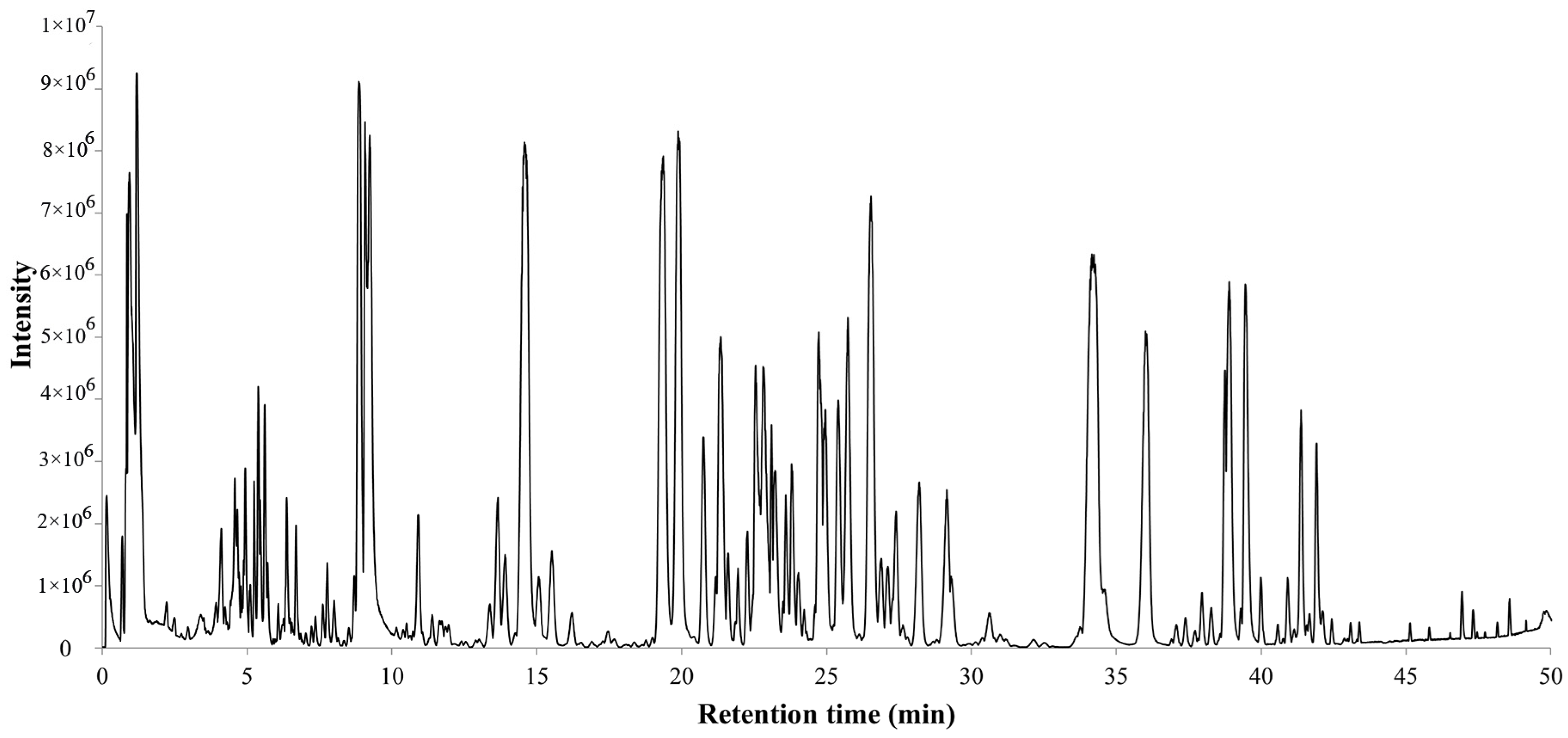
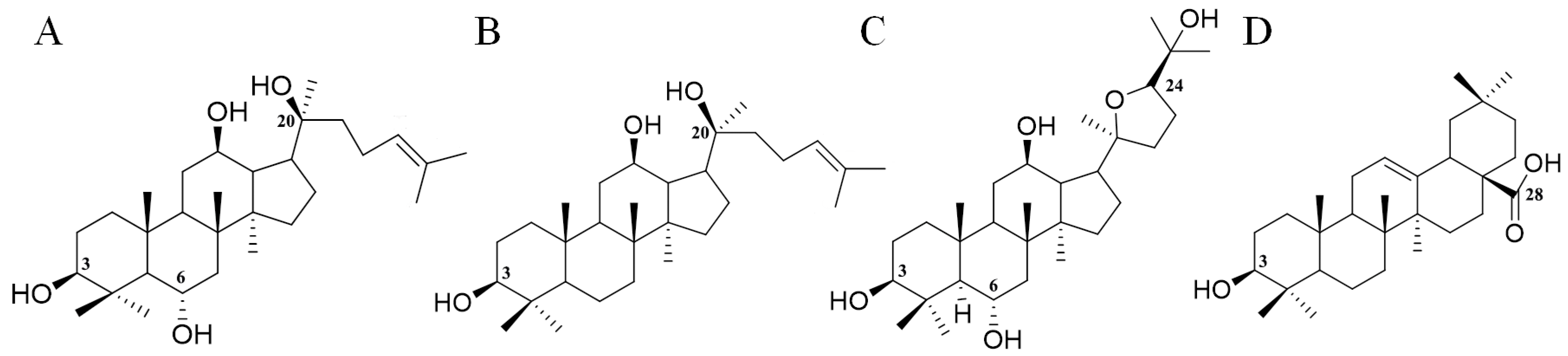
The total ion chromatogram of the PQ sample with LC-MS is shown in Figure 1. For ginsenosides, there were specific fragmentation rules in the MS/MS analysis [30]. The identification of ginsenosides was based on the accurate molecular weight, retention time, and MS/MS fragment ions. With different collision energies in the MS/MS analysis, ginsenosides were fragmented into aglycone and sugar chains. Characterized aglycone ions, sugar chain ions, and the neutral loss of sugar chains were found. For the PPT-, PPD-, OT-, or OA-type, the characterized aglycone ions were 475, 459, 491, and 455 (m/z), respectively. The chemical structures of the PPT-, PPD-, OT-, or OA-type aglycones are shown in Figure 2. The aglycone ions of other types of ginsenosides were complicated and varied. The characteristic ions for different sugar chains of O-glucose, O-rhamnose, O-xylose/arabinose, O-glucose-glucose, O-glucose-rhamnose, and O-glucose-xylose/arabinose were 161, 145, 131, 221/323, 205/307, and 191/293, respectively. In addition, acetyl and malonyl ginsenosides were characterized as having a neutral loss of 42 and 86. Then, a total of 175 ginsenosides were identified from the extracts of the PQ samples, which included 45 PPT-type, 49 PPD-type, 19 OT-type, 6 OA-type ginsenosides, and 56 other ginsenosides. The ginsenosides were then validated with the standards available. The detailed molecular weight, retention time, aglycone, sugar chains, and MS/MS fragment ions of the ginsenosides are listed in Table 1.
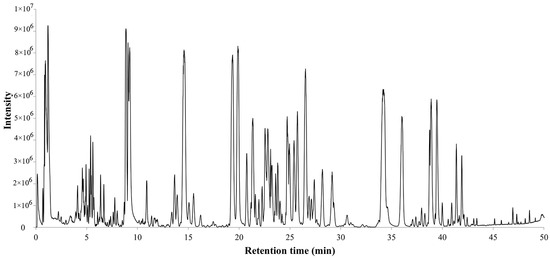
Figure 1.
Total ion chromatogram of steamed PQ with LC-MS in the negative ion mode.
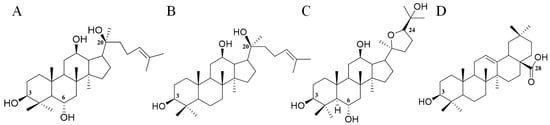
Figure 2.
The chemical structures of PPT- (A), PPD- (B), OT- (C), and OA- (D) type aglycones. Numbers represent the typical glycosylation sites.

Table 1.
Ginsenosides identified in the steamed PQ samples.
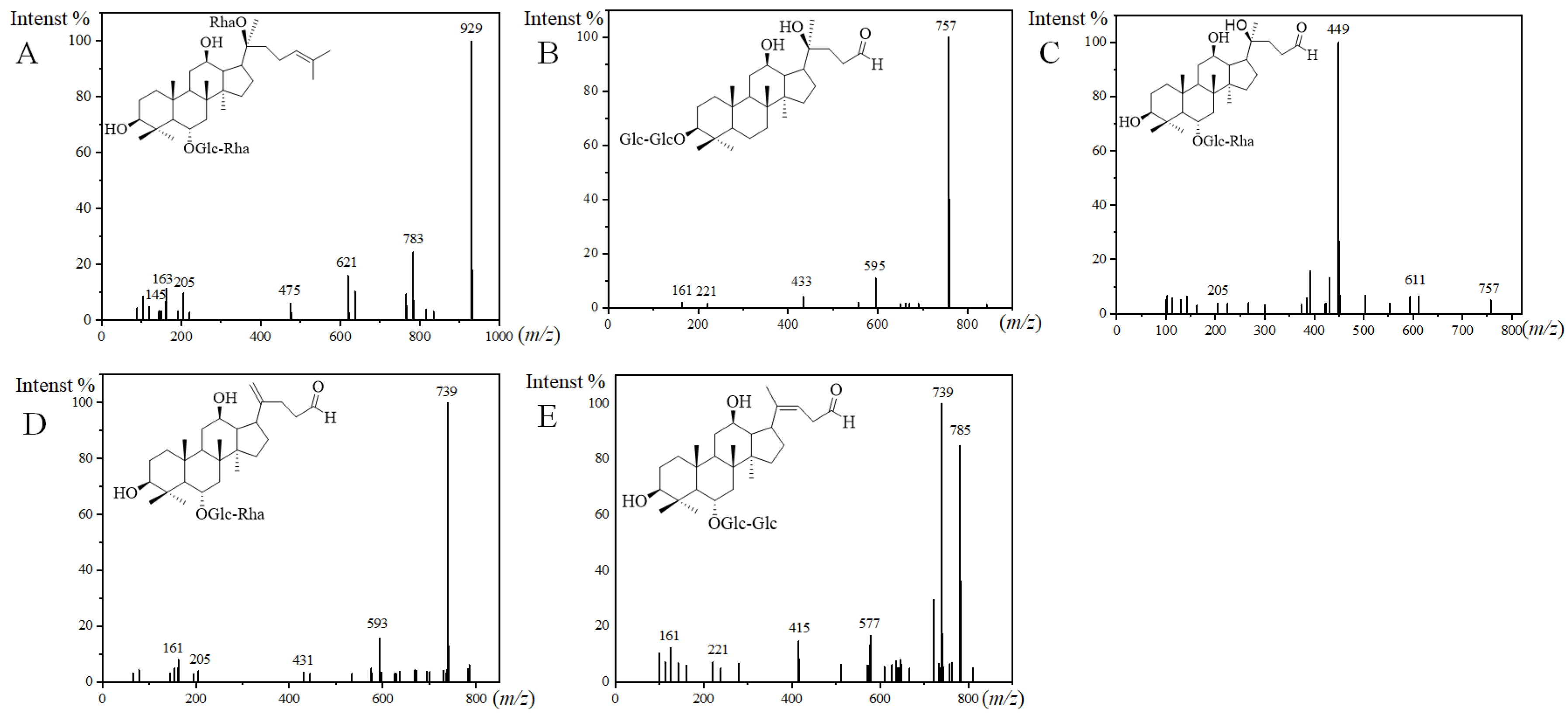
For the structures of the ginsenosides, the database CAS SciFindern was searched. Among them, 10 ginsenosides were identified in the steamed PQ for the first time. The aglycone or sugar chains were different compared with the ginsenosides reported previously. There was one new ginsenoside classified as the PPT-type, and it was named PPT-O-glc-rha/O-rha. PPT-O-glc-rha/O-rha (m/z 975.5534, [M+HCOO]−) was identified by fragment ions of 929, 783, 621, 475, 205, 163, 161, and 145 (Figure 3A). The fragment ion of 475 is the characteristic aglycone ion of PPT-type ginsenosides. The neutral loss of 929/783, 783/621, and 621/475 indicated two rhamnose and a glucose in the sugar chains. The fragment ion of 205 indicated a sugar chain of O-glucose-rhamnose.
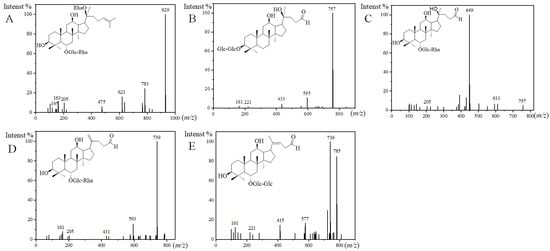
Figure 3.
MS2 spectra and the presumed structures of PPT-O-glc-rha/O-rha ((A), 50 eV), PQ-ginsenoside A ((B), 40 eV), PQ-ginsenoside B ((C), 70 eV), PQ-ginsenoside C ((D), 40 eV), and PQ-ginsenoside D ((E), 40 eV).
There were nine new ginsenosides classified as the other type, and they were named PQ-ginsenoside A, B, C, D, acetyl-PQ-ginsenoside A, and acetyl-PQ-ginsenoside D isomers. Three new aglycones were discovered in PQ-ginsenoside B, C, and D, and the m/z of them were 449, 431, and 415 in the MS/MS analysis. For example, the fragmentation rules of PQ-ginsenoside C were consistent with the general ginsenosides, and a new aglycone with m/z 431 was discovered. The m/z of the aglycone in PQ-ginsenoside A was 433. The aglycone with m/z 433 has been reported to be degraded from 459 in the PPD-type, and it was characterized as 25-, 26-, and 27-trinor-PPD-type in the floralginsenoside Kb from Panax ginseng [31,32]. The sugar chain of PQ-ginsenoside A was different from floralginsenoside Kb, and it was not found in the database of CAS SciFindern, so it was defined as a new ginsenoside. The sugar chains of PQ-ginsenoside A were the same as Rg3, and they were eluted at the same retention time. Rg3 is a PPD-type ginsenoside. Therefore, the aglycone structure in PQ-ginsenoside A were deduced as 25-, 26-, and 27-trinor-PPD-type degraded from Rg3 (Figure 3B). For PQ-ginsenoside B (aglycone, m/z, 449, Figure 3C), it has the same sugar chains and retention time with Rg2 (aglycone, m/z, 475). For PQ-ginsenoside C (aglycone, m/z, 431, Figure 3D), it has the same sugar chains and retention time with Rg6 (aglycone, m/z, 457). For PQ-ginsenoside D (aglycone, m/z, 415, Figure 3E) it has the same sugar chains and retention time with Rg5 (aglycone, m/z, 441). The molecular weight difference of 449/475, 431/457, and 415/441 were the same as the 433/459. Therefore, the aglycone structures of 449, 431, and 415 in PQ-ginsenoside B, C, and D were deduced to be degraded from 475, 457, and 441 in the Rg5, Rg6, and Rg5, characterized as 25-, 26-, and 27-trinor.
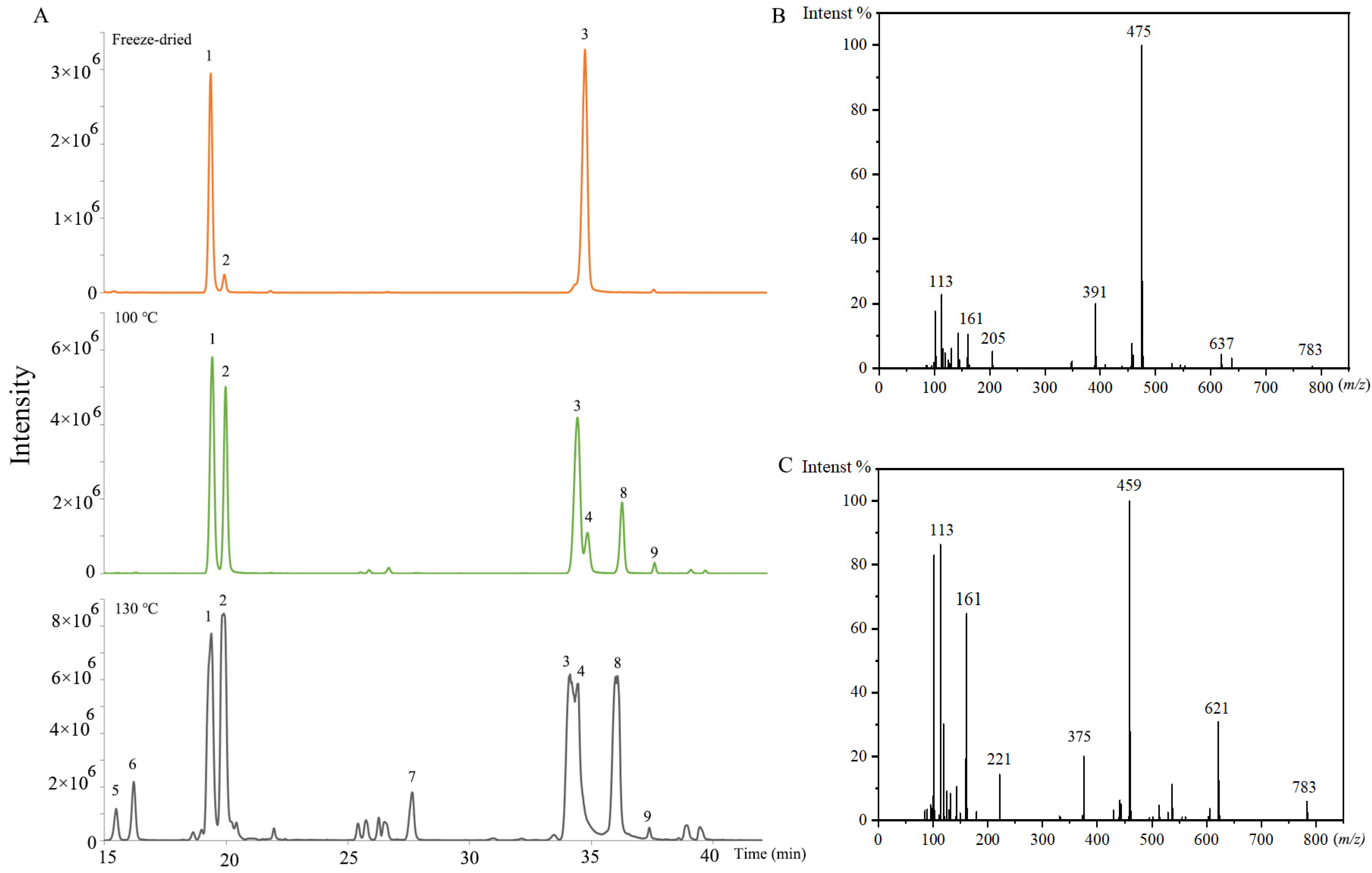
There were also many isomers identified. For the isomers, the fragmentation ions in the MS/MS analysis were the same. The aglycone and sugar chains of them were the same as the previous ginsenosides, but the attaching positions of the sugar chain and aglycone were different. The extracted ion chromatograms of 829.4943 (m/z) in the freeze-dried sample, the (100 °C, 2 h) sample, and the (130 °C, 2 h) sample are shown in Figure 4A. It was obvious that the ginsenosides contents and varieties were changed with the increase in steaming temperature. The ginsenoside Rg2 and Rg3 were PPT-type and PPD-type, respectively. The ginsenoside Rg2 isomers (No. 1, 2, 5, and 6) were identified by product ions of 783, 637, 475, 205, 161, and 145 (Figure 4B). The ginsenoside Rg3 isomers (No. 3, 4, 7, 8, and 9) were identified by product ions of 783, 621, 459, 221, and 161 (Figure 4C). The No. 1, 2, 3, and 4 were confirmed by standards as 20(S)-ginsenoside Rg2, 20(R)-ginsenoside Rg2, 20(S)-ginsenoside Rg3, and 20(R)-ginsenoside Rg3. For PPT-type ginsenosides, the glycosidic bonds were commonly at the aglycone C-6 and C-20 hydroxyl groups [33]. Ginsenoside Rg2 was C-6 linked. Therefore, No. 5 and 6 were deduced as sugar chains linking to the C-20 position, named as the ginsenoside Rg2 isomer. For PPD-type ginsenosides, the glycosidic bonds were commonly at the aglycone C-3 and C-20. Ginsenoside Rg3 was C-3 linked. Therefore, No. 7, 8, and 9 were speculated as sugar-chain-linking to the C-20 position, named the ginsenoside Rg3 isomers.
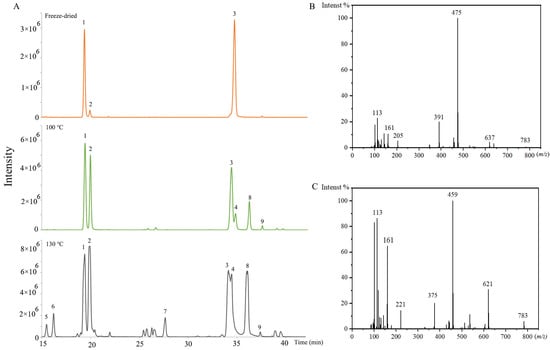
Figure 4.
(A) The extracted ion chromatogram of 829.4943 (m/z) in the freeze-dried sample, and steamed samples (100 °C, 2 h), and (130 °C, 2 h), No. 1, 2, 5, and 6 represent the ginsenoside Rg2 and its isomers, No. 3, 4, 7, 8, and 9 represent the ginsenoside Rg3 and its isomers. (B) MS2 spectrum of 20(S)-Ginsenoside Rg2. (C) MS2 spectrum of 20(S)-Ginsenoside Rg3 (70 eV).
2.2. Method Validation
The repeatability and precision of the analytical method were investigated by QC samples. The repeatability was investigated by six QC samples. The QC samples were analyzed continuously. The RSD of each peak was calculated among the six QC samples. The peak number and area were counted within different RSD ranges (0–10%, 10–20%, 20–30%, and >30%). In total, 98.2% of the peaks had an RSD value of less than 20%, while the accumulated peak area accounted for 99.3% of the total area (Figure S1A). For the intra-day precision analysis, six QC samples were analyzed every 4 h, and 98.1% of the peaks had an RSD less than 20%, while the accumulated peak area accounted for 99.3% of the total peak area (Figure S1B). For the inter-day precision analysis, 18 QC samples were analyzed for 3 days. The results show that 96.0% of the peaks had an RSD of less than 20%, while the accumulated peak area accounted for 98.6% of the total peak area (Figure S1C). These results indicated the good stability of the analytical method.
2.3. Difference between Steamed and Freeze-Dried Samples
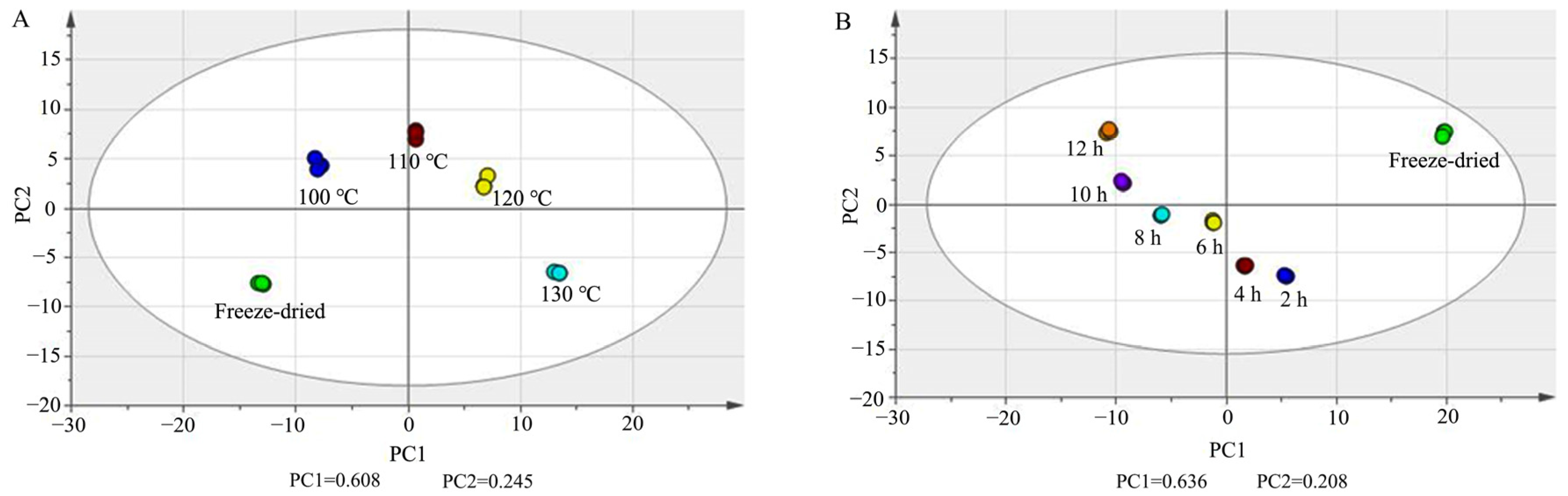
Principal component analysis (PCA) was carried out in PQ samples with different steaming times and temperatures with UV scaling. In Figure 5A, PC1 and PC2 were 0.608 and 0.245. In Figure 5B, PC1 and PC2 were 0.636 and 0.208. The score plots show that there was obvious separation among the freeze-dried samples and steamed samples.
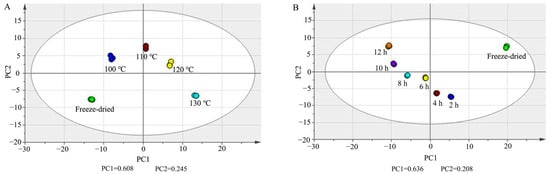
Figure 5.
PCA score plots of ginsenosides in PQ samples with different steaming temperatures (A) and times (B).
A non-parametric test was then performed, and the ratio was calculated between each of the two groups. p < 0.05 and ratio > 5 were set as the criteria to screen the differential ginsenosides (Tables S1 and S2). There were 51, 55, 58, 63, 67, and 75 differential ginsenosides found between the steamed samples of 2 h, 4 h, 6 h, 8 h, 10 h, and 12 h and freeze-dried samples, respectively. There were 48, 60, 74, and 89 differential ginsenosides found between steamed samples of 100 °C, 110 °C, 120 °C, and 130 °C and freeze-dried samples, respectively. This indicated that the steam temperature and time had a significant influence on the steamed PQ samples.
2.4. Influence of Steam Temperature and Time on the Ginsenosides Composition
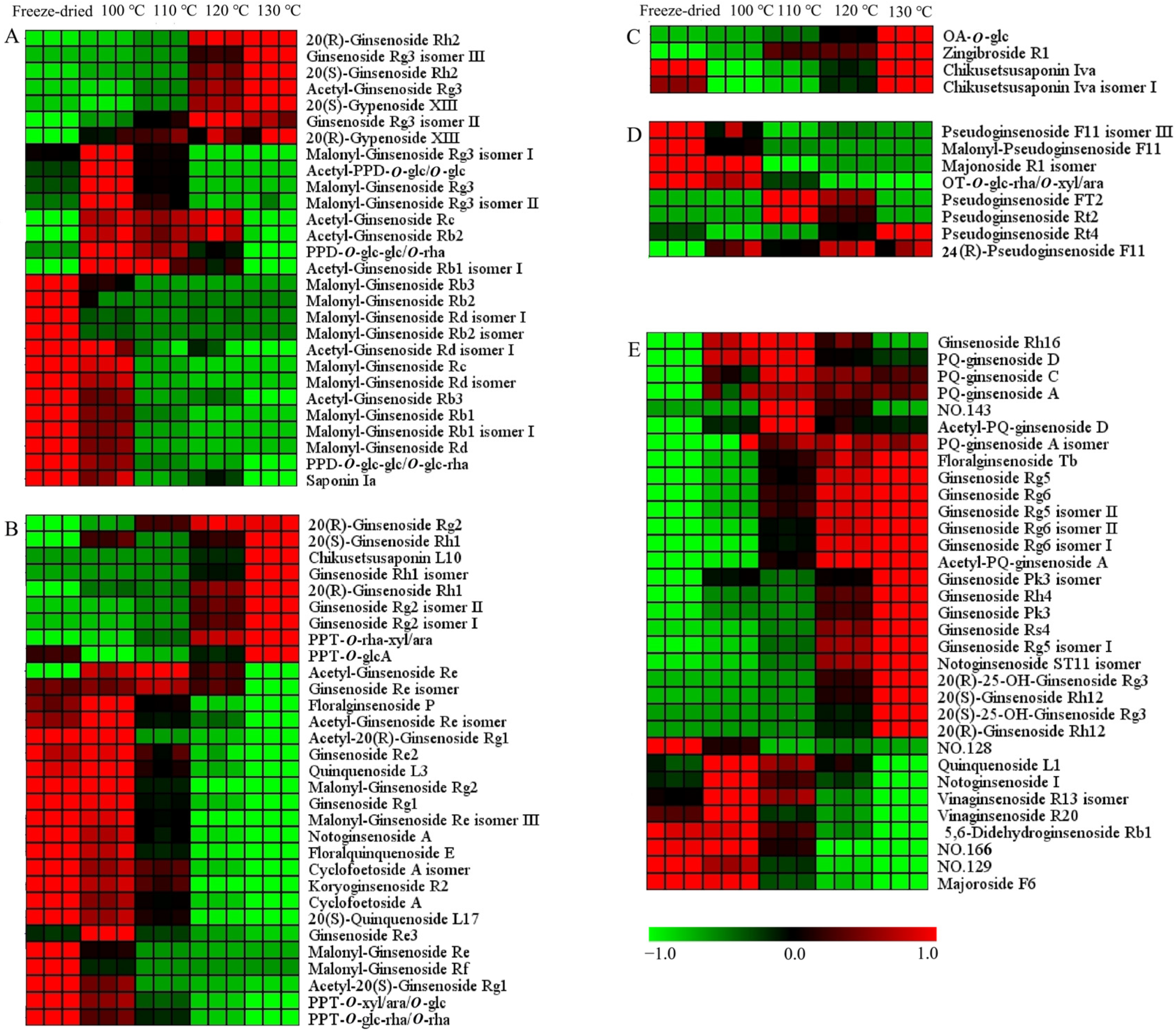
The hierarchical cluster analysis was carried out on the differential ginsenosides. The data were normalized, and the heat map is shown in Figure 6 and Figure S2. There were 104 and 86 differential ginsenosides screened for PQ samples with different steaming temperatures and times, respectively. With the increase in the steaming temperature, the content of 37 ginsenosides increased gradually, while the content of 36 ginsenosides decreased gradually. With the increase in the steaming time, the content of 42 ginsenosides increased gradually, while the content of 15 ginsenosides decreased gradually. In addition, there were some ginsenosides that changed with the steaming time and temperature, for example, climbing up and then declining. The steaming process not only changed the ginsenosides content, but also influenced their structure.
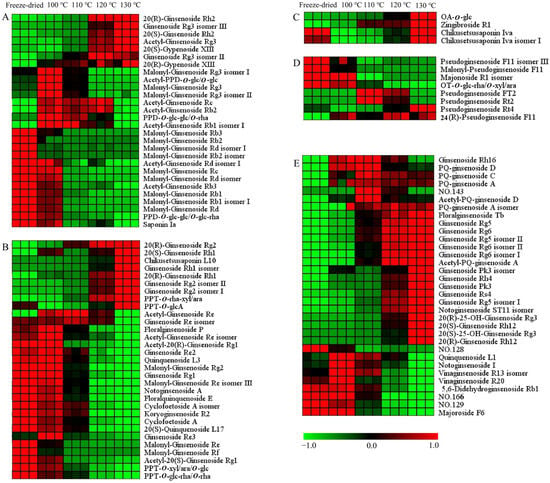
Figure 6.
Heat map of differential ginsenosides in PQ samples with different steaming temperatures. (A) PPD-type. (B) PPT-type. (C) OA-type. (D) OT-type. (E) Other type. No. 128 and 129 represent Dammarane-3,6,12,24,25-pentol, 20-(β-d-glucopyranosyloxy)-, (3β,6β,12β)-(ACI), and isomer. No. 143 represents β-d-Glucopyranoside, (3β,12β)-3,12,24,25-tetrahydroxy-20-(d-xylopyranosyloxy)dammaran-6-yl (ACI). No. 166 represents (3β,12β)-20-(β-d-Glucopyranosyloxy)-3,12,24,25-tetrahydroxydammaran-6-yl 2-O-(6-deoxy-α-l-β-d-mannopyranosyl)-β-d-glucopyranoside.
For the PPD-ginsenosides, the difference was shown in Figure 6A and Figure S2A. Firstly, the malonyl-ginsenosides Rb1, Rb2, Rb3, Rd, and Re decreased significantly with the steaming time and temperature. The rate among different samples was calculated. The content of malonyl-ginsenoside Rb3 in the freeze-dried sample was 2-, 323-, and 574-fold more than in the samples (100 °C, 2 h), (100 °C, 12 h), and (130 °C, 2 h), respectively. Malonyl-ginsenoside Rg3 was accumulated after the steaming process, and then decreased with the increase in the steaming time and temperature. Malonyl-ginsenoside Rg3 was increased by 5-fold in the sample (100 °C, 2 h), and then decreased by 26- and 47-fold in the samples (100 °C, 12 h) and (130 °C, 2 h), respectively. Secondly, the acetyl-ginsenosides showed a significant increase after the steaming process. Then, the acetyl-ginsenosides Rc, Rb3, Rb1, and Rd changed slightly with the steaming time, but showed an obviously decline with the increase in the steaming temperature. Acetyl-ginsenoside Rg3 was enhanced from steaming for 8 h at 120 °C. Thirdly, the levels of ginsenosides Rh2, Rg3, and Rb3 and gypenoside XIII were enhanced with the steaming process, while PPD-O-glc-glc/O-glc-rha and saponin Ia were decreased. And, the steaming time was not significant with the ratios. The content of 20(R)-ginsenoside Rh2 was extremely low in the freeze-dried sample, and then increased quickly after the steaming process. Although the content of 20(S)-ginsenoside Rh2 was also rising, the increase rate was low. The contents of 20(R)-ginsenoside Rh2 and 20(S)-ginsenoside Rh2 in the sample (100 °C, 2 h) were 22- and 4-fold more than in the freeze-dried sample, indicating the transformation from the S-configuration to the R-configuration. The contents of 20(R)-ginsenoside Rh2 in the samples (100 °C, 12 h) and (130 °C, 2 h) were 8 and 18 times more than in the sample (100 °C, 2 h), respectively. This shows that a high temperature enhances configuration transformation. The malonyl ginsenosides were sensitive to the heat process. The acetyl-ginsenoside could be produced by the decarboxylation of the malonyl group [34]. Therefore, with the decrease in the malonyl ginsenosides, the level of acetyl ginsenosides was enhanced. With the rise in temperature, acetyl ginsenosides were then degraded into neutral ginsenosides. The ginsenoside Rg3 could be produced from acetyl-ginsenoside Rg3, and could be further converted into ginsenoside Rh2 through the elimination of glucose at C-3 [35]. The acetyl-ginsenoside Rg3 could be produced from malonyl-ginsenoside Rg3. The possible transformation pathways are shown in Figure S3.
In the PPT-type (Figure 6B and Figure S2B), the malonyl ginsenosides Re, Rf, and Rg2 decreased with the steaming time and temperature. Acetyl-ginsenoside Re and its isomer showed a rising and then declining tendency, indicating the conversion of malonyl-ginsenosides Re. Configuration transformations were also found in the 20(S/R)-ginsenoside Rh1 and 20(S/R)-ginsenoside Rg2. The contents of 20(S)-ginsenoside Rh1 and 20(R)-ginsenoside Rh1 in the sample (130 °C, 2 h) were 15- and 750-fold more than in the freeze-dried sample, respectively. The contents of 20(S)-ginsenoside Rg2 and 20(R)-ginsenoside Rg2 in the sample (130 °C, 2 h) were 4- and 59-fold more than in the freeze-dried sample, respectively. PPT-O-rha-xyl/ara, chikusetsusaponin L10 and PPT-O-glcA were also increased, while ginsenoside Rg1, Re2, quinquenoside L3, PPT-O-xyl/ara/O-glc, PPT-O-glc-rha/O-rha, floralginsenoside P, floralquinquenoside E, Cyclofoetoside A, and 20(S)-quinquenoside L17 were decreased. The decreased ginsenosides had three or four sugars in the sugar chain, while the increased ginsenosides had one or two sugars. The sugar chains experienced hydrolysis and dehydration. The ginsenoside Re lost the C-6 sugar to generate ginsenoside Rg2 [36]. The PPT-O-glc-rha/O-rha lost the C-20 sugar to generate ginsenoside Rg2. The ginsenoside Rg2 lost the C-6 sugar to generate Rh1. The possible transformation pathways are shown in Figure S4.
For the OA-type ginsenosides, chikusetsusaponin Iva and isomer I had the same sugar chains (a glucose and a glucuronic acid attached to different positions of the aglycone). They were decreased after the steaming process (Figure 6C and Figure S2C). The content of chikusetsusaponin Iva and isomer I in the freeze-dried sample were 10 and 14 times higher than in the sample (100 °C, 2 h), respectively. Zingibroside R1 was different at the sugar chains in the structure compared with chikusetsusaponin Iva and isomer I. It had a disaccharide chain of glucose and glucuronic acid. OA-O-glc had glucose in the sugar chain. The contents of OA-O-glc and zingibroside R1 were increased after the steaming process. The contents of OA-O-glc and zingibroside R1 in the sample (100 °C, 2 h) were 12 and 6 times more than in the freeze-dried sample. The sugar chain of ginsenoside Ro has been reported to experience degradation to form the zingibroside R1 and chikusetsusaponin Iva [27]. Moreover, it has been deduced that the sugar chain of chikusetsusaponin Iva experiences degradation at C-3 to form OA-O-glc. With the increase in the steaming time and temperature, the content of them were also increased. From 2 h to 10 h, the contents of them increased continuously. At the 12 h, their content showed a little decline. The contents of chikusetsusaponin Iva, chikusetsusaponin Iva isomer I, zingibroside R1, and OA-O-glc in the sample (100 °C, 10 h) were 4, 3, 4, and 5 times more than in the sample (100 °C, 2 h), respectively. From 100 °C to 130 °C, the contents of them increased continuously. The contents of chikusetsusaponin Iva, chikusetsusaponin Iva isomer I, zingibroside R1, and OA-O-glc in the sample (130 °C, 2 h) were 9, 12, 4, and 29 times more than in the sample (100 °C, 2 h), respectively. The steaming temperature had a larger impact on the OA-type ginsenosides’ conversion than the steaming time. The possible transformation pathways are shown in Figure S5A.
OT-type ginsenosides are characteristic ingredients of PQ which different from other Panax species [27]. The differences are shown in Figure 6D and Figure S2D. In Figure 6D, Pseudoginsenoside F11 isomer III, malonyl-pseudoginsenoside F11, and OT-O-glc-rha/O-xyl/ara show a declining tendency, and pseudoginsenoside Rt4, Rt2, Ft2, and 24(R)-pseudoginsenoside F11 show a rising tendency. The contents of malonyl-pseudoginsenoside F11 and OT-O-glc-rha/O-xyl/ara in the sample (100 °C, 2 h) were 69 and 28 times more than in the sample (130 °C, 2 h). The content of malonyl-pseudoginsenoside F11 in the sample (100 °C, 2 h) was 27 times more than in the sample (100 °C, 12 h). The malonyl-pseudoginsenoside F11 and OT-O-glc-rha/O-xyl/ara underwent demalonylation and hydrolysis reactions to produce the pseudoginsenoside F11, respectively. Pseudoginsenoside Rt4 increased with the time and temperature. The contents of pseudoginsenoside Rt4 in the sample (100 °C, 12 h) and (130 °C, 2 h) was 5 and 10 times more than in the sample (100 °C, 2 h), respectively. The content of 24(R)-pseudoginsenoside F11 in the sample (130 °C, 2 h) was six times more than in the freeze-dried sample. The content of pseudoginsenoside Rt2 was the highest at 110 °C, and then declined. The sugar chain of 24(R)-pseudoginsenoside F11 (the sugar chain of glucose–rhamnose) and pseudoginsenoside Rt2 (the sugar chain of a glucose–xylose) underwent degradation to form the pseudoginsenoside Rt4 (the sugar chain of a glucose). These results indicate the degradation of the sugar chains with the steaming process, and the high temperature facilitated the degradation process. The possible transformation pathways are shown in Figure S5B.
In Figure 6E and Figure S2E, most other-type ginsenosides increase with the steaming process. Firstly, the steaming time and temperature have little difference in their influence on ginsenoside Rg5, floralginsenoside Tb, ginsenoside Rg6, ginsenoside Rg5 isomer II, and ginsenoside Rg6 isomer II. The contents of ginsenoside Rg5 in the samples (100 °C, 12 h) and (130 °C, 2 h) were five and four times more than in the sample (100 °C, 2 h), respectively. The contents of ginsenoside Rg6 in the samples (100 °C, 12 h) and (130 °C, 2 h) was four- and three-fold higher than in the sample (100 °C, 2 h), respectively. Secondly, the effect of the steaming temperature on the contents of ginsenoside Rs4, ginsenoside Rh4, ginsenoside Rg6 isomer I, 20(R)-ginsenoside Rh12, PQ-ginsenoside A, C, D, and acetyl-PQ-ginsenoside D was greater than that of the steaming time. The contents of ginsenoside Rs4 in the sample (100 °C, 12 h) and temperature sample (130 °C, 2 h) was 8 and 40 times that in the sample (100 °C, 2 h). Thirdly, the effect of the steaming time on the contents of ginsenoside Pk3, ginsenoside Rg5 isomer I, and 20(S)-ginsenoside Rh12 was greater than that of the steaming temperature. The increased rate of 20(S)-ginsenoside Rh12 was less than 20(R)-ginsenoside Rh12, indicating the transformation from an S-configuration to an R-configuration. With the steaming process, the protoginsenosides could be transformed into rare ginsenosides. The dehydration of the side chain of C17 resulted in the degradation of ginsenoside Rg3 to ginsenoside Rg5, respectively. Acetyl-ginsenoside Rg3 could be transformed into ginsenoside Rs4 by dehydration in the side chain of C17. Ginsenoside Rg2 could be transformed to ginsenoside Rg6 by dehydration in the side chain of C17. Then, the hydrolysis of the the rhamnosyl residue at C-6 of ginsenoside Rg6 can lead to its degradation into ginsenoside Rh4. The other types ginsenosides were mostly changed in the side chain at C17 of the aglycone (Figures S3 and S4). The ginsenosides underwent a series of chemical reactions, including dehydration, hydrolysis, isomerization, demalonylation, and deacetylation during the steaming process.
3. Materials and Methods
3.1. Chemical and Reagents
Acetonitrile and methanol were purchased from Merck (HPLC grade, Darmstadt, Germany). Formic acid was purchased from Honeywell (for mass spectrometry, Seelze, Germany). Ultrapure water was produced by a Direct-Q 8 UV-R water purification system (Millipore, Billerica, MA, USA). Ginsenosides Rb1, Ro, Rb2, Rd, Rg1, Rg5, Re, 20(S)-ginsenoside Rh1, 20(S)-ginsenoside Rg3, 20(R)-ginsenoside Rg3, 20(S)-ginsenoside Rg2, 20(R)-ginsenoside Rg2, 20(R)-ginsenoside Rh2, 20(S)-ginsenoside Rh2, and 24(S)-pseudoginsenoside F11 were purchased from Chengdu Desite Co., Ltd. (Chengdu, China).
3.2. Sample Information and Preparation
Four-year-old PQ roots were collected in the plantation farm of Weihai city, Shandong province, China. Fresh PQ roots were steamed at 100 °C for 2 h, 4 h, 6 h, 8 h, 10 h, and 12 h, respectively. And fresh PQ roots were steamed at 100 °C, 110 °C, 120 °C, and 130 °C for 2 h, respectively. Then the steamed PQ roots were cut into 2–3 mm slices and dried at 60 °C. The dried slices were then ground into powder. A batch of fresh roots were cut into 2–3 mm slices and freeze-dried as raw samples. Quality control (QC) samples were prepared by mixing equal weight of all samples.
PQ powder of 100 mg was accurately weighed into the Eppendorf tubes. Next, 1.5 mL extraction solution of methanol/water (1:1, v/v) was added. The mixture was vortexed for 5 min by the vortex oscillator (Digital Vortex-Genie 2, Scientific Industries, Bohemia, NY, USA). Then, the mixture was centrifuged for 10 min and the supernatant was taken out for LC-MS analysis. Three parallel samples were prepared.
3.3. LC-MS Analysis
The extract was analyzed by the UPLC system (Water, H-Class, Miford, MA, USA) coupled to a Q-TOF mass spectrometer equipped with an electrospray ionization interface (Bruker Impact II, Bremen, Germany). The LC separation was performed on an Agilent SB-Aq column (2.1 × 100 mm, 1.8 μm) with mobile phase A (0.1% formic acid in water) and mobile phase B (100% acetonitrile). The elution gradient was as follows: 0 min, 5% B; to 2 min, 7% B; to 3 min, 20% B; to 9 min, 24% B, and kept for 4 min; to 16 min, 26% B; to 18 min, 28% B; to 22 min, 34% B, and kept for 8 min; to 34 min, 36% B; to 35 min, 40% B; to 40 min, 50% B; to 50 min, 100% B, and kept for 5 min; to 55.1 min, 5%, and maintained for 5 min. The total run time was 60 min. The flow rate was 0.3 mL/min and column temperature was controlled at 35 °C. The mass data were acquired in the negative mode. The scan was set at a range of 50 to 1500 m/z. The capillary voltage was set at 3000 v for negative ion mode. The dry flow was set to 8 L/min, the nebulizer pressure was 2.0 bar, and the drying gas temperature was 200 °C. The prepulse storage was 8 μs, the collision RF was 750 Vpp, and the transfer time was 80 μs. The collision energies were set at 40–70 eV.
3.4. Data Analysis
MS-DIAL software (version 4.9.0)was used for date deconvolution and peak alignment. A dataset containing m/z, retention time and peak area was obtained. Principal component analysis (PCA) was performed using Simca 14.0 (Umetrics, Umeå, Sweden). The hierarchical cluster analysis and non-parametric test were performed using MultiExperiment Viewer software (version 4.9, Dana-Farber Cancer Institute, Boston, MA, USA). The select mode for non-parametric test was Wilcoxon and Mann–Whitney test (one factor, two experimental groups).
4. Conclusions
A comprehensive investigation was carried out on the ginsenoside identification and transformation of PQ samples with different steaming conditions. In total, 175 ginsenosides were identified, and the sugar chains were annotated based on UHPLC-QTOF-MS. New ginsenosides and isomers were discovered. The steaming process was an effective method to increase the chemical diversity of the ginsenosides. The types and contents of the ginsenosides were found to vary greatly. The content of acylated ginsenosides and protoginsenosides decreased, while the content of the rarest ginsenosides significantly increased after the steaming process. This study can deepen the understanding of the ginsenosides’ conversion in PQ during the steaming process. Since the definite functions of the individual ginsenoside have not been revealed clearly, further research is needed.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/molecules29030623/s1, Figure S1: The peak number and accumulated peak area with different RSD ranges for the repeatability (A), intra-day (B), and inter-day (C) precision. Figure S2: Heat map of differential ginsenosides in PQ samples with different steaming time. A, PPD-type. B, PPT-type. C, OA-type. D, OT-type. E, Other type. Figure S3: The deduced transformation pathways of PPD-type ginsenosides. Figure S4: The deduced transformation pathways of PPT-type ginsenosides. Figure S5: The deduced transformation pathways of OA-type ginsenosides (A) and OT-type ginsenosides (B). Table S1: The p and ratio of differential ginsenosides in the samples with different steaming times. Table S2: The p and ratio of differential ginsenosides in the samples with different steaming temperatures.
Author Contributions
Conceptualization, L.L. and X.W.; Methodology, J.F.; Validation, W.J. and F.L.; Formal Analysis, L.L.; Investigation, J.F.; Resources, X.W.; Data Curation, F.L.; Writing—Original Draft, J.F. and L.L.; Writing—Review and Editing, L.L. and X.W.; Visualization, W.J.; Supervision, F.L.; Project Administration, L.L.; Funding Acquisition, L.L. and X.W. All authors have read and agreed to the published version of the manuscript.
Funding
The work was funded by the National Natural Science Foundation of China (21904080), the New Innovation Team of Jinan (202228020), the Shandong Province Taishan Scholar Program (tstp20221138), the Key Research and Development Program of the Shandong Province (2022TZXD0034), and the Science, Education, and Industry Integration Innovation Pilot Project from Qilu University of Technology (Shandong Academy of Sciences) (2023PY048).
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Conflicts of Interest
The authors have declared no conflicts of interest.
References
- Xiong, H.; Zhang, A.; Zhao, Q.; Yan, G.; Sun, H.; Wang, X. Discovery of quality-marker ingredients of Panax quinquefolius driven by high-throughput chinmedomics approach. Phytomedicine 2020, 74, 152928. [Google Scholar] [CrossRef]
- Yang, Z.; Deng, J.; Liu, M.; He, C.; Feng, X.; Liu, S.; Wei, S. A review for discovering bioactive minor saponins and biotransformative metabolites in Panax quinquefolius L. Front. Pharmacol. 2022, 13, 972813. [Google Scholar] [CrossRef]
- Liu, L.; Xu, F.; Wang, Y. Traditional uses, chemical diversity and biological activities of Panax L. (Araliaceae): A review. J. Ethnopharmacol. 2020, 263, 112792. [Google Scholar] [CrossRef]
- Hong, H.; Kim, J.; Lim, T.; Song, Y.; Cho, C.; Jang, M. Mixing ratio optimization for functional complex extracts of rhodiolacrenulata, Panax quinquefolius, and astragalus membranaceus using mixturedesign and verification of immune functional efficacy in animal models. J. Funct. Foods. 2018, 40, 447–454. [Google Scholar] [CrossRef]
- Wei, G.; Yang, F.; Wei, F.; Zhang, L.; Gao, Y.; Qian, J.; Chen, Z.; Jia, Z.; Wang, Y.; Su, H.; et al. Metabolomes and transcriptomes revealed the saponin distribution in root tissues of Panax quinquefolius and Panax notoginseng. J. Ginseng Res. 2020, 44, 757–769. [Google Scholar] [CrossRef]
- Huang, L.; Ren, C.; Li, H.; Wu, Y. Recent progress on processing technologies, chemical components, and bioactivities of Chinese red ginseng, American red ginseng, and Korean red ginseng. Food Bioprocess Technol. 2022, 15, 47–71. [Google Scholar] [CrossRef]
- Yoo, K.; Lee, C.; Lo, M.; Moon, B. The hypoglycemic effects of American red ginseng (Panax quinquefolius L.) on a diabetic mouse model. J. Food Sci. 2012, 77, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Guo, N.; Bai, Y.; Huang, X.; Liu, X.; Cai, G.; Liu, S.; Guo, Y.; Gong, J. Comparison of the saponins in three processed American ginseng products by ultra-high performance liquid chromatography-quadrupole orbitrap tandem mass spectrometry and multivariate statistical analysis. Int. J. Anal. Chem. 2022, 2022, 6721937. [Google Scholar] [CrossRef]
- Wang, Y.; Li, H.; Li, Y.; Zhu, H.; Jin, Y. Identification of natural compounds targeting Annexin A2 with an anti-cancer effect. Protein Cell. 2018, 9, 568–579. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Xu, X.; Wang, X.; Wang, H.; Mi, Y.; Gao, X.; Guo, D.; Yang, W. Enhanced identification of ginsenosides simultaneously from seven Panax herbal extracts by data-dependent acquisition including a preferred precursor ions list derived from an in-house programmed virtual library. J. Agric. Food Chem. 2022, 70, 13796–13807. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, J.; Zuo, T.; Hu, Y.; Li, Z.; Wang, H.; Xu, X.; Yang, W.; Guo, D. Advances and challenges in ginseng research from 2011 to 2020: The phytochemistry, quality control, metabolism, and biosynthesis. Nat. Prod. Rep. 2022, 39, 875–909. [Google Scholar] [CrossRef]
- Chen, W.; Balan, P.; Popovich, D. Comparison of ginsenoside components of various tissues of new zealand forest-grown Asian ginseng (Panax Ginseng) and American ginseng (Panax Quinquefolium L.). Biomolecules 2020, 10, 372. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Fang, X.; Chen, D. Pharmacokinetics and bioavailability of ginsenoside Rb1 and Rg1 from Panax notoginseng in rats. J. Ethnopharmacol. 2003, 84, 187–192. [Google Scholar] [CrossRef]
- Fu, L.; Zhang, W.; Zhou, X.; Fu, J.; He, C. Tumor cell membrane-camouflaged responsive nanoparticles enable MRI-guided immuno-chemodynamic therapy of orthotopic osteosarcoma. Bioact. Mater. 2022, 17, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chiou, W.; Zhang, J. Comparison of the pharmacological effects of Panax ginseng and Panax quinquefolium. Acta Pharmacol. Sin. 2008, 29, 1103–1108. [Google Scholar] [CrossRef]
- Kim, S.; Kim, A. Anti-breast cancer activity of fine black ginseng (Panax ginseng Meyer) and ginsenoside Rg. J. Ginseng Res. 2015, 39, 125–134. [Google Scholar] [CrossRef]
- Liu, Y.; Fan, D. The preparation of ginsenoside Rg5, its antitumor activity against breast cancer cells and its targeting of PI3K. Nutrients 2020, 12, 246. [Google Scholar] [CrossRef] [PubMed]
- Yue, G.; Lee, J.; Li, L.; Chan, K.; Wong, E.; Chan, J.; Fung, K.; Lui, V.; Chiu, P.; Lau, C. Andrographis paniculata elicits anti-invasion activities by suppressing TM4SF3 gene expression and by anoikis-sensitization in esophageal cancer cells. Am. J. Cancer Res. 2015, 5, 3570–3587. [Google Scholar] [PubMed]
- Deng, X.; Zhao, J.; Qu, L.; Duan, Z.; Fu, R.; Zhu, C.; Fan, D. Ginsenoside Rh4 suppresses aerobic glycolysis and the expression of PD-L1 via targeting AKT in esophageal cancer. Biochem. Pharmacol. 2020, 178, 114038. [Google Scholar] [CrossRef]
- Huang, L.; Li, H.; Wu, Y. Processing technologies, phytochemistry, bioactivities and applications of black ginseng-a novel manufactured ginseng product: A comprehensive review. Food Chem. 2023, 407, 134714. [Google Scholar] [CrossRef]
- Zheng, M.; Xu, F.; Li, Y.; Xi, X.; Cui, X.; Han, C.; Zhang, X. Study on transformation of ginsenosides in different methods. BioMed. Res. Int. 2017, 2017, 8601027. [Google Scholar] [CrossRef]
- Liu, Z.; Wen, X.; Wang, C.; Li, W.; Huang, W.; Xiao, J.; Ruan, C.; Yuan, C. Remarkable impact of amino acids on ginsenoside transformationfrom fresh ginseng to red ginseng. J. Ginseng Res. 2020, 44, 424–434. [Google Scholar] [CrossRef]
- Hu, J.; Xu, X.; Li, W.; Wang, Y.; Liu, Y.; Wang, Z.; Wang, Y. Ginsenoside Rk1 ameliorates paracetamol-induced hepatotoxicity inmice through inhibition of inflammation, oxidative stress, nitrativestress and apoptosis. J. Ginseng Res. 2019, 43, 10–19. [Google Scholar] [CrossRef]
- Wang, C.; Aung, H.; Ni, M.; Wu, J.; Tong, R.; Wicks, S.; He, T.; Yuan, C. Red American ginseng: Ginsenoside constituents and antiproliferative activities of heat-processed Panax quinquefolius roots. Planta Med. 2007, 73, 669–674. [Google Scholar] [CrossRef]
- Sun, B.; Xu, M.; Li, Z.; Wang, Y.; Sung, C. UPLC-Q-TOF-MS/MS analysis for steaming times-dependent profiling of steamed Panax quinquefolius and its ginsenosides transformations induced by repetitious steaming. J. Ginseng Res. 2012, 36, 277–290. [Google Scholar] [CrossRef]
- Huang, X.; Liu, Y.; Zhang, Y.; Li, S.; Yue, H.; Chen, C.; Liu, S. Multicomponent assessment and ginsenoside conversions of Panax quinquefolium L. roots before and after steaming by HPLC-MSn. J. Ginseng Res. 2019, 43, 27–37. [Google Scholar] [CrossRef]
- Fan, W.; Yang, Y.; Li, L.; Fan, L.; Wang, Z.; Yang, L. Mass spectrometry-based profiling and imaging strategy, a fit-for-purpose tool for unveiling the transformations of ginsenosides in Panax notoginseng during processing. Phytomedicine 2022, 103, 154223. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Sun, L.; Zhang, Z.; Guo, Y.; Liu, S. Profiling and multivariate statistical analysis of Panax ginseng based on ultra-high-performance liquid chromatography coupled with quadrupole-time-of-flight mass spectrometry. J. Pharm. Biomed. Anal. 2015, 107, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Guo, Y.; Wang, Y.; Li, M.; Li, K.; Liu, X.; Fang, C.; Luo, J. Metabolomic Analysis Reveals Nutritional Diversity among Three Staple Crops and Three Fruits. Foods 2022, 11, 550. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, D.; Sun, C.; Li, Y.; Lu, H.; Wang, X. Comprehensive lipidome and metabolome profiling investigations of Panax quinquefolius and application in different growing regions using liquid chromatography coupled with mass spectrometry. J. Agric. Food Chem. 2021, 69, 6710–6719. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Hu, Y.; Wu, W.; Ye, M.; Guo, D. Saponins in the genus Panax L. (Araliaceae): A systematic review of their chemical diversity. Phytochemistry 2014, 106, 7–24. [Google Scholar] [CrossRef] [PubMed]
- Tung, N.; Song, G.; Nhiem, N.; Ding, Y.; Tai, B.; Jin, L.; Lim, C.; Hyun, J.; Park, C.; Kang, H.; et al. Dammarane-type saponins from the flower buds of Panax ginseng and their intracellular radical scavenging capacity. J. Agric. Food Chem. 2010, 58, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Park, H.; Lee, D.; Jayakodi, M.; Kim, N.; Koo, H.; Lee, S.; Kim, Y.; Kwon, S.; Yang, T. Integrated transcriptomic and metabolomic analysis of five Panax ginseng cultivars reveals the dynamics of ginsenoside biosynthesis. Front. Plant Sci. 2017, 8, 1048. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.; Bai, M.; Xu, J.; Kong, M.; Zhu, L.; Zhu, H.; Wang, Q.; Li, S. Discrimination of leaves of Panax ginseng and P. quinquefolius by ultrahigh performance liquid chromatography quadrupole/time-of-flightmass spectrometry based metabolomics approach. J. Pharm. Biomed. Anal. 2014, 97, 129–140. [Google Scholar] [CrossRef]
- Xu, X.; Gao, Y.; Xu, S.; Liu, H.; Xue, X.; Zhang, Y.; Zhang, H.; Liu, M.; Xiong, H.; Lin, R.; et al. Remarkable impact of steam temperature on ginsenosides transformation from fresh ginseng to red ginseng. J. Ginseng Res. 2018, 42, 277–287. [Google Scholar] [CrossRef]
- Wu, W.; Qin, Q.; Guo, Y.; Sun, J.; Liu, S. Studies on the chemical transformation of 20(S)-protopanaxatriol (PPT)-type ginsenosides Re, Rg2, and Rf using rapid resolution liquid chromatography coupled with quadruple-time-of-flight mass spectrometry (RRLC-Q-TOF-MS). J. Agric. Food Chem. 2012, 60, 10007–10014. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).