
Spinetoram-Induced Potential Neurotoxicity through Autophagy Mediated by Mitochondrial Damage



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
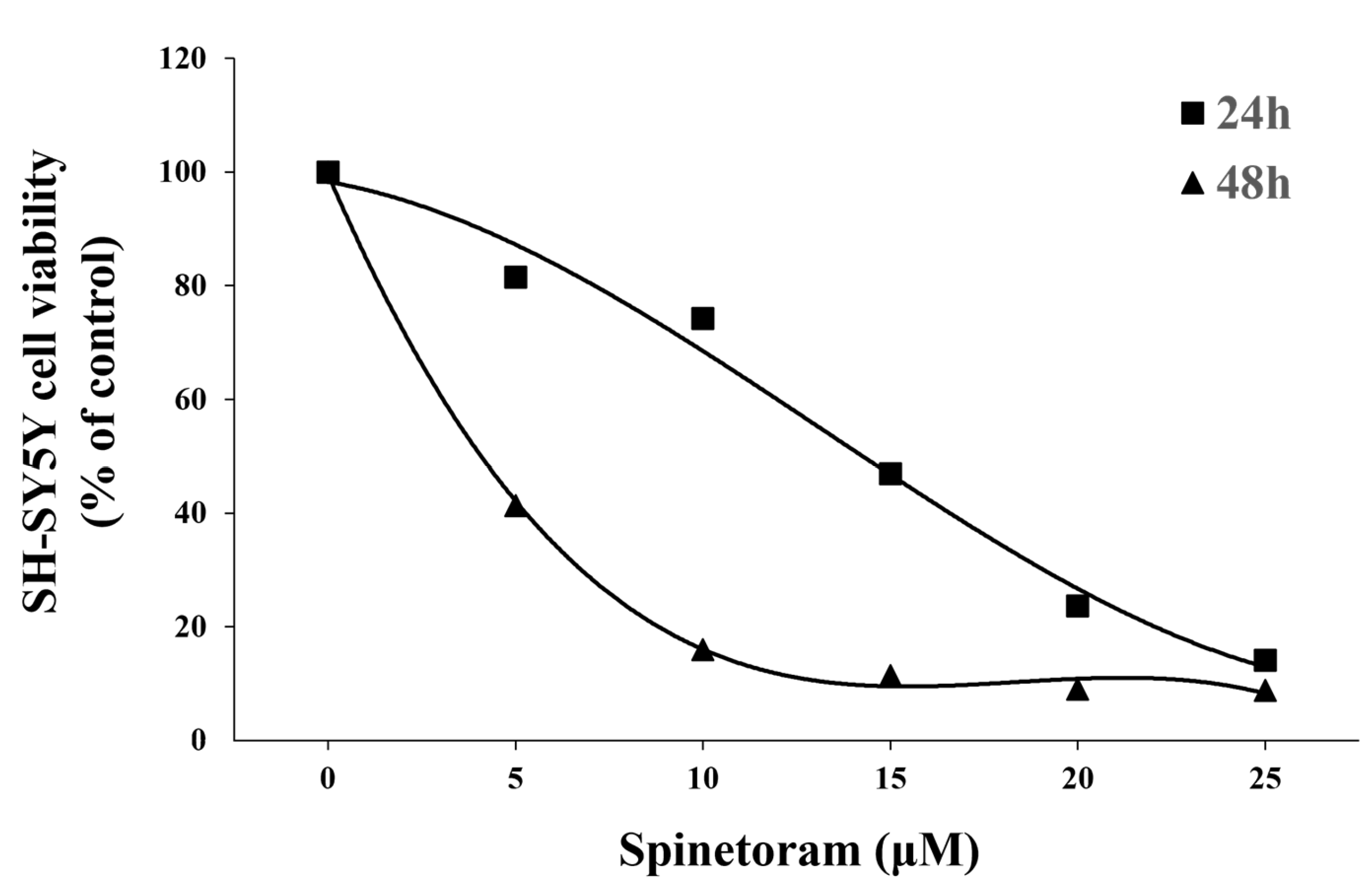
2.1. Spinetoram Exposure Inhibited SH-SY5Y Cell Viability
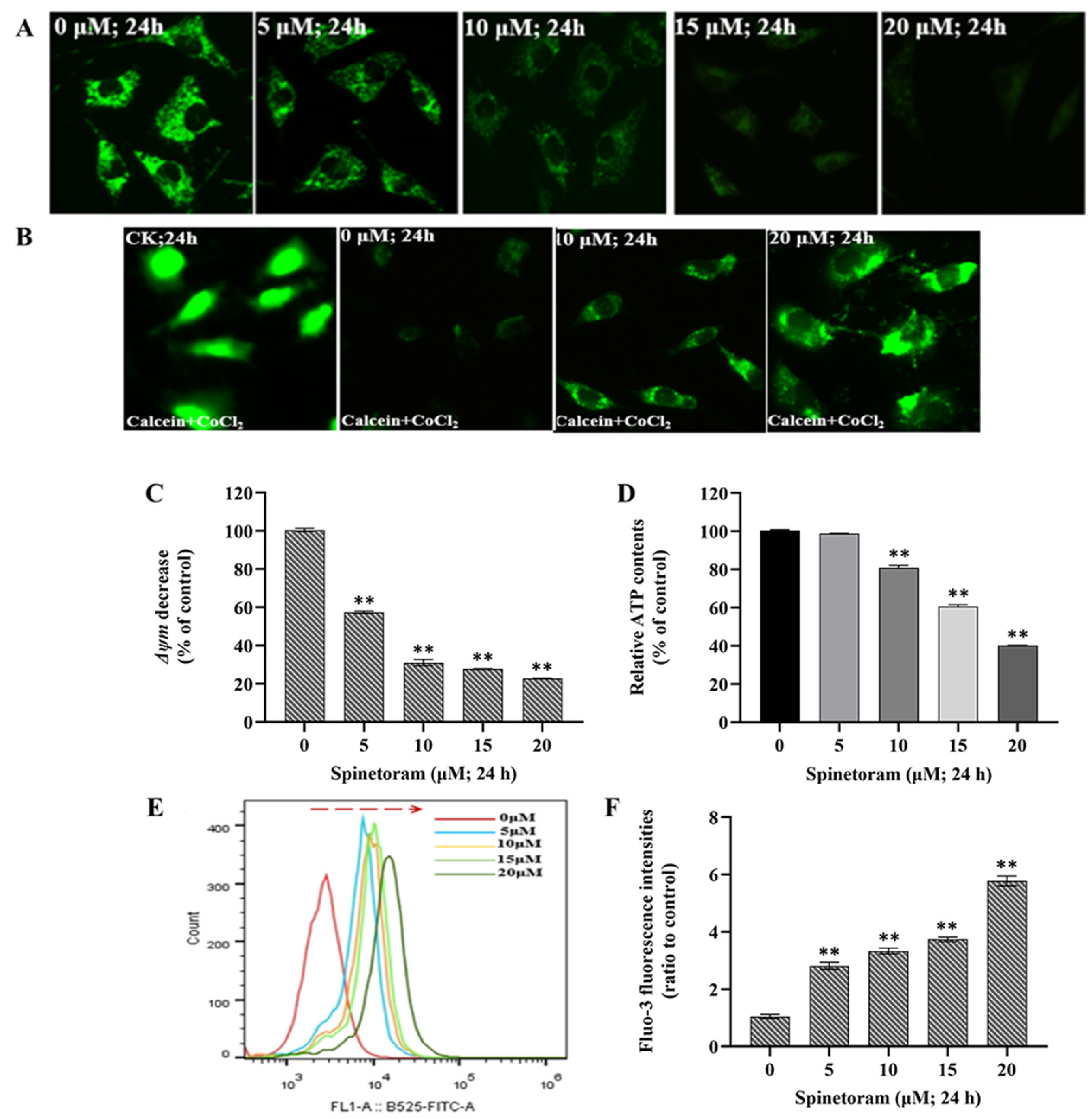
2.2. Spinetoram Induced Mitochondrial Dysfunction in SH-SY5Y Cells
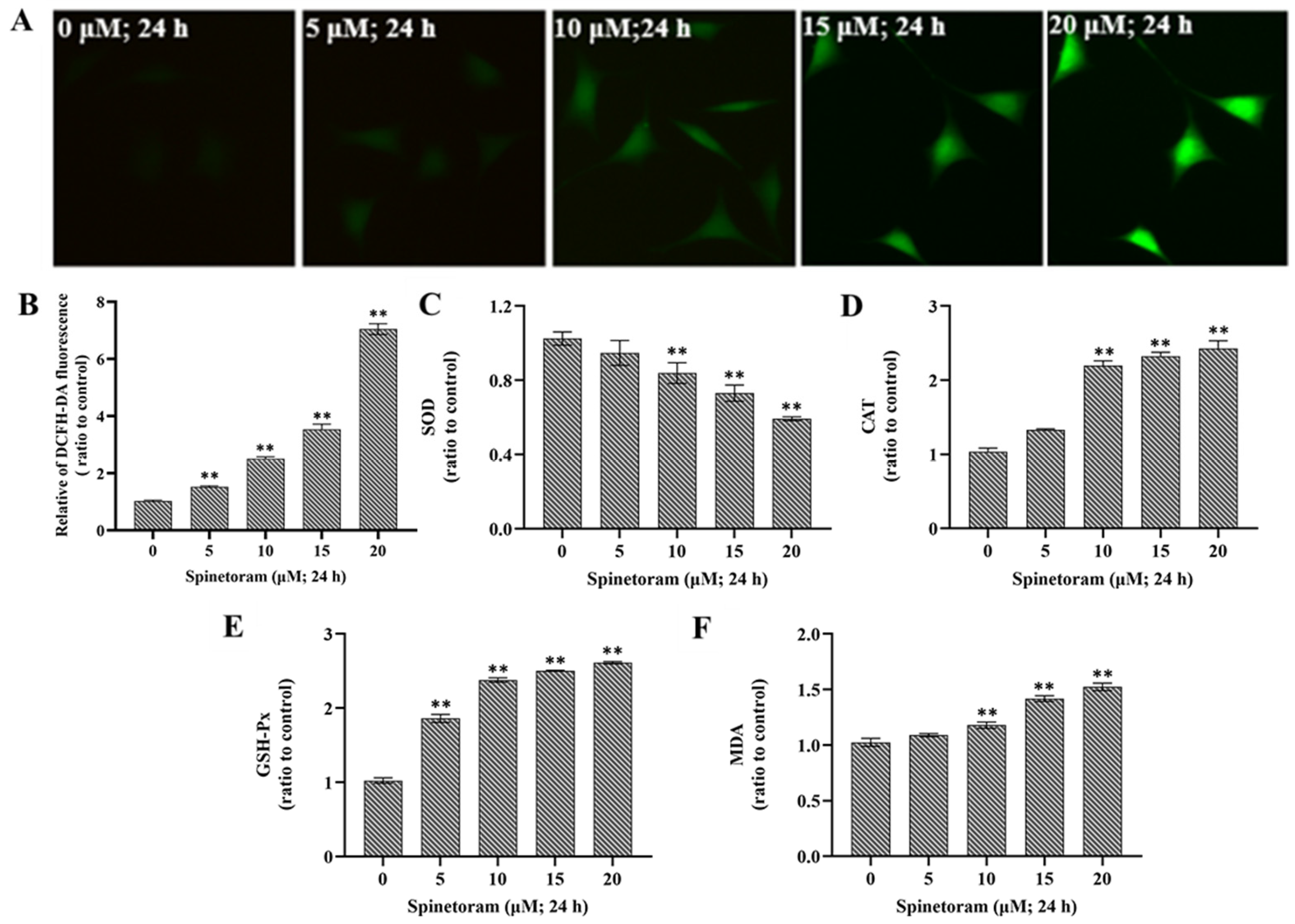
2.3. Spinetoram Induced Oxidative Damage in SH-SY5Y Cells
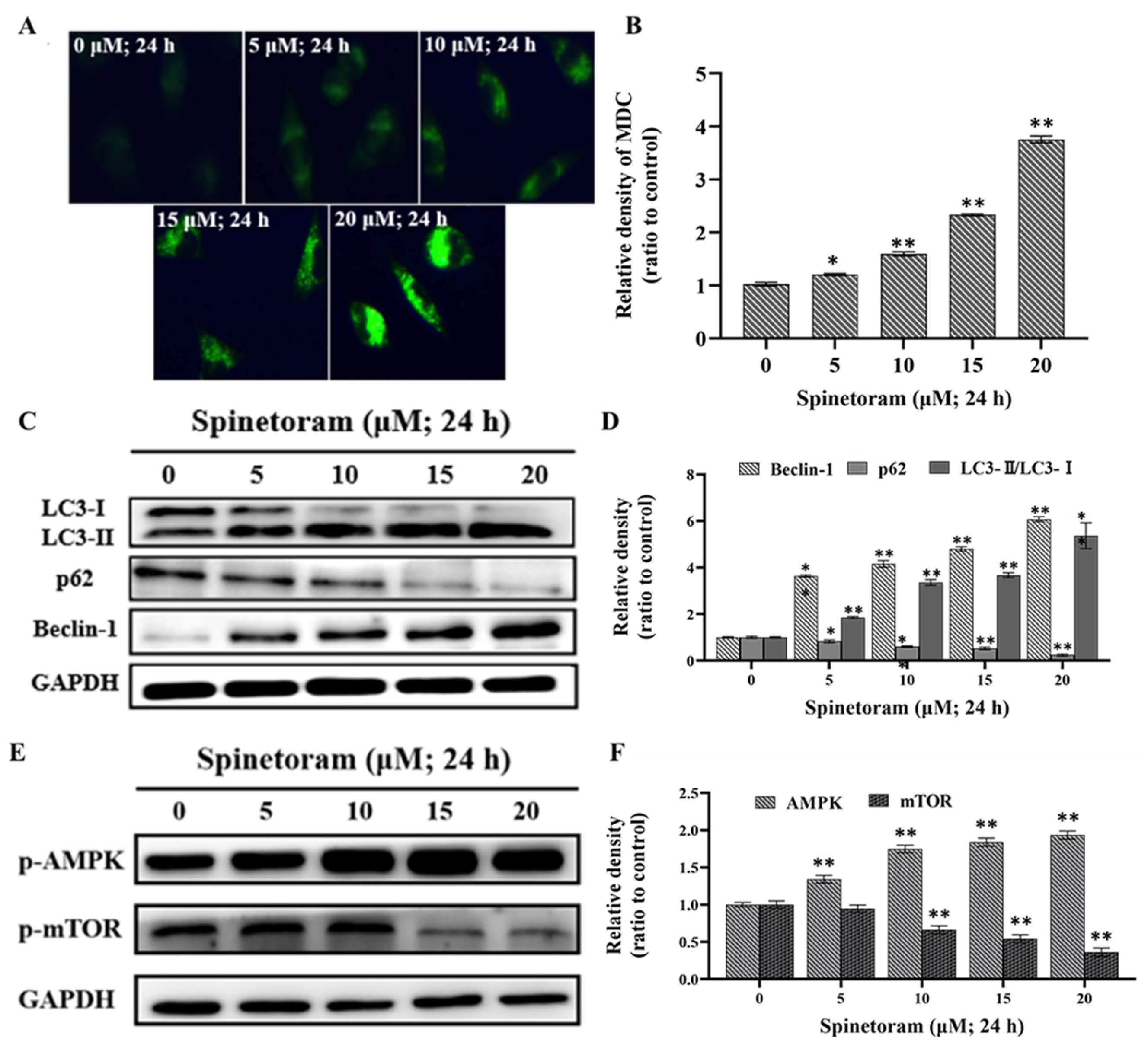
2.4. Spinetoram Induced Autophagy through AMPK/mTOR Signaling Pathways in SH-SY5Y Cells
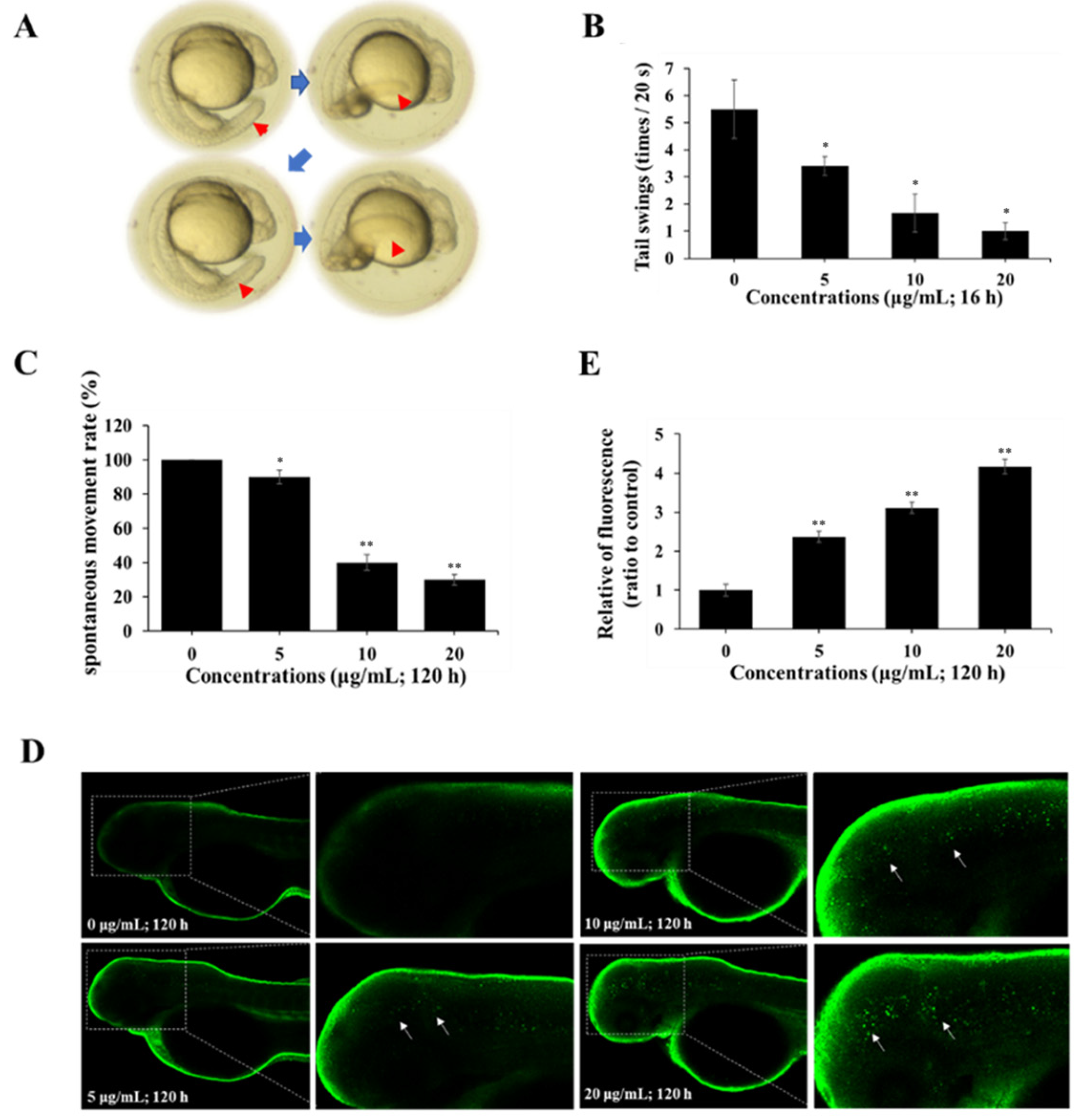
2.5. Spinetoram Altered Neuromotor Behavior in Zebrafish
2.6. Spinetoram Induced Autophagy in Zebrafish
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Cell Culture and Treatment
4.3. Cell Viability Assay
4.4. Mitochondrial Membrane Potential (ΔΨm) Analysis
4.5. Measurement of Mitochondrial Permeability Transition Pore (mPTP), Intracellular ATP and Ca2+ Level
4.6. Monodansylcadaverine (MDC) Staining Assay
4.7. Detection of ROS Content and Enzyme Activity
4.8. Western Blot Analysis
4.9. Zebrafish Husbandry
4.10. Neuromotor Behavior of Zebrafish
4.11. Immunofluorescence Assay
4.12. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saravi, S.S.S.; Shokrzadeh, M. Role of pesticides in human life in the modern age: A review. In Pesticides in the Modern World—Risks and Benefits; InTech: Rijeka, Croatia, 2011. [Google Scholar]
- Galm, U.; Sparks, T.C. Natural product derived insecticides: Discovery and development of spinetoram. J. Ind. Microbiol. Biotechnol. 2016, 43, 185–193. [Google Scholar] [CrossRef] [PubMed]
- DeAmicis, C.; Edwards, N.A.; Giles, M.B.; Harris, G.H.; Hewitson, P.; Janaway, L.; Ignatova, S. Comparison of preparative reversed phase liquid chromatography and countercurrent chromatography for the kilogram scale purification of crude spinetoram insecticide. J. Chromatogr. A 2011, 1218, 6122–6127. [Google Scholar] [CrossRef] [PubMed]
- Burgess, E.R.; King, B.H.; Geden, C.J. Oral and Topical Insecticide Response Bioassays and Associated Statistical Analyses Used Commonly in Veterinary and Medical Entomology. J. Insect Sci. 2020, 20, 6. [Google Scholar] [CrossRef] [PubMed]
- Sheele, J.M. A Preliminary Report Showing Spinosad and Fluralaner Are Able to Incapacitate Cimex lectularius L., the Common Bed Bug. Cureus 2020, 12, e7529. [Google Scholar] [CrossRef] [PubMed]
- Franc, M.; Bouhsira, E.; Bohm, C.; Wolken, S.; Wolf, O.; Lohlein, W.; Wiseman, S.; Hayes, B.; Schnitzler, B.; Fisher, M. Evaluation of spinosad for the oral treatment and control of flea infestations on cats in Europe. Vet. Rec. Open 2014, 1, e000047. [Google Scholar] [CrossRef] [PubMed]
- Paarlberg, T.; Winkle, J.; Rumschlag, A.J.; Young, L.M.; Ryan, W.G.; Snyder, D.E. Effectiveness and residual speed of flea kill of a novel spot on formulation of spinetoram (Cheristin (R)) for cats. Parasites Vectors 2017, 10, 59. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhong, Q.; Luo, F.; Wang, X.; Zhou, L.; Chen, Z.; Zhang, X. Residue degradation and metabolism of spinetoram in tea: A growing, processing and brewing risk assessment. Food Control 2021, 125, 107955. [Google Scholar] [CrossRef]
- Sparks, T.C.; Crouse, G.D.; Dripps, J.E.; Anzeveno, P.; Martynow, J.; Deamicis, C.V.; Gifford, J. Neural network-based QSAR and insecticide discovery: Spinetoram. J. Comput. Aided Mol. Des. 2008, 22, 393–401. [Google Scholar] [CrossRef]
- Liu, S.; Li, J.L.; Chen, J.; Wei, H.; Qiao, H.L.; Xu, R.; Guo, K.; Lin, C.; Jin, H.Y.; Ma, S.C.; et al. Residue degradation dynamics of spinetoram in wolfberry. China J. Chin. Mater. Med. 2016, 41, 1815–1818. [Google Scholar]
- Cheng, B.; Zhang, H.; Jia, K.; Li, E.; Zhang, S.; Yu, H.; Cao, Z.; Xiong, G.; Hu, C.; Lu, H. Effects of spinetoram on the developmental toxicity and immunotoxicity of zebrafish. Fish Shellfish Immunol. 2020, 96, 114–121. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, W.; Chen, H.; Gao, J.; Tao, L.; Li, Z.; Xu, W. The cytotoxic effects of spinetoram on human HepG2 cells by inducing DNA damage and mitochondria-associated apoptosis. Food Agric. Immunol. 2019, 30, 1020–1032. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, W.; Chen, H.; Gao, J.; Xu, Z.; Tao, L.; Li, Z.; Xu, W. Spinetoram confers its cytotoxic effects by inducing AMPK/mTOR-mediated autophagy and oxidative DNA damage. Ecotoxicol. Environ. Saf. 2019, 183, 109480. [Google Scholar] [CrossRef]
- Pellacani, C.; Eleftheriou, G. Neurotoxicity of antineoplastic drugs: Mechanisms, susceptibility, and neuroprotective strategies. Adv. Med. Sci. 2020, 65, 265–285. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. Guidelines for Neurotoxicity Risk Assessment; Federal Register; U.S. Environmental Protection Agency: Washington, DC, USA, 1998.
- Szpara, M.L.; Mangold, C.A.; Shipley, M.M. Differentiation of the SH-SY5Y Human Neuroblastoma Cell Line. J. Vis. Exp. 2016, 108, e53193. [Google Scholar]
- Krishna, A.; Biryukov, M.; Trefois, C.; Antony, P.M.; Hussong, R. Systems genomics evaluation of the SH-SY5Y neuroblastoma cell line as a model for Parkinson’s disease. BMC Genom. 2014, 15, 1154. [Google Scholar] [CrossRef]
- Dravid, A.; Raos, B.; Svirskis, D.; O’Carroll, S.J. Optimised techniques for high-throughput screening of differentiated SH-SY5Y cells and application for neurite outgrowth assays. Sci. Rep. 2021, 11, 23935. [Google Scholar] [CrossRef]
- Truong, L.; Reif, D.M.; St Mary, L.; Geier, M.C.; Truong, H.D.; Tanguay, R.L. Multidimensional in vivo hazard assessment using zebrafish. Toxicol. Sci. 2014, 137, 212–233. [Google Scholar] [CrossRef]
- Tierney, K.B. Behavioural assessments of neurotoxic effects and neurodegeneration in zebrafish. Biochim. Biophys. Acta 2011, 1812, 381–389. [Google Scholar] [CrossRef]
- Shi, R.; Guberman, M.; Kirshenbaum, L.A. Mitochondrial quality control: The role of mitophagy in aging. Trends Cardiovasc. Med. 2018, 28, 246–260. [Google Scholar] [CrossRef]
- Zhu, C.L.; Yao, R.Q.; Li, L.X.; Li, P.; Xie, J.; Wang, J.F.; Deng, X.M. Mechanism of Mitophagy and Its Role in Sepsis Induced Organ Dysfunction: A Review. Front. Cell Dev. Biol. 2021, 9, 664896. [Google Scholar] [CrossRef]
- Moreira, P.I.; Carvalho, C.; Zhu, X.; Smith, M.A.; Perry, G. Mitochondrial dysfunction is a trigger of Alzheimer’s disease pathophysiology. Biochim. Biophys. Acta 2010, 1802, 2–10. [Google Scholar] [CrossRef]
- Jung, H.; Kim, S.Y.; Canbakis Cecen, F.S.; Cho, Y.; Kwon, S.K. Dysfunction of Mitochondrial Ca(2+) Regulatory Machineries in Brain Aging and Neurodegenerative Diseases. Front. Cell Dev. Biol. 2020, 8, 599792. [Google Scholar] [CrossRef]
- Patlolla, A.K.; Barnes, C.; Yedjou, C.; Velma, V.R.; Tchounwou, P.B. Oxidative stress, DNA damage, and antioxidant enzyme activity induced by hexavalent chromium in Sprague-Dawley rats. Environ. Toxicol. 2009, 24, 66–73. [Google Scholar] [CrossRef]
- Liu, X.; Hussain, R.; Mehmood, K.; Tang, Z.; Zhang, H.; Li, Y. Mitochondrial-Endoplasmic Reticulum Communication-Mediated Oxidative Stress and Autophagy. Biomed. Res. Int. 2022, 2022, 6459585. [Google Scholar] [CrossRef] [PubMed]
- Khoubnasabjafari, M.; Ansarin, K.; Jouyban, A. Reliability of malondialdehyde as a biomarker of oxidative stress in psychological disorders. Bioimpacts 2015, 5, 123–127. [Google Scholar]
- Wang, S.; Zhang, K.; Yao, Y.; Li, J. Autophagy and Mitochondrial Homeostasis During Infection: A Double-Edged Sword. Front. Cell Dev. Biol. 2021, 9, 738932. [Google Scholar] [CrossRef] [PubMed]
- Tam, J.M.; Mansour, M.K.; Acharya, M.; Sokolovska, A.; Timmons, A.K.; Lacy-Hulbert, A.; Vyas, J.M. The Role of Autophagy-Related Proteins in Candida albicans Infections. Pathogens 2016, 5, 34. [Google Scholar] [CrossRef] [PubMed]
- Chifenti, B.; Locci, M.T.; Lazzeri, G.; Guagnozzi, M.; Dinucci, D.; Chiellini, F.; Filice, M.E.; Salerno, M.G.; Battini, L. Autophagy-related protein LC3 and Beclin-1 in the first trimester of pregnancy. Clin. Exp. Reprod. Med. 2013, 40, 33–37. [Google Scholar] [CrossRef]
- Liu, W.J.; Ye, L.; Huang, W.F.; Guo, L.J.; Xu, Z.G.; Wu, H.L.; Yang, C.; Liu, H.F. p62 links the autophagy pathway and the ubiqutin-proteasome system upon ubiquitinated protein degradation. Cell Mol. Biol. Lett. 2016, 21, 29. [Google Scholar] [CrossRef]
- Vega-Rubin-de-Celis, S.; Kinch, L.; Pena-Llopis, S. Regulation of Beclin 1-Mediated Autophagy by Oncogenic Tyrosine Kinases. Int. J. Mol. Sci. 2020, 21, 9210. [Google Scholar] [CrossRef]
- Kim, K.H.; Lee, J.Y.; Li, W.Y.; Lee, S.; Jeong, H.S.; Choi, J.Y.; Joo, M. The ethanol extract of Garcinia subelliptica Merr. induces autophagy. BMC Complement. Med. Ther. 2021, 21, 280. [Google Scholar] [CrossRef]
- Xia, S.; Zhu, X.; Yan, Y.; Zhang, T.; Chen, G.; Lei, D.; Wang, G. Developmental neurotoxicity of antimony (Sb) in the early life stages of zebrafish. Ecotoxicol. Environ. Saf. 2021, 218, 112308. [Google Scholar] [CrossRef] [PubMed]
- Raftery, T.D.; Volz, D.C. Abamectin induces rapid and reversible hypoactivity within early zebrafish embryos. Neurotoxicol. Teratol. 2015, 49, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.Y.; Wang, W.G.; Fu, W.; Zhang, F.; Feng, H.; Xu, W.P.; Tao, L.M.; Li, Z.; Zhang, Y.; Shao, X.S. The potential immunotoxicity of Emamectin Benzoate on the human THP-1 macrophages. Environ. Toxicol. 2022, 38, 500–510. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J.; Stainier DY, R.; Yelon, D. Screening mosaic F1 females for mutations affecting zebrafish heart induction and patterning. Dev. Genet. 1998, 22, 288–299. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, F.; Lu, J.; Li, M.; Yang, J.; Xu, W.; Jiang, X.; Zhang, Y. Spinetoram-Induced Potential Neurotoxicity through Autophagy Mediated by Mitochondrial Damage. Molecules 2024, 29, 253. https://doi.org/10.3390/molecules29010253
Chen F, Lu J, Li M, Yang J, Xu W, Jiang X, Zhang Y. Spinetoram-Induced Potential Neurotoxicity through Autophagy Mediated by Mitochondrial Damage. Molecules. 2024; 29(1):253. https://doi.org/10.3390/molecules29010253
Chicago/Turabian StyleChen, Fan, Jin Lu, Meng Li, Junwu Yang, Wenping Xu, Xufeng Jiang, and Yang Zhang. 2024. "Spinetoram-Induced Potential Neurotoxicity through Autophagy Mediated by Mitochondrial Damage" Molecules 29, no. 1: 253. https://doi.org/10.3390/molecules29010253
APA StyleChen, F., Lu, J., Li, M., Yang, J., Xu, W., Jiang, X., & Zhang, Y. (2024). Spinetoram-Induced Potential Neurotoxicity through Autophagy Mediated by Mitochondrial Damage. Molecules, 29(1), 253. https://doi.org/10.3390/molecules29010253