


The Chemopreventive Effects of Chlorogenic Acids, Phenolic Compounds in Coffee, against Inflammation, Cancer, and Neurological Diseases

Abstract
1. Introduction
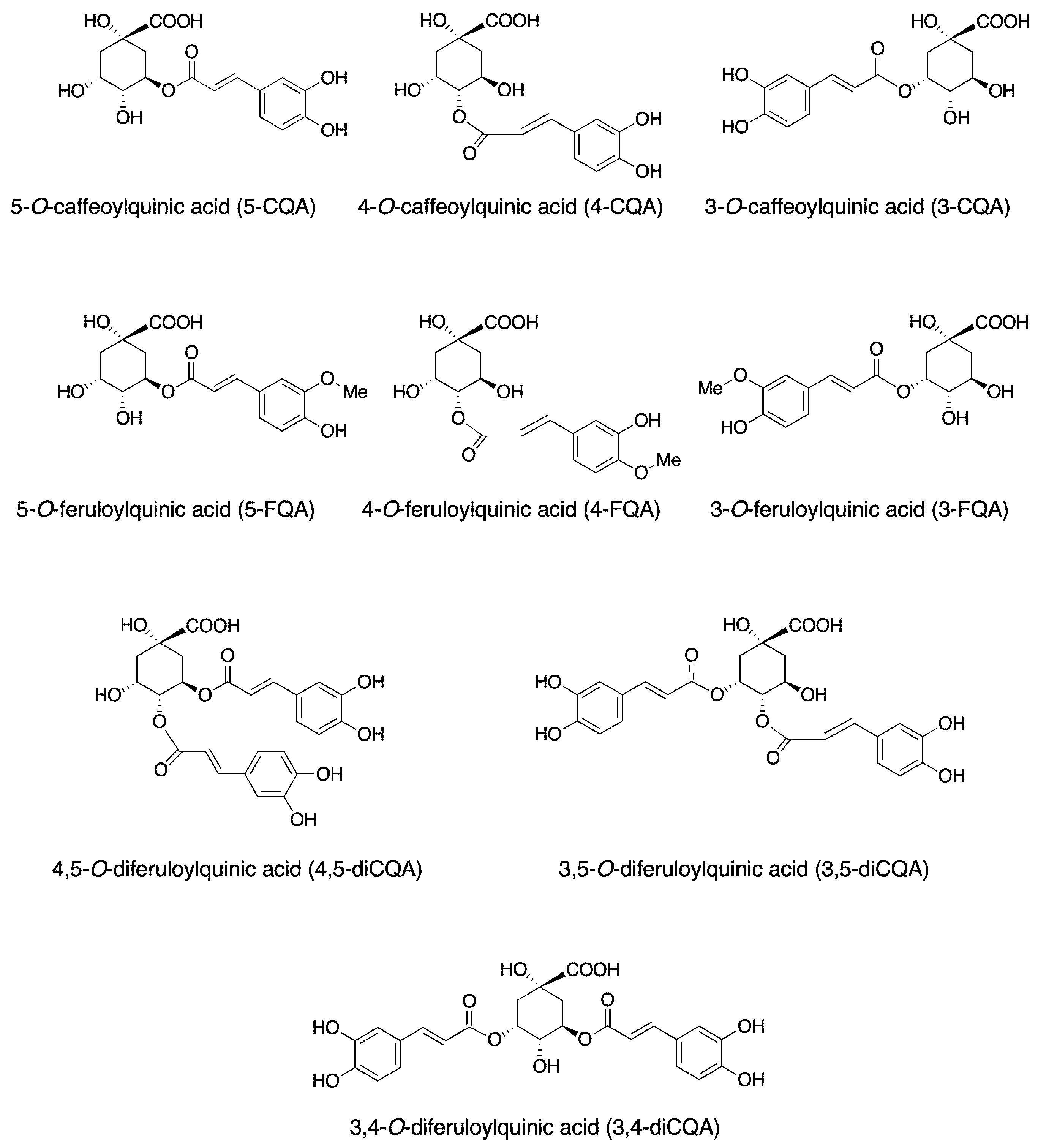
2. Chemical Ingredients of Coffee
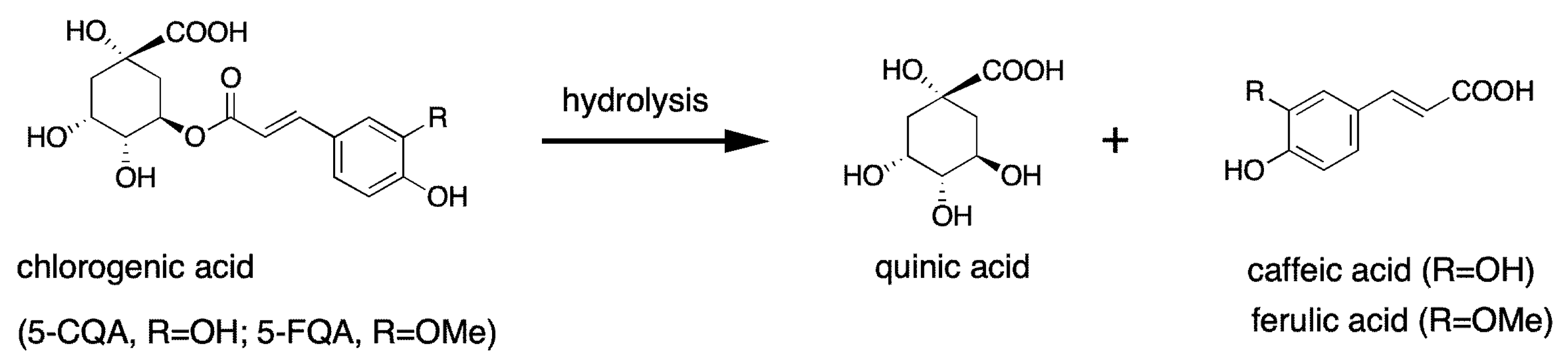
3. The Metabolism of Chlorogenic Acids
4. Anti-Inflammatory Activity of Chlorogenic Acids
5. Anti-Cancer Activity of Chlorogenic Acids
6. Chlorogenic Acid and Neurological Diseases
7. Membrane-Modulating Activity of Chlorogenic Acids
8. Conclusions and Future Perspective
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| caffeine-containing beverage (CCO) |
| Food and Agriculture Organization (FAO) |
| breast cancer susceptibility genes (BRCA) |
| 5-O-caffeoylquinic acid (5-CQA) |
| 3-O-caffeoylquinic acid (3-CQA) |
| 4-O-caffeoylquinic acid (4-CQA) |
| reactive oxygen species (ROS) |
| interleukin-8 (IL-8) |
| nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) |
| tumor necrosis factor-alpha (TNF-α) |
| intercellular adhesion molecule 1 (ICAM-1) |
| vascular cell adhesion molecule 1 (VCAM-1) |
| very late antigen-4 (VLA-4) |
| epidermal growth factor (EGF) |
| mitogen-activated protein kinase (MAP kinase) |
| extracellular-signal-regulated kinase (ERK) |
| jun amino-terminal kinases (JNK) |
| stress-activated protein kinases (SAPK) |
| matrix metalloproteinase (MMP) |
| tissue inhibitors of MMP (TIMP) |
| phosphatidylinositol 3-kinase (PI3K) |
| mammalian target of rapamycin (mTOR) |
| phosphatase and tensin homologue deleted on chromosome ten (PTEN) |
| breast cancer susceptibility gene 1 (BRCA1) |
| Bcl-2-associated X protein (Bax) |
| B-cell/CLL lymphoma-2 (Bcl-2) |
| B-cell lymphoma-extra-large (Bcl-xL) |
| T-cell factor and lymphoid enhancer factor (TCF/LEF) |
| leucine-rich repeat-containing G-protein coupled receptor 5 (LGR5) |
| nuclear factor erythroid 2-related factor 2 (Nrf2) |
| cyclic adenosine monophosphate (cAMP) |
| cyclic adenosine monophosphate responsive element binding protein (CREB) |
| glycosylphosphatidylinositol (GPI) |
| 1,2-di-istoyl-sn-glycero-3-phosphocholine (DMPC) |
| 1,2-di-O-tetradecyl-sn-glycero-3-phosphocholine (14:0 diether PC) |
| 1,2-dipalmitoyl-sn-glycero-3-phosphocholine (DPPC) |
| 1,2-di-O-hexadecyl-sn-glycero-3-phosphocholine (16:0 diether PC) |
| Fourier transform infrared spectroscopy (FT-IR) |
References
- Food and Agriculture Organization of the United Nations (FAO). Statistical Pocket Book, Coffee; FAO: Rome, Italy, 2015. [Google Scholar]
- Wolf, A.; Bray, G.A.; Popkin, B.M. A short history of beverages and how our body treats them. Obes. Rev. 2008, 9, 151–164. [Google Scholar] [CrossRef] [PubMed]
- United States Department of Agriculture Foreign Agricultural Service. Coffee: World Markets and Trade. 2022. Available online: https://www.fas.usda.gov/data/coffee-world-markets-and-trade (accessed on 31 January 2023).
- Gross, M. What coffee does to body and mind. Curr. Biol. 2021, 31, R311–R313. [Google Scholar] [CrossRef]
- Ludwig, I.A.; Mena, P.; Calani, L.; Cid, C.; Del Rio, D.; Lean, M.E.; Crozier, A. Variations in caffeine and chlorogenic acid contents of coffees: What are we drinking? Food Funct. 2014, 5, 1718–1726. [Google Scholar] [CrossRef]
- Mullee, A.; Romaguera, D.; Pearson-Stuttard, J.; Viallon, V.; Stepien, M.; Freisling, H.; Fagherazzi, G.; Mancini, F.R.; Boutron-Ruault, M.C.; Kühn, T.; et al. Association between soft drink consumption and mortality in 10 European countries. JAMA Intern. Med. 2019, 179, 1479–1490. [Google Scholar] [CrossRef]
- Zidan, N.S.; Omran, A.M.E.; Rezk, S.M.; Hebatallah, H.A.; Mohamed, I.S. Anti-Alzheimer’s disease potential of Arabian coffee versus Date palm seed extract in male rats. J. Food Biochem. 2022, 46, e14017. [Google Scholar] [CrossRef] [PubMed]
- Camandola, S.; Plick, N.; Mattson, M.P. Impact of coffee and cacao purine metabolites on neuroplasticity and neurodegenerative disease. Neurochem. Res. 2019, 44, 214–227. [Google Scholar] [CrossRef]
- Karpinska, J.; Świsłocka, R.; Lewandowski, W. A mystery of a cup of coffee; an insight look by chemist. BioFactors 2017, 43, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, M.C.; Cornelis, M.C. Caffeine in the diet: Country-level consumption guidelines. Nutrients 2018, 10, 1772. [Google Scholar] [CrossRef]
- Nkondjock, A. Coffee consumption and the risk of cancer: An overview. Cancer Lett. 2009, 277, 121–125. [Google Scholar] [CrossRef]
- Nkondjock, A.; Ghadirian, P.; Kotsopoulos, J.; Lubinski, J.; Lynch, H.; Kim-Sing, C.; Horsman, D.; Rosen, B.; Isaacs, C.; Weber, B.; et al. Coffee consumption and breast cancer risk among BRCA1 and BRCA2 mutation carriers. Int. J. Cancer 2006, 118, 103–107. [Google Scholar] [CrossRef]
- Kim, S.Y.; Yoo, D.M.; Min, C.; Choi, H.G. Association between coffee consumption/physical exercise and gastric, hepatic, colon, breast, uterine cervix, lung, thyroid, prostate, and bladder cancer. Nutrients 2021, 13, 3927. [Google Scholar] [CrossRef] [PubMed]
- Kawada, T. Coffee consumption and risk of cancers: Kidney as an example for the assessment. Clin. Nutr. 2022, 41, 3122. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wu, T.; Luo, G.; Chen, L. Exploring the casual association between coffee intake and bladder cancer risk using Mendelian Randomization. Front. Genet. 2022, 13, 992599. [Google Scholar] [CrossRef] [PubMed]
- Ferhatosmanoğlu, A.; Selcuk, L.B.; Arıca, D.A.; Ersöz, Ş.; Yaylı, S. Frequency of skin cancer and evaluation of risk factors: A hospital-based study from Turkey. J. Cosmet. Dermatol. 2022, 21, 6920–6927. [Google Scholar] [CrossRef]
- Li, B.H.; Yan, S.Y.; Li, X.H.; Huang, Q.; Luo, L.S.; Wang, Y.Y.; Huang, J.; Jin, Y.H.; Wang, Y.B. Coffee and caffeine consumption and risk of renal cell carcinoma: A Mendelian randomization study. Front. Nutr. 2022, 9, 898279. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Huang, J.; Wong, M.C.S. Associations of alcohol and coffee with colorectal cancer risk in East Asian populations: A Mendelian randomization study. Eur. J. Nutr. 2022, 62, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Carter, P.; Yuan, S.; Kar, S.; Vithayathil, M.; Mason, A.M.; Burgess, S.; Larsson, S.C. Coffee consumption and cancer risk: A Mendelian randomisation study. Clin. Nutr. 2022, 21, 2113–2123. [Google Scholar] [CrossRef]
- Soldato, D.; Havas, J.; Crane, T.E.; Presti, D.; Lapidari, P.; Rassy, N.; Pistilli, B.; Martin, E.; Del Mastro, L.; Martin, A.L.; et al. Coffee and tea consumption, patient-reported, and clinical outcomes in a longitudinal study of patients with breast cancer. Cancer 2022, 128, 3552–3563. [Google Scholar] [CrossRef]
- Crous-Bou, M.; Du, M.; Gunter, M.J.; Setiawan, V.W.; Schouten, L.J.; Shu, X.O.; Wentzensen, N.; Bertrand, K.A.; Cook, L.S.; Friedenreich, C.M.; et al. Coffee consumption and risk of endometrial cancer: A pooled analysis of individual participant data in the Epidemiology of Endometrial Cancer Consortium (E2C2). Am. J. Clin. Nutr. 2022, 116, 1219–1228. [Google Scholar] [CrossRef] [PubMed]
- Salaroli, L.B.; Ferreira, J.R.S.; Prado, C.B.D.; De Podestá, O.P.G.; Carvalho, A.L.; Mercante, A.M.D.C.; Toporcov, T.N. Cumulative coffee consumption as a protective factor for head and neck cancer in Brazil. Nutr. Cancer 2023, 75, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Azzeh, F.S.; Hasanain, D.M.; Qadhi, A.H.; Ghafouri, K.J.; Azhar, W.F.; Ghaith, M.M.; Aldairi, A.F.; Almasmoum, H.A.; Assaggaf, H.M.; Alhussain, M.H.; et al. Consumption of food components of the mediterranean diet decreases the risk of breast cancer in the Makkah Region, Saudi Arabia: A case-control study. Front. Nutr. 2022, 9, 863029. [Google Scholar] [CrossRef] [PubMed]
- Barré, T.; Fontaine, H.; Ramier, C.; Di Beo, V.; Pol, S.; Carrieri, P.; Marcellin, F.; Cagnot, C.; Dorival, C.; Zucman-Rossi, J.; et al. Elevated coffee consumption is associated with a lower risk of elevated liver fibrosis biomarkers in patients treated for chronic hepatitis B (ANRS CO22 Hepather cohort). Clin. Nutr. 2022, 41, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Imatoh, T.; Sawada, N.; Yamaji, T.; Iwasaki, M.; Inoue, M.; Tsugane, S.; JPHC Study Group. Association between coffee consumption and risk of prostate cancer in Japanese men: A population-based cohort study in Japan. Cancer Epidemiol. Biomark. Prev. 2022, 31, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Rhee, J.; Lim, R.K.; Purdue, M.P. Coffee consumption and risk of renal cancer: A meta-analysis of cohort evidence. Cancer Causes Control. 2022, 33, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Şemen, S.; Mercan, S.; Yayla, M.; Açıkkol, M. Elemental composition of green coffee and its contribution to dietary intake. Food Chem. 2017, 215, 92–100. [Google Scholar] [CrossRef]
- Clifford, M.N.; Kerimi, A.; Williamson, G. Bioavailability and metabolism of chlorogenic qcids (acyl-quinic acids) in humans. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1299–1352. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Su, C.; Chen, X.; Wang, Q.; Jiao, W.; Luo, H.; Tang, J.; Wang, W.; Li, S.; Guo, S. Chlorogenic acids in cardiovascular disease: A review of dietary consumption, pharmacology, and pharmacokinetics. J. Agric. Food Chem. 2020, 68, 6464–6484. [Google Scholar] [CrossRef]
- Clifford, M.N.; Wight, J. The measurement of feruloylquinic acids and, caffeoylquinic acids in coffee beans. Development of the technique and its, preliminary application to green coffee beans. J. Sci. Food Agric. 1976, 27, 73–84. [Google Scholar] [CrossRef]
- Jeon, J.S.; Kim, H.T.; Jeong, I.H.; Hong, S.R.; Oh, M.S.; Yoon, M.H.; Shim, J.H.; Jeong, J.H.; Abd El-Aty, A.M. Contents of chlorogenic acids and caffeine in various coffee-related products. J. Adv. Res. 2019, 17, 85–94. [Google Scholar] [CrossRef]
- Sanlier, N.; Atik, A.; Atik, I. Consumption of green coffee and the risk of chronic diseases. Crit. Rev. Food Sci. Nutr. 2019, 59, 2573–2585. [Google Scholar] [CrossRef]
- Awwad, S.; Issa, R.; Alnsour, L.; Albals, D.; Al-Momani, I. Quantification of caffeine and chlorogenic acid in green and roasted coffee samples using HPLC-DAD and evaluation of the effect of degree of roasting on their levels. Molecules 2021, 26, 7502. [Google Scholar] [CrossRef]
- Alnsour, L.; Issa, R.; Awwad, S.; Albals, D.; Al-Momani, I. Quantification of total phenols and antioxidants in coffee samples of different origins and evaluation of the effect of degree of roasting on their levels. Molecules 2022, 27, 1591. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Pan, X.; Jiang, L.; Chu, Y.; Gao, S.; Jiang, X.; Zhang, Y.; Chen, Y. The biological activity mechanism of chlorogenic acid and its applications in food industry: A review. Front Nutr. 2022, 9, 943911. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Atanasov, A.G.; Li, Y.; Kumar, N.; Bishayee, A. Chlorogenic acid for cancer prevention and therapy: Current status on efficacy and mechanisms of action. Pharmacol. Res. 2022, 186, 106505. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Nagase, N.; Tsuji, A.; Taniguchi, K.; Kitagishi, Y.; Matsuda, S. Comprehension of the relationship between autophagy and reactive oxygen species for superior cancer therapy with histone deacetylase inhibitors. Oxygen 2021, 1, 22–31. [Google Scholar] [CrossRef]
- Yoshikawa, S.; Taniguchi, K.; Sawamura, H.; Ikeda, Y.; Tsuji, A.; Matsuda, S. Roles of reactive oxygen species and autophagy in the pathogenesis of of cisplatin-induced acute kidney injury. Oxygen 2022, 2, 317–326. [Google Scholar] [CrossRef]
- Cheng, D.; Zhang, X.; Tang, J.; Kong, Y.; Wang, X.; Wang, S. Chlorogenic acid protects against aluminum toxicity via MAPK/Akt signaling pathway in murine RAW264.7 macrophages. J. Inorg. Biochem. 2019, 190, 113–120. [Google Scholar] [CrossRef]
- Effect of chlorogenic acid on the physicochemical and functional properties of Coregonus peled myofibrillar protein through hydroxyl radical oxidation. Molecules 2019, 24, 3205. [CrossRef]
- Miao, M.; Xiang, L. Pharmacological action and potential targets of chlorogenic acid. Adv. Pharmacol. 2020, 87, 71–88. [Google Scholar] [CrossRef]
- Ludwig, I.A.; Clifford, M.N.; Lean, M.E.; Ashihara, H.; Crozier, A. Coffee: Biochemistry and potential impact on health. Food Funct. 2014, 5, 1695–1717. [Google Scholar] [CrossRef] [PubMed]
- Hoelzl, C.; Knasmüller, S.; Wagner, K.H.; Elbling, L.; Huber, W.; Kager, N.; Ferk, F.; Ehrlich, V.; Nersesyan, A.; Neubauer, O.; et al. Instant coffee with high chlorogenic acid levels protects humans against oxidative damage of macromolecules. Mol. Nutr. Food Res. 2010, 54, 1722–1733. [Google Scholar] [CrossRef] [PubMed]
- Agudelo-Ochoa, G.M.; Pulgarín-Zapata, I.C.; Velásquez-Rodriguez, C.M.; Duque-Ramírez, M.; Naranjo-Cano, M.; Quintero-Ortiz, M.M.; Lara-Guzmán, O.J.; Munoz-Durango, K. Coffee consumption increases the antioxidant capacity of plasma and has no effect on the lipid profile or vascular function in healthy adults in a randomized controlled trial. J. Nutr. 2016, 146, 524–531. [Google Scholar] [CrossRef]
- Melamed, I.; Kark, J.D.; Spirer, Z. Coffee and the immune system. Int. J. Immunopharmacol. 1990, 12, 129–134. [Google Scholar] [CrossRef]
- Chen, D.; Pan, D.; Tang, S.; Tan, Z.; Zhang, Y.; Fu, Y.; Lü, G.; Huang, Q. Administration of chlorogenic acid alleviates spinal cord injury via TLR4/NF-kappaB and p38 signaling pathway antiinflammatory activity. Mol. Med. Rep. 2018, 17, 1340–1346. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.S.; Satsu, H.; Bae, M.J.; Zhao, Z.; Ogiwara, H.; Totsuka, M.; Shimizu, M. Anti-inflammatory effect of chlorogenic acid on the IL-8 production in Caco-2 cells and the dextran sulphate sodium-induced colitis symptoms in C57BL/6 mice. Food Chem. 2015, 168, 167–175. [Google Scholar] [CrossRef]
- Chang, W.C.; Chen, C.H.; Lee, M.F.; Chang, T.; Yu, Y.M. Chlorogenic acid attenuates adhesion molecules upregulation in IL-1beta-treated endothelial cells. Eur. J. Nutr. 2010, 49, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Hebeda, C.B.; Bolonheis, S.M.; Nakasato, A.; Belinati, K.; Souza, P.D.; Gouvea, D.R.; Lopes, N.P.; Farsky, S.H. Effects of chlorogenic acid on neutrophil locomotion functions in response to inflammatory stimulus. J. Ethnopharmacol. 2011, 135, 261–269. [Google Scholar] [CrossRef]
- Fuentes, E.; Caballero, J.; Alarcón, M.; Rojas, A.; Palomo, I. Chlorogenic acid inhibits human platelet activation and thrombus formation. PLoS ONE 2014, 9, e90699. [Google Scholar] [CrossRef]
- Arab, L. Epidemiologic evidence on coffee and cancer. Nutr. Cancer. 2010, 62, 271–283. [Google Scholar] [CrossRef]
- Hanahan, D. Hallmarks of cancer: New dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Pavlova, N.N.; Zhu, J.; Thompson, C.B. The hallmarks of cancer metabolism: Still emerging. Cell Metab. 2022, 34, 355–377. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Castranova, V.; Ding, M. Inhibition of activator protein-1, NF-kappaB, and MAPKs and induction of phase 2 detoxifying enzyme activity by chlorogenic acid. J. Biol. Chem. 2005, 280, 27888–27895. [Google Scholar] [CrossRef]
- Yagasaki, K.; Miura, Y.; Okauchi, R.; Furuse, T. Inhibitory effects of chlorogenic acid and its related compounds on the invasion of hepatoma cells in culture. Cytotechnology 2000, 33, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Pavlica, S.; Gebhardt, R. Protective effects of ellagic and chlorogenic acids against oxidative stress in PC12 cells. Free Radic. Res. 2005, 39, 1377–1390. [Google Scholar] [CrossRef] [PubMed]
- Egeblad, M.; Werb, Z. New functions for the matrix metalloproteinases in cancer progression. Nat. Rev. Cancer 2002, 2, 161–174. [Google Scholar] [CrossRef]
- Gobin, E.; Bagwell, K.; Wagner, J.; Mysona, D.; Sandirasegarane, S.; Smith, N.; Bai, S.; Sharma, A.; Schleifer, R.; She, J.X. A pan-cancer perspective of matrix metalloproteases (MMP) gene expression profile and their diagnostic/prognostic potential. BMC Cancer 2019, 19, 581. [Google Scholar] [CrossRef]
- Huang, H. Matrix metalloproteinase-9 (MMP-9) as a cancer biomarker and MMP-9 biosensors: Recent advances. Sensors 2018, 18, 3249. [Google Scholar] [CrossRef]
- Jin, U.H.; Lee, J.Y.; Kang, S.K.; Kim, J.K.; Park, W.H.; Kim, J.G.; Moon, S.K.; Kim, C.H. A phenolic compound, 5-caffeoylquinic acid (chlorogenic acid), is a new type and strong matrix metalloproteinase-9 inhibitor: Isolation and identification from methanol extract of Euonymus A latus. Life Sci. 2005, 77, 2760–2769. [Google Scholar] [CrossRef]
- Cabral-Pacheco, G.A.; Garza-Veloz, I.; Castruita-De la Rosa, C. The roles of matrix metalloproteinases and their inhibitors in human diseases. Int. J. Mol. Sci. 2020, 21, 9739. [Google Scholar] [CrossRef]
- Belkaid, A.; Currie, J.C.; Desgagnés, J.; Annabi, B. The chemopreventive properties of chlorogenic acid reveal a potential new role for the microsomal glucose-6-phosphate translocase in brain tumor progression. Cancer Cell Int. 2006, 6, 7. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, S.; Nakanishi, A.; Wada, Y.; Kitagishi, Y. Roles of PI3K/AKT/PTEN pathway as a target for pharmaceutical therapy. Open Med. Chem. J. 2013, 7, 23–29. [Google Scholar] [CrossRef]
- Nakanishi, A.; Kitagishi, Y.; Ogura, Y.; Matsuda, S. The tumor suppressor PTEN interacts with p53 in hereditary cancer. Int. J. Oncol. 2014, 44, 1813–1819. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, S.; Ikeda, Y.; Murakami, M.; Nakagawa, Y.; Tsuji, A.; Kitagishi, Y. Roles of PI3K/AKT/GSK3 pathway involved in psychiatric illnesses. Diseases 2019, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Murakami, M.; Nakagawa, Y.; Tsuji, A.; Kitagishi, Y.; Matsuda, S. Diet induces hepatocyte protection in fatty liver disease via modulation of PTEN signaling. Biomed. Rep. 2020, 12, 295–302. [Google Scholar] [CrossRef]
- Ikeda, Y.; Nagase, N.; Tsuji, A.; Kitagishi, Y.; Matsuda, S. Neuroprotection by dipeptidyl-peptidase-4 inhibitors and glucagon-like peptide-1 analogs via the modulation of AKT-signaling pathway in Alzheimer’s disease. World J. Biol. Chem. 2021, 12, 104–113. [Google Scholar] [CrossRef]
- Matsuda, S.; Murakami, M.; Ikeda, Y.; Nakagawa, Y.; Tsuji, A.; Kitagishi, Y. Role of tumor suppressor molecules in genomic perturbations and damaged DNA repair involved in the pathogenesis of cancer and neurodegeneration. Biomed. Rep. 2020, 13, 10. [Google Scholar] [CrossRef]
- Murakami, M.; Ikeda, Y.; Nakagawa, Y.; Tsuji, A.; Kitagishi, Y.; Matsuda, S. Special bioactive compounds and functional foods may exhibit neuroprotective effects in patients with dementia. Biomed. Rep. 2020, 13, 1. [Google Scholar] [CrossRef]
- Kitagishi, Y.; Matsuda, S. Redox regulation of tumor suppressor PTEN in cancer and aging. Int. J. Mol. Med. 2013, 31, 511–515. [Google Scholar] [CrossRef]
- Minami, A.; Nakanishi, A.; Ogura, Y.; Kitagishi, Y.; Matsuda, S. Connection between tumor suppressor BRCA1 and PTEN in damaged DNA repair. Front. Oncol. 2014, 4, 318. [Google Scholar] [CrossRef]
- Refolo, M.G.; Lippolis, C.; Carella, N.; Cavallini, A.; Messa, C.; D’Alessandro, R. Chlorogenic Acid Improves the Regorafenib Effects in Human Hepatocellular Carcinoma Cells. Int. J. Mol. Sci. 2018, 19, 1518. [Google Scholar] [CrossRef] [PubMed]
- Chlorogenic acid inhibits proliferation and induces apoptosis in A498 human kidney cancer cells via inactivating PI3K/Akt/mTOR signalling pathway. J. Pharm. Pharmacol. 2019, 71, 1100–1109. [CrossRef] [PubMed]
- Villota, H.; Santa-González, G.A.; Uribe, D.; Henao, I.C.; Arroyave-Ospina, J.C.; Barrera-Causil, C.J.; Pedroza-Díaz, J. Modulatory effect of chlorogenic acid and coffee extracts on Wnt/β-catenin pathway in colorectal cancer cells. Nutrients 2022, 14, 4880. [Google Scholar] [CrossRef] [PubMed]
- Villota, H.; Moreno-Ceballos, M.; Santa-González, G.A.; Uribe, D.; Castañeda, I.C.; Preciado, L.M.; Pedroza-Díaz, J. Biological impact of phenolic compounds from coffee on colorectal cancer. Pharmaceuticals 2021, 14, 761. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ping, Z.; Xuemei, G.; Hongjuan, M.; Yi, H.; Xiaoli, L.; Zhongxiang, Z. Chlorogenic acid regulates the proliferation and migration of high-grade serous ovarian cancer cells through modulating the miR199a5p/DDR1 axis. Acta Biochim. Pol. 2022, 69, 855–864. [Google Scholar] [CrossRef]
- Chen, Y.K.; Ngoc, N.T.M.; Chang, H.W.; Su, Y.F.; Chen, C.H.; Goan, Y.G.; Chen, J.Y.; Tung, C.W.; Hour, T.C. Chlorogenic acid inhibition of esophageal squamous cell carcinoma metastasis via EGFR/p-Akt/Snail signaling pathways. Anticancer Res. 2022, 42, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Feng, Y.; Li, Y.; Hu, Y.; Zhang, Q.; Huang, Y.; Shi, K.; Ran, C.; Hou, J.; Zhou, G.; et al. Chlorogenic acid decreases malignant characteristics of hepatocellular carcinoma cells by inhibiting DNMT1 expression. Front. Pharmacol. 2020, 11, 867. [Google Scholar] [CrossRef] [PubMed]
- Changizi, Z.; Moslehi, A.; Rohani, A.H.; Eidi, A. Chlorogenic acid inhibits growth of 4T1 breast cancer cells through involvement in Bax/Bcl2 pathway. J. Cancer Res. Ther. 2020, 16, 1435–1442. [Google Scholar] [CrossRef]
- Huang, S.; Wang, L.L.; Xue, N.N.; Li, C.; Guo, H.H.; Ren, T.K.; Zhan, Y.; Li, W.B.; Zhang, J.; Chen, X.G.; et al. Chlorogenic acid effectively treats cancers through induction of cancer cell differentiation. Theranostics 2019, 9, 6745–6763. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, K.; Izawa, Y.; Onodera, D.; Tagami, M. Chlorogenic acid regulates apoptosis and stem cell marker-related gene expression in A549 human lung cancer cells. Mol. Cell Biochem. 2018, 441, 9–19. [Google Scholar] [CrossRef]
- Yin, X.; He, X.; Wu, L.; Yan, D.; Yan, S. Chlorogenic acid, the main antioxidant in coffee, reduces radiation-induced apoptosis and DNA damage via NF-E2- related factor 2 (Nrf2) activation in hepatocellular carcinoma. Oxid. Med. Cell Longev. 2022, 2022, 4566949. [Google Scholar] [CrossRef]
- Bartolomeu, A.R.; Romualdo, G.R.; Lisón, C.G.; Besharat, Z.M.; Corrales, J.A.M.; Chaves, M.Á.G.; Barbisan, L.F. Caffeine and chlorogenic acid combination attenuate early-stage chemically induced colon carcinogenesis in mice: Involvement of oncomiR miR-21a-5p. Int. J. Mol. Sci. 2022, 23, 6292. [Google Scholar] [CrossRef]
- Chen, Y.; Abe, S.K.; Inoue, M.; Yamaji, T.; Iwasaki, M.; Nomura, S.; Hashizume, M.; Tsugane, S.; Sawada, N. JPHC Study Group. Green tea and coffee consumption and risk of kidney cancer in Japanese adults. Sci. Rep. 2022, 12, 20274. [Google Scholar] [CrossRef]
- Schuster, C.; Wolpert, N.; Moustaid-Moussa, N.; Gollahon, L.S. Combinatorial effects of the natural products Arctigenin, chlorogenic acid, and cinnamaldehyde commit oxidation assassination on breast cancer cells. Antioxidants 2022, 11, 591. [Google Scholar] [CrossRef]
- Socodato, R.; Portugal, C.C.; Canedo, T.; Domith, I.; Oliveira, N.A.; Paes-de-Carvalho, R.; Relvas, J.B.; Cossenza, M. c-Src deactivation by the polyphenol 3-O-caffeoylquinic acid abrogates reactive oxygen species-mediated glutamate release from microglia and neuronal excitotoxicity. Free Radic. Biol. Med. 2015, 79, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Mikami, Y.; Yamazawa, T. Chlorogenic acid, a polyphenol in coffee, protects neurons against glutamate neurotoxicity. Life Sci. 2015, 139, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Taram, F.; Winter, A.N.; Linseman, D.A. Neuroprotection comparison of chlorogenic acid and its metabolites against mechanistically distinct cell death-inducing agents in cultured cerebellar granule neuron. Brain Res. 2016, 1648, 69–80. [Google Scholar] [CrossRef]
- Xue, N.; Zhou, Q.; Ji, M.; Jin, J.; Lai, F.; Chen, J.; Zhang, M.; Jia, J.; Yang, H.; Zhang, J.; et al. Chlorogenic acid inhibits glioblastoma growth through repolarizating macrophage from M2 to M1 phenotype. Sci. Rep. 2017, 7, 39011. [Google Scholar] [CrossRef]
- Fukuyama, K.; Kakio, S.; Nakazawa, Y.; Kobata, K.; Funakoshi-Tago, M.; Suzuki, T.; Tamura, H. Roasted Coffee Reduces β-Amyloid Production by Increasing Proteasomal β-Secretase Degradation in Human Neuroblastoma SH-SY5Y Cells. Mol. Nutr. Food Res. 2018, 62, e1800238. [Google Scholar] [CrossRef]
- Unno, K.; Taguchi, K.; Hase, T.; Meguro, S.; Nakamura, Y. Coffee polyphenol, chlorogenic acid, suppresses brain aging and its effects are enhanced by milk fat globule membrane components. Int. J. Mol. Sci. 2022, 23, 5832. [Google Scholar] [CrossRef]
- Matsuda, S.; Nakagawa, Y.; Tsuji, A.; Kitagishi, Y.; Nakanishi, A.; Murai, T. Implications of PI3K/AKT/PTEN signaling on superoxide dismutases expression and in the pathogenesis of Alzheimer’s disease. Diseases 2018, 6, 28. [Google Scholar] [CrossRef]
- Londzin, P.; Zamora, M.; Kąkol, B.; Taborek, A.; Folwarczna, J. Potential of caffeine in Alzheimer’s disease-a review of experimental studies. Nutrients 2021, 13, 537. [Google Scholar] [CrossRef] [PubMed]
- Nakano, N.; Matsuda, S.; Ichimura, M.; Minami, A.; Ogino, M.; Murai, T.; Kitagishi, Y. PI3K/AKT signaling mediated by G protein-coupled receptors is involved in neurodegenerative Parkinson’s disease. Int. J. Mol. Med. 2017, 39, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Noyce, A.J.; Bestwick, J.P.; Silveira-Moriyama, L.; Hawkes, C.H.; Giovannoni, G.; Lees, A.J.; Schrag, A. Meta-analysis of early non- motor features and risk factors for Parkinson disease. Ann. Neurol. 2012, 72, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhao, Y.; Zheng, X.; Chen, Q.; Zhang, X. Chlorogenic acid alters the biological characteristics of basophil granulocytes by affecting the fluidity of the cell membrane and triggering pseudoallergic reactions. Int. J. Mol. Med. 2013, 32, 1273–1280. [Google Scholar] [CrossRef] [PubMed]
- Bonarska-Kujawa, D.; Cyboran-Mikołajczyk, S.; Kleszczyńska, H. Molecular mechanism of action of chlorogenic acid on erythrocyte and lipid membranes. Mol. Membr. Biol. 2015, 32, 46–54. [Google Scholar] [CrossRef]
- Cejas, J.P.; Rosa, A.S.; Nazareno, M.A.; Disalvo, E.A.; Frias, M.A. Interaction of chlorogenic acid with model lipid membranes and its influence on antiradical activity. Biochim. Biophys. Acta Biomembr. 2021, 1863, 183484. [Google Scholar] [CrossRef]
- Escribá, P.V.; Busquets, X.; Inokuchi, J.; Balogh, G.; Török, Z.; Horváth, I.; Harwood, J.L.; Vígh, L. Membrane lipid therapy: Modulation of the cell membrane composition and structure as a molecular base for drug discovery and new disease treatment. Prog. Lipid Res. 2015, 59, 38–53. [Google Scholar] [CrossRef]
- Domínguez Díaz, L.; Fernández-Ruiz, V.; Cámara, M. The frontier between nutrition and pharma: The international regulatory framework of functional foods, food supplements and nutraceuticals. Crit. Rev. Food Sci. Nutr. 2020, 60, 1738–1746. [Google Scholar] [CrossRef]
- Vega, E.N.; García-Herrera, P.; Ciudad-Mulero, M.; Dias, M.I.; Matallana-González, M.C.; Cámara, M.; Tardío, J.; Molina, M.; Pinela, J.C.S.P.; Pires, T.; et al. Wild sweet cherry, strawberry and bilberry as underestimated sources of natural colorants and bioactive compounds with functional properties. Food Chem. 2023, 414, 135669. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Effect | Mechanism | Cell | Reference |
|---|---|---|---|
| Anti-inflammatory effects | |||
| Adhesion molecule | ICAM-1, VCAM-1 | vascular endothelial cells | [48] |
| Chemotaxis | CD62L | neutrophil | [49] |
| Leukocyte rolling | CD62P | platelet | [50] |
| Anti-cancer effects | |||
| Proliferation | hepatoma | [56] | |
| Invasion | hepatoma | [57] | |
| MMP activity | MMP-9 | hepatoma | [61] |
| MMP activity | MMP-2 | glioma | [63] |
| Proliferation | PI3K/Akt/mTORC | hepatocellular carcinoma | [73] |
| Apoptosis | PI3K/Akt/mTORC | kidney cancer | [74] |
| Signaling | Wnt/β-catenin | colon cancer | [75] |
| Viability, migration | colorectal cancer | [76] | |
| Migration | DDR1 | ovarian cancer | [77] |
| Invasion | Akt | squamous cell carcinoma | [78] |
| Invasion | ERK, MMP-2/9 | hepatic cancer | [79] |
| Apoptosis | p53 | breast cancer | [80] |
| Apoptosis | p21 | breast cancer | [81] |
| Apoptosis | JNK | lung cancer | [82] |
| Apoptosis | Nrf2 | hepatocellular carcinoma | [83] |
| Carcinogenesis | mi-21a-5p | colon cancer | [84] |
| Neuroprotective effects | |||
| Glutamine release | c-Src | microglia | [87] |
| Glutamine release | neuron | [88] | |
| Cell viability | neuron | [89,90] | |
| Neurodegeneration | amyloid-β | neuron | [91] |
| Brain aging suppression | CREB | microglia | [92] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murai, T.; Matsuda, S. The Chemopreventive Effects of Chlorogenic Acids, Phenolic Compounds in Coffee, against Inflammation, Cancer, and Neurological Diseases. Molecules 2023, 28, 2381. https://doi.org/10.3390/molecules28052381
Murai T, Matsuda S. The Chemopreventive Effects of Chlorogenic Acids, Phenolic Compounds in Coffee, against Inflammation, Cancer, and Neurological Diseases. Molecules. 2023; 28(5):2381. https://doi.org/10.3390/molecules28052381
Chicago/Turabian StyleMurai, Toshiyuki, and Satoru Matsuda. 2023. "The Chemopreventive Effects of Chlorogenic Acids, Phenolic Compounds in Coffee, against Inflammation, Cancer, and Neurological Diseases" Molecules 28, no. 5: 2381. https://doi.org/10.3390/molecules28052381
APA StyleMurai, T., & Matsuda, S. (2023). The Chemopreventive Effects of Chlorogenic Acids, Phenolic Compounds in Coffee, against Inflammation, Cancer, and Neurological Diseases. Molecules, 28(5), 2381. https://doi.org/10.3390/molecules28052381