
Evaluation of the Potential of a Lectin Extracted from Polygonumpersicaria L. as a Biorational Agent against Sitophilusoryzae L.




Abstract
1. Introduction
2. Results
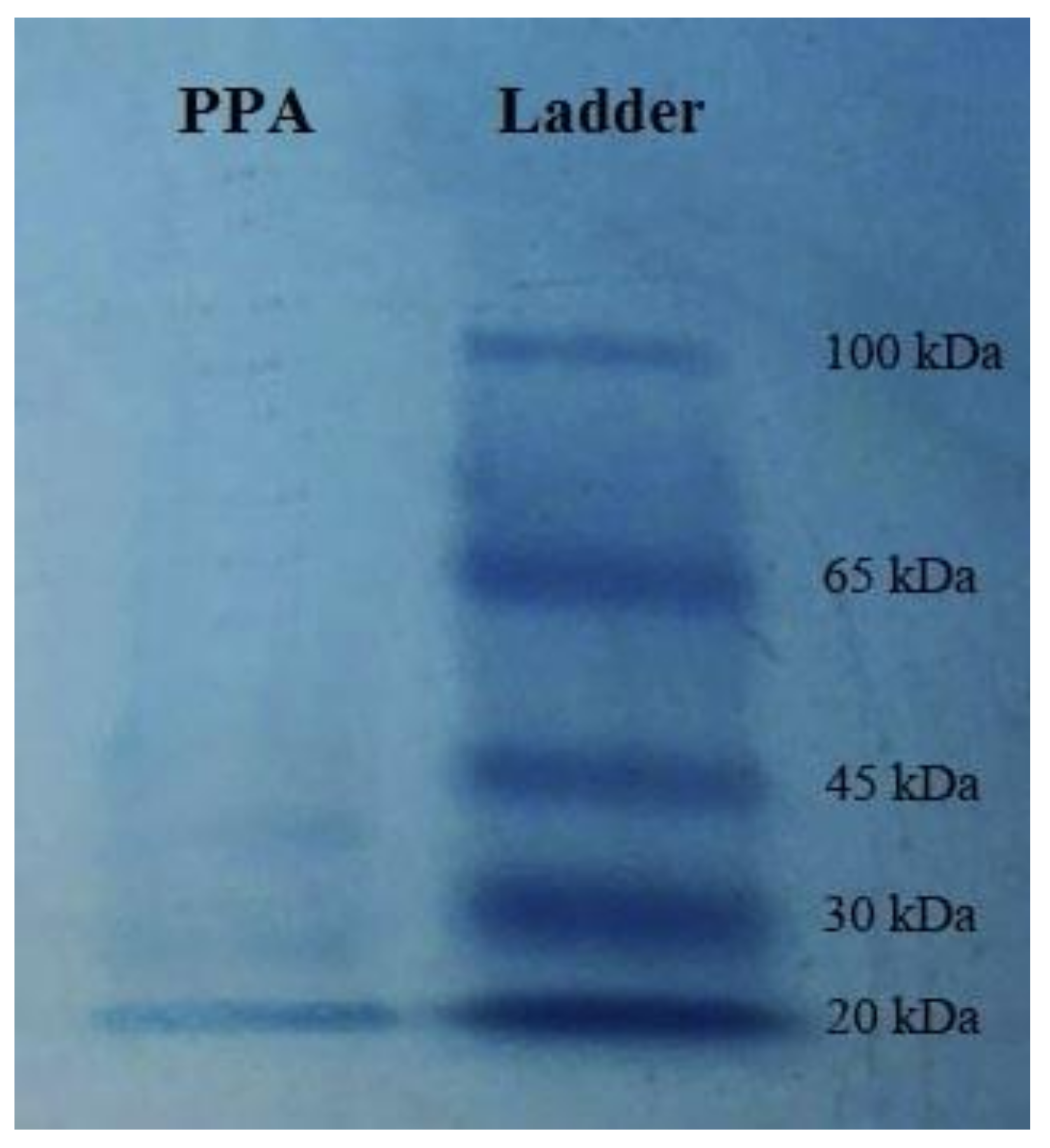
2.1. Purification of the Lectin
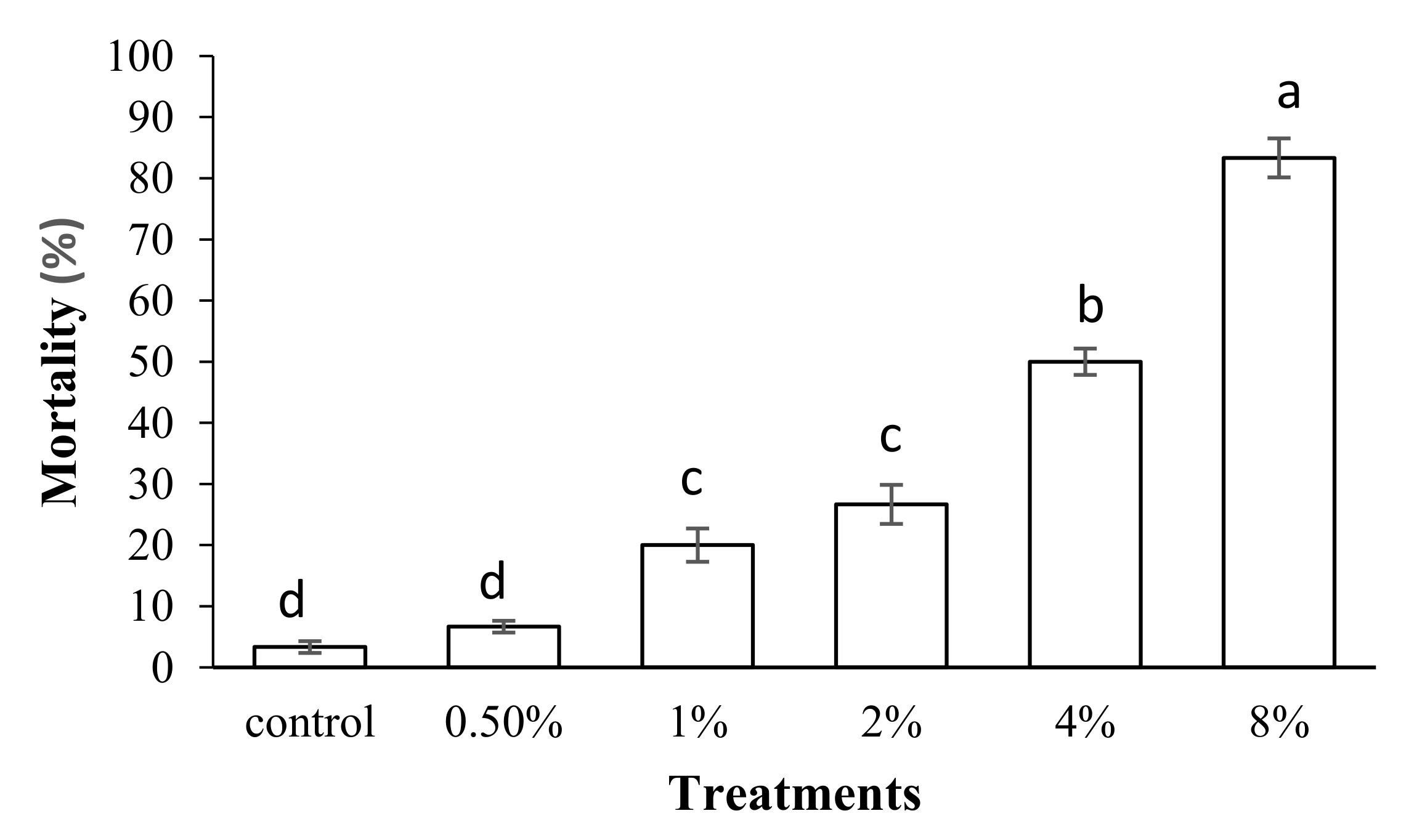
2.2. Bioassay
2.3. Effect on Digestive Enzymes of S. oryzae
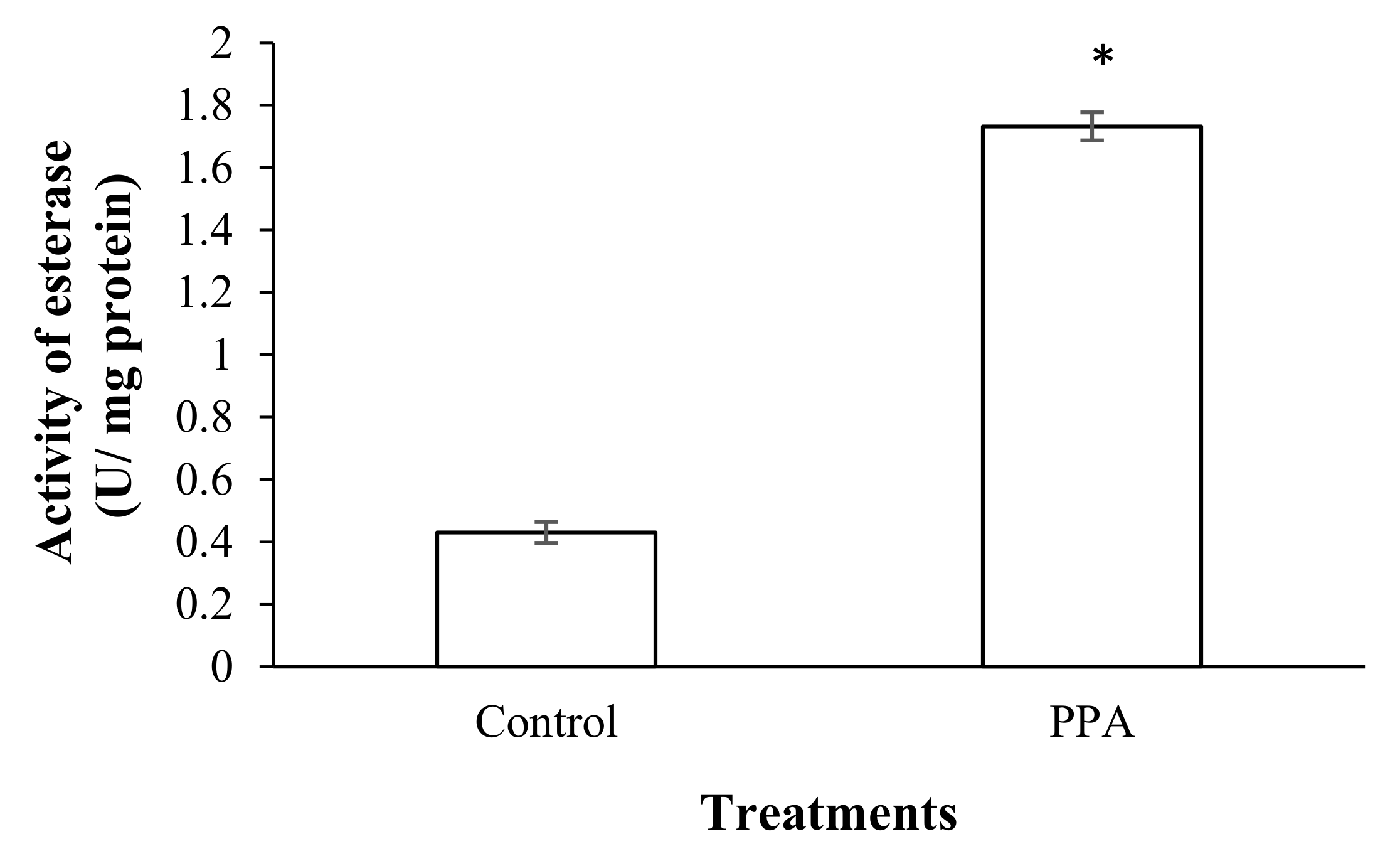
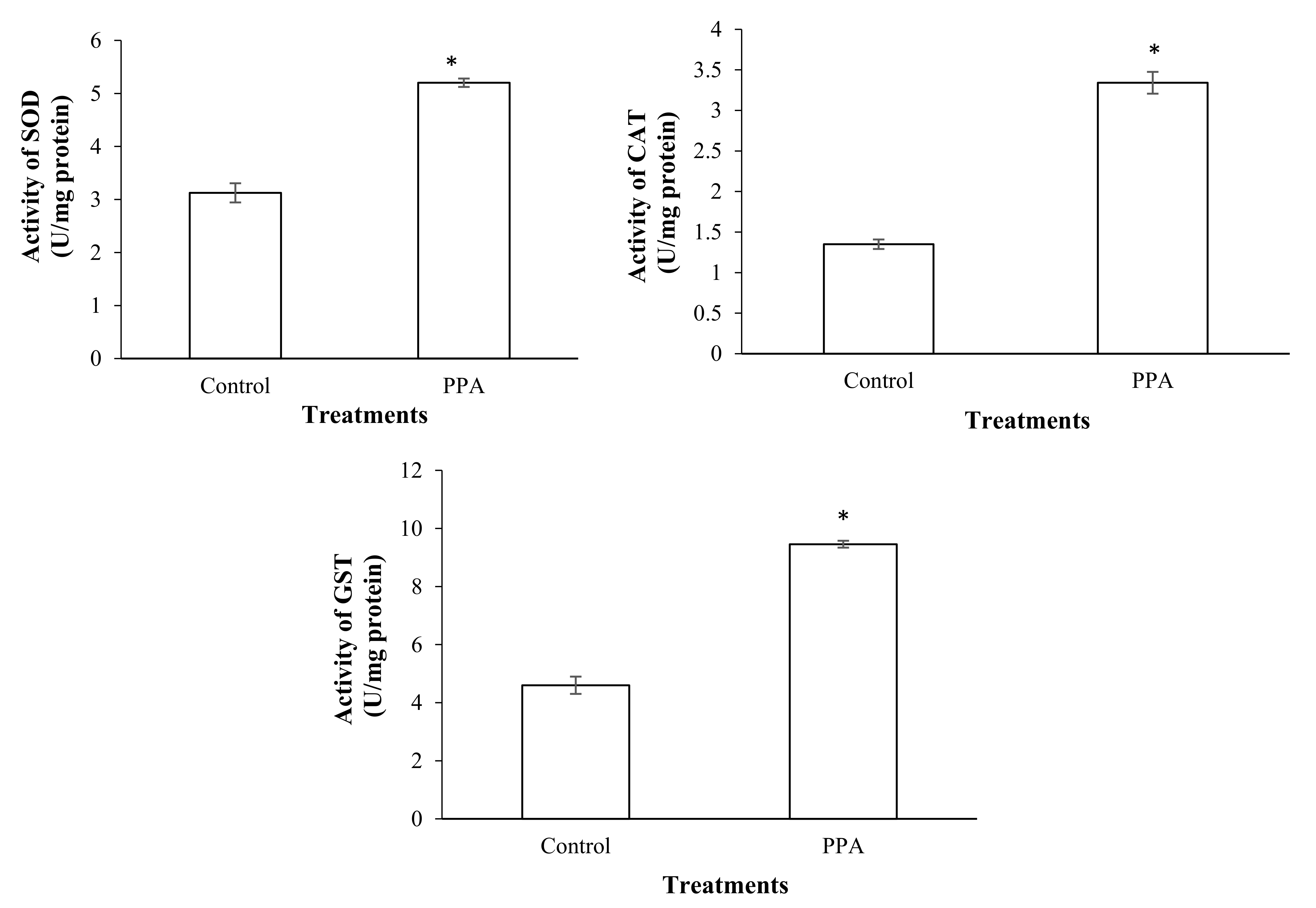
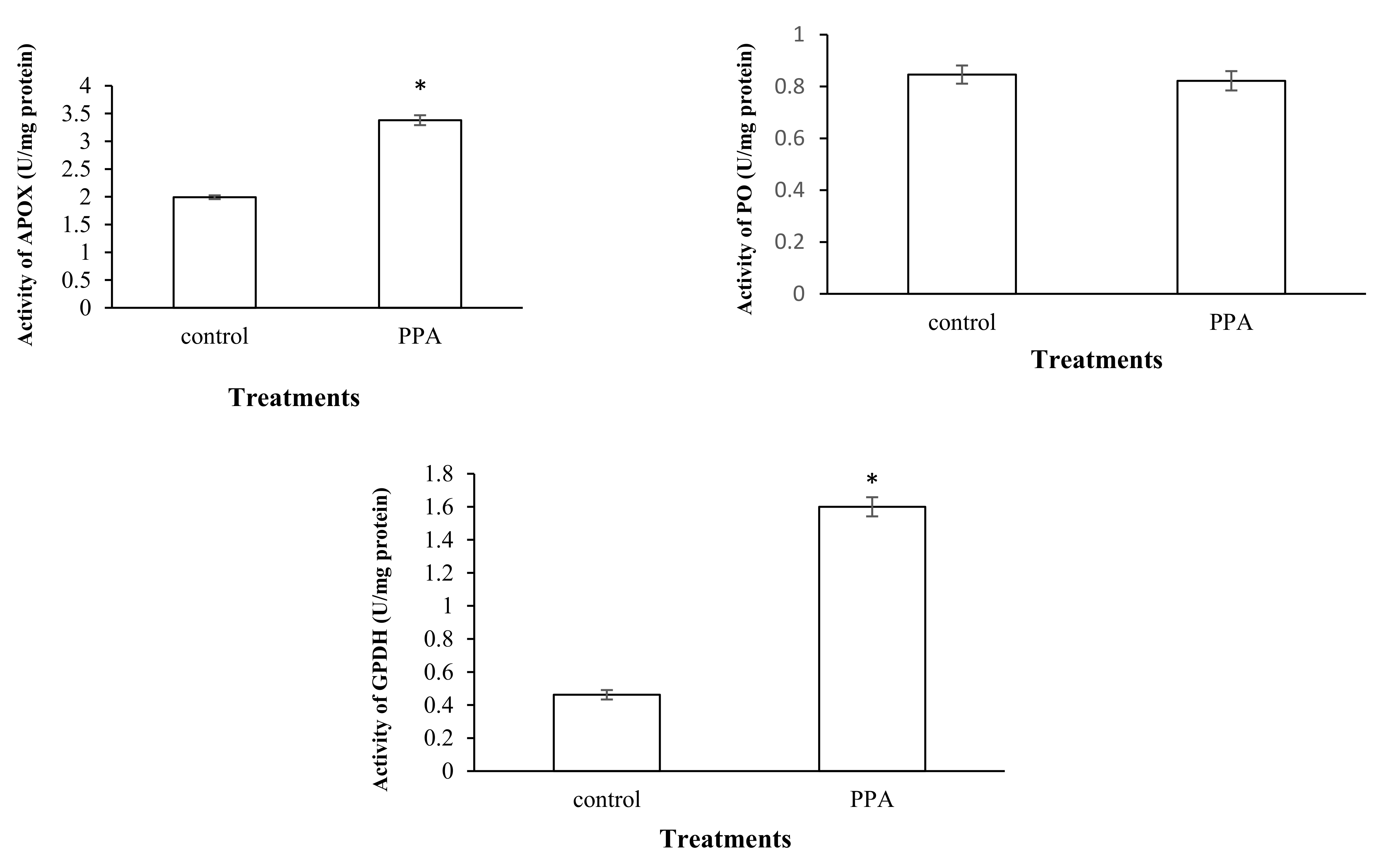
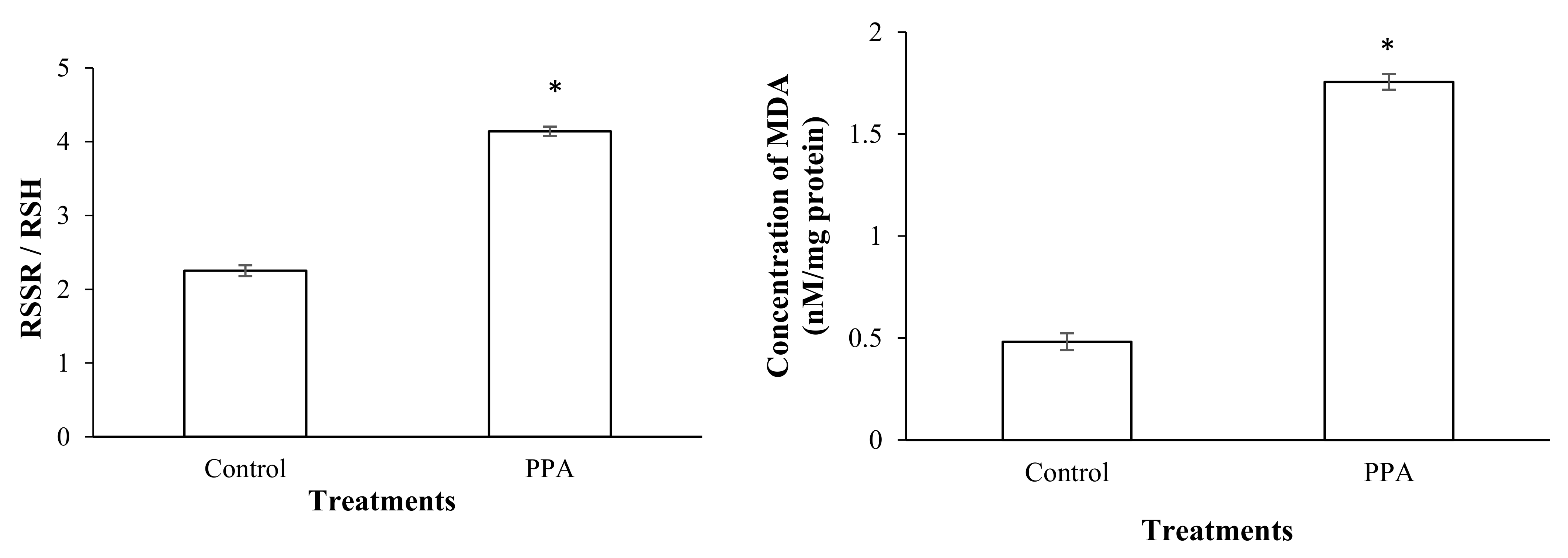
2.4. Effect on Detoxifying and Antioxidant Enzymes of S. oryzae
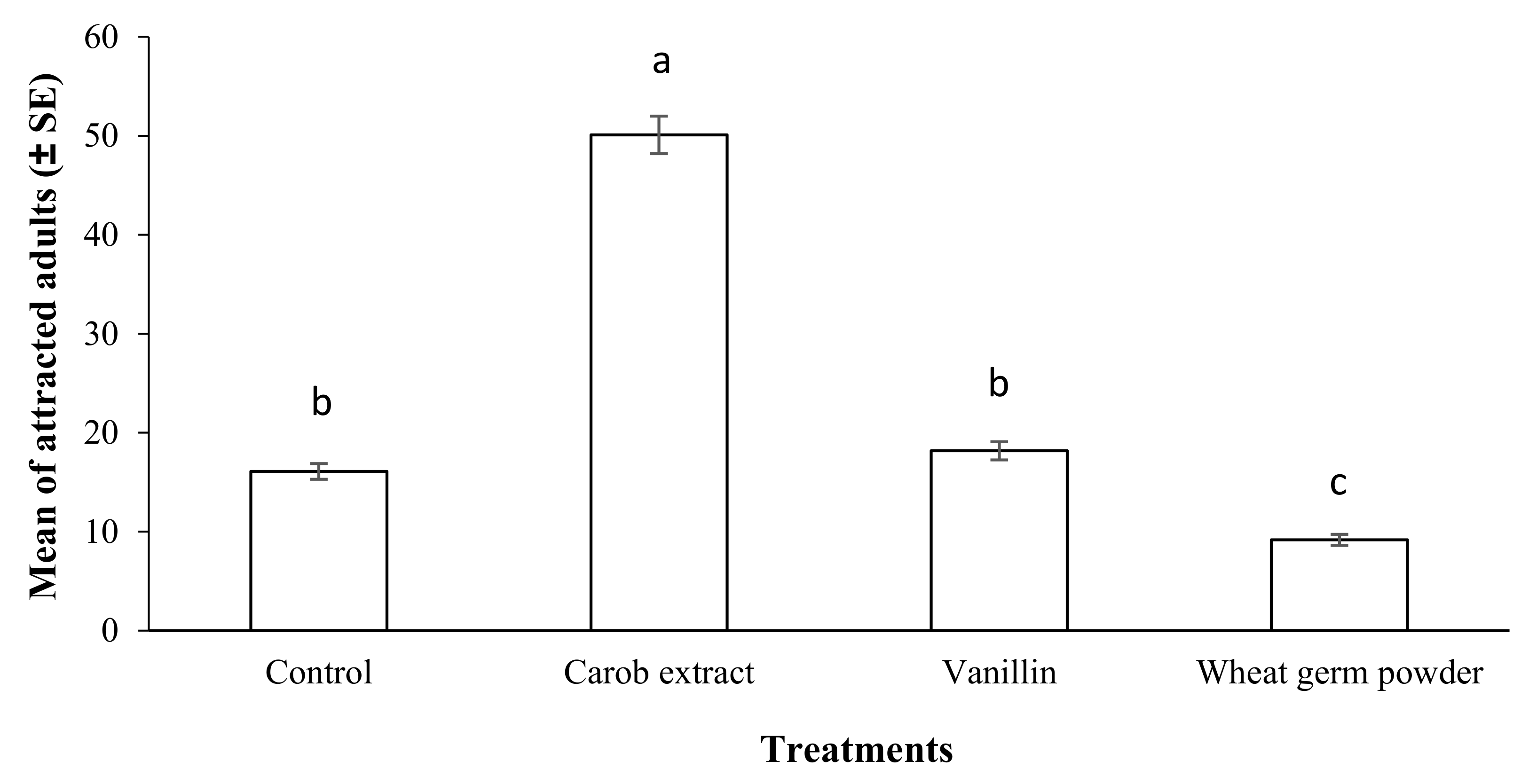
2.5. Olfactory Assay
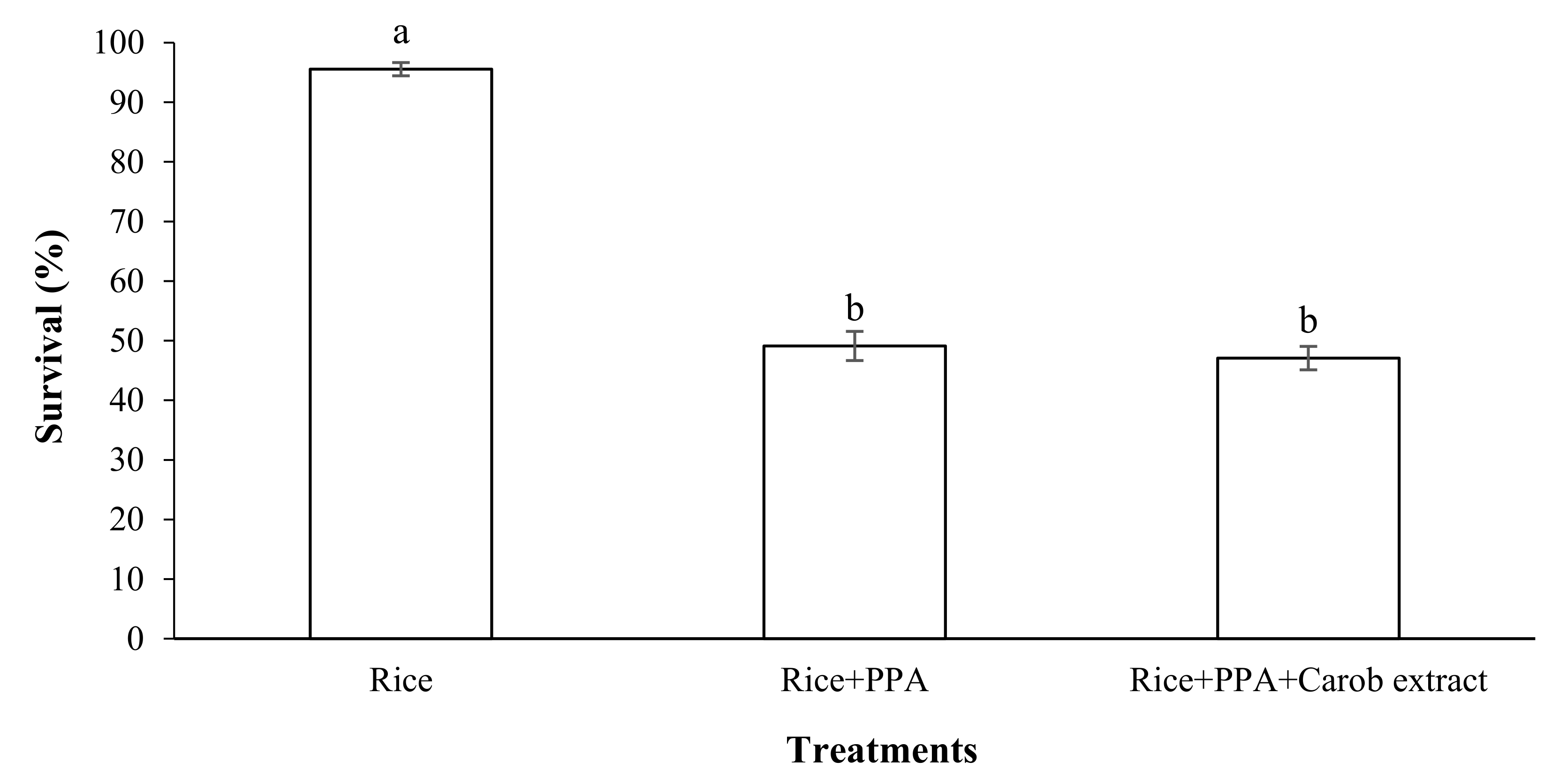
2.6. Effect of a Mixture of PPA and Food Attractant on the Survival of S. oryzae
3. Discussion
4. Materials and Methods
4.1. Insect Rearing
4.2. Purification of the Lectin
4.3. Bioassay
4.4. Effect on Digestive Enzymes Activities of S. oryzae
4.4.1. α-Amylase
4.4.2. α- and β-Glucosidases
4.4.3. Triacylglycerol (TAG)-Lipase
4.4.4. Serine Proteinases
4.4.5. Exopeptidases
4.5. Effect on Detoxifying and Antioxidant Enzymes of S. oryzae
4.5.1. Determination of General Esterase Activity
4.5.2. Glutathione S-Transferase
4.5.3. Catalase
4.5.4. Superoxide Dismutase
4.5.5. Peroxidase
4.5.6. Ascorbate Peroxidase
4.5.7. Glucose 6-Phosphate Dehydrogenase (GPDH)
4.5.8. RSSR/RSH Ratio
4.5.9. Malondialdehyde (MDA)
4.5.10. Protein Assay
4.6. Olfactometry
4.7. Effect of a Mixture of PPA and Food Attractant on S. oryzae Mortality
4.8. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Napoleão, T.H.; Belmonte, B.R.; Pontual, E.V.; Albuquerque, L.P.; Sa, R.A.; Paiva, L.M.; Coelho, L.C.B.B.; Paiva, P.M.G. Deleterious effects of Myracrodruon urundeuva leaf extract and lectin on the maize weevil, Sitophilus zeamais (Coleoptera, Curculionidae). J. Stored Prod. Res. 2013, 54, 26–33. [Google Scholar] [CrossRef]
- Yuan, Y.; Chen, C.; Zheng, C.; Wang, X.; Yang, G.; Wang, Q.; Zhang, Z. Residue of chlorpyrifos and cypermethrin in vegetables and probabilistic exposure assessment for consumers in Zhejiang Province, China. Food Control 2014, 36, 63–68. [Google Scholar] [CrossRef]
- Meyer-Baron, M.; Knapp, G.; Schäper, M.; van Thriel, C. Meta-analysis on occupational exposure to pesticides–neurobehavioral impact and dose–response relationships. Environ. Res. 2015, 136, 234–245. [Google Scholar] [CrossRef] [PubMed]
- Hertlein, M.B.; Thompson, G.D.; Subramanyam, B.; Athanassiou, C.G. Spinosad: A new natural product for stored grain protection. J. Stored Prod. Res. 2011, 47, 131–146. [Google Scholar] [CrossRef]
- Boyer, S.; Zhangand, H.; Lempérière, G. A review of control methods and resistance mechanisms in stored-product insects. Bull. Entomol. Res. 2012, 102, 213–229. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.; Harrington, P.; Turnbull, G. Neonicotinoid concentrations in arable soils after seed treatment applications in preceding years. Pest Manag. Sci. 2014, 70, 1780–1784. [Google Scholar] [CrossRef]
- Athanassiou, C.G.; Buchelos, C.T. Detection of stored-wheat beetle species and estimation of population density using unbaited probe traps and grain trier samples. Entomol. Exp. Appl. 2001, 98, 67–78. [Google Scholar] [CrossRef]
- Rojht, H.; Horvat, A.; Athanassiou, C.G.; Vayias, B.J.; Tomanović, Ž.; Trdan, S. Impact of geochemical composition of diatomaceous earth on its insecticidal activity against adults of Sitophilus oryzae (L.) (Coleoptera: Curculionidae). J. Pest Sci. 2010, 83, 429–436. [Google Scholar] [CrossRef]
- Koutsaviti, A.; Antonopoulou, V.; Vlassi, A.; Antonatos, S.; Michaelakis, A.; Papachristos, D.P.; Tzakou, O. Chemical composition and fumigant activity of essential oils from six plant families against Sitophilus oryzae (Col: Curculionidae). J. Pest Sci. 2018, 91, 873–886. [Google Scholar] [CrossRef]
- Benhalima, H.; Chaudhry, M.Q.; Mills, K.A.; Price, N.R. Phosphine resistance in stored-product insects collected from various grain storage facilities in Morocco. J. Stored Prod. Res. 2004, 40, 241–249. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Collins, P.J.; Duong, T.M.; Schlipalius, D.I.; Ebert, P.R. Genetic conservation of phosphine resistance in the rice Weevil Sitophilus oryzae (L.). J. Hered. 2016, 107, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, D.; Kar, A.; Giri, S.K. Insect pest management in stored pulses: An overview. Food Bioprocess Tech. 2015, 8, 239–265. [Google Scholar] [CrossRef]
- Daglish, G.J.; Nayak, M.K.; Pavic, H. Phosphine resistance in Sitophilus oryzae (L.) from eastern Australia: Inheritance, fitness and prevalence. J. Stored Prod. Res. 2014, 59, 237–244. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Collins, P.J.; Ebert, P.R. Inheritance and Characterization of Strong Resistance to Phosphine in Sitophilus oryzae (L.). PLoS ONE 2015, 10, e0124335. [Google Scholar] [CrossRef]
- Holloway, J.C.; Falk, M.G.; Emery, R.N.; Collins, P.J.; Nayak, M.K. Resistance to phosphine in Sitophilus oryzae in Australia: A national analysis of trends and frequencies over time and geographical spread. J. Stored Prod. Res. 2016, 69, 129–137. [Google Scholar] [CrossRef]
- Kim, K.; Yang, J.O.; Sung, J.Y.; Lee, J.Y.; Park, J.S.; Lee, H.S.; Lee, B.H.; Ren, Y.; Lee, D.W.; Lee, S.E. Minimization of energy transduction confers resistance to phosphine in the rice weevil, Sitophilus oryzae. Sci. Rep. 2019, 9, 14605. [Google Scholar] [CrossRef] [PubMed]
- Wakil, W.; Kavallieratos, N.G.; Usman, M.; Gulzar, S.; El-Shafie, H.A.F. Detection of Phosphine Resistance in Field Populations of Four Key Stored-Grain Insect Pests in Pakistan. Insects 2021, 12, 288. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [PubMed]
- Regnault-Roger, C.; Philogène, B.J. Past and current prospects for the use of botanicals and plant allelochemicals in integrated pest management. Pharm. Biol. 2008, 46, 41–52. [Google Scholar] [CrossRef]
- Kumar, P.; Mishra, S.; Malik, A.; Satya, S. Repellent, larvicidal and pupicidal properties of essential oils and their formulations against the housefly, Musca domestica. Med. Vet. Entomol. 2011, 25, 302–310. [Google Scholar] [CrossRef]
- Vandenborre, G.; Van Damme, E.J.M.; Smagghe, G. Natural products: Plant lectins as important tools in controlling pest insects. In Biorational Control of Arthropod Pests: Application and Resistance Management; Ishaaya, I., Horowitz, A.R., Eds.; Springer Science: Berlin, Germany, 2009; pp. 163–187. [Google Scholar]
- Michiels, K.; Van Damme, E.J.M.; Smagghe, G. Plant-insect interactions: What can we learn from plant lectins? Arch. Insect Biochem. Physiol. 2010, 73, 193–212. [Google Scholar] [CrossRef] [PubMed]
- Van Damme, E.J.M.; Lannoo, N.; Peumans, W.J. Plant lectins. Adv. Bot. Res. 2008, 48, 107–209. [Google Scholar] [CrossRef]
- Macedo, M.L.; Damico, D.C.; Freire, M.D.; Toyama, M.H.; Marangoni, S.; Novello, J.C. Purification and characterization of an N-acetylglucosamine-binding lectin from Koelreuteria paniculata seeds and its effect on the larval development of Callosobruchus maculatus (Coleoptera: Bruchidae) and Anagasta kuehniella (Lepidoptera: Pyralidae). J. Agric. Food Chem. 2003, 51, 2980–2986. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, A.; Van Damme, E.J.M.; Peumans, W.J.; Smagghe, G. Deterrent activity of plant lectins on cowpea weevil Callosobruchus maculatus (F.) oviposition. Phytochemistry 2006, 67, 2078–2084. [Google Scholar] [CrossRef][Green Version]
- Coelho, M.B.; Marangoni, S.; Macedo, M.L.R. Insecticidal action of Annona coriacea lectin against the flour moth Anagasta kuehniella and the rice moth Corcyra cephalonica (Lepidoptera: Pyralidae). Comp. Biochem. C Physiol. 2007, 146, 406–414. [Google Scholar] [CrossRef]
- Macedo, M.L.R.; Freire, M.G.M.; Novello, J.C.; Marangoni, S. Insecticidal action of Bauhinia monandra leaf lectin (BmoLL) against Anagasta kuehniella (Lepidoptera: Pyralidae), Zabrotes subfasciatus and Callosobruchus maculates (Coleoptera: Bruchidae). Comp. Biochem. Physiol. A 2007, 146, 486–498. [Google Scholar] [CrossRef]
- Oliveira, C.F.R.; Luz, L.A.; Paiva, P.M.G.; Coelho, L.C.B.B.; Marangoni, S.; Macedo, M.L.R. Evaluation of seed coagulant Moringa oleifera lectin (cMoL) as a bioinsecticidal tool with potential for the control of insects. Process Biochem. 2011, 46, 498–504. [Google Scholar] [CrossRef]
- Powell, K.S.; Spence, J.; Bharathi, M.; Gatehouse, J.A.; Gatehouse, A.M.R. Immunohistochemical and developmental studies to elucidate the mechanism of action of the snowdrop lectin on the rice brown planthopper, Nilaparvata lugens. J. Insect Physiol. 1998, 44, 529–539. [Google Scholar] [CrossRef]
- Zhu-Salzman, K.; Shade, R.E.; Koiwa, H.; Salzman, R.A.; Narasimhan, M.; Bressan, R.A.; Hasegawa, P.M.; Murdock, L.L. Carbohydrate binding and resistance to proteolysis control insecticidal activity of Griffonia simplicifolia lectin II. Proc. Natl. Acad. Sci. USA 1998, 95, 15123–15128. [Google Scholar] [CrossRef]
- Lima, J.E.; Sampaio, A.L.; Henriques, M.G.; Barja-Fidalgo, C. Lymphocyte activation and cytokine production by Pisum sativum agglutinin (PSA) in vivo and in vitro. Immunopharmacology 1999, 41, 147–155. [Google Scholar] [CrossRef]
- Zhu-Salzman, K.; Salzman, R.A. Functional mechanics of the plant defensive Griffonia simplicifolia lectin II: Resistance to proteolysis is independent of glycoconjugate binding in the insect gut. J. Econ. Entomol. 2001, 94, 1280–1284. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, V.; Hajizadeh, J.; Zibaee, A.; Jalali Sendi, J. Effect of Polygonum persicaria (Polygonales: Polygonaceae) extracted agglutinin on life table and antioxidant responses in Helicoverpa armigera (Lepidoptera: Noctuidae) larvae. J. Econ. Entomol. 2018, 111, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Dhang, P. Innovations in insect baiting and its role in reducing insecticide load in urban pest control. Int. Pest Control 2016, 58, 210–212. [Google Scholar]
- Weinzierl, R.; Henn, T.; Koehler, P.G.; Tucker, C.L. Insect attractants and traps. ENY277. 2005. Available online: http://ufdcimages.uflib.ufl (accessed on 15 December 2021).
- El-Sayed, A.M.; Suckling, D.M.; Byers, J.A.; Jang, E.B.; Wearing, C.H. Potential of “Lure and Kill” in Long-Term Pest Management and Eradication of Invasive Species. J. Econ. Entomol. 2009, 102, 815–835. [Google Scholar] [CrossRef] [PubMed]
- Collins, L.E.; Wakefield, M.E.; Chambers, J.; Cox, P.D. Progress towards a multi-species lure: Comparison of behavioural bioassay methods for multi-species attractants against three pests of stored grain. J. Stored Prod. Res. 2004, 40, 341–353. [Google Scholar] [CrossRef]
- Cogan, P.M.; Wakefield, M.E. Further developments in traps used to detect low-level infestations of beetle pests in bulk stored grain. In Stored Product Pest Control; Lawson, T.J., Ed.; BCPC: Croydon, UK, 1987; pp. 161–167. [Google Scholar]
- Roberts, G.T.; Chambers, J.; Wakefield, M.E. Response of Oryzaephilus surinamensis to carob extracts. Bull. OILB SROP 1993, 16, 314–319. [Google Scholar]
- Wakefield, M.E.; Morgan, C. The response of the saw-toothed grain beetle (Oryzaephilus surinamensis) to polyethylene tube lures containing carob volatiles in pitfall bioassay tests and bin trials. MAFF Cent. Sci. Lab. Rep. 1996, 71, 1–11. [Google Scholar]
- Ma, Q.H.; Tian, B.; Li, Y.L. Overexpression of a wheat jasmonate-regulated lectin increases pathogen resistance. Biochimie 2010, 92, 187–193. [Google Scholar] [CrossRef]
- Vandenborre, G.; Smagghe, G.; Van Damme, E.J.M. Plant lectins as defense proteins against phytophagous insects. Phytochemistry 2011, 72, 1538–1550. [Google Scholar] [CrossRef]
- Rahimi, V.; Hajizadeh, J.; Zibaee, A.; Jalali Sendi, J. Toxicity and physiological effects of an extracted lectin from Polygonum persicaria L. on Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Physiol. Mol. Plant Pathol. 2018, 101, 38–44. [Google Scholar] [CrossRef]
- Souza, C.S.; Procopio, T.F.; Belmonte, B.R.; Paiva, P.M.G.; Albuquerque, L.P.; Pontual, E.V.; Napoleao, T.H. Effects of Opuntia ficus-indica lectin on feeding, survival, and gut enzymes of maize weevil, Sitophilus zeamais. Appl. Biol. Chem. 2018, 61, 337–343. [Google Scholar] [CrossRef]
- Camaroti, J.R.S.L.; Almeida, W.A.; Belmonte, B.R.; Oliveira, A.P.S.; Lima, T.A.; Ferreira, M.R.A.; Paiva, P.M.G.; Soares, L.A.L.; Pontual, E.V.; Napoleao, T.H. Sitophilus zeamais adults have survival and nutrition affected by Schinus terebinthifolius leaf extract and its lectin (SteLL). Ind. Crops Prod. 2018, 116, 81–89. [Google Scholar] [CrossRef]
- Walski, T.; van Damme, E.J.M.; Smagghe, G. Penetration through the peritrophic matrix is a key to lectin toxicity against Tribolium castaneum. J. Insect Physiol. 2014, 70, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, L.P.; Procopio, T.F.; Guedes, C.C.S.; Pontual, E.V.; Paiva, P.M.G.; Napoleao, T.H. Antinutritional effects of the chitin-binding lectin from Microgramma vacciniifolia rhizome (MvRL) on Sitophilus zeamais. J. Stored Prod. Res. 2020, 88, 101652. [Google Scholar] [CrossRef]
- Sauvion, N.; Charles, H.; Febvay, G.; Rahbé, Y. Effects of jackbean lectin (ConA) on the feeding behaviour and kinetics of intoxication of the pea aphid, Acyrthosiphon pisum. Entomol. Exp. Appl. 2004, 110, 31–44. [Google Scholar] [CrossRef]
- Hamshou, M.; Smagghe, G.; Shahidi-Noghabi, S.; de Geyter, E.; Lannoo, N.; van Damme, E.J.M. Insecticidal properties of Sclerotinia sclerotiorum agglutinin and its interaction with insect tissues and cells. Insect Biochem. Mol. Biol. 2010, 40, 883–890. [Google Scholar] [CrossRef]
- Ramzi, S.; Sahragard, A.; Jalali Sendi, J.; Aalami, A. Effects of an extracted lectin from Citrullus colocynthis L. (Cucurbitaceae) on survival, digestion and energy reserves of Ectomyelois ceratoniae Zeller (Lepidoptera: Pyralidae). Front. Physiol. 2013, 12, 328. [Google Scholar] [CrossRef]
- Procópio, T.F.; Patriota, L.L.S.; de Moura, M.C.; da Silva, P.M.; de Oliveira, A.P.S.; Carvalho, L.V.N.; Lima, T.A.; Soares, T.; da Silva, T.D.; Coelho, L.C.B.B.; et al. CasuL: A new lectin isolated from Calliandra surinamensis leaf pinnulae with cytotoxicity to cancer cells, antimicrobial activity and antibiofilm effect. Int. J. Biol. Macromol. 2017, 98, 419–429. [Google Scholar] [CrossRef]
- Arteaga, I.T.; Gullien, J.L.C.; Olaya, E.M.; Gasca, T.G.; Zaragoza, M.V.A.; Garcia-Santoyo, V.; Castillo, J.A.T.; Aguirre, C.; Phinney, B.; Blanco-Labra, A. Characterization of Two Non-Fetuin-Binding Lectins from Tepary Bean (Phaseolus acutifolius) Seeds with Differential Cytotoxicity on Colon Cancer Cells. J. Glycobiol. 2016, 5, 1000117. [Google Scholar] [CrossRef]
- Mahdavi Moghdam, M.; Ghadamyari, M.; Talebi, K.; Memarizade, N. The effect of Artemisia annua L. (Asteraceae) essential oil on detoxify enzymes of two-spotted spider mite, Tetranychus urticae Kosh (Acari: Tetranychidae). J. Plant Prot. Res. 2012, 26, 233–242. [Google Scholar] [CrossRef]
- Valizadeh, B.; Sendi, J.J.; Zibaee, A.; Oftadeh, M. Effect of Neem based insecticide Achook® on mortality, biological and biochemical parameters of elm leaf beetle Xanthogaleruca luteola Mull (Col.: Chrysomelidae). J. Crop Prot. 2013, 2, 319–330. Available online: http://dorl.net/dor/20.1001.1.22519041.2013.2.3.1.3 (accessed on 15 December 2021).
- Adibmoradi, G.; Jalali Sendi, J.; Tirgari, S.; Imani, S.; Razavi-Nematolahi, A. Effect of 1,8-cineol on the biology and physiology of elm leaf beetle, Xanthogaleruca luteola (Col.: Chrysomelidae). J. Plant Prot. Res. 2018, 58, 420–430. [Google Scholar] [CrossRef]
- Gultekin, F.; Ozturk, M.; Akdogan, M. The effects of organophosphate insecticide chlorpyrifos-ethyl on lipid peroxidation and antioxidant enzymes (in vitro). Arch. Toxicol. 2000, 74, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Dubovskii, I.M.; Olifrenko, O.A.; Glupov, V.V. Level and activities of antioxidants in intestine of larvae Galleria mellonella L. (Lepidoptera, Pyralidae) at peroral infestation by bacteria Bacillus thuringiensis ssp. Galleriae. J. Evol. Biochem. Physiol. 2005, 41, 20–25. [Google Scholar] [CrossRef]
- Dubovskiy, I.M.; Martemyanov, V.V.; Vorontsova, Y.L.; Rantala, M.J.; Gryzanova, E.V.; Glupov, V.V. Effect of bacterial infection on antioxidant activity and lipid peroxidation in the midgut of Galleria mellonella L. larvae (Lepidoptera, Pyralidae). Comp. Biochem. Physiol. 2008, 148, 1–5. [Google Scholar] [CrossRef]
- Felton, G.W.; Summers, C.B. Antioxidant systems in insects. Arch. Insect Biochem. Physiol. 1995, 29, 187–197. [Google Scholar] [CrossRef]
- Barbehenn, R.V. Gut-based antioxidant enzymes in a polyphagous and a graminivorous grasshopper. J. Chem. Ecol. 2002, 28, 1329–1347. [Google Scholar] [CrossRef]
- Lyakhovich, V.V.; Vavilin, V.A.; Zenkov, N.K.; Menshchikova, E.B. Active defense under oxidative stress. The antioxidant responsive element. Biochemistry 2006, 71, 962–974. [Google Scholar] [CrossRef]
- Asada, K. Radical production and scavenging in the chloroplasts. Photosynth. Environ. 1996, 5, 123–150. [Google Scholar] [CrossRef]
- Nation, J.L. Insect Physiology and Biochemistry, 2nd ed.; CRC Press: London, UK, 2008. [Google Scholar]
- Wang, Y.; Oberley, L.W.; Murhammer, D.W. Evidence of oxidative stress following the viral infection of two Lepidopteran insect cell lines. Free Rad. Biol. Med. 2001, 31, 1448–1455. [Google Scholar] [CrossRef]
- Obeng-Ofori, D. The behaviour of 9 stored product beetles at pitfall trap arenas and their capture in millet. Entomol. Exp. Appl. 1993, 66, 161–169. [Google Scholar] [CrossRef]
- Wakefield, M.E.; Bryning, G.P.; Chambers, J. Progress towards a lure to attract three stored product weevils, Sitophilus zeamais Motschulsky, S. oryzae (L.) and S. granarius (L.) (Coleoptera: Curculionidae). J. Stored Prod. Res. 2005, 41, 145–161. [Google Scholar] [CrossRef]
- Obeng-Ofori, D. Behavioural responses of three stored product Coleoptera species to extract of carob (Locust bean), Ceratonia siliqua. Entomol. Exp. Appl. 1993, 68, 9–13. [Google Scholar] [CrossRef]
- Kim, S.I.; Roh, J.Y.; Kim, D.H.; Lee, H.S.; Ahn, Y.J. Insecticidal activities of aromatic plant extracts and essential oils against Sitophilus oryzae and Callosobruchus chinensis. J. Stored Prod. Res. 2003, 39, 293–303. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Bernfeld, P. Amylases, α and β. Meth. Enzymol. 1955, 1, 149–158. [Google Scholar] [CrossRef]
- Silva, C.P.; Terra, W.R.M. An α-glucosidase from perimicrovillar membranes of Dysdercus peruvianus (Hemiptera: Pyrrhocoridae) midgut cells: Purification and properties. Insect Biochem. Mol. Biol. 1995, 25, 487–494. [Google Scholar] [CrossRef]
- Tsujita, T.; Ninomiya, H.; Okuda, H. p-Nitrophenyl butyrate 1 hydrolyzing activity of hormone-sensitive lipase from bovine adipose tissue. J. Lipid Res. 1989, 30, 997–1004. [Google Scholar] [CrossRef]
- Oppert, B.; Kramer, K.J.; Mc Gaughey, W.H. Rapid microplate assay of proteinase mixtures. Biotechnology 1997, 23, 70–72. [Google Scholar] [CrossRef]
- Han, Z.; Graham, D.; Moores, I.D.; Alan, L. Devonshire association between biochemical markers and insecticide resistance in the cotton aphid, Aphis gossypii Glover. Pestic. Biochem. Physiol. 1998, 62, 164–171. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione-S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- McCord, J.M.; Fridovich, I. Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [CrossRef]
- Addy, S.K.; Goodman, R.N. Polyphenol oxidase and peroxidase in apple leaves inoculated with a virulent or an avirulent strain for Ervinia amylovora. Indian Phytopathol. 1972, 25, 575–579. [Google Scholar]
- Balinsky, D.; Bernstein, R.E. The purification and properties of glucose-6-phosphate dehydrogenase from human erythrocytes. Biochim. Biophys. Act. 1963, 67, 313–315. [Google Scholar] [CrossRef]
- Khramtsov, V.V.; Yelinova, V.I.; Glazachev, Y.U.I.; Reznikov, V.A.; Zimmer, G.; Weiner, L.; Berezina, T.; Martin, V.; Volodarsky, L. Quantitative determination and reversible modification of thiols using imidazolidine biradical disulfide labl. J. Biochem. Biophys. Meth. 1997, 35, 115–128. [Google Scholar] [CrossRef]
- Bar-Or, D.; Rael, L.T.; Lau, E.P.; Rao, N.K.; Thomas, G.W.; Winkler, J.V.; Yukl, R.L.; Kingston, R.G.; Curtis, C.G. An analog of the human albumin N-terminus (Asp-Ala-His-Lys) prevents formation of copperinduced reactive oxygen species. Biochem. Biophys. Res. Commun. 2001, 284, 856–862. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Bashir, T.; Birkinshaw, L.A.; Hall, D.R.; Hodges, R.J. Host odors enhance the responses of adult Rhyzopertha dominica to male-produced aggregation pheromone. Entomol. Exp. Appl. 2001, 101, 273–280. [Google Scholar] [CrossRef]
- SAS Institute. SAS/STAT User’s Guide for Personal Computers; SAS Institute: Cary, NC, USA, 2010. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N | LC30 with 95% Confidence Limit (%) | LC50 with 95% Confidence Limit (%) | Slope ± SE | χ2 (df = 3) | R2 |
|---|---|---|---|---|---|
| 120 | 1.82 (1.19–2.52) | 3.68 (2.65–5.78) | 1.717 ± 0.149 | 5.28 | 0.95 |
| Enzymes | Treatments | |
|---|---|---|
| Control | PPA | |
| α-Amylase (U/mg protein) | 4.28 ± 0.13 * | 1.58 ± 0.10 |
| α-Glucosidase (U/mg protein) | 0.76 ± 0.02 * | 0.33 ± 0.02 |
| β-Glucosidase (U/mg protein) | 0.71 ± 0.01 | 0.72 ± 0.01 |
| TAG-Lipase (U/mg protein) | 1.27 ± 0.06 * | 0.26 ± 0.02 |
| Trypsin (U/mg protein) | 0.45 ± 0.01 * | 0.19 ± 0.03 |
| Chymotrypsin (U/mg protein) | 0.46 ± 0.01 * | 0.17 ± 0.01 |
| Elastase (U/mg protein) | 5.03 ± 0.06 * | 2.63 ± 0.15 |
| Carboxypeptidase (U/mg protein) | 4.88 ± 0.19 * | 1.49 ± 0.12 |
| Aminopeptidase (U/mg protein) | 5.11 ± 0.05 * | 2.26 ± 0.27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khoobdel, M.; Rahimi, V.; Ebadollahi, A.; Krutmuang, P. Evaluation of the Potential of a Lectin Extracted from Polygonumpersicaria L. as a Biorational Agent against Sitophilusoryzae L. Molecules 2022, 27, 793. https://doi.org/10.3390/molecules27030793
Khoobdel M, Rahimi V, Ebadollahi A, Krutmuang P. Evaluation of the Potential of a Lectin Extracted from Polygonumpersicaria L. as a Biorational Agent against Sitophilusoryzae L. Molecules. 2022; 27(3):793. https://doi.org/10.3390/molecules27030793
Chicago/Turabian StyleKhoobdel, Mehdi, Vahid Rahimi, Asgar Ebadollahi, and Patcharin Krutmuang. 2022. "Evaluation of the Potential of a Lectin Extracted from Polygonumpersicaria L. as a Biorational Agent against Sitophilusoryzae L." Molecules 27, no. 3: 793. https://doi.org/10.3390/molecules27030793
APA StyleKhoobdel, M., Rahimi, V., Ebadollahi, A., & Krutmuang, P. (2022). Evaluation of the Potential of a Lectin Extracted from Polygonumpersicaria L. as a Biorational Agent against Sitophilusoryzae L. Molecules, 27(3), 793. https://doi.org/10.3390/molecules27030793