
Pro-Inflammatory Interactions of Dolutegravir with Human Neutrophils in an In Vitro Study

 , , ,
, , ,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. ROS Production
2.2. Elastase Release
2.3. Cytosolic Ca2+ Fluxes
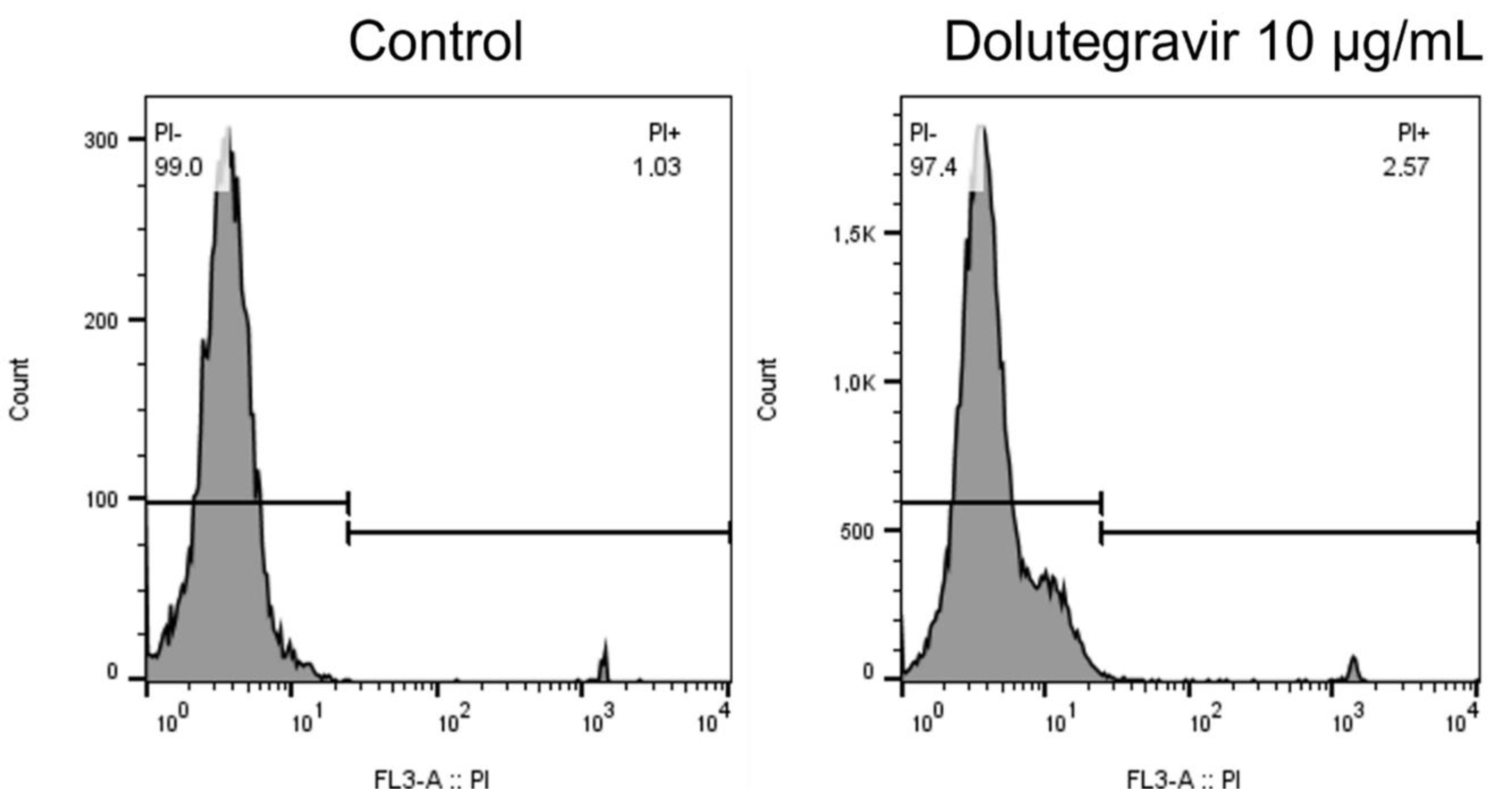
2.4. Cell Viability
2.5. Hemolytic Activity of Dolutegravir
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
4.2. Chemicals and Reagents
4.3. Preparation of Neutrophils
4.4. Measurement of Reactive Oxygen Species
4.5. Elastase Release
4.6. Cytosolic Ca2+ Fluxes
4.7. Cell Viability
4.8. Hemolytic Assay
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Taha, H.; Das, A.; Das, S. Clinical effectiveness of dolutegravir in the treatment of HIV. Infect. Drug Resist. 2015, 8, 339–352. [Google Scholar] [CrossRef] [PubMed]
- WHO. WHO Recommends Dolutegravir as Preferred HIV Treatment Option in All Populations. 2019. Available online: who.int/news/item/22-07-2019-who-recommends-dolutegravir-as-preferred-hiv-treatment-option-in-all-populations (accessed on 22 April 2021).
- Hauser, A.; Kusejko, K.; Johnson, L.F.; Günthard, H.F.; Riou, J.; Wandeler, G.; Egger, M.; Kouyos, R.D. Impact of scaling up dolutegravir on antiretroviral resistance in South Africa: A modeling study. PLoS Med. 2020, 17, e1003397. [Google Scholar] [CrossRef] [PubMed]
- Menard, A.; Meddeb, L.; Tissot-Dupont, H.; Ravaux, I.; Dhiver, C.; Mokhtari, S.; Tomei, C.; Brouqui, P.; Colson, P.; Stein, A. Dolutegravir and weight gain: An unexpected bothering side effect? AIDS 2017, 31, 1499–1500. [Google Scholar] [CrossRef] [PubMed]
- Bourgi, K.; Rebeiro, P.F.; Turner, M.; Castilho, J.L.; Hulgan, T.; Raffanti, S.P.; Koethe, J.R.; Sterling, T.R. Greater weight gain in treatment-naive persons starting dolutegravir-based antiretroviral therapy. Clin. Infect. Dis. 2020, 70, 1267–1274. [Google Scholar] [CrossRef]
- Venter, W.D.F.; Moorhouse, M.; Sokhela, S.; Fairlie, L.; Mashabane, N.; Masenya, M.; Serenata, C.; Akpomiemie, G.; Qavi, A.; Chandiwana, N.; et al. Dolutegravir plus two different prodrugs of tenofovir to treat HIV. N. Engl. J. Med. 2019, 381, 803–815. [Google Scholar] [CrossRef]
- Burns, J.E.; Stirrup, O.T.; Dunn, D.; Runcie-Unger, I.; Milinkovic, A.; Candfield, S.; Lukha, H.; Severn, A.; Waters, L.; Edwards, S.; et al. No overall change in the rate of weight gain after switching to an integrase-inhibitor in virologically suppressed adults with HIV. AIDS 2020, 34, 109–114. [Google Scholar] [CrossRef]
- Esber, A.L.; Chang, D.; Iroezindu, M.; Bahemana, E.; Kibuuka, H.; Owuoth, J.; Singoei, V.; Maswai, J.; Dear, N.F.; Crowell, T.A.; et al. Weight gain during the dolutegravir transition in the African Cohort Study. J. Int. AIDS Soc. 2022, 25, e25899. [Google Scholar] [CrossRef]
- Godfrey, C.; Bremer, A.; Alba, D.; Apovian, C.; Koethe, J.R.; Koliwad, S.; Lewis, D.; Lo, J.; McComsey, G.A.; Eckard, A.; et al. Obesity and fat metabolism in human immunodeficiency virus-infected individuals: Immunopathogenic mechanisms and clinical implications. J. Infect. Dis. 2019, 220, 420–431. [Google Scholar] [CrossRef]
- Hepler, C.; Gupta, R.K. The expanding problem of adipose depot remodeling and postnatal adipocyte progenitor recruitment. Mol. Cell Endocrinol. 2017, 445, 95–108. [Google Scholar] [CrossRef]
- Lehman, H.K.; Segal, B.H. The role of neutrophils in host defense and disease. J. Allergy Clin. Immunol. 2020, 145, 1535–1544. [Google Scholar] [CrossRef]
- Teng, T.S.; Ji, A.L.; Ji, X.Y.; Li, Y.Z. Neutrophils and immunity: From bactericidal action to being conquered. J. Immunol. Res. 2017, 2017, 9671604. [Google Scholar] [CrossRef] [PubMed]
- Peiró, T.; Patel, D.F.; Akthar, S.; Gregory, L.G.; Pyle, C.J.; Harker, J.A.; Birrell, M.A.; Lloyd, C.M.; Snelgrove, R.J. Neutrophils drive alveolar macrophage IL-1β release during respiratory viral infection. Thorax 2018, 73, 546–556. [Google Scholar] [CrossRef]
- Nijhuis, J.; Rensen, S.S.; Slaats, Y.; van Dielen, F.M.; Buurman, W.A.; Greve, J.W. Neutrophil activation in morbid obesity, chronic activation of acute inflammation. Obesity 2009, 17, 2014–2018. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Su, S.; Wang, X.; Barnes, V.; De Miguel, C.; Ownby, D.; Pollock, J.; Snieder, H.; Chen, W.; Wang, X. Obesity is associated with more activated neutrophils in African American male youth. Int. J. Obes. 2015, 39, 26–32. [Google Scholar] [CrossRef]
- Brotfain, E.; Hadad, N.; Shapira, Y.; Avinoah, E.; Zlotnik, A.; Raichel, L.; Levy, R. Neutrophil functions in morbidly obese subjects. Clin. Exp. Immunol. 2015, 181, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Madzime, M.; Rossouw, T.M.; Theron, A.J.; Anderson, R.; Steel, H.C. Interactions of HIV and antiretroviral therapy with neutrophils and platelets. Front. Immunol. 2021, 12, 634386. [Google Scholar] [CrossRef]
- Al Mamun Bhuyan, A.; Signoretto, E.; Bissinger, R.; Lang, F. Enhanced eryptosis following exposure to dolutegravir. Cell Physiol. Biochem. 2016, 39, 639–650. [Google Scholar] [CrossRef]
- Wang, W.J.; Mao, L.F.; Lai, H.L.; Wang, Y.W.; Jiang, Z.B.; Li, W.; Huang, J.M.; Xie, Y.J.; Xu, C.; Liu, P.; et al. Dolutegravir derivative inhibits proliferation and induces apoptosis of non-small cell lung cancer cells via calcium signaling pathway. Pharmacol. Res. 2020, 161, 105129. [Google Scholar] [CrossRef]
- Borregaard, N.; Lollike, K.; Kjeldsen, L.; Sengeløv, H.; Bastholm, L.; Nielsen, M.H.; Bainton, D.F. Human neutrophil granules and secretory vesicles. Eur. J. Haematol. 1993, 51, 187–198. [Google Scholar] [CrossRef]
- Song, I.; Borland, J.; Arya, N.; Wynne, B.; Piscitelli, S. Pharmacokinetics of dolutegravir when administered with mineral supplements in healthy adult subjects. J. Clin. Pharmacol. 2015, 55, 490–496. [Google Scholar] [CrossRef]
- Madzime, M.; Theron, A.J.; Anderson, R.; Tintinger, G.R.; Steel, H.C.; Meyer, P.W.A.; Nel, J.G.; Feldman, C.; Rossouw, T.M. Dolutegravir potentiates platelet activation by a calcium-dependent, ionophore-like mechanism. J. Immunotoxicol. 2022, 19, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Grießinger, J.A.; Hauptstein, S.; Laffleur, F.; Netsomboon, K.; Bernkop-Schnürch, A. Evaluation of the impact of multivalent metal ions on the permeation behavior of Dolutegravir sodium. Drug Dev. Ind. Pharm. 2016, 42, 1118–1126. [Google Scholar] [CrossRef] [PubMed]
- Thierry, E.; Lebourgeois, S.; Simon, F.; Delelis, O.; Deprez, E. Probing resistance mutations in retroviral integrases by direct measurement of dolutegravir fluorescence. Sci. Rep. 2017, 7, 14067. [Google Scholar] [CrossRef] [PubMed]
- Scully, T.; Ettela, A.; LeRoith, D.; Gallagher, E.J. Obesity, type 2 diabetes, and cancer risk. Front. Oncol. 2021, 10, 615375. [Google Scholar] [CrossRef]
- Herrin, M.; Tate, J.P.; Akgün, K.M.; Butt, A.A.; Crothers, K.; Freiberg, M.S.; Gibert, C.L.; Leaf, D.A.; Rimland, D.; Rodriguez-Barradas, M.C.; et al. Weight gain and incident diabetes among HIV-infected veterans initiating antiretroviral therapy compared with uninfected individuals. J. Acquir. Immune Defic. Syndr. 2016, 73, 228–236. [Google Scholar] [CrossRef]
- Achhra, A.C.; Sabin, C.; Ryom, L.; Hatleberg, C.; Antonella d’Aminio, M.; de Wit, S.; Phillips, A.; Pradier, C.; Weber, R.; Reiss, P.; et al. Body mass index and the risk of serious non-AIDS events and all-cause mortality in treated HIV-Positive individuals: D: A: D Cohort Analysis. J. Acquir. Immune Defic. Syndr. 2018, 78, 579–588. [Google Scholar] [CrossRef]
- Kumar, S.; Samaras, K. The impact of weight gain during HIV treatment on risk of pre-diabetes, diabetes mellitus, cardiovascular disease, and mortality. Front. Endocrinol. 2018, 9, 705. [Google Scholar] [CrossRef]
- Domingo, P.; Quesada-Lopez, T.; Villarroya, J.; Cairo, M.; Del Mar Gutierrez, M.; Mateo, M.G.; Mur, I.; Corbacho, N.; Domingo, J.C.; Villarroya, F.; et al. Differential effects of dolutegravir, bictegravir and raltegravir in adipokines and inflammation markers on human adipocytes. Life Sci. 2022, 308, 120948. [Google Scholar] [CrossRef]
- Minkenberg, I.; Ferber, E. Lucigenin-dependent chemiluminescence as a new assay for NAD(P)H-oxidase activity in particulate fractions of human polymorphonuclear leukocytes. J. Immunol. Methods 1984, 71, 61–67. [Google Scholar] [CrossRef]
- Beatty, K.; Robertie, P.; Senior, R.M.; Travis, J. Determination of oxidized alpha-1-proteinase inhibitor in serum. J. Lab. Clin. Med. 1982, 100, 186–192. [Google Scholar] [CrossRef]
- Grynkiewicz, G.; Poenie, M.; Tsien, R.Y. A new generation of Ca2+ indicators with greatly improved fluorescence properties. J. Biol. Chem. 1985, 260, 3440–3450. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Theron, A.J.; Anderson, R.; Madzime, M.; Rossouw, T.M.; Steel, H.C.; Meyer, P.W.A.; Cholo, M.C.; Kwofie, L.L.I.; Feldman, C.; Tintinger, G.R. Pro-Inflammatory Interactions of Dolutegravir with Human Neutrophils in an In Vitro Study. Molecules 2022, 27, 9057. https://doi.org/10.3390/molecules27249057
Theron AJ, Anderson R, Madzime M, Rossouw TM, Steel HC, Meyer PWA, Cholo MC, Kwofie LLI, Feldman C, Tintinger GR. Pro-Inflammatory Interactions of Dolutegravir with Human Neutrophils in an In Vitro Study. Molecules. 2022; 27(24):9057. https://doi.org/10.3390/molecules27249057
Chicago/Turabian StyleTheron, Annette J., Ronald Anderson, Morris Madzime, Theresa M. Rossouw, Helen C. Steel, Pieter W. A. Meyer, Moloko C. Cholo, Luyanda L. I. Kwofie, Charles Feldman, and Gregory R. Tintinger. 2022. "Pro-Inflammatory Interactions of Dolutegravir with Human Neutrophils in an In Vitro Study" Molecules 27, no. 24: 9057. https://doi.org/10.3390/molecules27249057
APA StyleTheron, A. J., Anderson, R., Madzime, M., Rossouw, T. M., Steel, H. C., Meyer, P. W. A., Cholo, M. C., Kwofie, L. L. I., Feldman, C., & Tintinger, G. R. (2022). Pro-Inflammatory Interactions of Dolutegravir with Human Neutrophils in an In Vitro Study. Molecules, 27(24), 9057. https://doi.org/10.3390/molecules27249057