
Relationships between X-ray Diffraction Peaks, Molecular Components, and Heat Properties of C-Type Starches from Different Sweet Potato Varieties


 and
and
Abstract
:1. Introduction
2. Results and Discussion
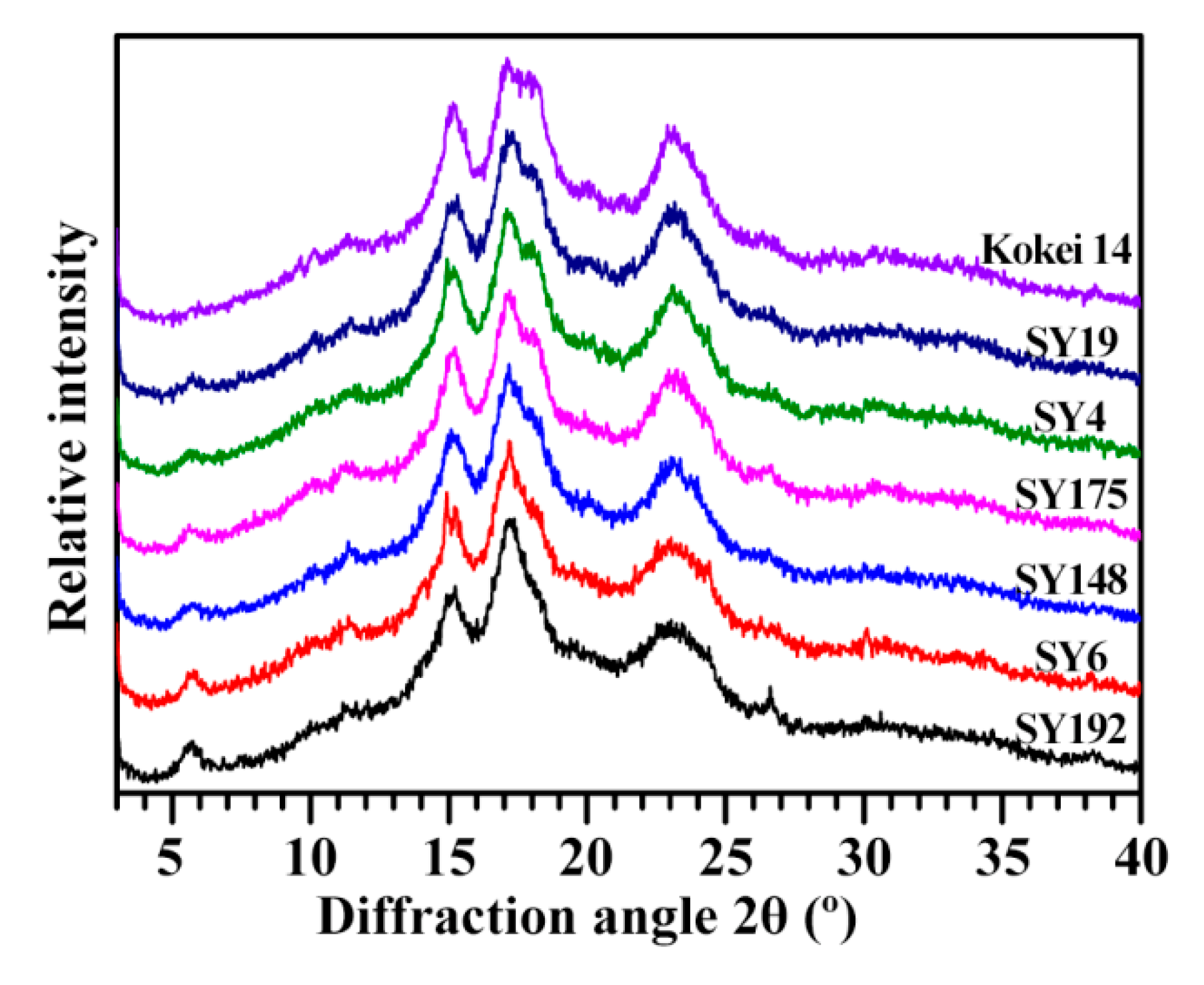
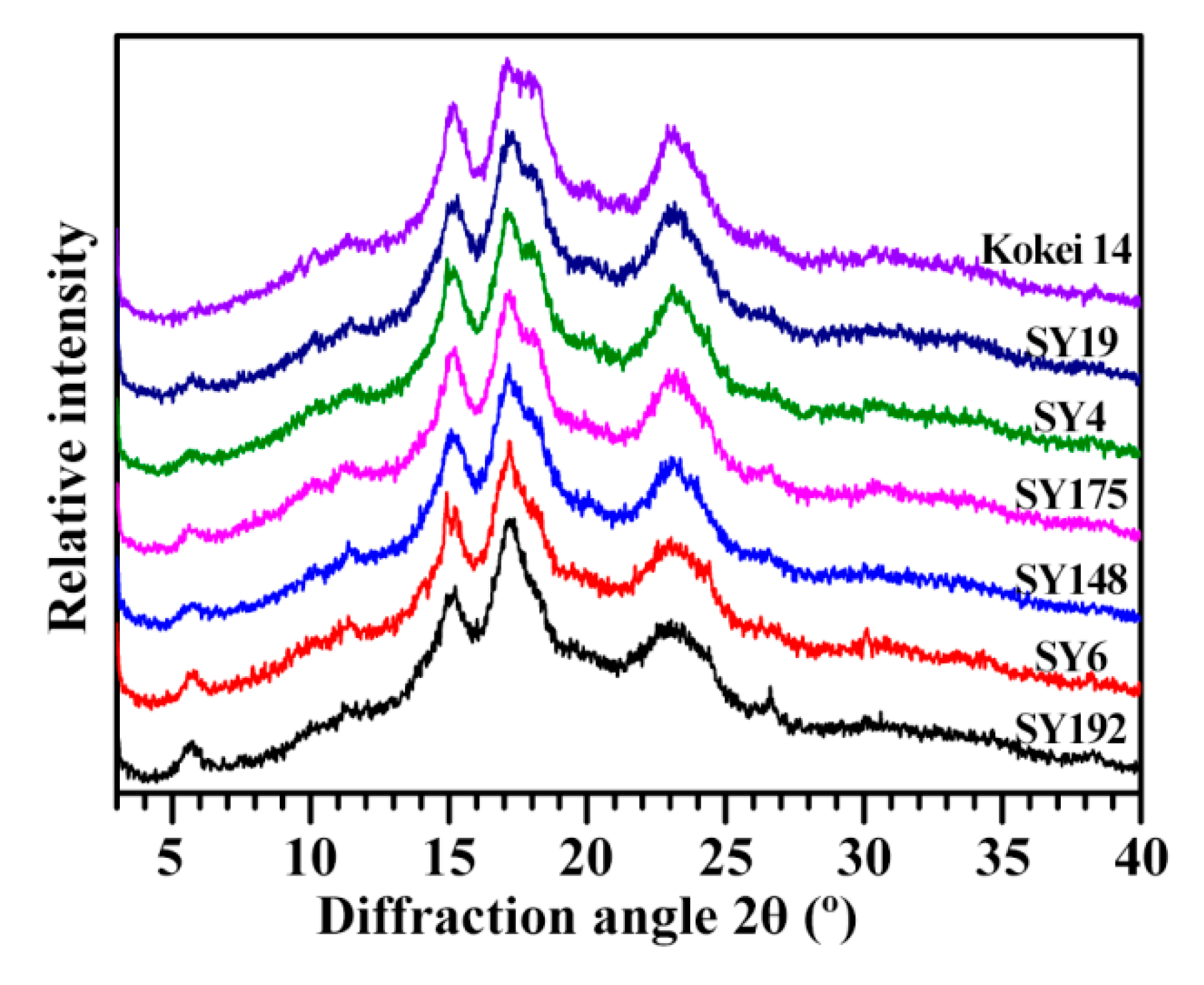
2.1. X-ray Diffraction Peaks of C-Type Starches
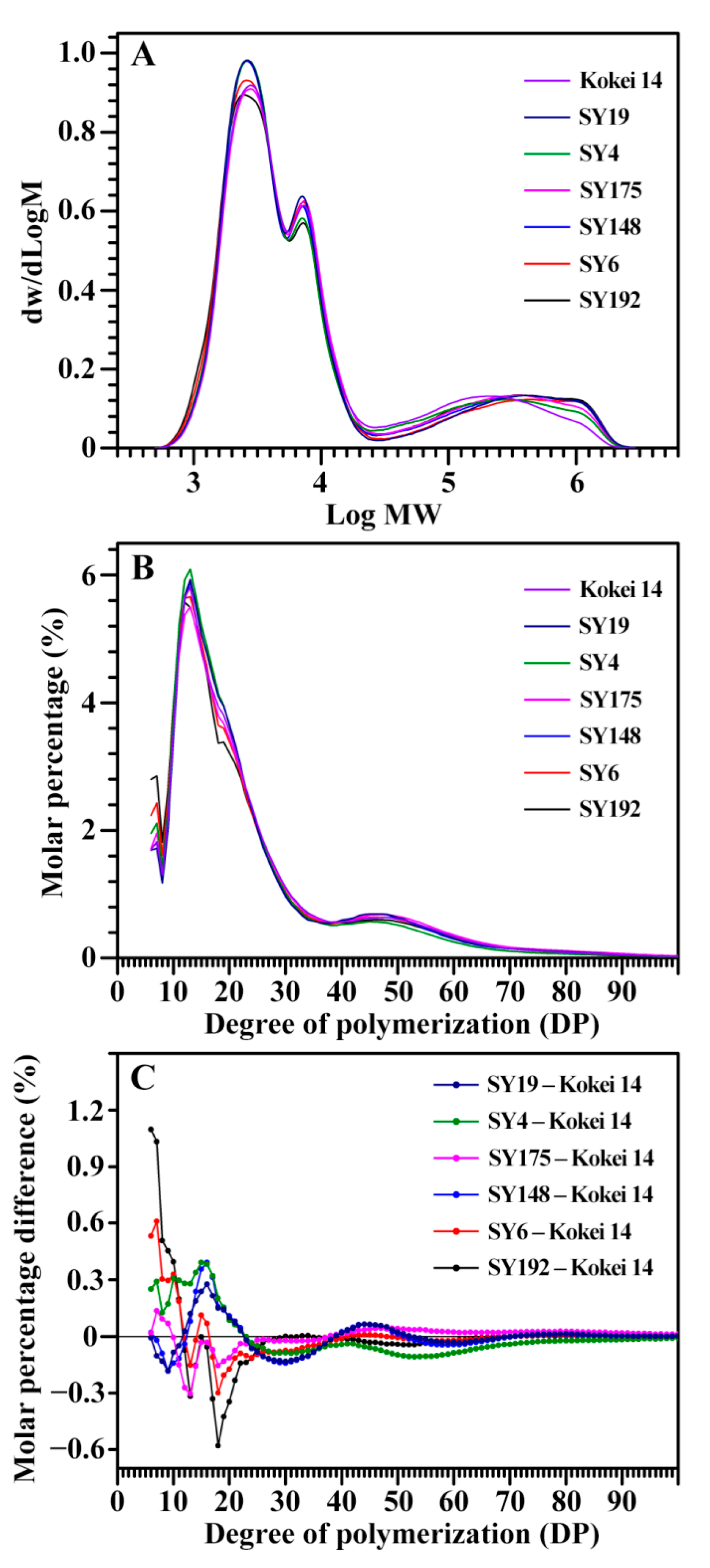
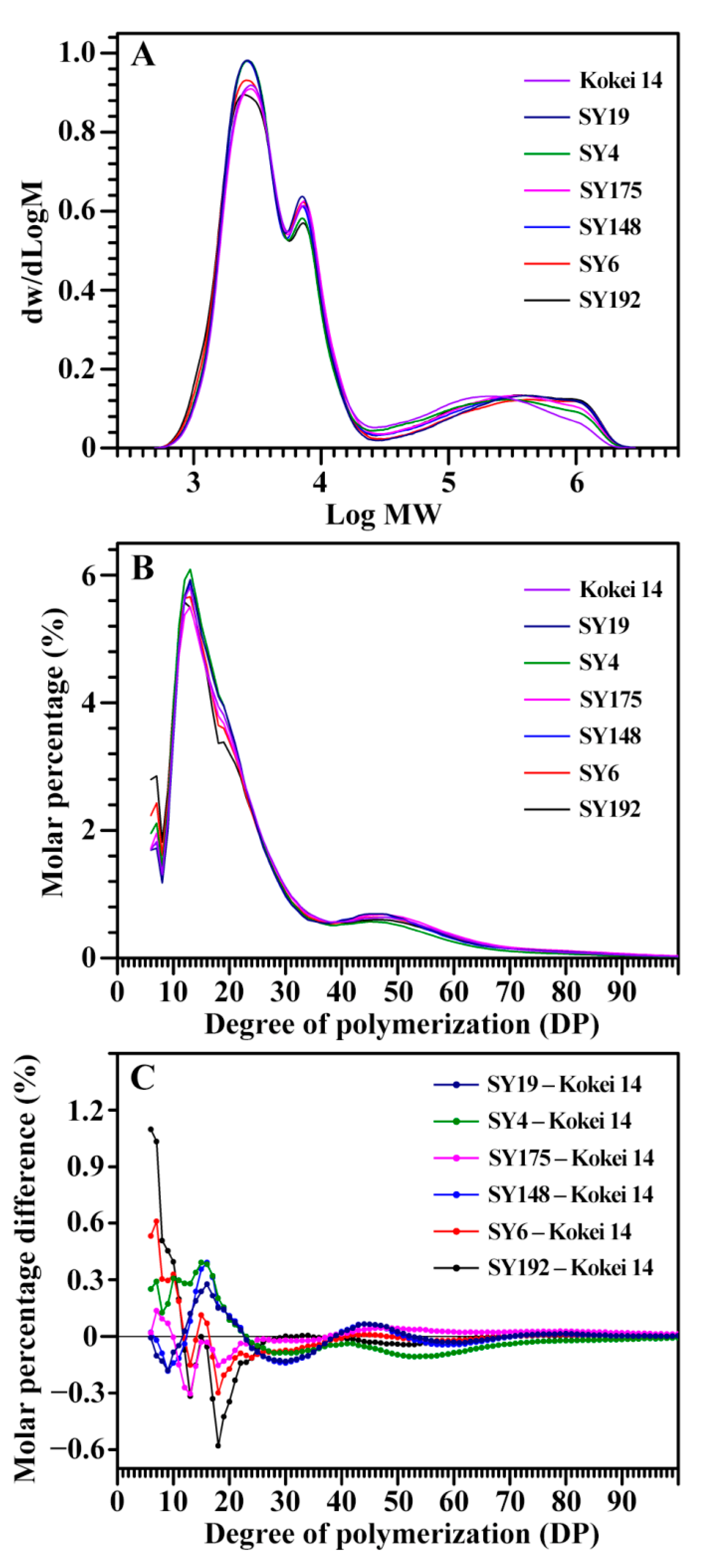
2.2. Molecular Components of C-Type Starches
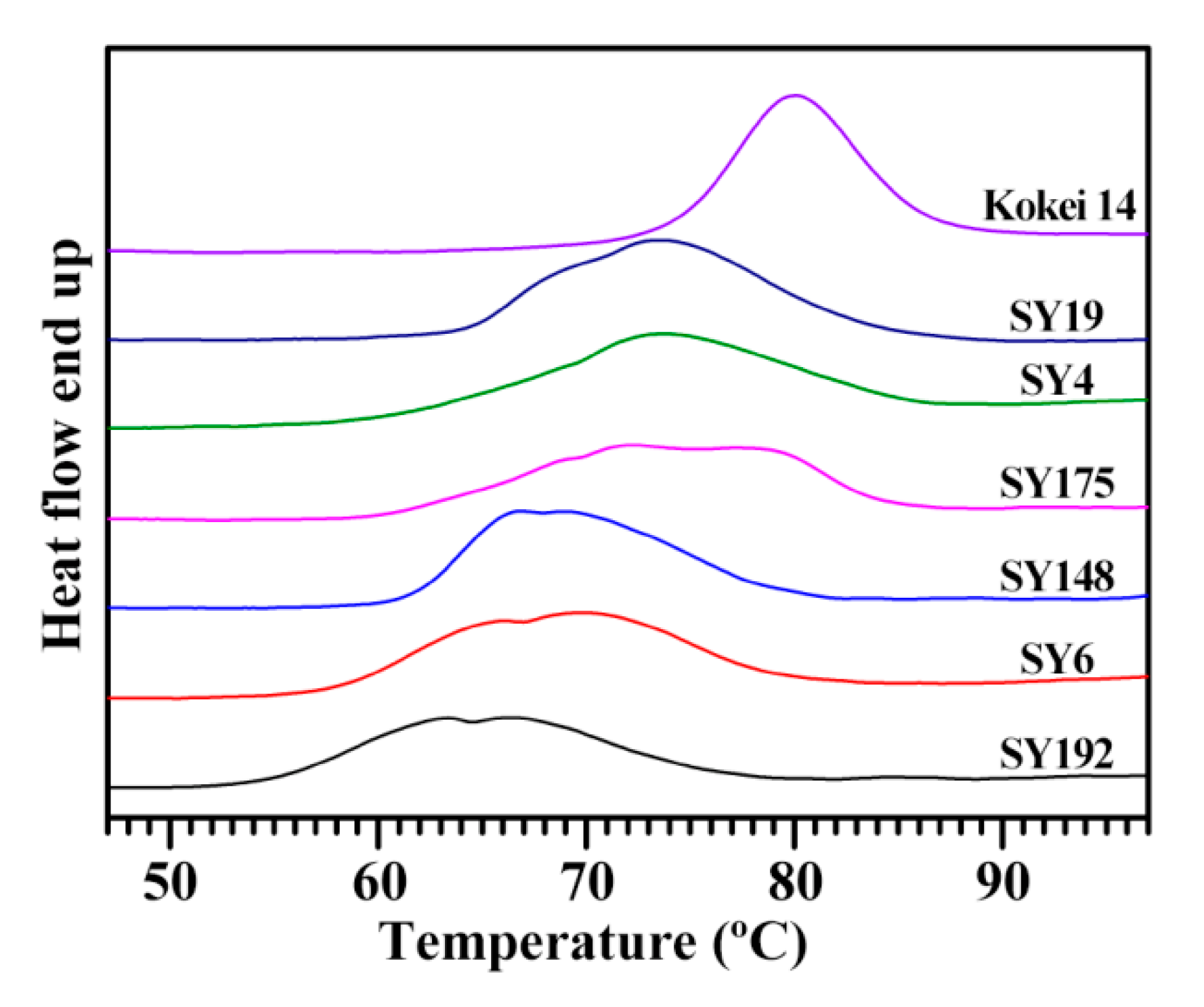
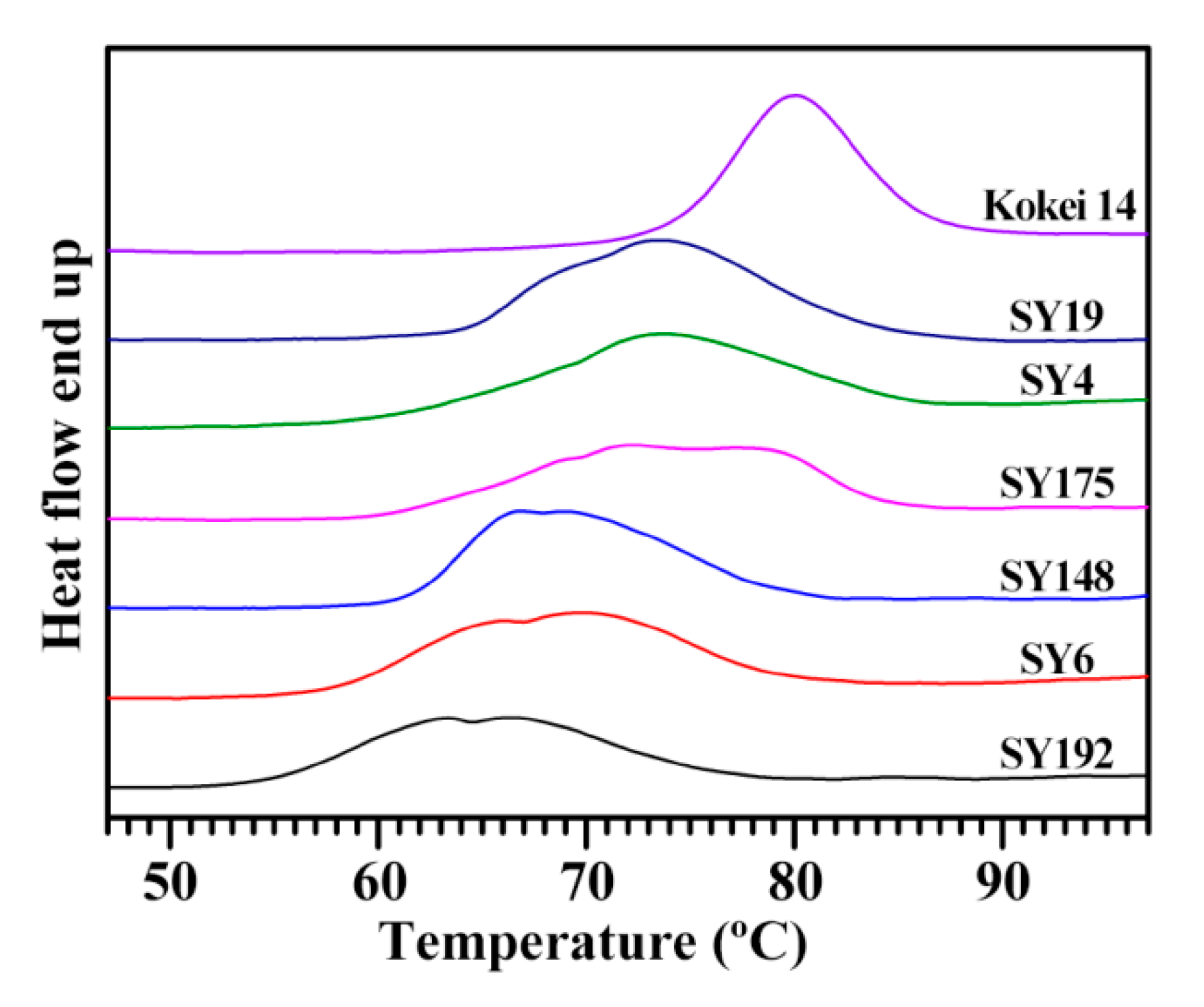
2.3. Heat Properties of C-Type Starches
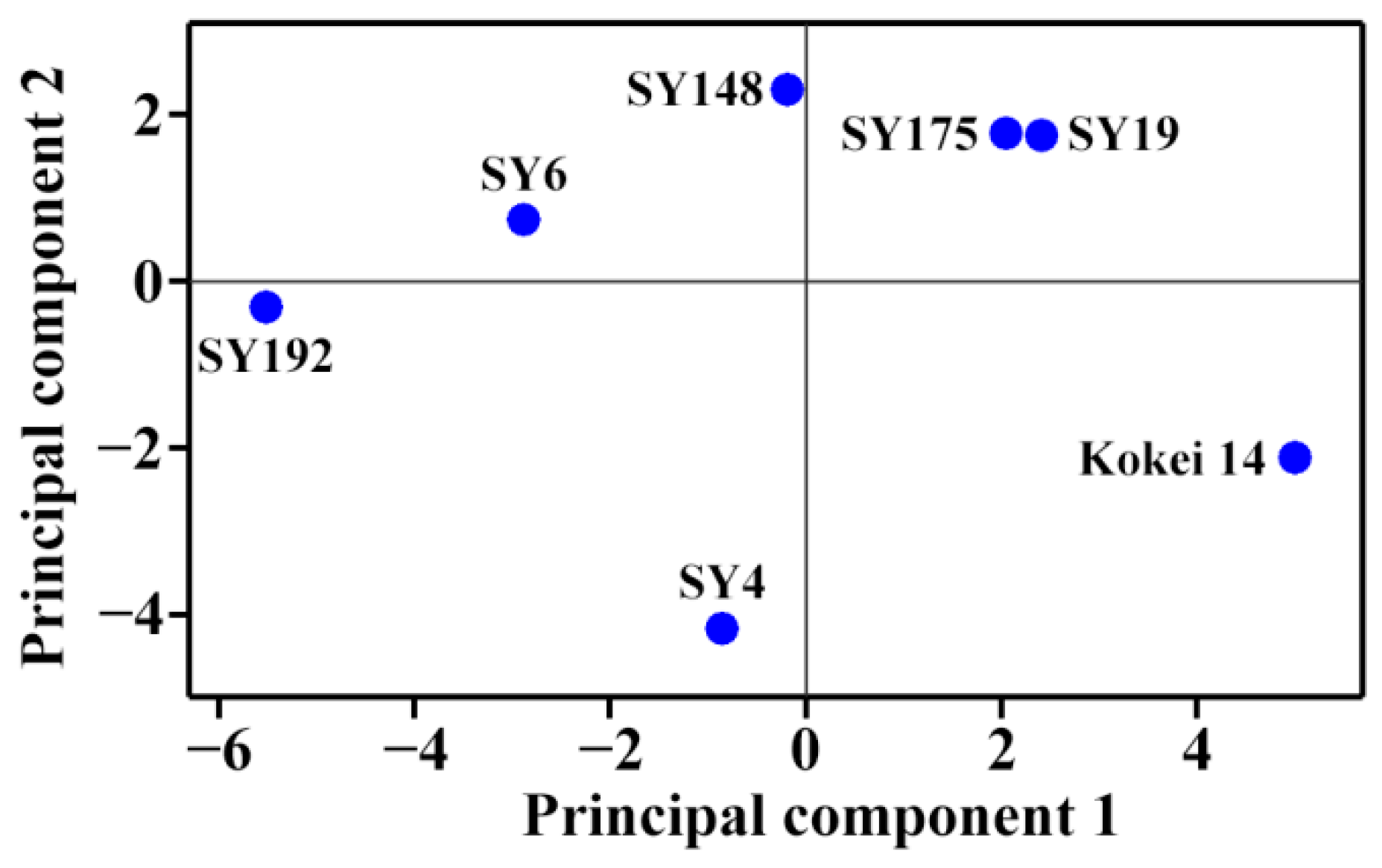
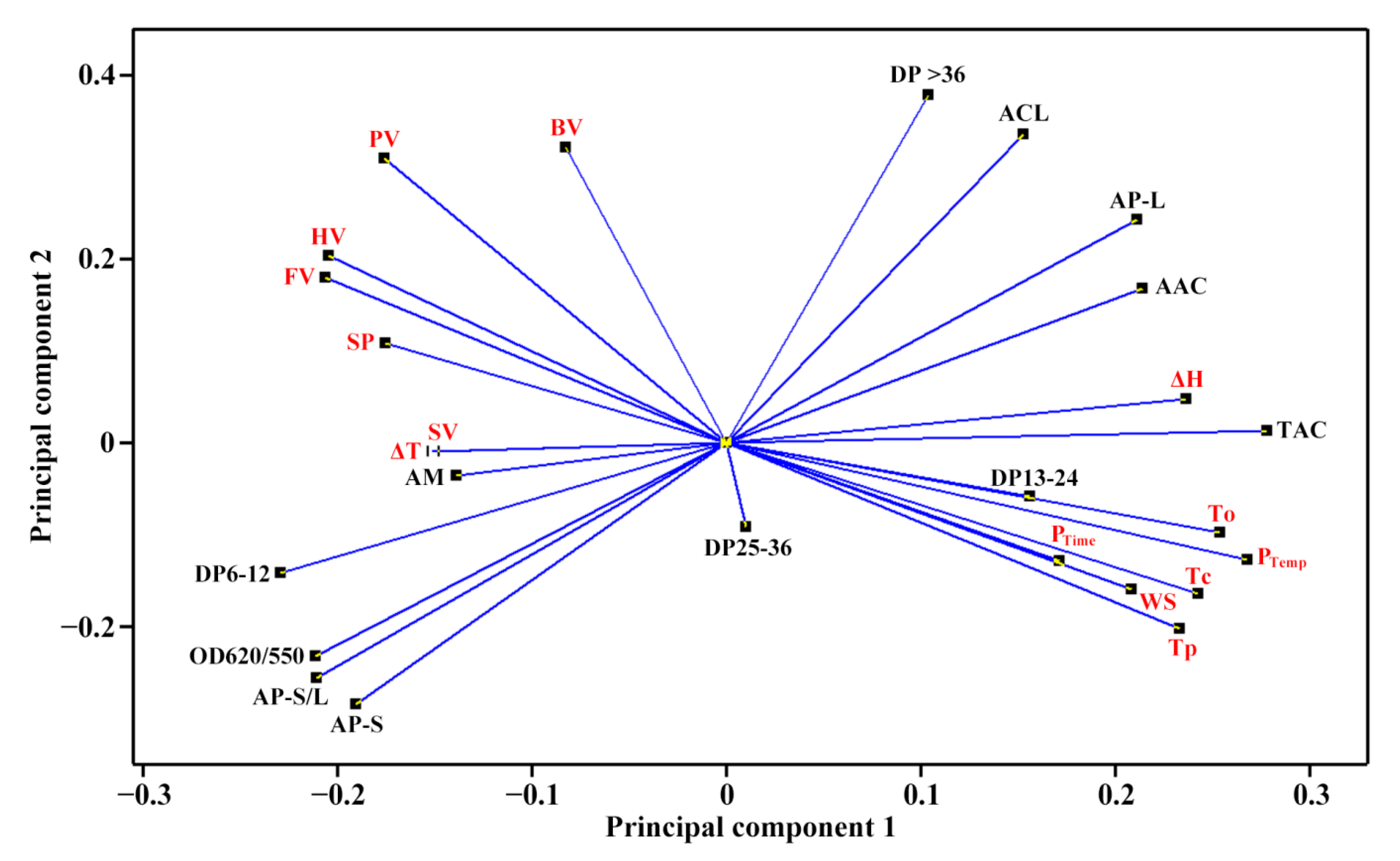
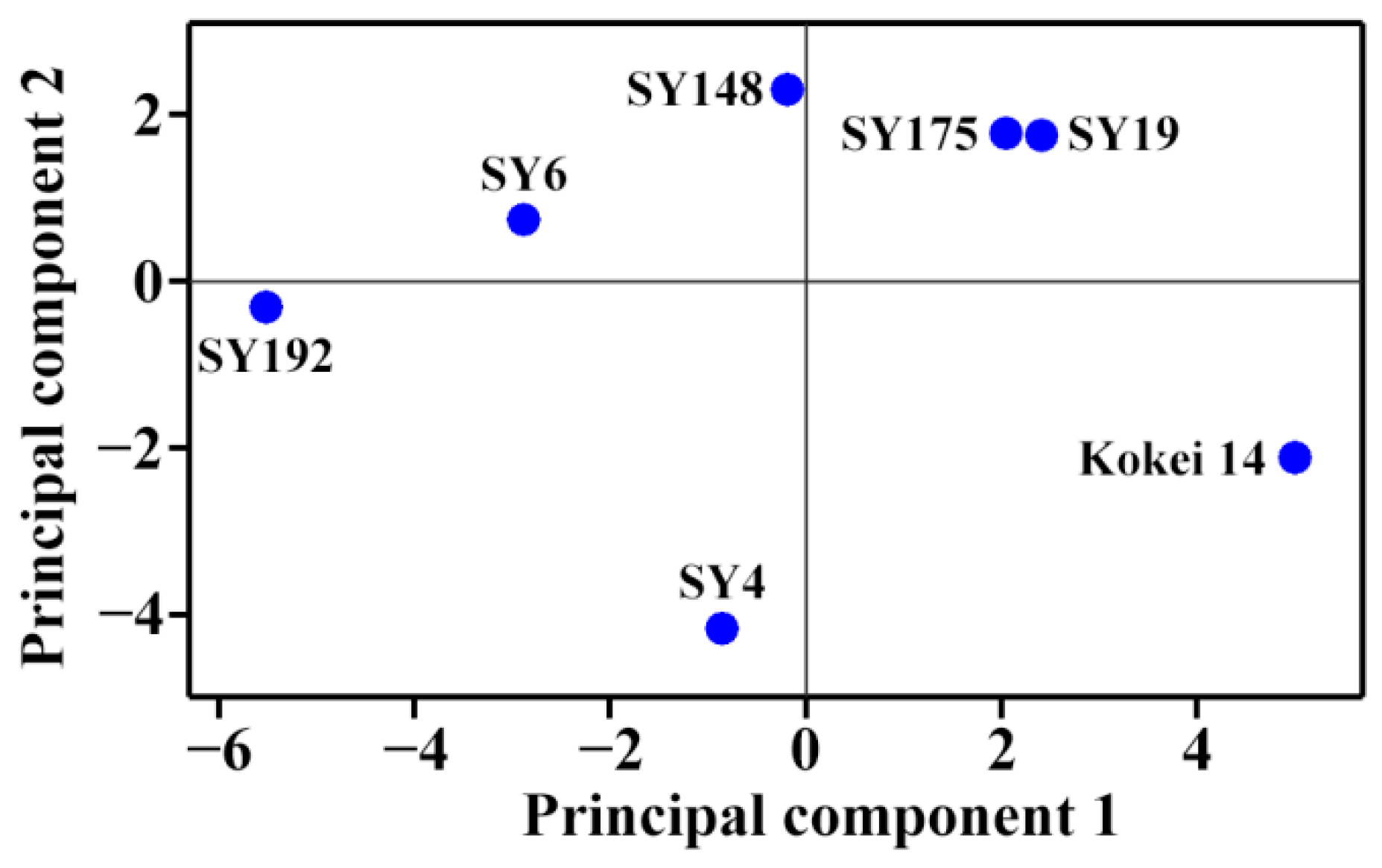
2.4. Principal Component Analysis of Molecular Components and Heat Properties of C-Type Starches
3. Materials and Methods
3.1. Plant Materials
3.2. Isolation of Starch
3.3. Measurement of Starch–Iodine Absorption and Amylose Content
3.4. Molecular Weight Distribution Analysis of Starch
3.5. Chain Length Distribution Analysis of Amylopectin
3.6. Crystalline Structure Analysis of Starch
3.7. Thermal Property Analysis of Starch
3.8. Measurement of Swelling Power and Water Solubility of Starch
3.9. Pasting Property Analysis of Starch
3.10. Principal Component Analysis (PCA)
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Guo, Z.; Jia, X.; Zhao, B.; Zeng, S.; Xiao, J.; Zheng, B. C-type starches and their derivatives: Structure and function. Ann. N. Y. Acad. Sci. 2017, 1398, 47–61. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Wei, C. Progress in C-type starches from different plant sources. Food Hydrocoll. 2017, 73, 162–175. [Google Scholar] [CrossRef]
- Ouyang, Q.; Wang, X.; Xiao, Y.; Luo, F.; Lin, Q.; Ding, Y. Structural changes of A-, B- and C-type starches of corn, potato and pea as influenced by sonication temperature and their relationships with digestibility. Food Chem. 2021, 358, 129858. [Google Scholar] [CrossRef] [PubMed]
- Barreto, F.F.V.; Bello-Perez, L.A. Chemical, structural, technological properties and applications of Andean tuber starches: A review. Food Rev. Int. 2021. [Google Scholar] [CrossRef]
- Dereje, B. Composition, morphology and physicochemical properties of starches derived from indigenous Ethiopian tuber crops: A review. Int. J. Biol. Macromol. 2021, 187, 911–921. [Google Scholar] [CrossRef]
- Buléon, A.; Gérard, C.; Riekel, C.; Vuong, R.; Chanzy, H. Details of the crystalline ultrastructure of C-starch granules revealed by synchrotron microfocus mapping. Macromolecules 1998, 31, 6605–6610. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.; Qin, F.; Zhou, W.; Yu, H.; Xu, B.; Chen, C.; Zhu, L.; Wang, Y.; Gu, M.; Liu, Q. Granule structure and distribution of allomorphs in C-type high-amylose rice starch granule modified by antisense RNA inhibition of starch branching enzyme. J. Agric. Food Chem. 2010, 58, 11946–11954. [Google Scholar] [CrossRef]
- Cai, C.; Cai, J.; Man, J.; Yang, Y.; Wang, Z.; Wei, C. Allomorph distribution and granule structure of lotus rhizome C-type starch during gelatinization. Food Chem. 2014, 142, 408–415. [Google Scholar] [CrossRef]
- Fan, X.; Zhao, L.; Zhang, L.; Xu, B.; Wei, C. A new allomorph distribution of C-type starch from root tuber of Apios fortunei. Food Hydrocoll. 2017, 66, 334–342. [Google Scholar] [CrossRef]
- Guo, K.; Liu, T.; Xu, A.; Zhang, L.; Bian, X.; Wei, C. Structural and functional properties of starches from root tubers of white, yellow, and purple sweet potatoes. Food Hydrocoll. 2019, 89, 829–836. [Google Scholar] [CrossRef]
- Zhu, F.; Wang, S. Physicochemical properties, molecular structure, and uses of sweetpotato starch. Trends Food Sci. Technol. 2014, 36, 68–78. [Google Scholar] [CrossRef]
- Dhital, S.; Shrestha, A.K.; Hasjim, J.; Gidley, M.J. Physicochemical and structural properties of maize and potato starches as a function of granule size. J. Agric. Food Chem. 2011, 59, 10151–10161. [Google Scholar] [CrossRef] [PubMed]
- Genkina, N.K.; Wasserman, L.A.; Noda, T.; Tester, R.F.; Yuryev, V.P. Effects of annealing on the polymorphic structure of starches from sweet potatoes (Ayamurasaki and Sunnyred cultivars) grown at various soil temperatures. Carbohydr. Res. 2004, 339, 1093–1098. [Google Scholar] [CrossRef] [PubMed]
- Guo, K.; Zhang, L.; Bian, X.; Cao, Q.; Wei, C. A-, B- and C-type starch granules coexist in root tuber of sweet potato. Food Hydrocoll. 2020, 98, 105279. [Google Scholar] [CrossRef]
- Kim, J.; Ren, C.; Shin, M. Physicochemical properties of starch isolated from eight different varieties of Korean sweet potatoes. Starch-Stärke 2013, 65, 923–930. [Google Scholar] [CrossRef]
- Wang, H.; Yang, Q.; Ferdinand, U.; Gong, X.; Qu, Y.; Gao, W.; Ivanistau, A.; Feng, B.; Liu, M. Isolation and characterization of starch from light yellow, orange, and purple sweet potatoes. Int. J. Biol. Macromol. 2020, 160, 660–668. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, L.; Bian, X.; Guo, K.; Zhou, L.; Wei, C. Characterization and comparative study of starches from seven purple sweet potatoes. Food Hydrocoll. 2018, 80, 168–176. [Google Scholar] [CrossRef]
- Zhu, F.; Xie, Q. Rheological and thermal properties in relation to molecular structure of New Zealand sweetpotato starch. Food Hydrocoll. 2018, 83, 165–172. [Google Scholar] [CrossRef]
- Frost, K.; Kaminski, D.; Kriwan, G.; Lascaris, E.; Shanks, R. Crystallinity and structure of starch using wide angle X-ray scattering. Carbohydr. Polym. 2009, 78, 543–548. [Google Scholar] [CrossRef]
- Pérez, S.; Baldwin, P.M.; Gallant, D.J. Structural features of starch granules I. In Starch: Chemistry and Technology, 3rd ed.; BeMiller, J., Whistler, R., Eds.; Elsevier Inc.: New York, NY, USA, 2009; pp. 149–192. [Google Scholar]
- Noda, T.; Kobayashi, T.; Suda, I. Effect of soil temperature on starch properties of sweet potatoes. Carbohydr. Polym. 2001, 44, 239–246. [Google Scholar] [CrossRef]
- Man, J.; Lin, L.; Wang, Z.; Wang, Y.; Liu, Q.; Wei, C. Different structures of heterogeneous starch granules from high-amylose rice. J. Agric. Food Chem. 2014, 62, 11254–11263. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Man, J.; Huang, J.; Liu, Q.; Wei, W.; Wei, C. Relationship between structure and functional properties of normal rice starches with different amylose contents. Carbohydr. Polym. 2015, 125, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Guo, D.; Zhao, L.; Zhang, X.; Wang, J.; Zhang, F.; Wei, C. Comparative structure of starches from high-amylose maize inbred lines and their hybrids. Food Hydrocoll. 2016, 52, 19–28. [Google Scholar] [CrossRef]
- Lin, L.; Zhang, Q.; Zhang, L.; Wei, C. Evaluation of the molecular structural parameters of normal rice starch and their relationships with its thermal and digestion properties. Molecules 2017, 22, 1526. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Sreenivasulu, N.; Liu, Q. Waxy editing: Old meets new. Trends Plant Sci. 2020, 25, 963–966. [Google Scholar] [CrossRef]
- Lin, L.; Cai, C.; Gilbert, R.G.; Li, E.; Wang, J.; Wei, C. Relationships between amylopectin molecular structures and functional properties of different-sized fractions of normal and high-amylose maize starches. Food Hydrocoll. 2016, 52, 359–368. [Google Scholar] [CrossRef] [Green Version]
- Duan, W.; Zhang, H.; Xie, B.; Wang, B.; Zhang, L. Impacts of nitrogen fertilization rate on the root yield, starch yield and starch physicochemical properties of the sweet potato cultivar Jishu 25. PLoS ONE 2019, 14, e0221351. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.H.; Lee, Y.T. Physicochemical and structural properties of different colored sweet potato starches. Starch 2017, 69, 1600001. [Google Scholar] [CrossRef]
- Osundahunsi, O.F.; Fagbemi, T.N.; Kesselman, E.; Shimoni, E. Comparison of the physicochemical properties and pasting characteristics of flour and starch from red and white sweet potato cultivars. J. Agric. Food Chem. 2003, 51, 2232–2236. [Google Scholar] [CrossRef]
- Collado, L.S.; Mabesa, R.C.; Corke, H. Genetic variation in the physical properties of sweet potato starch. J. Agric. Food Chem. 1999, 47, 4195–5201. [Google Scholar] [CrossRef]
- Shar, T.; Sheng, Z.H.; Ali, U.; Fiaz, S.; Wei, X.J.; Xie, L.H.; Jiao, C.A.; Ali, F.; Shao, G.N.; Hu, S.K.; et al. Mapping quantitative trait loci associated with starch paste viscosity attributes by using double haploid populations of rice (Oryza sativa L.). J. Int. Agric. 2020, 19, 1691–1703. [Google Scholar] [CrossRef]
- Singh, N.; Kaur, L.; Ezekiel, R.; Guraya, H.S. Microstructural, cooking and textural characteristics of potato (Solanum tuberosum L) tubers in relation to physicochemical and functional properties of their flours. J. Sci. Food Agric. 2005, 85, 1275–1284. [Google Scholar] [CrossRef]
- Abegunde, O.K.; Mu, T.H.; Chen, J.W.; Deng, F.M. Physicochemical characterization of sweet potato starches popularly used in Chinese starch industry. Food Hydrocoll. 2013, 33, 169–177. [Google Scholar] [CrossRef]
- Hellemans, T.; Abera, G.; De Leyn, I.; Van der Meeren, P.; Dewettinck, K.; Eeckhout, M.; De Meulenaer, B.; Van Bockstaele, F. Composition, granular structure, and pasting properties of native starch extracted from Plectranthus edulis (Oromo dinich) tubers. J. Food Sci. 2017, 82, 2794–2804. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Guo, K.; Zhang, B.; Wei, C. Comparison of physicochemical properties of very small granule starches from endosperms of dicotyledon plants. Int. J. Biol. Macromol. 2020, 154, 818–825. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhao, L.; Shi, L.; Lin, L.; Cao, Q.; Wei, C. Sizes, components, crystalline structure, and thermal properties of starches from sweet potato varieties originating from different countries. Molecules 2022, 27, 1905. [Google Scholar] [CrossRef]
- Lin, L.; Zhao, S.; Li, E.; Guo, D.; Wei, C. Structural properties of starch from single kernel of high-amylose maize. Food Hydrocoll. 2022, 124, 107349. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession ID | RI of Diffraction Peak (Counts) | RC of Diffraction Peak (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| RI5.6° | RI15° | RI23° | RC5.6° | RC15° | RC17–18° | RC23° | RCTotal | |
| SY192 | 146 ± 9 a | 716 ± 6 d | 592 ± 2 f | 1.23 ± 0.18 a | 3.50 ± 0.10 d | 9.70 ± 0.14 de | 8.35 ± 0.28 b | 22.84 ± 0.31 cd |
| SY6 | 115 ± 7 b | 761 ± 1 bc | 617 ± 4 e | 0.74 ± 0.08 b | 3.63 ± 0.12 cd | 9.27 ± 0.01 e | 8.15 ± 0.22 b | 21.96 ± 0.46 d |
| SY148 | 80 ± 1 c | 751 ± 2 c | 639 ± 2 d | 0.56 ± 0.08 bc | 3.94 ± 0.06 bc | 11.17 ± 0.01 bc | 8.35 ± 0.40 b | 24.23 ± 0.65 bc |
| SY175 | 79 ± 2 c | 778 ± 3 bc | 676 ± 5 c | 0.46 ± 0.04 bc | 3.71 ± 0.21 cd | 10.21 ± 0.25 d | 9.77 ± 0.08 a | 24.21 ± 0.41 bc |
| SY4 | 69 ± 2 c | 788 ± 12 b | 706 ± 8 b | 0.34 ± 0.05 cd | 4.40 ± 0.01 a | 10.88 ± 0.08 c | 9.02 ± 0.28 ab | 24.84 ± 0.28 ab |
| SY19 | 67 ± 4 c | 751 ± 15 c | 708 ± 3 b | 0.33 ± 0.10 cd | 4.16 ± 0.10 ab | 11.50 ± 0.33 ab | 9.69 ± 0.20 a | 25.93 ± 0.45 a |
| Kokei 14 | 38 ± 3 d | 826 ± 6 a | 727 ± 5 a | 0.09 ± 0.01 d | 4.20 ± 0.12 ab | 11.92 ± 0.13 a | 9.10 ± 0.41 ab | 25.49 ± 0.34 ab |
| Sig. | 0.558 | 0.906 | 0.547 | 0.516 | 0.713 | 0.830 | 0.275 | 0.723 |
| Accession ID | OD620/550 | AAC (%) | TAC (%) | Components of Isoamylase-Debranched Starch | |||
|---|---|---|---|---|---|---|---|
| AM (%) | AP-L (%) | AP-S (%) | AP-S/L | ||||
| SY192 | 1.208 ± 0.011 a | 21.8 ± 0.2 c | 14.7 ± 0.3 d | 20.7 ± 0.1 a | 21.5 ± 0.2 c | 57.8 ± 0.3 ab | 2.69 ± 0.04 a |
| SY6 | 1.203 ± 0.018 ab | 20.2 ± 0.3 c | 15.2 ± 0.7 d | 19.3 ± 1.0 ab | 23.8 ± 0.7 ab | 56.9 ± 0.3 abc | 2.39 ± 0.06 b |
| SY148 | 1.186 ± 0.001 abc | 28.5 ± 0.2 a | 17.2 ± 0.5 bc | 19.9 ± 0.6 ab | 23.9 ± 0.1 ab | 56.3 ± 0.5 bcd | 2.36 ± 0.01 b |
| SY175 | 1.146 ± 0.015 d | 29.2 ± 1.3 a | 17.7 ± 0.7 bc | 19.8 ± 0.2 ab | 25.1 ± 0.1 a | 55.1 ± 0.3 d | 2.19 ± 0.02 b |
| SY4 | 1.220 ± 0.018 a | 21.5 ± 1.0 c | 16.2 ± 0.1 cd | 19.5 ± 0.3 ab | 22.2 ± 0.1 bc | 58.3 ± 0.3 a | 2.62 ± 0.01 a |
| SY19 | 1.160 ± 0.012 cd | 25.7 ± 0.8 b | 18.2 ± 0.4 ab | 18.7 ± 0.4 b | 25.3 ± 0.9 a | 56.0 ± 0.5 cd | 2.21 ± 0.10 b |
| Kokei 14 | 1.169 ± 0.019 bcd | 28.3 ± 0.2 a | 19.7 ± 0.4 a | 19.7 ± 0.3 ab | 24.2 ± 1.1 ab | 56.2 ± 0.9 bcd | 2.33 ± 0.14 b |
| Sig. | 0.787 | 0.156 | 0.925 | 0.871 | 0.440 | 0.810 | 0.331 |
| Accession ID | DP6-12 (%) | DP13-24 (%) | DP25-36 (%) | DP > 36 (%) | ACL (DP) |
|---|---|---|---|---|---|
| SY192 | 25.1 ± 0.5 a | 44.0 ± 0.6 e | 13.9 ± 0.1 a | 17.0 ± 0.1 ab | 23.5 ± 0.1 ab |
| SY6 | 23.5 ± 0.2 b | 45.7 ± 0.4 d | 13.0 ± 0.2 b | 17.7 ± 0.4 ab | 23.7 ± 0.1 ab |
| SY148 | 20.2 ± 0.5 d | 48.9 ± 0.3 a | 12.7 ± 0.1 b | 18.1 ± 0.7 ab | 24.1 ± 0.4 ab |
| SY175 | 21.1 ± 0.1 cd | 46.2 ± 0.3 cd | 13.8 ± 0.1 a | 18.8 ± 0.4 a | 24.6 ± 0.2 a |
| SY4 | 22.4 ± 0.7 bc | 49.0 ± 0.6 a | 13.0 ± 0.1 b | 15.6 ± 1.4 b | 22.9 ± 0.7 b |
| SY19 | 20.3 ± 0.4 d | 48.4 ± 0.4 ab | 12.8 ± 0.1 b | 18.4 ± 0.7 a | 24.3 ± 0.3 a |
| Kokei 14 | 21.4 ± 0.3 cd | 47.4 ± 0.1 bc | 13.8 ± 0.3 a | 17.5 ± 0.2 ab | 24.0 ± 0.1 ab |
| Sig. | 0.438 | 0.467 | 0.066 | 0.583 | 0.924 |
| RI5.6° | RI15° | RI23° | RC5.6° | RC15° | RC17–18° | RC23° | RCTotal | |
|---|---|---|---|---|---|---|---|---|
| OD620/550 | 0.486 | −0.270 | −0.431 | 0.466 | −0.006 | −0.393 | −0.735 | −0.490 |
| AAC | −0.613 | 0.370 | 0.436 | −0.521 | 0.175 | 0.614 | 0.524 | 0.574 |
| TAC | −0.903 ** | 0.703 | 0.822 * | −0.856 * | 0.571 | 0.855 * | 0.651 | 0.824 * |
| AM | 0.530 | −0.306 | −0.578 | 0.635 | −0.533 | −0.346 | −0.448 | −0.447 |
| AP-L | −0.566 | 0.329 | 0.471 | −0.601 | 0.144 | 0.376 | 0.615 | 0.440 |
| AP-S | 0.409 | −0.232 | −0.272 | 0.400 | 0.123 | −0.263 | −0.548 | −0.301 |
| AP-S/L | 0.544 | −0.324 | −0.432 | 0.573 | −0.091 | −0.350 | −0.596 | −0.404 |
| DP6-12 | 0.792 * | −0.397 | −0.643 | 0.764 * | −0.547 | −0.734 | −0.570 | −0.731 |
| DP13-24 | −0.764 * | 0.424 | 0.676 | −0.752 | 0.864 * | 0.746 | 0.311 | 0.711 |
| DP25-36 | 0.161 | 0.161 | −0.068 | 0.209 | −0.385 | −0.179 | 0.160 | −0.128 |
| DP > 36 | −0.094 | −0.116 | −0.044 | −0.087 | −0.384 | 0.042 | 0.335 | 0.067 |
| ACL | −0.265 | 0.040 | 0.139 | −0.241 | −0.240 | 0.211 | 0.491 | 0.250 |
| Accession ID | Thermal Property Parameters | SP (g/g) | WS (%) | ||||
|---|---|---|---|---|---|---|---|
| To (°C) | Tp (°C) | Tc (°C) | ΔT (°C) | ΔH (J/g) | |||
| SY192 | 54.4 ± 0.2 f | 65.8 ± 0.7 e | 76.6 ± 1.2 d | 22.2 ± 1.0 b | 12.6 ± 0.3 c | 26.2 ± 0.5 a | 10.4 ± 0.8 b |
| SY6 | 57.9 ± 0.1 e | 69.9 ± 0.1 d | 79.9 ± 0.1 c | 22.0 ± 0.1 b | 14.9 ± 0.3 b | 27.1 ± 0.3 a | 10.7 ± 0.6 b |
| SY148 | 61.0 ± 0.4 c | 66.8 ± 0.1 e | 79.7 ± 0.1 c | 18.7 ± 0.3 c | 14.7 ± 0.3 b | 27.6 ± 0.4 a | 11.0 ± 0.2 ab |
| SY175 | 60.3 ± 0.1 d | 72.5 ± 0.4 c | 84.7 ± 0.6 b | 24.4 ± 0.6 a | 14.7 ± 0.2 b | 24.5 ± 0.4 b | 10.7 ± 0.1 b |
| SY4 | 60.3 ± 0.1 d | 74.0 ± 0.3 b | 85.8 ± 0.8 ab | 25.5 ± 0.9 a | 14.4 ± 0.3 b | 26.4 ± 0.4 a | 11.8 ± 0.4 ab |
| SY19 | 64.0 ± 0.1 b | 73.6 ± 0.1 b | 84.8 ± 0.4 b | 20.8 ± 0.4 b | 16.8 ± 0.5 a | 26.4 ± 0.9 a | 12.3 ± 0.6 a |
| Kokei 14 | 73.8 ± 0.1 a | 80.0 ± 0.1 a | 86.9 ± 0.1 a | 13.1 ± 0.1 d | 16.2 ± 0.1 a | 24.0 ± 0.3 b | 12.4 ± 0.8 a |
| Sig. | 0.232 | 0.728 | 0.311 | 0.424 | 0.528 | 0.328 | 0.192 |
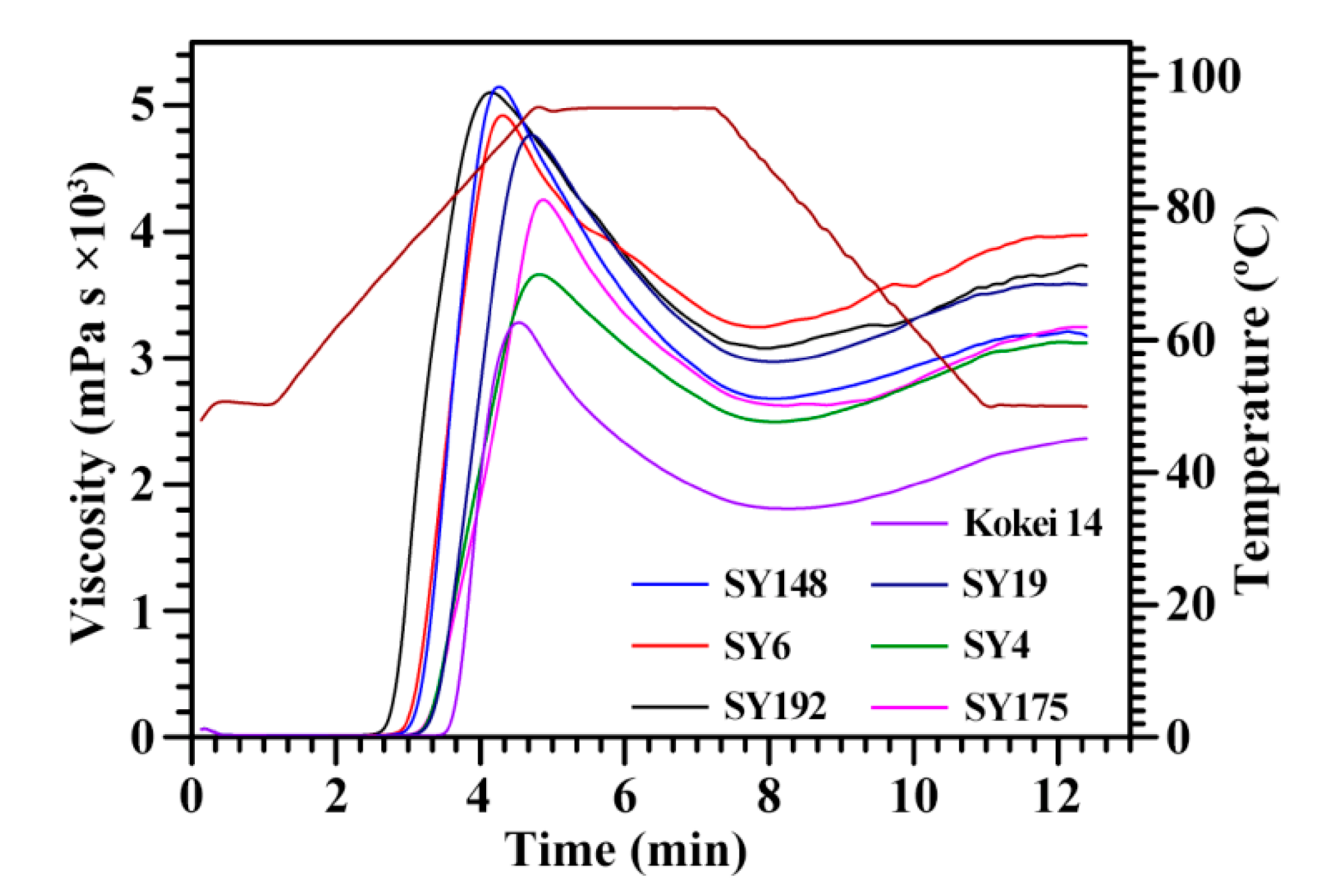
| Accession ID | PV (mPa·s) | HV (mPa·s) | BV (mPa·s) | FV (mPa·s) | SV (mPa·s) | PTime (min) | PTemp (°C) |
|---|---|---|---|---|---|---|---|
| SY192 | 5140 ± 19 a | 3062 ± 16 b | 2078 ± 4 b | 3722 ± 48 b | 660 ± 65 b | 4.18 ± 0.08 e | 70.4 ± 0.5 d |
| SY6 | 4944 ± 51 b | 3220 ± 59 a | 1724 ± 9 d | 3993 ± 42 a | 773 ± 34 a | 4.31 ± 0.08 d | 73.6 ± 0.5 c |
| SY148 | 5152 ± 36 a | 2648 ± 26 c | 2503 ± 14 a | 3148 ± 44 de | 500 ± 35 d | 4.27 ± 0.01 de | 74.2 ± 0.1 c |
| SY175 | 4257 ± 52 c | 2579 ± 35 c | 1678 ± 29 d | 3215 ± 27 d | 636 ± 9 bc | 4.82 ± 0.04 a | 76.6 ± 0.1 b |
| SY4 | 3651 ± 31 d | 2478 ± 13 d | 1173 ± 18 f | 3100 ± 36 e | 622 ± 23 bc | 4.82 ± 0.04 a | 76.5 ± 0.1 b |
| SY19 | 4877 ± 58 b | 3011 ± 42 b | 1867 ± 32 c | 3622 ± 37 c | 611 ± 31 bc | 4.67 ± 0.01 b | 77.1 ± 0.5 b |
| Kokei 14 | 3265 ± 50 e | 1789 ± 16 e | 1476 ± 36 e | 2341 ± 20 f | 552 ± 6 cd | 4.56 ± 0.04 c | 81.4 ± 0.1 a |
| Sig. | 0.139 | 0.439 | 0.980 | 0.658 | 0.816 | 0.293 | 0.857 |
| RI5.6° | RI15° | RI23° | RC5.6° | RC15° | RC17–18° | RC23° | RCTotal | |
|---|---|---|---|---|---|---|---|---|
| To | −0.879 ** | 0.819 * | 0.804 * | −0.844 * | 0.631 | 0.827 * | 0.419 | 0.718 |
| Tp | −0.834 * | 0.900 ** | 0.902 ** | −0.860 * | 0.682 | 0.635 | 0.597 | 0.670 |
| Tc | −0.904 ** | 0.845 * | 0.974 ** | −0.934 ** | 0.784 * | 0.672 | 0.766 * | 0.782 * |
| ΔT | 0.458 | −0.424 | −0.280 | 0.378 | −0.201 | −0.599 | 0.097 | −0.333 |
| ΔH | −0.793 * | 0.560 | 0.760 * | −0.832 * | 0.596 | 0.679 | 0.543 | 0.682 |
| SP | 0.457 | −0.641 | −0.534 | 0.433 | −0.122 | −0.305 | −0.609 | −0.390 |
| WS | −0.825 * | 0.608 | 0.883 ** | −0.816 * | 0.879 ** | 0.870 * | 0.490 | 0.862 * |
| PV | 0.716 | −0.918 ** | −0.779 * | 0.721 | −0.651 | −0.500 | −0.433 | −0.517 |
| HV | 0.779 * | −0.854 * | −0.698 | 0.708 | −0.584 | −0.705 | −0.329 | −0.598 |
| BV | 0.397 | −0.672 | −0.601 | 0.486 | −0.501 | −0.095 | −0.400 | −0.246 |
| FV | 0.789 * | −0.803 * | −0.691 | 0.703 | −0.601 | −0.759 * | −0.330 | −0.643 |
| SV | 0.596 | −0.263 | −0.433 | 0.456 | −0.506 | −0.822 * | −0.226 | −0.690 |
| PTime | −0.650 | 0.579 | 0.799 * | −0.702 | 0.608 | 0.377 | 0.857 * | 0.628 |
| PTemp | −0.950 ** | 0.907 ** | 0.933 ** | −0.953 ** | 0.721 | 0.770 * | 0.625 | 0.766 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhao, L.; Lin, L.; Li, E.; Cao, Q.; Wei, C. Relationships between X-ray Diffraction Peaks, Molecular Components, and Heat Properties of C-Type Starches from Different Sweet Potato Varieties. Molecules 2022, 27, 3385. https://doi.org/10.3390/molecules27113385
Li Y, Zhao L, Lin L, Li E, Cao Q, Wei C. Relationships between X-ray Diffraction Peaks, Molecular Components, and Heat Properties of C-Type Starches from Different Sweet Potato Varieties. Molecules. 2022; 27(11):3385. https://doi.org/10.3390/molecules27113385
Chicago/Turabian StyleLi, Yibo, Lingxiao Zhao, Lingshang Lin, Enpeng Li, Qinghe Cao, and Cunxu Wei. 2022. "Relationships between X-ray Diffraction Peaks, Molecular Components, and Heat Properties of C-Type Starches from Different Sweet Potato Varieties" Molecules 27, no. 11: 3385. https://doi.org/10.3390/molecules27113385
APA StyleLi, Y., Zhao, L., Lin, L., Li, E., Cao, Q., & Wei, C. (2022). Relationships between X-ray Diffraction Peaks, Molecular Components, and Heat Properties of C-Type Starches from Different Sweet Potato Varieties. Molecules, 27(11), 3385. https://doi.org/10.3390/molecules27113385