
Elucidation of Interaction between Whey Proteins and Proanthocyanidins and Its Protective Effects on Proanthocyanidins during In-Vitro Digestion and Storage

Abstract
:1. Introduction
2. Results and Discussion
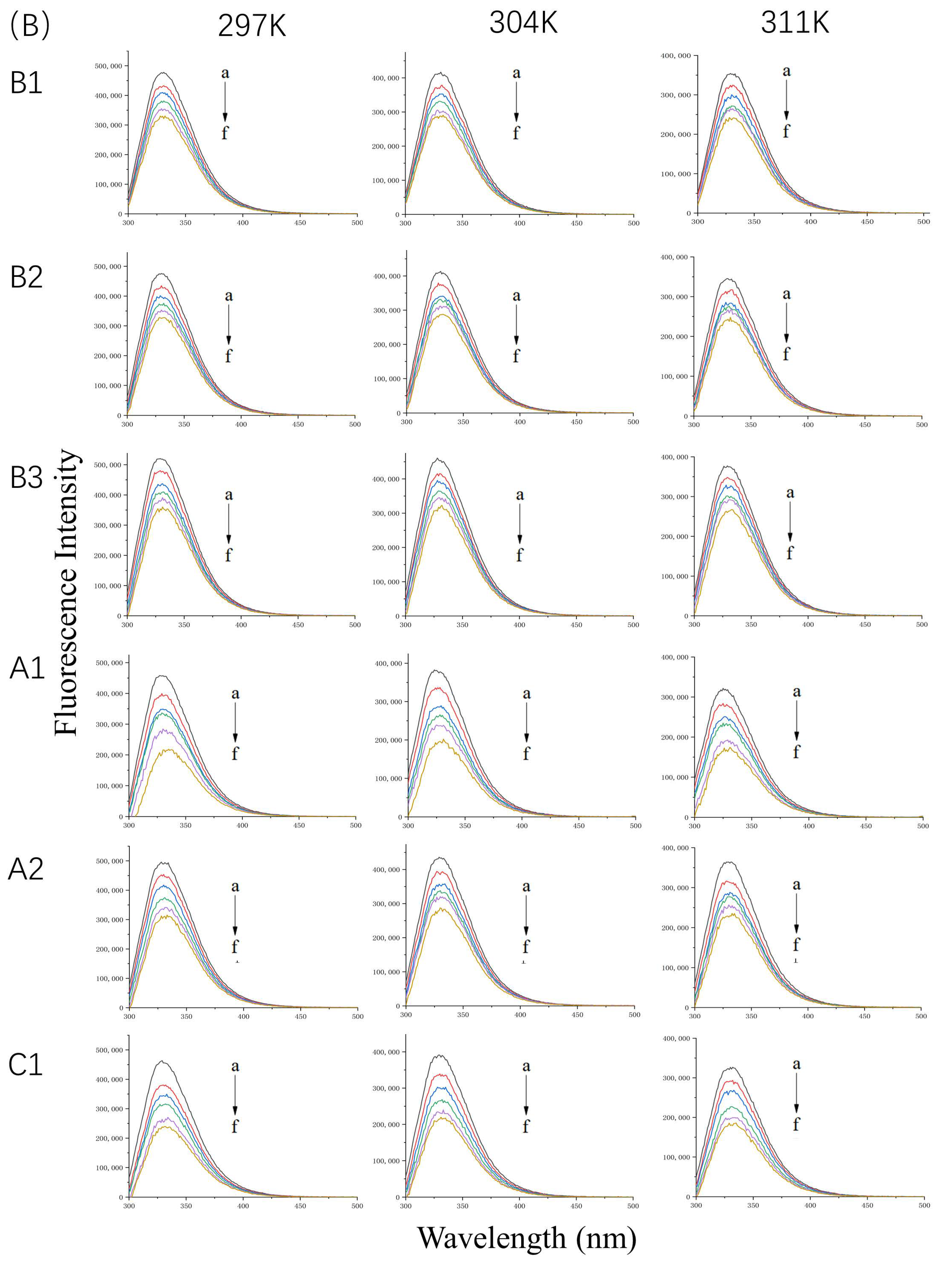
2.1. Analysis of Fluorescence Quenching
2.1.1. Fluorescence Spectra of WPI Bound to Proanthocyanidins
2.1.2. Fluorescence Spectra of β-LG or α-LA Bound to Proanthocyanidins
2.1.3. Binding Ability of β-LG or α-LA to Proanthocyanidins
2.1.4. Thermodynamic Parameters and Binding Forces between Proteins and Proanthocyanidins
2.2. Fourier-Transform Infrared Spectroscopy Analysis
2.3. Circular Dichroism Spectroscopy Analysis
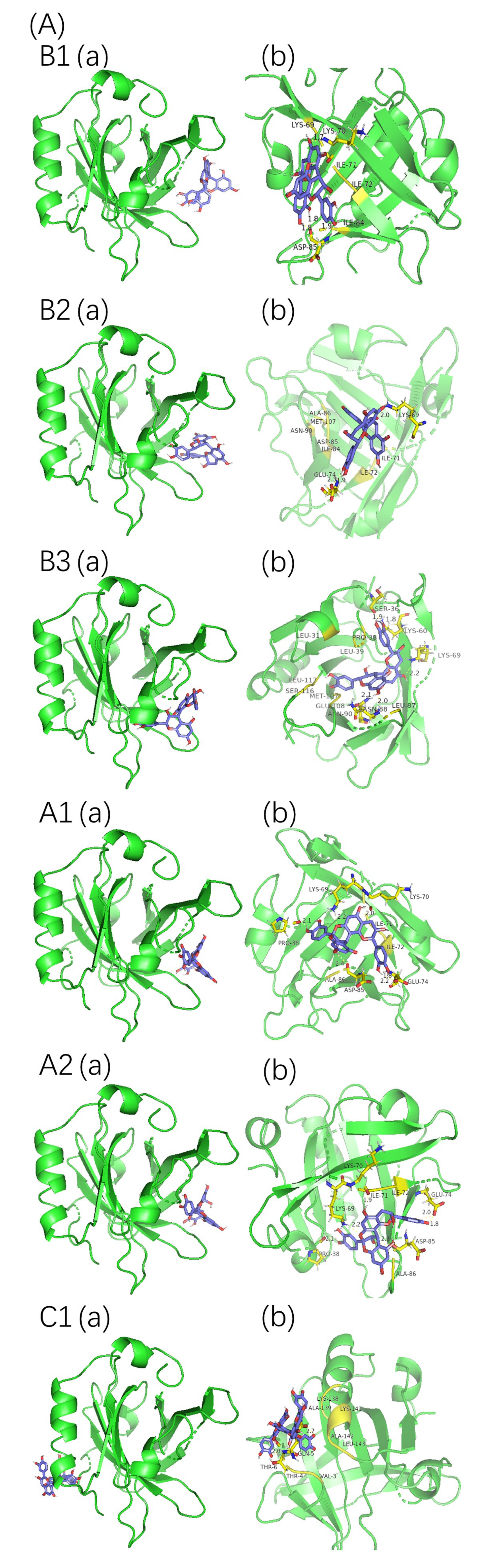
2.4. Molecular Docking Analysis
2.5. Effects of WPI on the Stability of Proanthocyanidins during In Vitro Gastrointestinal Digestion
2.6. Effect of WPI on the Stability of Proanthocyanidins during Storage at Room Temperature
3. Materials and Methods
3.1. Materials
3.2. Fluorescence Spectroscopy
3.3. Fourier-Transform Infrared Spectroscopy
3.4. Circular Dichroism Spectroscopy
3.5. Molecular Docking
3.6. Assessment of Stability of Proanthocyanidin in WPI–Proanthocyanidin Mixture during In Vitro Gastrointestinal Digestion
3.7. Assessment of the Storage Stability of Proanthocyanidins in WPI–Proanthocyanidin Mixture
3.8. High-Performance Liquid Chromatography (HPLC) Analysis of Proanthocyanidins
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hellström, J.K.; Törrönen, A.R.; Mattila, P.H. Proanthocyanidins in Common Food Products of Plant Origin. J. Agric. Food Chem. 2009, 57, 7899–7906. [Google Scholar] [CrossRef]
- Lai, R.; Xian, D.; Xiong, X.; Yang, L.; Song, J.; Zhong, J. Proanthocyanidins: Novel treatment for psoriasis that reduces oxidative stress and modulates Th17 and Treg cells. Redox Rep. 2018, 23, 130–135. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.-L.; Pan, C.; Wang, L.; Ding, L.; Guo, K.; Wang, H.-Z.; Xu, A.-M.; Gao, S. Protective effects of grape seed proanthocyanidins on cardiovascular remodeling in DOCA-salt hypertension rats. J. Nutr. Biochem. 2015, 26, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, X.; Dai, T.; Liu, C.; Li, T.; McClements, D.J.; Chen, J.; Liu, J. Proanthocyanidins, Isolated from Choerospondias axillaris Fruit Peels, Exhibit Potent Antioxidant Activities in Vitro and a Novel Anti-angiogenic Property in Vitro and in Vivo. J. Agric. Food Chem. 2016, 64, 3546–3556. [Google Scholar] [CrossRef] [PubMed]
- Kruger, M.J.; Davies, N.; Myburgh, K.H.; Lecour, S. Proanthocyanidins, anthocyanins and cardiovascular diseases. Food Res. Int. 2014, 59, 41–52. [Google Scholar] [CrossRef]
- Ou, K.; Gu, L. Absorption and metabolism of proanthocyanidins. J. Funct. Foods 2014, 7, 43–53. [Google Scholar] [CrossRef]
- Unusan, N. Proanthocyanidins in grape seeds: An updated review of their health benefits and potential uses in the food industry. J. Funct. Foods 2020, 67, 103861. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Abu-Izneid, T.; Iahtisham Ul, H.; Patel, S.; Pan, X.; Naz, S.; Sanches Silva, A.; Saeed, F.; Rasul Suleria, H.A. Proanthocyanidins: A comprehensive review. Biomed. Pharmacother. 2019, 116, 108999. [Google Scholar] [CrossRef]
- Zang, Z.; Chou, S.; Tian, J.; Lang, Y.; Shen, Y.; Ran, X.; Gao, N.; Li, B. Effect of whey protein isolate on the stability and antioxidant capacity of blueberry anthocyanins: A mechanistic and in vitro simulation study. Food Chem. 2021, 336, 127700. [Google Scholar] [CrossRef]
- Sun, X.M.; Wang, C.N.; Guo, M.R. Interactions between whey protein or polymerized whey protein and soybean lecithin in model system. J. Dairy Sci. 2018, 101, 9680–9692. [Google Scholar] [CrossRef] [Green Version]
- Playne, M.J.; Bennett, L.E.; Smithers, G.W. Functional dairy foods and ingredients. Aust. J. Dairy Technol. 2003, 58, 242. [Google Scholar]
- Wang, B.; Heinonen, M. Protein–Tannin Interactions of Tryptic Digests of α-Lactalbumin and Procyanidins. J. Agric. Food Chem. 2017, 65, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; McClements, D.J.; Decker, E.A. Antioxidant Activity of a Proanthocyanidin-Rich Extract from Grape Seed in Whey Protein Isolate Stabilized Algae Oil-in-Water Emulsions. J. Agric. Food Chem. 2004, 52, 5272–5276. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, R.; Xie, B.; Sun, Z.; McClements, D.J. Lotus seedpod proanthocyanidin-whey protein complexes: Impact on physical and chemical stability of β-carotene-nanoemulsions. Food Res. Int. 2020, 127, 108738. [Google Scholar] [CrossRef] [PubMed]
- Albani, J.R.; Vogelaer, J.; Bretesche, L.; Kmiecik, D. Tryptophan 19 residue is the origin of bovine β-lactoglobulin fluorescence. J. Pharm. Biomed. Anal. 2014, 91, 144–150. [Google Scholar] [CrossRef]
- Al-Hanish, A.; Stanic-Vucinic, D.; Mihailovic, J.; Prodic, I.; Minic, S.; Stojadinovic, M.; Radibratovic, M.; Milcic, M.; Cirkovic Velickovic, T. Noncovalent interactions of bovine α-lactalbumin with green tea polyphenol, epigalocatechin-3-gallate. Food Hydrocoll. 2016, 61, 241–250. [Google Scholar] [CrossRef]
- Lakowicz, J.R.; Weber, G. Quenching of fluorescence by oxygen. Probe for structural fluctuations in macromolecules. Biochemistry 1973, 12, 4161–4170. [Google Scholar]
- Hu, Y.; Liu, Y.; Zhao, R.; Dong, J.; Qu, S. Spectroscopic studies on the interaction between methylene blue and bovine serum albumin. J. Photochem. Photobiol. A Chem. 2006, 179, 324–329. [Google Scholar] [CrossRef]
- Zuo, H.; Tang, L.; Li, S.; Huang, J. Combined multispectroscopic and molecular docking investigation on the interaction between delphinidin-3-O-glucoside and bovine serum albumin. Luminescence 2015, 30, 110–117. [Google Scholar] [CrossRef]
- van de Weert, M. Fluorescence Quenching to Study Protein-ligand Binding: Common Errors. J. Fluoresc. 2010, 20, 625–629. [Google Scholar] [CrossRef]
- Prigent, S.V.E.; Voragen, A.G.J.; van Koningsveld, G.A.; Baron, A.; Renard, C.M.G.C.; Gruppen, H. Interactions between globular proteins and procyanidins of different degrees of polymerization. J. Dairy Sci. 2009, 92, 5843–5853. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.; Tang, C.; Sun, X.; Zhang, J. The interaction mechanism of β-casein with oligomeric proanthocyanidins and its effect on proanthocyanidin bioaccessibility. Food Hydrocoll. 2021, 113, 106485. [Google Scholar] [CrossRef]
- Bohin, M.C.; Vincken, J.-P.; Westphal, A.H.; Tripp, A.M.; Dekker, P.; van der Hijden, H.T.W.M.; Gruppen, H. Interaction of flavan-3-ol derivatives and different caseins is determined by more than proline content and number of proline repeats. Food Chem. 2014, 158, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Prigent, S.V.E.; Gruppen, H.; Visser, A.J.W.G.; van Koningsveld, G.A.; de Jong, G.A.H.; Voragen, A.G.J. Effects of Non-covalent Interactions with 5-O-Caffeoylquinic Acid (Chlorogenic Acid) on the Heat Denaturation and Solubility of Globular Proteins. J. Agric. Food Chem. 2003, 51, 5088–5095. [Google Scholar] [CrossRef] [PubMed]
- Jakobek, L. Interactions of polyphenols with carbohydrates, lipids and proteins. Food Chem. 2015, 175, 556–567. [Google Scholar] [CrossRef]
- Freitas, V.D.; Mateus, N. Nephelometric study of salivary protein–tannin aggregates. J. Sci. Food Agric. 2002, 82, 113–119. [Google Scholar] [CrossRef]
- Ross, P.D.; Subramanian, S. Thermodynamics of protein association reactions: Forces contributing to stability. Biochemistry 1981, 20, 3096–3102. [Google Scholar] [CrossRef]
- Zhang, Y.; Wright, E.; Zhong, Q. Effects of pH on the Molecular Binding between β-Lactoglobulin and Bixin. J. Agric. Food Chem. 2013, 61, 947–954. [Google Scholar] [CrossRef]
- He, Z.; Xu, M.; Zeng, M.; Qin, F.; Chen, J. Interactions of milk α- and β-casein with malvidin-3-O-glucoside and their effects on the stability of grape skin anthocyanin extracts. Food Chem. 2016, 199, 314–322. [Google Scholar] [CrossRef]
- Zeng, H.Y.; Cai, L.H.; Cai, X.L.; Wang, Y.J.; Li, Y.Q. Structure characterization of protein fractions from lotus (Nelumbo nucifera) seed. J. Mol. Struct. 2011, 1001, 139–144. [Google Scholar] [CrossRef]
- Yampolskaya, G.P.; Tarasevich, B.N.; Elenskii, A.A. Secondary Structure of Globular Proteins in Adsorption Layers at the Solution-Air Interface by the Data of Fourier Transform IR Spectroscopy. Colloid J. 2005, 67, 385–391. [Google Scholar] [CrossRef]
- Agyare, K.K.; Damodaran, S. pH-Stability and Thermal Properties of Microbial Transglutaminase-Treated Whey Protein Isolate. J. Agric. Food Chem. 2010, 58, 1946–1953. [Google Scholar] [CrossRef] [PubMed]
- Kelly, S.M.; Price, N.C. The Use of Circular Dichroism in the Investigation of Protein Structure and Function. Curr. Protein Pept. Sci. 2000, 1, 349–384. [Google Scholar] [CrossRef] [Green Version]
- Kanakis, C.D.; Hasni, I.; Bourassa, P.; Tarantilis, P.A.; Polissiou, M.G.; Tajmir-Riahi, H.-A. Milk β-lactoglobulin complexes with tea polyphenols. Food Chem. 2011, 127, 1046–1055. [Google Scholar] [CrossRef]
- Chakraborty, S.; Ittah, V.; Bai, P.; Luo, L.; Haas, E.; Peng, Z.Y. Structure and Dynamics of the α-Lactalbumin Molten Globule: Fluorescence Studies Using Proteins Containing a Single Tryptophan Residue. Biochemistry 2001, 40, 7228–7238. [Google Scholar] [CrossRef] [PubMed]
- Roufic, S.; Gauthier, S.F.; Leng, X.; Turgeon, S.L. Thermodynamics of binding interactions between bovine β-lactoglobulin A and the antihypertensive peptide β-Lg f142-148. Biomacromolecules 2006, 7, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Delavari, B.; Saboury, A.A.; Atri, M.S.; Ghasemi, A.; Bigdeli, B.; Khammari, A.; Maghami, P.; Moosavi-Movahedi, A.A.; Haertlé, T.; Goliaei, B. Alpha-lactalbumin: A new carrier for vitamin D3 food enrichment. Food Hydrocoll. 2015, 45, 124–131. [Google Scholar] [CrossRef]
- Xie, F.; Zhang, W.; Gong, S.; Gu, X.; Lan, X.; Wu, J.; Wang, Z. Investigating lignin from Canna edulis ker residues induced activation of α-amylase: Kinetics, interaction, and molecular docking. Food Chem. 2019, 271, 62–69. [Google Scholar] [CrossRef]
- Pineda-Vadillo, C.; Nau, F.; Dubiard, C.G.; Cheynier, V.; Meudec, E.; Sanz-Buenhombre, M.; Guadarrama, A.; Tóth, T.; Csavajda, É.; Hingyi, H.; et al. In vitro digestion of dairy and egg products enriched with grape extracts: Effect of the food matrix on polyphenol bioaccessibility and antioxidant activity. Food Res. Int. 2016, 88, 284–292. [Google Scholar] [CrossRef]
- Zhu, Q.Y.; Holt, R.R.; Lazarus, S.A.; Ensunsa, J.L.; Hammerstone, J.F.; Schmitz, H.H.; Keen, C.L. Stability of the Flavan-3-ols Epicatechin and Catechin and Related Dimeric Procyanidins Derived from Cocoa. J. Agric. Food Chem. 2002, 50, 1700–1705. [Google Scholar] [CrossRef]
- Moser, S.; Chegeni, M.; Jones, O.G.; Liceaga, A.; Ferruzzi, M.G. The effect of milk proteins on the bioaccessibility of green tea flavan-3-ols. Food Res. Int. 2014, 66, 297–305. [Google Scholar] [CrossRef]
- Hernández-Ledesma, B.; Dávalos, A.; Bartolomé, B.; Amigo, L. Preparation of Antioxidant Enzymatic Hydrolysates from α-Lactalbumin and β-Lactoglobulin. Identification of Active Peptides by HPLC-MS/MS. J. Agric. Food Chem. 2005, 53, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Lamothe, S.; Azimy, N.; Bazinet, L.; Couillard, C.; Britten, M. Interaction of green tea polyphenols with dairy matrices in a simulated gastrointestinal environment. Food Funct. 2014, 5, 2621–2631. [Google Scholar] [CrossRef]
- Ozdal, T.; Capanoglu, E.; Altay, F. A review on protein-phenolic interactions and associated changes. Food Res. Int. 2013, 51, 954–970. [Google Scholar] [CrossRef]
- van der Burg-Koorevaar, M.C.D.; Miret, S.; Duchateau, G.S.M.J.E. Effect of Milk and Brewing Method on Black Tea Catechin Bioaccessibility. J. Agric. Food Chem. 2011, 59, 7752–7758. [Google Scholar] [CrossRef] [PubMed]
- Quan, W.; Qie, X.; Chen, Y.; Zeng, M.; Qin, F.; Chen, J.; He, Z. Effect of milk addition and processing on the antioxidant capacity and phenolic bioaccessibility of coffee by using an in vitro gastrointestinal digestion model. Food Chem. 2020, 308, 125598. [Google Scholar] [CrossRef]
- Rashidinejad, A.; Birch, E.J.; Sun-Waterhouse, D.; Everett, D.W. Addition of milk to tea infusions: Helpful or harmful? Evidence from in vitro and in vivo studies on antioxidant properties. Crit. Rev. Food Sci. Nutr. 2017, 57, 3188–3196. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Xu, M.; Zeng, M.; Qin, F.; Chen, J. Preheated milk proteins improve the stability of grape skin anthocyanins extracts. Food Chem. 2016, 210, 221–227. [Google Scholar] [CrossRef]
- Chung, C.; Rojanasasithara, T.; Mutilangi, W.; McClements, D.J. Enhanced stability of anthocyanin-based color in model beverage systems through whey protein isolate complexation. Food Res. Int. 2015, 76, 761–768. [Google Scholar] [CrossRef]
- Liang, L.; Subirade, M. Study of the acid and thermal stability of β-lactoglobulin–ligand complexes using fluorescence quenching. Food Chem. 2012, 132, 2023–2029. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Verzelloni, E.; Bertolini, D.; Conte, A. In vitro bio-accessibility and antioxidant activity of grape polyphenols. Food Chem. 2010, 120, 599–606. [Google Scholar] [CrossRef]
- Condict, L.; Kaur, J.; Hung, A.; Ashton, J.; Kasapis, S. Combined spectroscopic, molecular docking and quantum mechanics study of β-casein and ferulic acid interactions following UHT-like treatment. Food Hydrocoll. 2019, 89, 351–359. [Google Scholar] [CrossRef]
- Kaur, J.; Katopo, L.; Hung, A.; Ashton, J.; Kasapis, S. Combined spectroscopic, molecular docking and quantum mechanics study of β-casein and p-coumaric acid interactions following thermal treatment. Food Chem. 2018, 252, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Ma, Y. Mechanistic and conformational studies on the interaction of food dye amaranth with human serum albumin by multispectroscopic methods. Food Chem. 2013, 136, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Du, W.; Jin, J.; Du, Q. Preservation of (−)-Epigallocatechin-3-gallate Antioxidant Properties Loaded in Heat Treated β-Lactoglobulin Nanoparticles. J. Agric. Food Chem. 2012, 60, 3477–3484. [Google Scholar] [CrossRef] [PubMed]
- Jia, J.; Gao, X.; Hao, M.; Tang, L. Comparison of binding interaction between β-lactoglobulin and three common polyphenols using multi-spectroscopy and modeling methods. Food Chem. 2017, 228, 143–151. [Google Scholar] [CrossRef]
- Liu, J. Interaction of chlorogenic acid with milk proteins analyzed by spectroscopic and modeling methods. Spectrosc. Lett. 2016, 49, 44–50. [Google Scholar] [CrossRef]
- Zhou, X.M.; Lü, W.J.; Su, L.; Shan, Z.J.; Chen, X.G. Binding of Phthalate Plasticizers to Human Serum Albumin in Vitro: A Multispectroscopic Approach and Molecular Modeling. J. Agric. Food Chem. 2012, 60, 1135–1145. [Google Scholar] [CrossRef]
- Wootton-Beard, P.C.; Moran, A.; Ryan, L. Stability of the total antioxidant capacity and total polyphenol content of 23 commercially available vegetable juices before and after in vitro digestion measured by FRAP, DPPH, ABTS and Folin-Ciocalteu methods. Food Res. Int. 2011, 44, 217–224. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| T/K | Ksv (×104 L mol−1) | Kq (×1012 L mol−1 s−1) | n | Ka (×103 L mol−1) | ∆H (KJ mol−1) | ∆S (KJ mol−1 K−1) | ∆G (KJ mol−1) | |
|---|---|---|---|---|---|---|---|---|
| A1 | 297 | 1.61 ± 0.08 aB | 1.61 ± 0.08 aB | 1.06 ± 0.04 | 29.51 ± 4.47 aA | −40.45 | −0.05 | −25.45 |
| 304 | 1.59 ± 0.06 a | 1.59 ± 0.06 a | 1.04 ± 0.03 | 22.22 ± 1.97 a | −25.09 | |||
| 311 | 1.52 ± 0.07 a | 1.52 ± 0.07 a | 1.00 ± 0.06 | 14.51 ± 3.27 ab | −24.74 | |||
| A2 | 297 | 1.16 ± 0.02 aC | 1.16 ± 0.02 aC | 1.04 ± 0.03 | 18.59 ± 2.41 aB | −134.21 | −0.37 | −24.40 |
| 304 | 1.06 ± 0.01 b | 1.06 ± 0.01 b | 0.95 ± 0.02 | 6.50 ± 0.53 b | −21.82 | |||
| 311 | 1.01 ± 0.03 c | 1.01 ± 0.03 c | 0.82 ± 0.03 | 1.61 ± 0.22 b | −19.23 | |||
| B1 | 297 | 0.90 ± 0.01 aD | 0.90 ± 0.01 aD | 0.95 ± 0.04 | 5.60 ± 0.75 cD | 55.03 | 0.26 | −21.30 |
| 304 | 0.86 ± 0.02 b | 0.86 ± 0.02 b | 1.02 ± 0.01 | 10.81 ± 0.23 b | −23.10 | |||
| 311 | 0.85 ± 0.01 b | 0.85 ± 0.01 b | 1.05 ± 0.03 | 14.97 ± 0.40 a | −24.90 | |||
| B2 | 297 | 0.88 ± 0.01 aD | 0.88 ± 0.01 aD | 0.98 ± 0.01 | 7.54 ± 0.29 aCD | −72.97 | −0.17 | −22.48 |
| 304 | 0.85 ± 0.02 b | 0.85 ± 0.02 b | 0.91 ± 0.05 | 3.40 ± 0.56 b | −21.29 | |||
| 311 | 0.82 ± 0.01 c | 0.82 ± 0.01 c | 0.87 ± 0.03 | 2.02 ± 0.24 c | −20.10 | |||
| B3 | 297 | 0.95 ± 0.02 aD | 0.95 ± 0.02 aD | 0.95 ± 0.04 | 5.59 ± 0.69 bD | 39.53 | 0.21 | −22.84 |
| 304 | 0.82 ± 0.01 b | 0.82 ± 0.01 b | 1.01 ± 0.02 | 9.19 ± 0.50 ab | −24.31 | |||
| 311 | 0.80 ± 0.01 b | 0.80 ± 0.01 b | 1.03 ± 0.02 | 11.35 ± 0.75 a | −25.78 | |||
| C1 | 297 | 1.93 ± 0.04 aA | 1.93 ± 0.04 aA | 0.92 ± 0.05 | 8.30 ± 1.49 bCD | 103.77 | 0.42 | −22.27 |
| 304 | 1.53 ± 0.02 b | 1.53 ± 0.02 b | 1.03 ± 0.03 | 22.07 ± 2.08 b | −25.24 | |||
| 311 | 1.47 ± 0.03 b | 1.47 ± 0.03 b | 1.13 ± 0.03 | 55.44 ± 6.38 a | −28.21 |
| T/K | Ksv (×104 L mol−1) | Kq (×1012 L mol−1 s−1) | n | Ka (×104 L mol−1) | ∆H (KJ mol−1) | ∆S (KJ mol−1 K−1) | ∆G (KJ mol−1) | |
|---|---|---|---|---|---|---|---|---|
| A1 | 297 | 2.14 ± 0.08 aC | 2.14 ± 0.08 aC | 1.19 ± 0.05 | 14.72 ± 2.36 aA | −112.12 | −0.28 | −29.13 |
| 304 | 1.78 ± 0.02 b | 1.78 ± 0.02 b | 1.08 ± 0.00 | 4.01 ± 0.06 b | −27.17 | |||
| 311 | 1.68 ± 0.05 c | 1.68 ± 0.05 c | 1.02 ± 0.04 | 1.90 ± 0.27 b | −25.22 | |||
| A2 | 297 | 1.61 ± 0.05 aD | 1.61 ± 0.05 aD | 0.79 ± 0.04 | 0.19 ± 0.03 bD | 41.89 | 0.20 | −18.56 |
| 304 | 1.58 ± 0.05 a | 1.58 ± 0.05 a | 0.82 ± 0.04 | 0.26 ± 0.4 b | −19.97 | |||
| 311 | 1.40 ± 0.03 b | 1.40 ± 0.03 b | 0.88 ± 0.04 | 0.41 ± 0.06 a | −21.40 | |||
| B1 | 297 | 2.52 ± 0.05 aA | 2.52 ± 0.05 aA | 1.07 ± 0.04 | 5.04 ± 0.71 bB | 67.90 | 0.32 | −26.67 |
| 304 | 2.46 ± 0.05 a | 2.46 ± 0.05 a | 1.13 ± 0.02 | 9.21 ± 0.63 ab | −28.90 | |||
| 311 | 2.15 ± 0.03 b | 2.15 ± 0.03 b | 1.19 ± 0.09 | 18.85 ± 5.88 a | −31.13 | |||
| B2 | 297 | 2.11 ± 0.05 aC | 2.11 ± 0.05 aC | 1.06 ± 0.06 | 4.13 ± 0.85 aBC | −32.94 | −0.02 | −26.12 |
| 304 | 1.77 ± 0.04 b | 1.77 ± 0.04 b | 1.05 ± 0.04 | 2.89 ± 0.35 ab | −25.96 | |||
| 311 | 1.57 ± 0.03 c | 1.57 ± 0.03 c | 1.03 ± 0.04 | 2.20 ± 0.27 b | −25.80 | |||
| B3 | 297 | 1.26 ± 0.03 aE | 1.26 ± 0.03 aE | 1.01 ± 0.04 | 1.46 ± 0.20 aCD | 13.61 | 0.13 | −23.61 |
| 304 | 1.22 ± 0.03 a | 1.22 ± 0.03 a | 1.03 ± 0.03 | 1.62 ± 0.20 a | −24.49 | |||
| 311 | 1.11 ± 0.02 b | 1.11 ± 0.02 b | 1.05 ± 0.03 | 1.86 ± 0.21 a | −25.37 | |||
| C1 | 297 | 2.33 ± 0.06 aB | 2.33 ± 0.06 aB | 1.07 ± 0.05 | 4.95 ± 0.87 aB | −29.10 | −0.01 | −26.60 |
| 304 | 2.09 ± 0.05 b | 2.09 ± 0.05 b | 1.05 ± 0.04 | 3.66 ± 0.49 a | −26.54 | |||
| 311 | 1.98 ± 0.03 b | 1.98 ± 0.03 b | 1.04 ± 0.04 | 2.89 ± 0.45 a | −26.48 |
| Samples | α-Helix% | β-Sheet% | β-Turn% | Coil% |
|---|---|---|---|---|
| β-LG | 30.77 | 23.79 | 32.18 | 13.26 |
| A1+β-LG | 37.93 | 19.02 | 26.60 | 16.45 |
| A2+β-LG | 32.50 | 24.27 | 25.62 | 17.61 |
| B1+β-LG | 37.44 | 28.85 | 13.53 | 20.18 |
| B2+β-LG | 38.65 | 27.47 | 13.64 | 20.23 |
| B3+β-LG | 29.64 | 23.31 | 33.68 | 13.37 |
| C1+β-LG | 37.59 | 28.83 | 13.79 | 19.78 |
| α-LA | 28.18 | 25.13 | 19.50 | 27.19 |
| A1+α-LA | 47.08 | 23.55 | 15.31 | 14.07 |
| A2+α-LA | 20.20 | 32.15 | 31.04 | 16.61 |
| B1+α-LA | 44.95 | 21.54 | 18.66 | 14.84 |
| B2+α-LA | 21.31 | 35.19 | 18.92 | 24.58 |
| B3+α-LA | 40.28 | 15.93 | 31.61 | 12.18 |
| C1+α-LA | 26.94 | 30.32 | 22.10 | 20.64 |
| Samples | α-Helix% | β-Sheet% | β-Turn% | Coil% |
|---|---|---|---|---|
| β-LG | 27.00 ± 2.42 c | 29.73 ± 6.80 a | 18.80 ± 2.75 ab | 24.67 ± 3.71 ab |
| A1+β-LG | 31.07 ± 1.85 bc | 15.37 ± 4.10 bc | 24.83 ± 2.49 a | 28.70 ± 0.89 a |
| A2+β-LG | 41.97 ± 2.90 a | 30.70 ± 7.63 a | 10.97 ± 2.56 c | 16.40 ± 3.40 c |
| B1+β-LG | 32.97 ± 1.37 bc | 26.60 ± 3.36 abc | 18.40 ± 1.89 b | 22.03 ± 1.13 bc |
| B2+β-LG | 37.13 ± 0.55 ab | 18.37 ± 0.52 abc | 21.77 ± 1.53 ab | 22.73 ± 0.73 ab |
| B3+β-LG | 36.07 ± 2.48 ab | 14.30 ± 5.05 c | 24.67 ± 1.35 a | 25.00 ± 1.51 ab |
| C1+β-LG | 28.67 ± 1.56 c | 27.03 ± 0.49 abc | 19.30 ± 0.75 ab | 25.03 ± 0.84 ab |
| α-LA | 27.60 ± 1.25 A | 23.83 ± 2.77 BC | 12.57 ± 0.67 A | 36.07 ± 1.44 B |
| A1+α-LA | 16.87 ± 1.24 B | 42.00 ± 3.05 A | 0.00 ± 0.00 C | 41.13 ± 1.87 A |
| A2+α-LA | 28.27 ± 1.50 A | 29.43 ± 3.25 B | 7.27 ± 2.09 B | 35.03 ± 0.92 B |
| B1+α-LA | 25.77 ± 0.79 A | 24.40 ± 2.38 BC | 13.87 ± 0.87 A | 36.00 ± 1.42 B |
| B2+α-LA | 25.83 ± 2.02 A | 29.53 ± 4.01 B | 8.47 ± 1.34 B | 36.20 ± 2.98 B |
| B3+α-LA | 25.63 ± 1.01 A | 21.80 ± 1.37 BC | 15.23 ± 0.24 A | 37.37 ± 0.67 AB |
| C1+α-LA | 17.00 ± 0.40 A | 20.17 ± 1.44 C | 15.30 ± 1.38 A | 37.57 ± 0.26 AB |
| Complex | Hydrogen Bonds | H-Bond Distance (Å) | Hydrophobic Residues | Other Residues | ΔG (kJ mol−1) | Intermolecular Energy (kJ mol−1) | Torsional Energy (kJ·mol−1) |
|---|---|---|---|---|---|---|---|
| A1+β-LG | Pro38, Lys69, Lys70, Glu74 *, Asp85 | 1.8–2.4 | Ala86, Ile71, Ile72 | - | −28.13 | −41.86 | 13.73 |
| A2+β-LG | Pro38, Lys69, Lys70, Glu74 *, Asp85 | 1.8–2.3 | Ala86, Ile71, Ile72 | - | −26.96 | −40.73 | 13.73 |
| B1+β-LG | Lys70, Asp85 # | 1.7–1.9 | Ile71, Ile72, Ile84 | Lys69 | −14.06 | −30.31 | 16.25 |
| B2+β-LG | Lys69, Glu74 | 1.9–2.3 | Ile71, Ile72, Ile84, Ala86, Met107 | Asp85, Asn90 | −19.46 | −35.71 | 16.25 |
| B3+β-LG | Ser36, Lys60, Lys69, Asn88, Asn90 | 1.8–2.2 | Leu31, Pro38, Leu39, Leu87, Met107, Leu117 | Glu108, Ser116 | −25.70 | −41.94 | 16.25 |
| C1+β-LG | Gln5, Thr6 | 2.0–2.7 | Val3, Ala139, Ala142, Leu143 | Thr4, Lys138, Lys141 | −15.57 | −40.56 | 25.01 |
| Complex | Hydrogen Bonds | H-Bond Distance (Å) | Hydrophobic Residues | Other Residues | ΔG (kJ·mol−1) | Intermolecular Energy (kJ·mol−1) | Torsional Energy (kJ·mol−1) |
|---|---|---|---|---|---|---|---|
| A1+α-LA | Ser112 *, Asp133 *, Trp118 | 1.8–3.0 | Pro109, Leu110 | His32, Lys108, Cys111, Gln117 | −24.91 | −38.64 | 13.73 |
| A2+α-LA | Ile33, Asp46 *, Tyr103, Trp104 | 1.8–2.1 | Val42 | Ser45, His47, Glu49, Gln54, Lys58, Lys108 | −20.89 | −34.62 | 13.73 |
| B1+α-LA | Lys5 *, Glu121 * | 1.7–2.0 | Phe31, Trp118 | Thr4, Cys6, Tyr36, Tyr119, Cys120 | −19.51 | −35.75 | 16.25 |
| B2+α-LA | Asn44, Ser45, Thr66, Gln68 # | 1.9–2.2 | Val67 | Lys43, Asp46, His47 | −19.17 | −35.41 | 16.25 |
| B3+α-LA | Tyr103, Asp59 | 1.8–2.1 | Ile33, Val42, Trp104, Ala106 | Asn44, Ser45, His47, Glu49, Gln54, Asn56, Lys58, Lys108 | −20.93 | −37.13 | 16.25 |
| C1+α-LA | Asp102 | 2.0 | Ile21, Ile101, Leu105, Pro109 | Glu25, His107, Lys108, Ser112 | −15.07 | −40.06 | 25.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, C.; Tan, B.; Sun, X. Elucidation of Interaction between Whey Proteins and Proanthocyanidins and Its Protective Effects on Proanthocyanidins during In-Vitro Digestion and Storage. Molecules 2021, 26, 5468. https://doi.org/10.3390/molecules26185468
Tang C, Tan B, Sun X. Elucidation of Interaction between Whey Proteins and Proanthocyanidins and Its Protective Effects on Proanthocyanidins during In-Vitro Digestion and Storage. Molecules. 2021; 26(18):5468. https://doi.org/10.3390/molecules26185468
Chicago/Turabian StyleTang, Chenyu, Bing Tan, and Xiangjun Sun. 2021. "Elucidation of Interaction between Whey Proteins and Proanthocyanidins and Its Protective Effects on Proanthocyanidins during In-Vitro Digestion and Storage" Molecules 26, no. 18: 5468. https://doi.org/10.3390/molecules26185468
APA StyleTang, C., Tan, B., & Sun, X. (2021). Elucidation of Interaction between Whey Proteins and Proanthocyanidins and Its Protective Effects on Proanthocyanidins during In-Vitro Digestion and Storage. Molecules, 26(18), 5468. https://doi.org/10.3390/molecules26185468