
Identification and Evaluation of Aromatic Volatile Compounds in 26 Cultivars and 8 Hybrids of Freesia hybrida


Abstract
:
1. Introduction
2. Results
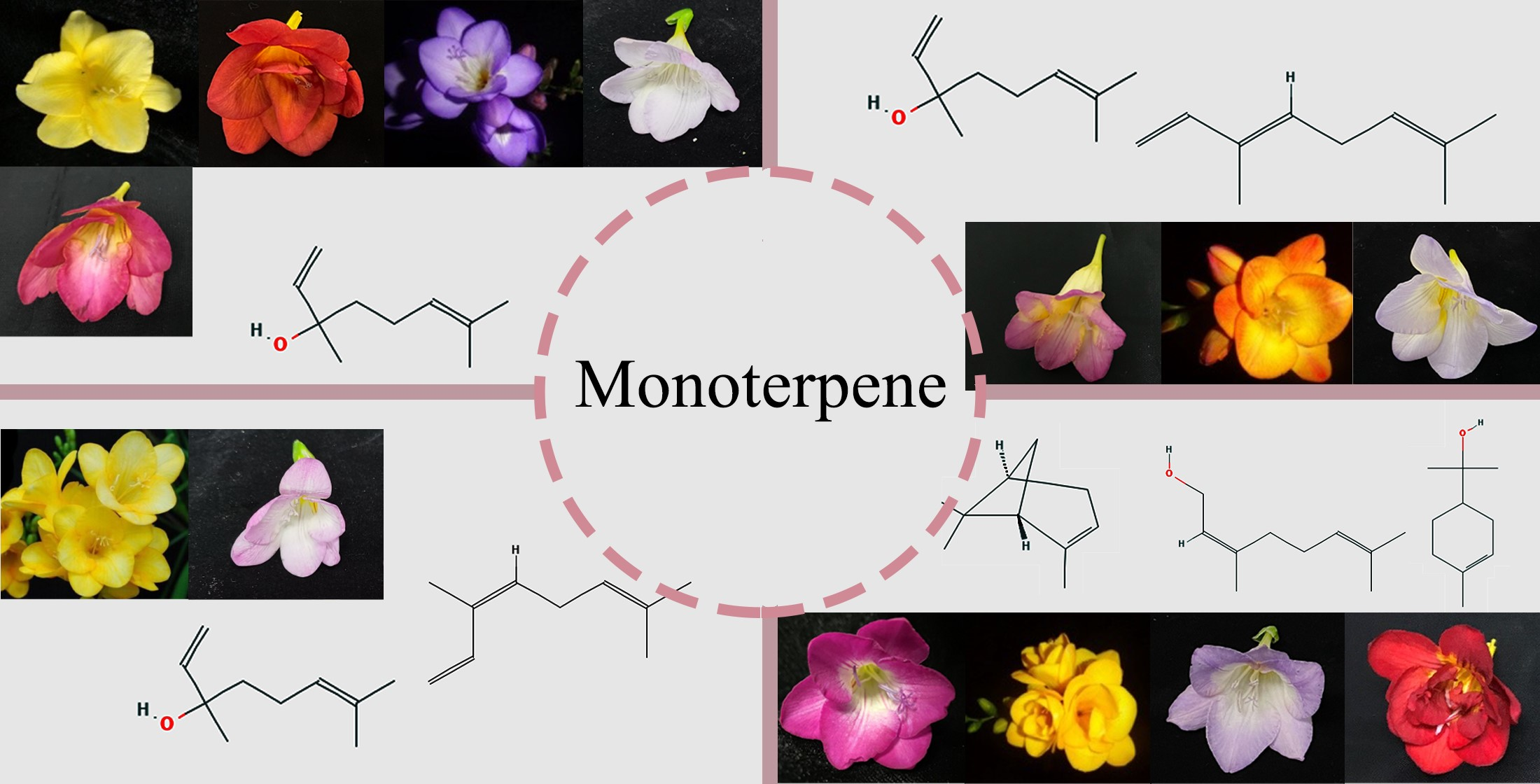
2.1. Comparative Analysis and Evaluation of VOCs
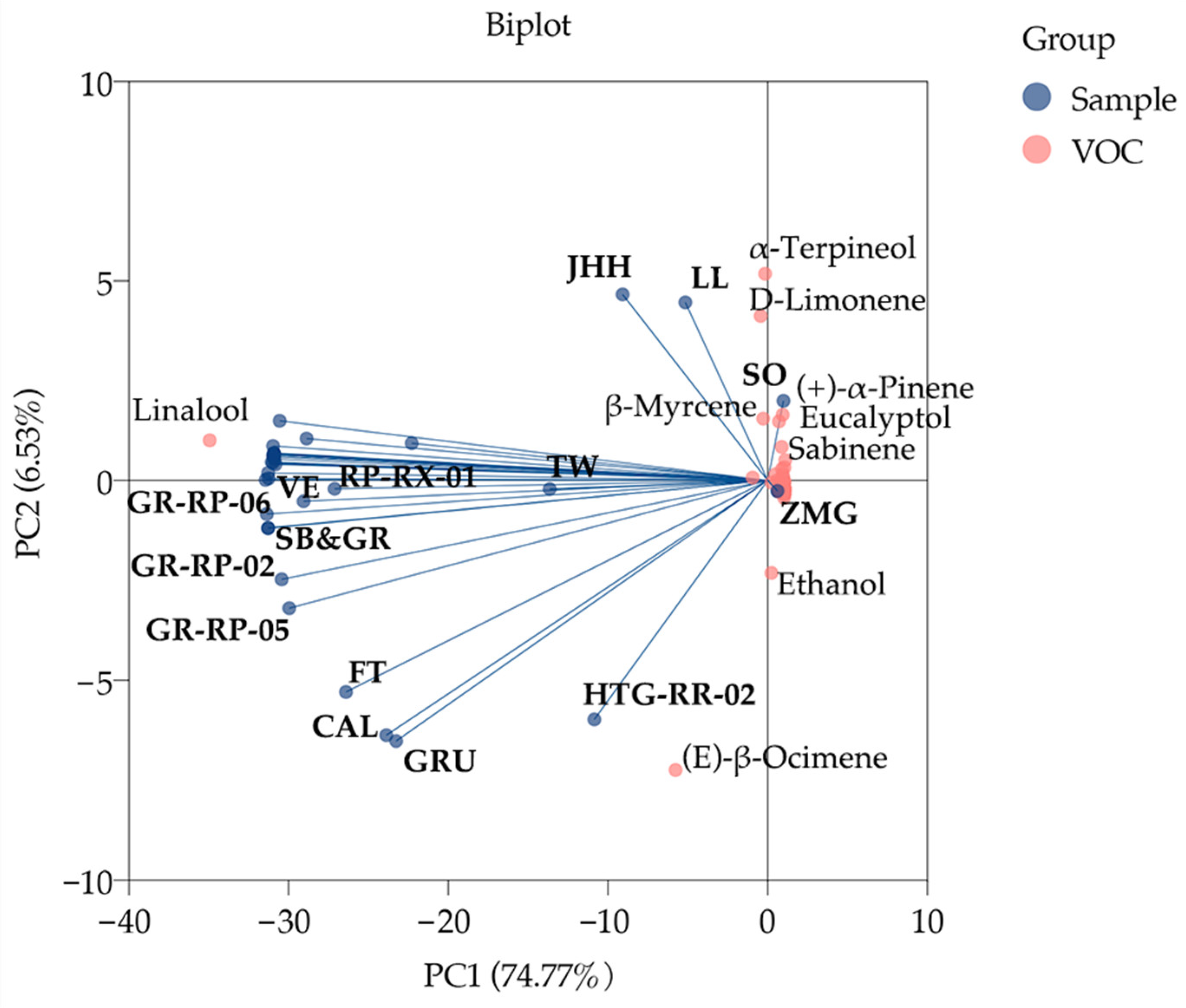
2.2. Pattern of the Floral Composition in 34 Germplasms
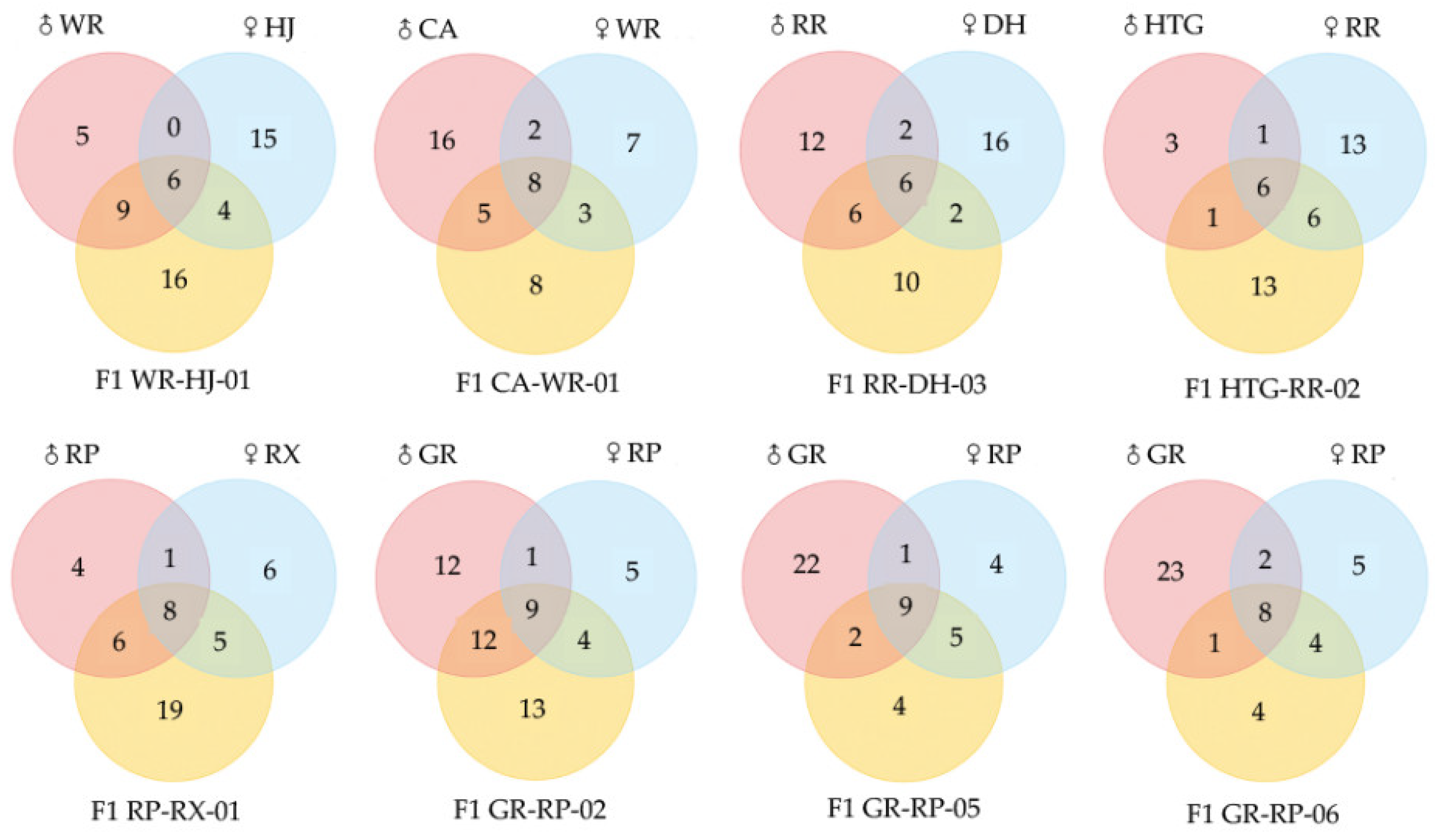
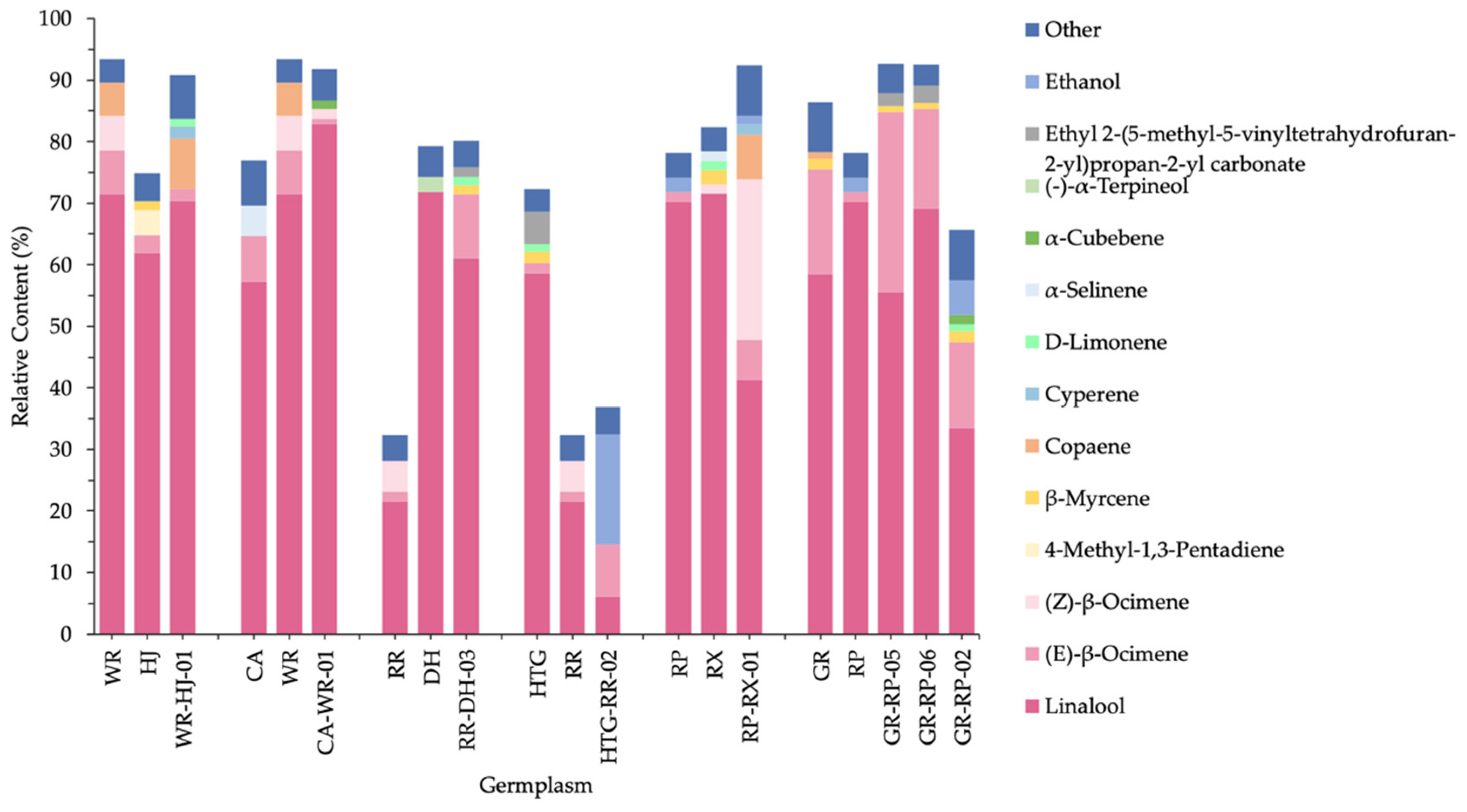
2.3. Comparison of the VOCs of Hybrids and Parental Species in F. hybrida
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. HS-SPME-GC-MS
4.3. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dudareva, N.; Negre, F.; Nagegowda, D.A.; Orlova, I. Plant volatiles: Recent advances and future perspectives. Crit. Rev. Plant Sci. 2006, 25, 417–440. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Gershenzon, J. The Chemical Diversity of Floral Scent. In Biology of Floral Scent, 2nd ed.; Pichersky, E., Dudareva, N., Eds.; CRC Press: Boca Raton, FL, USA, 2020; pp. 57–78. [Google Scholar] [CrossRef]
- Effah, E.; Holopainen, J.K.; McCormick, A.C. Potential roles of volatile organic compounds in plant competition. Perspect. Plant Ecol. Evol. Syst. 2019, 38, 58–63. [Google Scholar] [CrossRef]
- Pichersky, E.; Gershenzon, J. The formation and function of plant volatiles: Perfumes for pollinator attraction and defense. Curr. Opin. Plant Biol. 2002, 5, 237–243. [Google Scholar] [CrossRef]
- Kessler, D.; Gase, K.; Baldwin, I.T. Field experiments with transformed plants reveal the sense of floral scents. Science 2008, 321, 1200–1202. [Google Scholar] [CrossRef] [Green Version]
- Junker, R.R.; Blüthgen, N. Floral scents repel facultative flower visitors, but attract obligate ones. Ann. Bot. 2010, 105, 777–782. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, K.; Matsui, K.; Takabayashi, J. Uptake and Conversion of Volatile Compounds in Plant–Plant Communication. In Deciphering Chemical Language of Plant Communication, 1st ed.; Blande, J.D., Glinwood, R., Eds.; Springer: Cham, Switzerland, 2016; pp. 305–316. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; Coppola, R.; Feo, V.D. Essential oils and antifungal activity. Pharmaceuticals 2017, 10, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, A.A.; Pinho, D.B.; Cardeal, Z.L.; Menezes, H.C.; De Queiroz, M.V.; Pereira, O.L. Simplicillium coffeanum, a new endophytic species from Brazilian coffee plants, emitting antimicrobial volatiles. Phytotaxa 2018, 333, 188–198. [Google Scholar] [CrossRef]
- Vallarino, J.G.; Erban, A.; Fehrle, I.; Fernie, A.R.; Kopka, J.; Osorio, S. Acquisition of Volatile Compounds by Gas Chromatography–Mass Spectrometry (GC-MS). In Plant Metabolomics, 1st ed.; António, C., Ed.; Humana Press: New York, NY, USA, 2018; Volume 1778, pp. 225–239. [Google Scholar] [CrossRef]
- Bergougnoux, V.; Caissard, J.-C.; Jullien, F.; Magnard, J.-L.; Scalliet, G.; Cock, J.M.; Hugueney, P.; Baudino, S. Both the adaxial and abaxial epidermal layers of the rose petal emit volatile scent compounds. Planta 2007, 226, 853–866. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Yu, C.; Cheng, B.; Wan, H.; Luo, L.; Pan, H.; Zhang, Q. Volatile compound analysis and aroma evaluation of tea-scented roses in China. Ind. Crop. Prod. 2020, 155, 112735. [Google Scholar] [CrossRef]
- Kong, Y.; Bai, J.; Lang, L.; Bao, F.; Dou, X.; Wang, H.; Shang, H. Variation in floral scent compositions of different lily hybrid groups. J. Am. Soc. Hort. Sci. 2017, 142, 175–183. [Google Scholar] [CrossRef] [Green Version]
- Scalliet, G.; Piola, F.; Douady, C.J.; Réty, S.; Raymond, O.; Baudino, S.; Bordji, K.; Bendahmane, M.; Dumas, C.; Cock, J.M.; et al. Scent evolution in Chinese roses. Proc. Natl. Acad. Sci. USA 2008, 105, 5927–5932. [Google Scholar] [CrossRef] [Green Version]
- Wongchaochant, S.; Inamoto, K.; Doi, M. Analysis of flower scent of Freesia species and cultivars. Acta Hortic. 2005, 673, 595–601. [Google Scholar] [CrossRef]
- Wang, L. Freesia. In Flower Breeding and Genetics, 1st ed.; Anderson, N.O., Ed.; Springer: Dordrecht, The Netherlands, 2007; pp. 665–693. [Google Scholar] [CrossRef]
- Manning, J.C.; Goldblatt, P.; Duncan, G.D.; Forest, F.; Kaiser, R.; Tatarenko, I. Botany and Horticulture of the Genus Freesia (Iridaceae), 1st ed.; South African National Biodiversity Institute: Pretoria, South Africa, 2010; pp. 92–95. [Google Scholar]
- Harada, K.; Mihara, S. The Volatile constituents of Freesia flower (Freesia hybrida Hort.). Agric. Biol. Chem. 1984, 48, 2843–2845. [Google Scholar] [CrossRef]
- Yang, D.; Gao, X.; Wang, M.; Ao, M.; Wang, L. Extraction methods and analysis of the essential oil from Red Freesia hybrida. J. Northeast Norm. Univ. (Nat. Sci.) 2010, 42, 106–110, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Ao, M.; Liu, B.F.; Wang, L. Volatile compound in cut and un-cut flowers of tetraploid Freesia hybrida. Nat. Prod. Res. 2013, 27, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Gao, X.; Xue, Y.; Hui, Y.; Chen, F.; Su, Q.; Wang, L. Volatile Compounds in the Flowers of Freesia Parental Species and Hybrids. J. Integr. Plant Biol. 2007, 49, 1714–1718. [Google Scholar] [CrossRef]
- Stashenko, E.E.; Martínez, J.R. Sampling flower scent for chromatographic analysis. J. Sep. Sci. 2008, 31, 2022–2031. [Google Scholar] [CrossRef] [PubMed]
- Manzo, A.; Panseri, S.; Vagge, I.; Giorgi, A. Volatile Fingerprint of Italian Populations of Orchids Using Solid Phase Microextraction and Gas Chromatography Coupled with Mass Spectrometry. Molecules 2014, 19, 7913–7936. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Bao, F.; Yang, Y.; Hu, L.; Ding, A.; Ding, A.; Wang, J.; Cheng, T.; Zhang, Q. A Comparative Analysis of Floral Scent Compounds in Intraspecific Cultivars of Prunus mume with Different Corolla Colours. Molecules 2019, 25, 145. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Wang, H.; Wang, X.; Li, K.; Dong, M.; Li, Y.; Zhu, Q.; Shang, F. Mechanism of floral scent production in Osmanthus fragrans and the production and regulation of its key floral constituents, β-ionone and linalool. Hortic. Res. 2019, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lin, R.Y.; Zhong, H.Q.; Huang, M.L.; Luo, Y.H.; Lin, B. Aromatics in Flowers of Freesia hybrida. Fujian J. Agric. Sci. 2016, 31, 1216–1220, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Kishimoto, K. The Characters of Flower Scents in Carnations. In The Carnation Genome, 1st ed.; Onozaki, T., Yagi, M., Eds.; Springer: Singapore, 2020; pp. 147–157. [Google Scholar] [CrossRef]
- Oyama-Okubo, N.; Ando, T.; Watanabe, N.; Marchesi, E.; Uchida, K.; Nakayama, M. Emission mechanism of floral scent in Petunia axillaris. Biosci. Biotechnol. Biochem. 2014, 69, 773–777. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, S.M.; Kolosova, N.; Kish, C.M.; Wood, K.V.; Dudareva, N.; Jenks, M.A. Cuticle characters and volatile emissions of petals in Antirrhinum majus. Physiol. Plant. 2003, 117, 435–443. [Google Scholar] [CrossRef]
- Yue, Y.; Wang, L.; Yu, R.; Chen, F.; He, J.; Li, X.; Yu, Y.; Fan, Y. Coordinated and high-Level expression of biosynthetic pathway genes is responsible for the production of a major floral scent compound methyl benzoate in Hedychium coronarium. Front. Plant Sci. 2021, 12, 650582. [Google Scholar] [CrossRef]
- Liu, B.F.; Gao, F.Z.; Fang, Q.; Wang, L. Determination of red Freesia flower volatiles with indirect headspace solid phase microextraction coupled to gas chromatography and mass spectrometry. Chin. J. Anal. Chem. 2016, 44, 444–450, (In Chinese with English Abstract). [Google Scholar] [CrossRef]
- Aros, D.; Garrido, N.; Rivas, C.; Medel, M.; Müller, C.; Rogers, H.; Úbeda, C. Floral scent evaluation of three cut flowers through sensorial and gas chromatography analysis. Agronomy 2020, 10, 131. [Google Scholar] [CrossRef] [Green Version]
- Zarzo, M. Effect of functional group and carbon chain length on the odor detection threshold of aliphatic compounds. Sensors 2012, 12, 4105–4112. [Google Scholar] [CrossRef] [PubMed]
- Van Gemert, L.J. Compilations of Odour Threshold Values in Air, Water and Other Media, 2nd ed.; Oliemans Punter & Partners BV: Zeist, The Netherlands, 2011. [Google Scholar]
- Paulo, C.D.L.; Bittrich, V.; Shepherd, G.J.; Lopes, A.V.; Marsaioli, A.J. The ecological and taxonomic importance of flower volatiles of Clusia species (Guttiferae). Phytochemistry 2001, 56, 443–452. [Google Scholar] [CrossRef]
- Oyama-Okubo, N.; Tsuji, T. Analysis of floral scent compounds and classification by scent quality in tulip cultivars. J. Jpn. Soc. Hortic. Sci. 2013, 82, 344–353. [Google Scholar] [CrossRef] [Green Version]
- Angerosa, F.; Basti, C.; Vito, R. Virgin olive oil volatile compounds from lipoxygenase pathway and characterization of some italian cultivars. J. Agric. Food Chem. 1999, 47, 836–839. [Google Scholar] [CrossRef] [PubMed]
- Rjiba, I.; Debbou, S.; Gazzah, N.; Chreif, I.; Hammami, M. Profiles of volatile compounds from nine new hybrids obtained by controlled crossings on olive Chemlali cultivar and Mediterranean varieties. Nat. Prod. Res. 2009, 23, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, M.; Jürgens, A.; Campbell, D.R. Floral scent in natural hybrids of Ipomopsis (Polemoniaceae) and their parental species. Ann. Bot. 2014, 113, 533–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rieseberg, L.H.; Archer, M.A.; Wayne, R.K. Transgressive segregation, adaptation and speciation. Heredity 1999, 83, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Zu, P.; Blanckenhorn, W.U.; Schiestl, F.P. Heritability of floral volatiles and pleiotropic responses to artificial selection in Brassica rapa. New Phytol. 2015, 209, 1208–1219. [Google Scholar] [CrossRef] [PubMed]
- The Good Scents Company (tgsc) Information System. Available online: http://www.thegoodscentscompany.com/ (accessed on 21 June 2021).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Code | Germplasm Name | Source 1 | Sample Type |
|---|---|---|---|---|
| 1 | AN | ‘Ancona’ | a | Cultivar |
| 2 | CAL | ‘Calvados’ | a | Cultivar |
| 3 | SB | ‘Summer Beach’ | a | Cultivar |
| 4 | PP | ‘Pink Passion’ | a | Cultivar |
| 5 | VE | ‘Versailles’ | a | Cultivar |
| 6 | HTG | ‘SN 2 Hongtaige’ | b | Cultivar |
| 7 | RX | ‘SN Ruxiang’ | b | Cultivar |
| 8 | ZMG | ‘SN Zimeigui’ | b | Cultivar |
| 9 | SO | ‘Soleil’ | a | Cultivar |
| 10 | JHH | ‘SN Jinhuanghou’ | b | Cultivar |
| 11 | FS | ‘Fragrant Sunburst’ | a | Cultivar |
| 12 | LL | ‘Lovely Lavander’ | a | Cultivar |
| 13 | SN | ‘Snoozy’ | a | Cultivar |
| 14 | HJ | ‘SN Huangjin’ | b | Cultivar |
| 15 | GR | ‘Gold River’ | a | Cultivar |
| 16 | WR | ‘White River’ | a | Cultivar |
| 17 | RP | ‘Red Passion’ | a | Cultivar |
| 18 | CA | ‘Castor’ | a | Cultivar |
| 19 | MA | ‘Mandarine’ | a | Cultivar |
| 20 | GRU | ‘Grumpy’ | a | Cultivar |
| 21 | RR | ‘Red River’ | a | Cultivar |
| 22 | TW | ‘Tweety’ | a | Cultivar |
| 23 | CH | ‘SN Chenghuang’ | b | Cultivar |
| 24 | DXQ | ‘SN Danxueqing’ | b | Cultivar |
| 25 | DH | ‘SN Dahong’ | b | Cultivar |
| 26 | FT | ‘SN Feitao’ | b | Cultivar |
| 27 | WR-HJ-01 | ‘White River × SN Huangjin—01’ | b | Hybrid |
| 28 | RP-RX-01 | ‘Red Passion × SN Ruxiang—01’ | b | Hybrid |
| 29 | CA-WR-01 | ‘Castor × White River—01’ | b | Hybrid |
| 30 | HTG-RR-02 | ‘SN Hongtaige × Red River—01’ | b | Hybrid |
| 31 | RR-DH-03 | ‘Red River × SN Dahong—03’ | b | Hybrid |
| 32 | GR-RP-02 | ‘Gold River × Red Passion—02’ | b | Hybrid |
| 33 | GR-RP-05 | ‘Gold River × Red Passion—05’ | b | Hybrid |
| 34 | GR-RR-06 | ‘Gold River × Red Passion—06’ | b | Hybrid |
| No. | Name | Frequency |
|---|---|---|
| 1 | Linalool | 6 |
| 2 | β-Myrcene | 6 |
| 3 | d-Limonene | 6 |
| 4 | (E)-β-Ocimene | 6 |
| 5 | α-Terpineol | 4 |
| 6 | (+)-α-Pinene | 3 |
| 7 | Alloocimene | 3 |
| 8 | Terpinolene | 2 |
| 9 | (E)-Dehydroxylinalool Oxide | 2 |
| 10 | Rosefuran | 1 |
| 11 | (E)-Linalool Oxide (Pyranoid) | 1 |
| 12 | Ethanol | 1 |
| 13 | Cosmene | 1 |
| 14 | Dihydro-β-Ionone | 1 |
| 15 | β-Elemene | 1 |
| 16 | Selina-4(15),7(11)-Diene | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weng, S.; Fu, X.; Gao, Y.; Liu, T.; Sun, Y.; Tang, D. Identification and Evaluation of Aromatic Volatile Compounds in 26 Cultivars and 8 Hybrids of Freesia hybrida. Molecules 2021, 26, 4482. https://doi.org/10.3390/molecules26154482
Weng S, Fu X, Gao Y, Liu T, Sun Y, Tang D. Identification and Evaluation of Aromatic Volatile Compounds in 26 Cultivars and 8 Hybrids of Freesia hybrida. Molecules. 2021; 26(15):4482. https://doi.org/10.3390/molecules26154482
Chicago/Turabian StyleWeng, Shidan, Xueqing Fu, Yu Gao, Tianlei Liu, Yi Sun, and Dongqin Tang. 2021. "Identification and Evaluation of Aromatic Volatile Compounds in 26 Cultivars and 8 Hybrids of Freesia hybrida" Molecules 26, no. 15: 4482. https://doi.org/10.3390/molecules26154482
APA StyleWeng, S., Fu, X., Gao, Y., Liu, T., Sun, Y., & Tang, D. (2021). Identification and Evaluation of Aromatic Volatile Compounds in 26 Cultivars and 8 Hybrids of Freesia hybrida. Molecules, 26(15), 4482. https://doi.org/10.3390/molecules26154482