
Endoplasmic Reticulum Stress Provocation by Different Nanoparticles: An Innovative Approach to Manage the Cancer and Other Common Diseases

 ,
,  , ,
, , 
Abstract
1. Introduction
2. An Overview of Endoplasmic Reticulum (ER) Stress
3. Endoplasmic Reticulum Stress Mediated Diseases
4. Endoplasmic Reticulum Stress and Cancer
5. Endoplasmic Reticulum Stress as a Novel Target to Fight against Cancer Cells
6. Physico-Chemical Characteristics of Nanoparticles (NPs) and Their Role on Biological Systems
7. Nanoparticles Used as Toxicological and Therapeutic Agents Including ER Stress
7.1. Silver Nanoparticles
7.2. Gold Nanoparticles
7.3. Iron Oxide Nanoparticles
7.4. Manganese Nanoparticles
7.5. Titanium Oxide Nanoparticles
7.6. Zinc Oxide Nanoparticles
7.7. Quartz and Silica Nanoparticles
7.8. Graphene Oxide Nanoparticles
7.9. Lipid Nanoparticles
7.10. Enzyme Assembly Based Nanoparticles
7.11. Carbon Nanotubes
8. Summary
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ASK1 | apoptosis signal regulating kinase1 |
| ATF6 | activating transcription factor 6 |
| BiP | binding immunoglobulin protein |
| CHOP | C/EBP homologues protein |
| CNTs | carbon nanotubes |
| eIF2α | eukaryotic initiation factor 2α |
| EISA | enzyme instructed self-assembly |
| ER | Endoplasmic Reticulum |
| ERAD | endoplasmic reticulum associated protein degradation |
| GADD34 | growth arrest and DNA damage-inducible protein |
| GRP 78 | glucose regulated protein |
| GSH | glutathione |
| HSP | heat shock protein |
| HUVEC | human umbilical vein endothelial cells |
| IRE1α | inositol-requiring protein 1α |
| JNK | c-Jun N-terminal kinase |
| NPs | nanoparticles |
| OS | oxidative stress |
| PDI | protein disulfide isomerase |
| PEG | polyethylene glycol |
| PERK | protein kinase RNA-like ER kinase |
| ROS | reactive oxygen species |
| SOD 1 | superoxide dismutase 1 |
| SREBP 1c | sterol regulatory elementary binding protein 1c |
| TNFα | tumor necrosis factor α |
| TRAF2 | tumor necrosis factor-receptor associated factor 2 |
| UPR | unfolded protein response |
| Xbp1 | X-box binding protein 1 |
References
- Iurlaro, R.; Muñoz-Pinedo, C. Cell death induced by endoplasmic reticulum stress. FEBS J. 2016, 283, 2640–2652. [Google Scholar] [CrossRef] [PubMed]
- Braakman, I.; Hebert, D.N. Protein folding in the endoplasmic reticulum. Cold Spring Harb. Perspect. Biol. 2013, 5, a013201. [Google Scholar] [CrossRef] [PubMed]
- Luo, B.; Lee, A.S. The critical roles of endoplasmic reticulum chaperones and unfolded protein response in tumorigenesis and anticancer therapies. Oncogene 2013, 32, 805–818. [Google Scholar] [CrossRef] [PubMed]
- Lukas, J.; Pospech, J.; Oppermann, C.; Hund, C.; Iwanov, K.; Pantoom, S.; Petters, J.; Frech, M.; Seemann, S.; Thiel, F.G.; et al. Role of endoplasmic reticulum stress and protein misfolding in disorders of the liver and pancreas. Adv. Med. Sci. 2019, 64, 315–323. [Google Scholar] [CrossRef]
- Cao, S.S.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress in cell fate decision and human disease. Antioxid. Redox Signal. 2014, 21, 396–413. [Google Scholar] [CrossRef]
- Houck, S.A.; Ren, H.Y.; Madden, V.J.; Bonner, J.N.; Conlin, M.P.; Janovick, J.A.; Conn, P.M.; Cyr, D.M. Quality control autophagy degrades soluble ERAD-resistant conformers of the misfolded membrane protein GnRHR. Mol. Cell 2014, 54, 166–179. [Google Scholar] [CrossRef]
- Gessner, D.K.; Schlegel, G.; Ringseis, R.; Schwarz, F.J.; Eder, K. Up-regulation of endoplasmic reticulum stress induced genes of the unfolded protein response in the liver of periparturient dairy cows. BMC Vet. Res. 2014, 10, 46. [Google Scholar] [CrossRef]
- Malhotra, J.D.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress: A vicious cycle or a double-edged sword? Antioxid. Redox Signal. 2007, 9, 2277–2293. [Google Scholar] [CrossRef]
- Tse, G.; Yan, B.P.; Chan, Y.W.; Tian, X.Y.; Huang, Y. Reactive oxygen species, endoplasmic reticulum stress and mitochondrial dysfunction: The link with cardiac arrhythmogenesis. Front. Physiol. 2016, 7, 313. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Endoplasmic reticulum stress and the inflammatory basis of metabolic disease. Cell 2010, 140, 900–917. [Google Scholar] [CrossRef]
- Liu, Z.W.; Zhu, H.T.; Chen, K.L.; Dong, X.; Wei, J.; Qiu, C.; Xue, J.H. Protein kinase RNA-like endoplasmic reticulum kinase (PERK) signaling pathway plays a major role in reactive oxygen species (ROS)-mediated endoplasmic reticulum stress-induced apoptosis in diabetic cardio-myopathy. Cardiovasc. Diabetol. 2013, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Korennykh, A.V.; Behrman, S.L.; Walter, P. Mammalian endoplasmic reticulum stress sensor IRE1 signals by dynamic clustering. Proc. Natl. Acad. Sci. USA 2010, 107, 16113–16118. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Lerner, A.G.; Walle, L.V.; Upton, J.P.; Xu, W.; Hagen, A.; Backes, B.J.; Oakes, S.A.; Papa, F.R. IRE1α kinase activation modes control alternate endoribonuclease outputs to determine divergent cell fates. Cell 2009, 138, 562–575. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.H.; Iwakoshi, N.N.; Glimcher, L.H. XBP-1 regulates a subset of endoplasmic reticulum resident chaperone genes in the unfolded protein response. Mol. Cell. Biol. 2003, 23, 7448–7459. [Google Scholar] [CrossRef] [PubMed]
- Shaffer, A.; Shapiro-Shelef, M.; Iwakoshi, N.N.; Lee, A.-H.; Qian, S.-B.; Zhao, H.; Yu, X.; Yang, L.; Tan, B.K.; Rosenwald, A.; et al. XBP1, Downstream of Blimp-1, Expands the Secretory Apparatus and Other Organelles, and Increases Protein Synthesis in Plasma Cell Differentiation. Immunity 2004, 21, 81–93. [Google Scholar] [CrossRef]
- Nishitoh, H.; Matsuzawa, A.; Tobiume, K.; Saegusa, K.; Takeda, K.; Inoue, K.; Hori, S.; Kakizuka, A.; Ichijo, H. ASK1 is essential for endoplasmic reticulum stress-induced neuronal cell death triggered by expanded polyglutamine repeats. Genes Dev. 2002, 16, 1345–1355. [Google Scholar] [CrossRef]
- Barr, R.K.; Bogoyevitch, M.A. The c-Jun N-terminal protein kinase family of mitogen-activated protein kinases (JNK MAPKs). Int. J. Biochem. Cell Biol. 2001, 33, 1047–1063. [Google Scholar] [CrossRef]
- Tong, Q.; Wu, L.; Jiang, T.; Ou, Z.; Zhang, Y.; Zhu, D. Inhibition of endoplasmic reticulum stress-activated IRE1α-TRAF2-caspase-12 apoptotic pathway is involved in the neuroprotective effects of telmisartan in the rotenone rat model of Parkinson’s disease. Eur. J. Pharmacol. 2016, 776, 106–115. [Google Scholar] [CrossRef]
- Wang, P.; Li, J.; Tao, J.; Sha, B. The luminal domain of the ER stress sensor protein PERK binds misfolded proteins and thereby triggers PERK oligomerization. J. Biol. Chem. 2018, 293, 4110–4121. [Google Scholar] [CrossRef]
- Anderson, P.; Kedersha, N. Visibly stressed: The role of eIF2, TIA-1, and stress granules in protein translation. Cell Stress Chaperones 2002, 7, 213–221. [Google Scholar] [CrossRef]
- McQuiston, A.; Diehl, J.A. Recent insights into PERK-dependent signaling from the stressed endoplasmic reticulum. F1000Research 2017, 6, 1897. [Google Scholar] [CrossRef] [PubMed]
- Harding, H.P.; Novoa, I.; Zhang, Y.; Zeng, H.; Wek, R.; Schapira, M.; Ron, D. Regulated Translation Initiation Controls Stress-Induced Gene Expression in Mammalian Cells. Mol. Cell 2000, 6, 1099–1108. [Google Scholar] [CrossRef]
- Liao, Y.; Gu, F.; Mao, X.; Niu, Q.; Wang, H.; Sun, Y.; Song, C.; Qiu, X.; Tan, L.; Ding, C. Regulation of de novo translation of host cells by manipulation of PERK/PKR and GADD34-PP1 activity during Newcastle disease virus infection. J. Gen. Virol. 2016, 97, 867–879. [Google Scholar] [CrossRef]
- Horimoto, S.; Ninagawa, S.; Okada, T.; Koba, H.; Sugimoto, T.; Kamiya, Y.; Kato, K.; Takeda, S.; Mori, K. The unfolded protein response transducer ATF6 represents a novel transmembrane-type endoplasmic reticulum-associated degradation substrate requiring both mannose trimming and SEL1L protein. J. Biol. Chem. 2013, 288, 31517–31527. [Google Scholar] [CrossRef] [PubMed]
- Stirling, J.; O’Hare, P. CREB4, a transmembrane bZip transcription factor and potential new substrate for regulation and cleavage by S1P. Mol. Biol. Cell 2006, 17, 413–426. [Google Scholar] [CrossRef]
- Chen, Y.; Brandizzi, F. IRE1: ER stress sensor and cell fate executor. Trends Cell Biol. 2013, 23, 547–555. [Google Scholar] [CrossRef]
- Chen, X.; Shen, J.; Prywes, R. The luminal domain of ATF6 senses endoplasmic reticulum (ER) stress and causes translocation of ATF6 from the ER to the Golgi. J. Biol. Chem. 2002, 277, 13045–13052. [Google Scholar] [CrossRef]
- Ye, J.; Rawson, R.B.; Komuro, R.; Chen, X.; Davé, U.P.; Prywes, R.; Brown, M.S.; Goldstein, J.L. ER Stress Induces Cleavage of Membrane-Bound ATF6 by the Same Proteases that Process SREBPs. Mol. Cell 2000, 6, 1355–1364. [Google Scholar] [CrossRef]
- Teske, B.F.; Wek, S.A.; Bunpo, P.; Cundiff, J.K.; McClintick, J.N.; Anthony, T.G.; Wek, R.C. The eIF2 kinase PERK and the integrated stress response facilitate activation of ATF6 during endoplasmic reticulum stress. Mol. Biol. Cell 2011, 22, 4390–4405. [Google Scholar] [CrossRef]
- Nakanishi, K.; Sudo, T.; Morishima, N. Endoplasmic reticulum stress signaling transmitted by ATF6 mediates apoptosis during muscle development. J. Cell Biol. 2005, 169, 555–560. [Google Scholar] [CrossRef]
- Ozcan, L.; Tabas, I. Role of endoplasmic reticulum stress in metabolic disease and other disorders. Annu. Rev. Med. 2012, 18, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.; Choi, S.K.; Galan, M.; Kassan, M.; Partyka, M.; Kadowitz, P.; Henrion, D.; Trebak, M.; Belmadani, S.; Matrougui, K. Chronic inhibition of endoplasmic reticulum stress and inflammation prevents ischaemia-induced vascular pathology in type II diabetic mice. J. Pathol. 2012, 227, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Imai, Y.; Soda, M.; Takahashi, R. Parkin suppresses unfolded protein stress-induced cell death through its E3 ubiquitin-protein ligase activity. J. Biol. Chem. 2000, 275, 35661–35664. [Google Scholar] [CrossRef]
- Nijholt, D.A.; de Graaf, T.R.; van Haastert, E.S.; Oliveira, A.O.; Berkers, C.R.; Zwart, R.; Ovaa, H.; Baas, F.; Hoozemans, J.J.; Scheper, W. Endoplasmic reticulum stress activates autophagy but not the proteasome in neuronal cells: Implications for Alzheimer’s disease. Cell Death Differ. 2011, 18, 1071–1081. [Google Scholar] [CrossRef] [PubMed]
- Tsukano, H.; Gotoh, T.; Endo, M.; Miyata, K.; Tazume, H.; Kadomatsu, T.; Yano, M.; Iwawaki, T.; Kohno, K.; Araki, K.; et al. The Endoplasmic Reticulum Stress-C/EBP Homologous Protein Pathway-Mediated Apoptosis in Macrophages Contributes to the Instability of Atherosclerotic Plaques. Arter. Thromb. Vasc. Biol. 2010, 30, 1925–1932. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, L.; Zhang, X.; Xu, L.; Xie, B.; Shi, H.; Duan, Z.; Zhang, H.; Ren, F. Kaempferol protects mice from d-GalN/LPS-induced acute liver failure by regulating the ER stress-Grp78-CHOP signaling pathway. Biomed. Pharmacother. 2019, 111, 468–475. [Google Scholar] [CrossRef]
- Kammoun, H.L.; Chabanon, H.; Hainault, I.; Luquet, S.; Magnan, C.; Koike, T.; Ferré, P.; Foufelle, F. GRP78 expression inhibits insulin and ER stress-induced SREBP-1c activation and reduces hepatic steatosis in mice. J. Clin. Investig. 2009, 119, 1201–1215. [Google Scholar] [CrossRef]
- Yeganeh, B.; Moghadam, A.R.; Alizadeh, J.; Wiechec, E.; Alavian, S.M.; Hashemi, M.; Geramizadeh, B.; Samali, A.; Lankarani, K.B.; Post, M.; et al. Hepatitis B and C virus-induced hepatitis: Apoptosis, autophagy, and unfolded protein response. World J. Gastroenterol. 2015, 21, 13225. [Google Scholar] [CrossRef]
- Saxena, S.; Cabuy, E.; Caroni, P. A role for motoneuron subtype–selective ER stress in disease manifestations of FALS mice. Nat. Neurosci. 2009, 12, 627–636. [Google Scholar] [CrossRef]
- So, A.Y.-L.; De La Fuente-Ortega, E.; Walter, P.; Shuman, M.; Bernales, S. The unfolded protein response during prostate cancer development. Cancer Metastasis Rev. 2009, 28, 219–223. [Google Scholar] [CrossRef]
- Delie, F.; Petignat, P.; Cohen, M. GRP78 Protein Expression in Ovarian Cancer Patients and Perspectives for a Drug-Targeting Approach. J. Oncol. 2012, 2012, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Su, R.; Li, Z.; Li, H.; Song, H.; Bao, C.; Wei, J.; Cheng, L. Grp78 promotes the invasion of hepatocellular carcinoma. BMC Cancer 2010, 10, 20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Tseng, C.-C.; Tsai, Y.-L.; Fu, X.; Schiff, R.; Lee, A.S. Cancer Cells Resistant to Therapy Promote Cell Surface Relocalization of GRP78 Which Complexes with PI3K and Enhances PI(3,4,5)P3 Production. PLoS ONE 2013, 8, e80071. [Google Scholar] [CrossRef]
- Overley-Adamson, B.; Artlett, C.M.; Stephens, C.; Sassi-Gaha, S.; Weis, R.D.; Thacker, J.D. Targeting the unfolded protein response, XBP1, and the NLRP3 inflammasome in fibrosis and cancer. Cancer Biol. Ther. 2014, 15, 452–462. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Déry, M.-A.; Jodoin, J.; Ursini-Siegel, J.; Aleynikova, O.; Ferrario, C.; Hassan, S.; Basik, M.; Leblanc, A.C. Endoplasmic reticulum stress induces PRNP prion protein gene expression in breast cancer. Breast Cancer Res. 2013, 15, R22. [Google Scholar] [CrossRef] [PubMed]
- Nagelkerke, A.; Bussink, J.; Mujcic, H.; Wouters, B.G.; Lehmann, S.; Sweep, F.C.; Span, N.P. Hypoxia stimulates migration of breast cancer cells via the PERK/ATF4/LAMP3-arm of the unfolded protein response. Breast Cancer Res 2013, 15, R2. [Google Scholar] [CrossRef]
- Ming, J.; Ruan, S.; Wang, M.; Ye, D.; Fan, N.; Meng, Q.; Tian, B.; Huang, T. A novel chemical, STF-083010, reverses tamoxifen-related drug resistance in breast cancer by inhibiting IRE1/XBP1. Oncotarget 2015, 6, 40692. [Google Scholar] [CrossRef]
- Pike, L.R.G.; Singleton, D.C.; Buffa, F.M.; Abramczyk, O.; Phadwal, K.; Li, J.; Simon, A.K.; Murray, J.T.; Harris, A.L. Transcriptional up-regulation of ULK1 by ATF4 contributes to cancer cell survival. Biochem. J. 2012, 449, 389–400. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, L.; Zhao, Y.; Li, H.; Xiao, H.; Fu, R.; Zhao, C.; Wu, H.; Li, Z. Cell-surface GRP78 facilitates colorectal cancer cell migration and invasion. Int. J. Biochem. Cell Biol. 2013, 45, 987–994. [Google Scholar] [CrossRef]
- Lee, H.K.; Xiang, C.; Cazacu, S.; Finniss, S.; Kazimirsky, G.; Lemke, N.; Lehman, N.L.; Rempel, S.A.; Mikkelsen, T.; Brodie, C. GRP78 is overexpressed in glioblastomas and regulates glioma cell growth and apoptosis. Neuro-Oncology 2008, 10, 236–243. [Google Scholar] [CrossRef]
- Mahoney, D.J.; Lefebvre, C.; Allan, K.; Brun, J.; Sanaei, C.A.; Baird, S.; Pearce, N.; Grönberg, S.; Wilson, B.; Prakesh, M.; et al. Virus-Tumor Interactome Screen Reveals ER Stress Response Can Reprogram Resistant Cancers for Oncolytic Virus-Triggered Caspase-2 Cell Death. Cancer Cell 2011, 20, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; McGrath, B.; Cavener, D.R. PERK regulates the proliferation and development of insulin-secreting beta-cell tumors in the endocrine pancreas of mice. PLoS ONE 2009, 4, e8008. [Google Scholar] [CrossRef]
- Niu, Z.; Wang, M.; Zhou, L.; Yao, L.; Liao, Q.; Zhao, Y. Elevated GRP78 expression is associated with poor prognosis in patients with pancreatic cancer. Sci. Rep. 2015, 5, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.S. GRP78 Induction in Cancer: Therapeutic and Prognostic Implications. Cancer Res. 2007, 67, 3496–3499. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Cheng, D.; Zhou, S.-M.; Zhu, B.; Hu, T.; Yang, Q.-C. Overexpression of X-Box Binding Protein 1 (XBP1) Correlates to Poor Prognosis and Up-Regulation of PI3K/mTOR in Human Osteosarcoma. Int. J. Mol. Sci. 2015, 16, 28635–28646. [Google Scholar] [CrossRef]
- Kraskiewicz, H.; Fitzgerald, U. InterfERing with endoplasmic reticulum stress. Trends Pharmacol. Sci. 2012, 33, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Schleicher, S.M.; Moretti, L.; Varki, V.; Lu, B. Progress in the unraveling of the endoplasmic reticulum stress/autophagy pathway and cancer: Implications for future therapeutic approaches. Drug Resist. Update 2010, 13, 79–86. [Google Scholar] [CrossRef]
- Healy, S.J.; Gorman, A.M.; Mousavi-Shafaei, P.; Gupta, S.; Samali, A. Targeting the endoplasmic reticulum-stress response as an anticancer strategy. Eur. J. Pharmacol. 2009, 625, 234–246. [Google Scholar] [CrossRef]
- Cho, H.-Y.; Thomas, S.; Golden, E.B.; Gaffney, K.J.; Hofman, F.M.; Chen, T.C.; Louie, S.G.; Petasis, N.A.; Schönthal, A.H. Enhanced killing of chemo-resistant breast cancer cells via controlled aggravation of ER stress. Cancer Lett. 2009, 282, 87–97. [Google Scholar] [CrossRef]
- Shen, K.; Johnson, D.W.; Vesey, D.A.; McGuckin, M.A.; Gobe, G.C. Role of the unfolded protein response in determining the fate of tumor cells and the promise of multi-targeted therapies. Cell Stress Chaperones 2018, 23, 317–334. [Google Scholar] [CrossRef]
- Seo, S.; Richter, A.; Blechschmidt, A.-M.; Bougoudis, I.; Burrows, J.P. First high resolution BrO column retrievals from TROPOMI. J. Cancer 2018, 2018, 1–26. [Google Scholar] [CrossRef]
- Suh, D.H.; Kim, M.K.; Kim, H.S.; Chung, H.H.; Song, Y.S. Unfolded protein response to autophagy as a promising druggable target for anticancer therapy. Ann. N. Y. Acad. Sci. 2012, 1271, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Martelli, A.M.; Paganelli, F.; Chiarini, F.; Evangelisti, C.; McCubrey, J.A. The Unfolded Protein Response: A Novel Therapeutic Target in Acute Leukemias. Cancers 2020, 12, 333. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Nie, G.; Meng, H.; Xia, T.; Nel, A.; Zhao, Y. Physicochemical Properties Determine Nanomaterial Cellular Uptake, Transport, and Fate. Acc. Chem. Res. 2013, 46, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Stenzel, M.H. Entry of nanoparticles into cells: The importance of nanoparticle properties. Polym. Chem. 2018, 9, 259–272. [Google Scholar] [CrossRef]
- Jiang, W.; Kim, B.Y.S.; Rutka, J.T.; Chan, W.C.W. Nanoparticle-mediated cellular response is size-dependent. Nat. Nanotechnol. 2008, 3, 145–150. [Google Scholar] [CrossRef]
- Bartczak, D.; Louafi, F.; Millar, T.M.; Kanaras, A.G.; Sanchez-Elsner, T. Receptor-Mediated Interactions between Colloidal Gold Nanoparticles and Human Umbilical Vein Endothelial Cells. Small 2010, 7, 388–394. [Google Scholar] [CrossRef]
- Muro, S.; Garnacho, C.; Champion, J.A.; Leferovich, J.; Gajewski, C.; Schuchman, E.H.; Mitragotri, S.; Muzykantov, V.R. Control of Endothelial Targeting and Intracellular Delivery of Therapeutic Enzymes by Modulating the Size and Shape of ICAM-1-targeted Carriers. Mol. Ther. 2008, 16, 1450–1458. [Google Scholar] [CrossRef]
- Unfried, K.; Sydlik, U.; Bierhals, K.; Weissenberg, A.; Abel, J. Carbon nanoparticle-induced lung epithelial cell proliferation is mediated by receptor-dependent Akt activation. Am. J. Physiol. Cell. Mol. Physiol. 2008, 294, L358–L367. [Google Scholar] [CrossRef]
- Zhang, D.; Neumann, O.; Wang, H.; Yuwono, V.M.; Barhoumi, A.; Perham, M.; Hartgerink, J.D.; Wittung-Stafshede, P.; Halas, N.J. Gold Nanoparticles Can Induce the Formation of Protein-based Aggregates at Physiological pH. Nano Lett. 2009, 9, 666–671. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.J.; Liang, M.; Monteiro, M.; Toth, I.; Minchin, R.F. Nanoparticle-induced unfolding of fibrinogen promotes Mac-1 receptor activation and inflammation. Nat. Nanotechnol. 2011, 6, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Christen, V.; Capelle, M.; Fent, K. Silver nanoparticles induce endoplasmatic reticulum stress response in zebrafish. Toxicol. Appl. Pharmacol. 2013, 272, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Piao, M.J.; Kim, K.C.; Kim, A.D.; Choi, J.-Y.; Choi, J.; Hyun, J.W. Endoplasmic reticulum stress signaling is involved in silver nanoparticles-induced apoptosis. Int. J. Biochem. Cell Biol. 2012, 44, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, A.; Das, J.; Manna, P.; Sil, P.C. Nano-copper induces oxidative stress and apoptosis in kidney via both extrinsic and intrinsic pathways. Toxicology 2011, 290, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.A.; Alhadlaq, H.A.; Ahmad, J.; Al-Khedhairy, A.A.; Musarrat, J.; Ahamed, M. Copper Oxide Nanoparticles Induced Mitochondria Mediated Apoptosis in Human Hepatocarcinoma Cells. PLoS ONE 2013, 8, e69534. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.-Y.; Huang, Y.-H.; Chao, Y.-L.; Hu, K.-Y.; Chin, L.-T.; Chou, S.-H.; Hour, A.-L.; Yao, Y.-D.; Tu, C.-S.; Liang, Y.-J.; et al. Identification of the Nanogold Particle-Induced Endoplasmic Reticulum Stress by Omic Techniques and Systems Biology Analysis. ACS Nano 2011, 5, 9354–9369. [Google Scholar] [CrossRef]
- Xia, T.; Zhao, Y.; Sager, T.; George, S.; Pokhrel, S.; Li, N.; Schoenfeld, D.; Meng, H.; Lin, S.; Wang, X.; et al. Decreased Dissolution of ZnO by Iron Doping Yields Nanoparticles with Reduced Toxicity in the Rodent Lung and Zebrafish Embryos. ACS Nano 2011, 5, 1223–1235. [Google Scholar] [CrossRef]
- Kim, C.K.; Kim, T.; Choi, I.; Soh, M.; Kim, D.; Kim, Y.-J.; Jang, H.; Yang, H.-S.; Kim, J.Y.; Park, H.-K.; et al. Ceria Nanoparticles that can Protect against Ischemic Stroke. Angew. Chem. Int. Ed. 2012, 51, 11039–11043. [Google Scholar] [CrossRef]
- Conde, J.; Doria, G.; Baptista, P. Noble metal nanoparticles applications in cancer. J. Drug Deliv. 2012, 2012. [Google Scholar] [CrossRef]
- Khan, A.A.; Allemailem, K.S.; Almatroodi, S.A.; Almatroudi, A.; Rahmani, A.H. Recent strategies towards the surface modification of liposomes: An innovative approach for different clinical applications. 3 Biotech 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Dabbaghi, M.; Oskuee, R.K.; Hashemi, K.; Goli, A.A. Evaluating polyethyleneimine/DNA nanoparticles-mediated damage to cellular organelles using endoplasmic reticulum stress profile. Artif. Cells Nanomed. Biotechnol. 2017, 46, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Shen, Y.; Li, X.; Jiang, Q.; Cheng, S.; Gu, Y.; Liu, L.; Cao, Y. The endoplasmic reticulum stress inducer thapsigargin enhances the toxicity of ZnO nanoparticles to macrophages and macrophage-endothelial co-culture. Environ. Toxicol. Pharmacol. 2017, 50, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Erbis, S.; Ok, Z.; Isaacs, J.A.; Benneyan, J.C.; Kamarthi, S. Review of Research Trends and Methods in Nano Environmental, Health, and Safety Risk Analysis. Risk Anal. 2016, 36, 1644–1665. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Cheng, S.; Chen, G.; Shen, Y.; Li, X.; Jiang, Q.; Li, J.; Cao, Y. The effects of endoplasmic reticulum stress inducer thapsigargin on the toxicity of ZnO or TiO2 nanoparticles to human endothelial cells. Toxicol. Mech. Methods 2017, 27, 191–200. [Google Scholar] [CrossRef]
- Hirsch, I.; Weiwad, M.; Prell, E.; Ferrari, D.M. ERp29 deficiency affects sensitivity to apoptosis via impairment of the ATF6–CHOP pathway of stress response. Apoptosis 2013, 19, 801–815. [Google Scholar] [CrossRef]
- Hoseinzadeh, E.; Makhdoumi, P.; Taha, P.; Hossini, H.; Stelling, J.; Kamal, M.A.; Ashraf, G.M. A Review on Nano-Antimicrobials: Metal Nanoparticles, Methods and Mechanisms. Curr. Drug Metab. 2017, 18, 120–128. [Google Scholar] [CrossRef]
- Ghosh, C.; Nandi, A.; Basu, S. Supramolecular self-assembly of triazine-based small molecules: Targeting the endoplasmic reticulum in cancer cells. Nanoscale 2019, 11, 3326–3335. [Google Scholar] [CrossRef]
- Sydlik, S.A.; Jhunjhunwala, S.; Webber, M.J.; Anderson, D.G.; Langer, R. In Vivo Compatibility of Graphene Oxide with Differing Oxidation States. ACS Nano 2015, 9, 3866–3874. [Google Scholar] [CrossRef]
- Mallick, A.; Nandi, A.; Basu, S. Polyethylenimine coated graphene oxide nanoparticles for targeting mitochondria in cancer cells. ACS Appl. Bio Mater. 2019, 2, 14–19. [Google Scholar] [CrossRef]
- Chen, L.; Yokel, R.A.; Hennig, B.; Toborek, M. Manufactured Aluminum Oxide Nanoparticles Decrease Expression of Tight Junction Proteins in Brain Vasculature. J. Neuroimmune Pharmacol. 2008, 3, 286–295. [Google Scholar] [CrossRef]
- Jalal, M.; Ansari, M.A.; Shukla, A.K.; Ali, S.G.; Khan, H.M.; Pal, R.; Alam, J.; Cameotra, S.S. Green synthesis and antifungal activity of Al2O3NPs against fluconazole-resistant Candida spp isolated from a tertiary care hospital. RSC Adv. 2016, 6, 107577–107590. [Google Scholar] [CrossRef]
- Radziun, E.; Wilczyńska, J.D.; Książek, I.; Nowak, K.; Anuszewska, E.; Kunicki, A.; Olszyna, A.R.; Ząbkowski, T. Assessment of the cytotoxicity of aluminium oxide nanoparticles on selected mammalian cells. Toxicol. In Vitro 2011, 25, 1694–1700. [Google Scholar] [CrossRef]
- Ansari, M.A.; Khan, H.M.; Alzohairy, M.A.; Jalal, M.; Ali, S.G.; Pal, R.; Musarrat, J. Green synthesis of Al2O3 nanoparticles and their bactericidal potential against clinical isolates of multi-drug resistant Pseudomonas aeruginosa. World J. Microbiol. Biotechnol. 2014, 31, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Alshatwi, A.A.; Subbarayan, P.V.; Ramesh, E.; Al-Hazzani, A.A.; AlSaif, M.A.; Alwarthan, A.A. Al2O3Nanoparticles Induce Mitochondria-Mediated Cell Death and Upregulate the Expression of Signaling Genes in Human Mesenchymal Stem Cells. J. Biochem. Mol. Toxicol. 2012, 26, 469–476. [Google Scholar] [CrossRef]
- Prashanth, P.; Raveendra, R.; Krishna, R.H.; Ananda, S.; Bhagya, N.; Nagabhushana, B.; Lingaraju, K.; Naika, H.R. Synthesis, characterizations, antibacterial and photoluminescence studies of solution combustion-derived α-Al2O3 nanoparticles. J. Asian Ceram. Soc. 2015, 3, 345–351. [Google Scholar] [CrossRef]
- Balasubramanyam, A.; Sailaja, N.; Mahboob, M.; Rahman, M.F.; Hussain, S.M.; Grover, P. In vivo genotoxicity assessment of aluminium oxide nanomaterials in rat peripheral blood cells using the comet assay and micronucleus test. Mutagenesis 2009, 24, 245–251. [Google Scholar] [CrossRef]
- Rajan, Y.C.; Inbaraj, B.S.; Chen, B.-H. Synthesis and characterization of poly(γ-glutamic acid)-based alumina nanoparticles with their protein adsorption efficiency and cytotoxicity towards human prostate cancer cells. RSC Adv. 2015, 5, 15126–15139. [Google Scholar] [CrossRef]
- Kim, Y.J.; Choi, H.S.; Song, M.K.; Youk, D.Y.; Kim, J.H.; Ryu, J.C. Genotoxicity of aluminum oxide (Al2O3) nanoparticle in mammalian cell lines. Mol. Cell. Toxicol. 2009, 5, 172–178. [Google Scholar]
- Beyzay, F.; Zavaran-Hosseini, A.; Soudi, S. Alpha Alumina Nanoparticle Conjugation to Cysteine Peptidase A and B: An Efficient Method for Autophagy Induction. Avicenna J. Med. Biotechnol. 2017, 9, 71–81. [Google Scholar]
- Ahamed, M.; Siddiqui, M.A.; Akhtar, M.J.; Ahmad, I.; Pant, A.B.; Alhadlaq, H.A. Genotoxic potential of copper oxide nanoparticles in human lung epithelial cells. Biochem. Biophys. Res. Commun. 2010, 396, 578–583. [Google Scholar] [CrossRef]
- Kamble, S.; Utage, B.; Mogle, P.; Kamble, R.; Hese, S.; Dawane, B.; Gacche, R.N. Evaluation of Curcumin Capped Copper Nanoparticles as Possible Inhibitors of Human Breast Cancer Cells and Angiogenesis: A Comparative Study with Native Curcumin. AAPS PharmSciTech 2015, 17, 1030–1041. [Google Scholar] [CrossRef] [PubMed]
- Magrez, A.; Kasas, S.; Salicio, V.; Pasquier, N.; Seo, J.W.; Celio, M.; Catsicas, S.; Schwaller, B.; Forró, L. Cellular Toxicity of Carbon-Based Nanomaterials. Nano Lett. 2006, 6, 1121–1125. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, R.; Basu, T. Metallic copper nanoparticles induce apoptosis in a human skin melanoma A-375 cell line. Nanotechnology 2017, 28, 105101. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Hartono, D.; Ong, C.-N.; Bay, B.-H.; Yung, L.-Y.L. Autophagy and oxidative stress associated with gold nanoparticles. Biomaterials 2010, 31, 5996–6003. [Google Scholar] [CrossRef]
- Arvizo, R.; Bhattacharya, R.; Mukherjee, P. Gold nanoparticles: Opportunities and challenges in nanomedicine. Expert Opin. Drug Deliv. 2010, 7, 753–763. [Google Scholar] [CrossRef]
- Boyoglu, C.; He, Q.; Willing, G.; Boyoglu-Barnum, S.; Dennis, V.A.; Pillai, S.R.; Singh, S.R. Microscopic Studies of Various Sizes of Gold Nanoparticles and Their Cellular Localizations. ISRN Nanotechnol. 2013, 2013, 1–13. [Google Scholar] [CrossRef]
- Felson, D.T.; Anderson, J.J.; Meenan, R.F. The comparative efficacy and toxicity of second-line drugs in rheumatoid arthritis. Arthritis Rheum. 1990, 3, 1449–1461. [Google Scholar] [CrossRef]
- Coradeghini, R.; Gioria, S.; García, C.P.; Nativo, P.; Franchini, F.; Gilliland, D.; Ponti, J.; Rossi, F. Size-dependent toxicity and cell interaction mechanisms of gold nanoparticles on mouse fibroblasts. Toxicol. Lett. 2013, 217, 205–216. [Google Scholar] [CrossRef]
- Mukherjee, P.; Bhattacharya, R.; Bone, N.; Lee, Y.K.; Patra, C.R.; Wang, S.; Lu, L.; Secreto, C.; Banerjee, P.C.; Yaszemski, M.J.; et al. Potential therapeutic application of gold nanoparticles in B-chronic lymphocytic leukemia (BCLL): Enhancing apoptosis. J. Nanobiotechnol. 2007, 5, 4. [Google Scholar] [CrossRef]
- Boyles, M.S.; Kristl, T.; Andosch, A.; Zimmermann, M.; Tran, N.; Casals, E.; Himly, M.; Puntes, V.; Huber, C.G.; Lutz-Meindl, U.; et al. Chitosan functionalisation of gold nanoparticles encourages particle uptake and induces cytotoxicity and pro-inflammatory conditions in phagocytic cells, as well as enhancing particle interactions with serum components. J. Nanobiotechnol. 2015, 13, 84. [Google Scholar] [CrossRef]
- Das, S.K.; Das, A.R.; Guha, A.K. Gold nanoparticles: Microbial synthesis and application in water hygiene management. Langmuir 2009, 25, 8192–8199. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Huang, Y.-W.; Zhou, X.-D.; Ma, Y. In vitro toxicity of silica nanoparticles in human lung cancer cells. Toxicol. Appl. Pharmacol. 2006, 217, 252–259. [Google Scholar] [CrossRef] [PubMed]
- Pattani, V.P.; Tunnell, J.W. Nanoparticle-mediated photothermal therapy: A comparative study of heating for different particle types. Lasers Surg. Med. 2012, 44, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Li, Y.; Liu, X.; Jin, M.; Zhang, L.; Du, Z.; Guo, C.; Huang, P.; Sun, Z. Cytotoxicity and mitochondrial damage caused by silica nanoparticles. Toxicol. In Vitro 2011, 25, 1619–1629. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, L.R.; Stafford, R.J.; Bankson, J.A.; Sershen, S.R.; Rivera, B.; Price, R.E.; Hazle, J.D.; Halas, N.J.; West, J.L. Nanoshell-mediated near-infrared thermal therapy of tumors under magnetic resonance guidance. Proc. Natl. Acad. Sci. USA 2003, 100, 13549–13554. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.M.; Hess, K.; Gearhart, J.; Geiss, K.; Schlager, J. In vitro toxicity of nanoparticles in BRL 3A rat liver cells. Toxicol. In Vitro 2005, 19, 975–983. [Google Scholar] [CrossRef]
- Locatelli, E.; Naddaka, M.; Uboldi, C.; Loudos, G.; Fragogeorgi, E.; Molinari, V.; Pucci, A.; Tsotakos, T.; Psimadas, D.; Ponti, J.; et al. Targeted delivery of silver nanoparticles and alisertib: In vitro and in vivo synergistic effect against glioblastoma. Nanomedicine 2014, 9, 839–849. [Google Scholar] [CrossRef]
- Foldbjerg, R.B.; Dang, D.A.; Autrup, H. Cytotoxicity and genotoxicity of silver nanoparticles in the human lung cancer cell line, A549. Arch. Toxicol. 2011, 85, 743–750. [Google Scholar] [CrossRef]
- Kleinauskas, A.; Rocha, S.; Sahu, S.; Sun, Y.-P.; Juzenas, P. Carbon-core silver-shell nanodots as sensitizers for phototherapy and radiotherapy. Nanotechnology 2013, 24, 325103. [Google Scholar] [CrossRef]
- Haase, A.; Tentschert, J.; Jungnickel, H.; Graf, P.; Mantion, A.; Draude, F.; Plendl, J.; Goetz, M.E.; Galla, S.; Mašić, A.; et al. Toxicity of silver nanoparticles in human macrophages: Uptake, intracellular distribution and cellular responses. J. Phys. Conf. Ser. 2011, 304, 012030. [Google Scholar] [CrossRef]
- Braun, G.B.; Friman, T.; Pang, H.-B.; Pallaoro, A.; de Mendoza, T.H.; Willmore, A.-M.A.; Kotamraju, V.R.; Mann, A.P.; She, Z.-G.; Sugahara, K.N.; et al. Etchable plasmonic nanoparticle probes to image and quantify cellular internalization. Nat. Mater. 2014, 13, 904–911. [Google Scholar] [CrossRef] [PubMed]
- De Berardis, B.; Civitelli, G.; Condello, M.; Lista, P.; Pozzi, R.; Arancia, G.; Meschini, S. Exposure to ZnO nanoparticles induces oxidative stress and cytotoxicity in human colon carcinoma cells. Toxicol. Appl. Pharmacol. 2010, 246, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Namvar, F.; Rahman, H.S.; Mohamad, R.; Azizi, S.; Tahir, P.M.; Chartrand, M.S.; Yeap, S.K. Cytotoxic Effects of Biosynthesized Zinc Oxide Nanoparticles on Murine Cell Lines. Evid.-Based Complement. Altern. Med. 2015, 2015, 593014. [Google Scholar] [CrossRef] [PubMed]
- Osman, I.F.; Baumgartner, A.; Cemeli-Carratala, E.; Fletcher, J.N.; Anderson, D. Genotoxicity and cytotoxicity of zinc oxide and titanium dioxide in HEp-2 cells. Nanomedicine 2010, 5, 1193–1203. [Google Scholar] [CrossRef]
- Tian, L.; Lin, B.; Wu, L.; Li, K.; Liu, H.; Yan, J.; Liu, X.; Xi, Z. Neurotoxicity induced by zinc oxide nanoparticles: Age-related differences and interaction. Sci. Rep. 2015, 5, 16117. [Google Scholar] [CrossRef]
- Sharma, V.; Singh, P.; Pandey, A.K.; Dhawan, A. Induction of oxidative stress, DNA damage and apoptosis in mouse liver after sub-acute oral exposure to zinc oxide nanoparticles. Mutat. Res. Toxicol. Environ. Mutagen. 2012, 745, 84–91. [Google Scholar] [CrossRef]
- Zhang, Y.; Nayak, T.R.; Hong, H.; Cai, W. Biomedical applications of zinc oxide nanomaterials. Curr. Mol. Med. 2013, 1, 633–1645. [Google Scholar] [CrossRef]
- Guan, R.; Kang, T.; Lu, F.; Zhang, Z.; Shen, H.; Liu, M.-Q. Cytotoxicity, oxidative stress, and genotoxicity in human hepatocyte and embryonic kidney cells exposed to ZnO nanoparticles. Nanoscale Res. Lett. 2012, 7, 602. [Google Scholar] [CrossRef]
- Antoine, T.E.; Hadigal, S.R.; Yakoub, A.M.; Mishra, Y.K.; Bhattacharya, P.; Haddad, C.; Valyi-Nagy, T.; Adelung, R.; Prabhakar, B.S.; Shukla, D. Intravaginal Zinc Oxide Tetrapod Nanoparticles as Novel Immunoprotective Agents against Genital Herpes. J. Immunol. 2016, 196, 4566–4575. [Google Scholar] [CrossRef]
- Huang, C.-C.; Aronstam, R.S.; Chen, D.-R.; Huang, Y.-W. Oxidative stress, calcium homeostasis, and altered gene expression in human lung epithelial cells exposed to ZnO nanoparticles. Toxicol. In Vitro 2010, 24, 45–55. [Google Scholar] [CrossRef]
- Khan, Y.A.; Singh, B.R.; Ullah, R.; Mobin, M.; Naqvi, A.H.; Abidi, S.M.A. Anthelmintic Effect of Biocompatible Zinc Oxide Nanoparticles (ZnO NPs) on Gigantocotyle explanatum, a Neglected Parasite of Indian Water Buffalo. PLoS ONE 2015, 10, e0133086. [Google Scholar] [CrossRef] [PubMed]
- Naqvi, S.; Samim, M.; Abdin, M.Z.; Ahmed, F.J.; Maitra, A.N.; Prashant, C.K.; Dinda, A.K. Concentration-dependent toxicity of iron oxide nanoparticles mediated by increased oxidative stress. Int. J. Nanomed. 2010, 5, 983–989. [Google Scholar] [CrossRef] [PubMed]
- P.Melanconab, M.; Lu, W.; Zhongb, M.; Zhoub, M.; Liangd, G.; Elliott, A.M.; Hazle, J.D.; Myers, J.N.; Lia, C.; Stafford, R.J. Targeted multifunctional gold-based nanoshells for magnetic resonance-guided laser ablation of head and neck cancer. Biomaterials 2011, 32, 7600–7608. [Google Scholar] [CrossRef] [PubMed]
- Jeng, H.A.; Swanson, J. Toxicity of Metal Oxide Nanoparticles in Mammalian Cells. J. Environ. Sci. Health Part A 2006, 41, 2699–2711. [Google Scholar] [CrossRef]
- Anselmo, A.C.; Mitragotri, S. A Review of Clinical Translation of Inorganic Nanoparticles. AAPS J. 2015, 17, 1041–1054. [Google Scholar] [CrossRef]
- Ge, Y.; Zhang, Y.; He, S.; Nie, F.; Teng, G.; Gu, N. Fluorescence Modified Chitosan-Coated Magnetic Nanoparticles for High-Efficient Cellular Imaging. Nanoscale Res. Lett. 2009, 4, 287–295. [Google Scholar] [CrossRef]
- Jordan, A.; Scholz, R.; Maier-Hauff, K.; Van Landeghem, F.K.; Waldoefner, N.; Teichgraeber, U.; Pinkernelle, J.; Bruhn, H.; Neumann, F.; Thiesen, B.; et al. The effect of thermotherapy using magnetic nanoparticles on rat malignant glioma. J. Neuro-Oncol. 2006, 78, 7–14. [Google Scholar] [CrossRef]
- Delcroix, G.J.-R.; Jacquart, M.; Lemaire, L.; Sindji, L.; Franconi, F.; Le Jeune, J.-J.; Montero-Menei, C.N. Mesenchymal and neural stem cells labeled with HEDP-coated SPIO nanoparticles: In vitro characterization and migration potential in rat brain. Brain Res. 2009, 1255, 18–31. [Google Scholar] [CrossRef]
- Jurgons, R.; Seliger, C.; Hilpert, A.; Trahms, L.; Odenbach, S.; Alexiou, C. Drug loaded magnetic nanoparticles for cancer therapy. J. Phys. Condens. Matter 2006, 18, S2893–S2902. [Google Scholar] [CrossRef]
- Liu, R.; Yin, L.; Pu, Y.; Liang, G.; Zhang, J.; Su, Y.; Xiao, Z.; Ye, B. Pulmonary toxicity induced by three forms of titanium dioxide nanoparticles via intra-tracheal instillation in rats. Prog. Nat. Sci. 2009, 19, 573–579. [Google Scholar] [CrossRef]
- Fujiwara, R.; Luo, Y.; Sasaki, T.; Fujii, K.; Ohmori, H.; Kuniyasu, H. Cancer Therapeutic Effects of Titanium Dioxide Nanoparticles Are Associated with Oxidative Stress and Cytokine Induction. Pathobiology 2015, 82, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, K.; Davoren, M.; Boertz, J.; Schins, R.P.; Hoffmann, E.; Dopp, E. Titanium dioxide nanoparticles induce oxidative stress and DNA-adduct formation but not DNA-breakage in human lung cells. Part. Fibre Toxicol. 2009, 6, 17. [Google Scholar] [CrossRef] [PubMed]
- Sortino, S. Photoactivated nanomaterials for biomedical release applications. J. Mater. Chem. 2012, 22, 301–318. [Google Scholar] [CrossRef]
- Lee, Y.-H.; Cheng, F.-Y.; Chiu, H.-W.; Tsai, J.-C.; Fang, C.-Y.; Chen, C.-W.; Wang, Y.-J. Cytotoxicity, oxidative stress, apoptosis and the autophagic effects of silver nanoparticles in mouse embryonic fibroblasts. Biomaterials 2014, 35, 4706–4715. [Google Scholar] [CrossRef]
- Braydich-Stolle, L.; Hussain, S.; Schlager, J.J.; Hofmann, M.-C. In Vitro Cytotoxicity of Nanoparticles in Mammalian Germline Stem Cells. Toxicol. Sci. 2005, 88, 412–419. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, R.; Ni, M.; Gill, P.; Lee, A.S. Cell surface relocalization of the endoplasmic reticulum chaperone and unfolded protein response regulator GRP78/BiP. J. Biol. Chem. 2010, 285, 15065–15075. [Google Scholar] [CrossRef]
- Xu, Z.; Bu, Y.; Chitnis, N.; Koumenis, C.; Fuchs, S.Y.; Diehl, J.A. miR-216b regulation of c-Jun mediates GADD153/CHOP-dependent apoptosis. Nat. Commun. 2016, 7, 1–12. [Google Scholar] [CrossRef]
- Hetz, C.; Bernasconi, P.; Fisher, J.; Lee, A.-H.; Bassik, M.C.; Antonsson, B.; Brandt, G.S.; Iwakoshi, N.N.; Schinzel, A.; Glimcher, L.H.; et al. Proapoptotic BAX and BAK Modulate the Unfolded Protein Response by a Direct Interaction with IRE1. Science 2006, 312, 572–576. [Google Scholar] [CrossRef]
- Noh, M.R.; Kim, J.I.; Han, S.J.; Lee, T.-J.; Park, K.M. C/EBP homologous protein (CHOP) gene deficiency attenuates renal ischemia/reperfusion injury in mice. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2015, 1852, 1895–1901. [Google Scholar] [CrossRef]
- Li, Y.; Bhalli, J.A.; Ding, W.; Yan, J.; Pearce, M.G.; Sadiq, R.; Cunningham, C.K.; Jones, M.Y.; Monroe, W.A.; Howard, P.C.; et al. Cytotoxicity and genotoxicity assessment of silver nanoparticles in mouse. Nanotoxicology 2013, 8, 36–45. [Google Scholar] [CrossRef]
- De Jong, W.H.; Van Der Ven, L.T.; Sleijffers, A.; Park, M.V.; Jansen, E.H.; Van Loveren, H.; Vandebriel, R.J. Systemic and immunotoxicity of silver nanoparticles in an intravenous 28 days repeated dose toxicity study in rats. Biomaterials 2013, 34, 8333–8343. [Google Scholar] [CrossRef] [PubMed]
- Hackenberg, S.; Scherzed, A.; Kessler, M.; Hummel, S.; Technau, A.; Froelich, K.; Ginzkey, C.; Koehler, C.; Hagen, R.; Kleinsasser, N. Silver nanoparticles: Evaluation of DNA damage, toxicity and functional impairment in human mesenchymal stem cells. Toxicol. Lett. 2011, 201, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Zhao, L.; Bai, R.; Liu, Y.; Han, L.; Xu, Z.; Chen, F.; Autrup, H.; Long, D.; Chen, C. Silver nanoparticles induced oxidative and endoplasmic reticulum stresses in mouse tissues: Implications for the development of acute toxicity after intravenous administration. Toxicol. Res. 2016, 5, 602–608. [Google Scholar] [CrossRef] [PubMed]
- 5Kozuka, C.; Shimizu-Okabe, C.; Takayama, C.; Nakano, K.; Morinaga, H.; Kinjo, A.; Fukuda, K.; Kamei, A.; Yasuoka, A.; Kondo, T.; et al. Marked augmentation of PLGA nanoparticle-induced metabolically beneficial impact of γ-oryzanol on fuel dyshomeostasis in genetically obese-diabetic ob/ob mice. Drug Deliv. 2017, 24, 558–568. [Google Scholar] [CrossRef]
- Kuang, H.; Yang, P.; Yang, L.; Aguilar, Z.P.; Xu, H. Size dependent effect of ZnO nanoparticles on endoplasmic reticulum stress signaling pathway in murine liver. J. Hazard. Mater. 2016, 317, 119–126. [Google Scholar] [CrossRef]
- Li, J.; Zhou, Y.; Zhang, W.; Bao, C.; Xie, Z. Relief of oxidative stress and cardiomyocyte apoptosis by using curcumin nanoparticles. Colloids Surf. B Biointerfaces 2017, 153, 174–182. [Google Scholar] [CrossRef]
- Spyratou, E.; Makropoulou, M.; Efstathopoulos, E.; Georgakilas, A.; Sihver, L. Recent Advances in Cancer Therapy Based on Dual Mode Gold Nanoparticles. Cancers 2017, 9, 173. [Google Scholar] [CrossRef]
- Li, J.; Gupta, S.; Li, C. Research perspectives: Gold nanoparticles in cancer theranostics. Quant. Imaging Med. Surg. 2013, 3, 284–291. [Google Scholar]
- Herold, I.J.D.D.M. Gold microspheres: A selective technique for producing biologically effective dose enhancement. Int. J. Radiat. Biol. 2000, 76, 1357–1364. [Google Scholar] [CrossRef]
- Hainfeld, J.F.; Smilowitz, H.M.; O’Connor, M.J.; Dilmanian, F.A.; Slatkin, D.N. Gold nanoparticle imaging and radiotherapy of brain tumors in mice. Nanomedicine 2013, 8, 1601–1609. [Google Scholar] [CrossRef]
- Chang, M.-Y.; Shiau, A.-L.; Chen, Y.-H.; Chang, C.-J.; Chen, H.H.W.; Wu, C.-L. Increased apoptotic potential and dose-enhancing effect of gold nanoparticles in combination with single-dose clinical electron beams on tumor-bearing mice. Cancer Sci. 2008, 99, 1479–1484. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-J.; Wang, C.-H.; Chen, S.-T.; Chen, H.-H.; Leng, W.-H.; Chien, C.-C.; Wang, C.-L.; Kempson, I.M.; Hwu, Y.; Lai, T.-C.; et al. Enhancement of cell radiation sensitivity by pegylated gold nanoparticles. Phys. Med. Biol. 2010, 55, 931–945. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Tamura, A.; Murotani, H.; Oishi, M.; Jinji, Y.; Matsuishi, K.; Nagasaki, Y. Large payloads of gold nanoparticles into the polyamine network core of stimuli-responsive PEGylated nanogels for selective and noninvasive cancer photothermal therapy. Nanoscale 2010, 2, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Oishi, M.; Nagasaki, Y. Synthesis, characterization, and biomedical applications of core–shell-type stimuli-responsive nanogels—Nanogel composed of poly[2-(N,N-diethylamino)ethyl methacrylate] core and PEG tethered chains. React. Funct. Polym. 2007, 67, 1311–1329. [Google Scholar] [CrossRef]
- Pan, Y.; Leifert, A.; Ruau, D.; Neuss, S.; Bornemann, J.; Schmid, G.; Brandau, W.; Simon, U.; Jahnen-Dechent, W. Gold Nanoparticles of Diameter 1.4 nm Trigger Necrosis by Oxidative Stress and Mitochondrial Damage. Small 2009, 5, 2067–2076. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fang, X.; Liang, W. Pegylated Phospholipid Micelles Induce Endoplasmic Reticulum-Dependent Apoptosis of Cancer Cells but not Normal Cells. ACS Nano 2012, 6, 5018–5030. [Google Scholar] [CrossRef]
- Zhou, J.; Yang, F.; Zhou, Q.; Yang, K.; Chen, W. Magnetic resonance imaging tracking of ultra small superparamagnetic iron oxide labeled rabbit dendritic cells. Exp. Biol. Med. 2013, 239, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Smits, L.P.; Coolen, B.F.; Panno, M.D.; Runge, J.H.; Nijhof, W.H.; Verheij, J.; Nieuwdorp, M.; Stoker, J.; Beuers, U.H.; Nederveen, A.J.; et al. Noninvasive Differentiation between Hepatic Steatosis and Steatohepatitis with MR Imaging Enhanced with USPIOs in Patients with Nonalcoholic Fatty Liver Disease: A Proof-of-Concept Study. Radiology 2016, 278, 782–791. [Google Scholar] [CrossRef]
- Kanematsu, M.; Kondo, H.; Goshima, S.; Kato, H.; Tsuge, U.; Hirose, Y.; Kim, M.-J.; Moriyama, N. Imaging Liver Metastases: Review and Update. Eur. J. Radiol. 2006, 58, 217–228. [Google Scholar] [CrossRef]
- Tanimoto, A.; Kuribayashi, S. Application of Superparamagnetic Iron Oxide to Imaging of Hepatocellular Carcinoma. Eur. J. Radiol. 2006, 58, 200–216. [Google Scholar] [CrossRef]
- Rock, K.L.; Kono, H. The Inflammatory Response to Cell Death. Annu. Rev. Pathol. Mech. Dis. 2008, 3, 99–126. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Jiang, S.; Yao, H.; Zhang, L.; Yang, C.; Zhan, D.; Lin, G.; Zeng, Y.; Xia, Y.; Lin, Z.; et al. Endoplasmic reticulum stress mediates inflammatory response triggered by ultra-small superparamagnetic iron oxide nanoparticles in hepatocytes. Nanotoxicology 2018, 12, 1198–1214. [Google Scholar] [CrossRef] [PubMed]
- Sangaiya, P.; Jayaprakash, R. A Review on Iron Oxide Nanoparticles and Their Biomedical Applications. J. Supercond. Nov. Magn. 2018, 31, 3397–3413. [Google Scholar] [CrossRef]
- Rajendran, K.; Karunagaran, V.; Mahanty, B.; Sen, S. Biosynthesis of hematite nanoparticles and its cytotoxic effect on HepG2 cancer cells. Int. J. Biol. Macromol. 2015, 74, 376–381. [Google Scholar] [CrossRef]
- Calero, M.; Chiappi, M.; Lazaro-Carrillo, A.; Rodriguez, M.J.; Chichon, F.J.; Crosbie-Staunton, K.; Prina-Mello, A.; Volkov, Y.; Villanueva, A.; Carrascosa, J.L. Characterization of interaction of magnetic nanoparticles with breast cancer cells. J. Nanobiotechnol. 2015, 13, 1–15. [Google Scholar] [CrossRef]
- Abrams, D.B.; Witkiewitz, K.; Fulmer, T.; Tanenbaum, M.L.; Commissariat, P.; Kupperman, E.; Baek, R.N.; Gonzalez, J.S.; Heaney, J.; Kline, C.; et al. Apple Shaped. Encycl. Behav. Med. 2013, 281, 124. [Google Scholar] [CrossRef]
- Kossatz, S.; Grandke, J.; Couleaud, P.; Latorre, A.; Aires, A.; Crosbie-Staunton, K.; Ludwig, R.; Dahring, H.; Ettelt, V.; Lazaro-Carrillo, A.; et al. Efficient treatment of breast cancer xenografts with multifunctionalized iron oxide nanoparticles combining magnetic hyperthermia and anti-cancer drug delivery. Breast Cancer Res. 2015, 17, 1–17. [Google Scholar] [CrossRef]
- Nagamuthu, S.; Vijayakumar, S.; Muralidharan, G. Synthesis of Mn3O4/Amorphous Carbon Nanoparticles as Electrode Material for High Performance Supercapacitor Applications. Energy Fuels 2013, 27, 3508. [Google Scholar] [CrossRef]
- Parveen, A.; Rizvi, S.H.M.; Tiwari, S.; Mahdi, A.A. Manganese Nanoparticles Induce Endoplasmic Reticulum Stress Mediated Cell Death in Primary Hippocampal Neuron Culture. Free Radic. Biol. Med. 2017, 112, 77. [Google Scholar] [CrossRef]
- Pradhan, S.; Patra, P.; Das, S.; Chandra, S.; Mitra, S.; Dey, K.K.; Akbar, S.; Palit, P.; Goswami, A. Photochemical Modulation of Biosafe Manganese Nanoparticles on Vigna radiata: A Detailed Molecular, Biochemical, and Biophysical Study. Environ. Sci. Technol. 2013, 47, 13122–13131. [Google Scholar] [CrossRef] [PubMed]
- Sanfins, E.; Dairou, J.; Rodrigues-Lima, F.; Dupret, J.-M. Nanoparticle-protein interactions: From crucial plasma proteins to key enzymes. J. Phys. Conf. Ser. 2011, 304, 012039. [Google Scholar] [CrossRef]
- Marano, S.; Hussain, F.; Rodrigues-Lima, A.; Baeza-Squiban, A.; Boland, S. Nanoparticles: Molecular targets and cell signalling. Arch. Toxicol. 2011, 85, 733. [Google Scholar] [CrossRef] [PubMed]
- Alarifi, S.; Ali, D.; Alkahtani, S. Oxidative Stress-Induced DNA Damage by Manganese Dioxide Nanoparticles in Human Neuronal Cells. BioMed Res. Int. 2017, 2017, 5478790. [Google Scholar] [CrossRef] [PubMed]
- Yi, X.; Zhao, W.; Li, J.; Zhang, B.; Yu, Q.; Li, M. Mn3O4nanoparticles cause endoplasmic reticulum stress-dependent toxicity to Saccharomyces cerevisiae. RSC Adv. 2017, 7, 46028–46035. [Google Scholar] [CrossRef]
- Li, J.; Wang, X.; Jiang, H.; Lü, X.; Zhu, Y.; Chen, B. New strategy of photodynamic treatment of TiO2 nanofibers combined with celastrol for HepG2 proliferation in vitro. Nanoscale 2011, 3, 3115–3122. [Google Scholar] [CrossRef]
- Reeves, J.F.; Davies, S.J.; Dodd, N.J.; Jha, A.N. Hydroxyl radicals (OH) are associated with titanium dioxide (TiO2) nanoparticle-induced cytotoxicity and oxidative DNA damage in fish cells. Mutat. Res. 2008, 640, 113–122. [Google Scholar] [CrossRef]
- Tang, Y.; Wang, F.; Jin, C.; Liang, H.; Zhong, X.; Yang, Y. Mitochondrial injury induced by nanosized titanium dioxide in A549 cells and rats. Environ. Toxicol. Pharmacol. 2013, 36, 66–72. [Google Scholar] [CrossRef]
- Oesch, F.; Landsiedel, R. Genotoxicity investigations on nanomaterials. Arch. Toxicol. 2012, 86, 985–994. [Google Scholar] [CrossRef]
- Van Vliet, A.R.; Verfaillie, T.; Agostinis, P. New functions of mitochondria associated membranes in cellular signaling. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2014, 1843, 2253–2262. [Google Scholar] [CrossRef]
- Verma, S.K.; Jha, E.; Panda, P.K.; Thirumurugan, A.; Parashar, S.K.S.; Patro, S.; Suar, M. Mechanistic Insight into Size-Dependent Enhanced Cytotoxicity of Industrial Antibacterial Titanium Oxide Nanoparticles on Colon Cells Because of Reactive Oxygen Species Quenching and Neutral Lipid Alteration. ACS Omega 2018, 3, 1244–1262. [Google Scholar] [CrossRef]
- Shukla, R.K.; Sharma, V.; Pandey, A.K.; Singh, S.; Sultana, S.; Dhawan, A. ROS-mediated genotoxicity induced by titanium dioxide nanoparticles in human epidermal cells. Toxicol. In Vitro 2011, 25, 231–241. [Google Scholar] [CrossRef]
- Aliakbari, F.; Hosseinali, S.H.; Sarokhalil, Z.K.; Shahpasand, K.; Saboury, A.A.; Akhtari, K.; Falahati, M. Reactive oxygen species generated by titanium oxide nanoparticles stimulate the hemoglobin denaturation and cytotoxicity against human lymphocyte cell. J. Biomol. Struct. Dyn. 2019, 37, 4875–4881. [Google Scholar] [CrossRef] [PubMed]
- Malhotra, J.D.; Kaufman, R.J. ER Stress and Its Functional Link to Mitochondria: Role in Cell Survival and Death. Cold Spring Harb. Perspect. Biol. 2011, 3, a004424. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.-N.; Yoon, T.-J.; Minai-Tehrani, A.; Kim, J.-E.; Park, S.J.; Jeong, M.S.; Ha, S.-W.; Lee, J.-K.; Kim, J.S.; Cho, M.-H. Zinc oxide nanoparticle induced autophagic cell death and mitochondrial damage via reactive oxygen species generation. Toxicol. In Vitro 2013, 27, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- Murdock, R.C.; Braydich-Stolle, L.; Schrand, A.M.; Schlager, J.J.; Hussain, S.M. Characterization of nanomaterial dispersion in solution prior to in vitro exposure using dynamic light scattering technique. Toxicol. Sci. 2008, 10, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Kahn, B.B.; Alquier, T.; Carling, D.; Hardie, D.G. AMP-activated protein kinase: Ancient energy gauge provides clues to modern understanding of metabolism. Cell Metab. 2005, 1, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Huo, L.; Shi, X.; Bai, R.; Zhang, Z.; Zhao, Y.; Chang, Y.; Chen, C. Endoplasmic Reticulum Stress Induced by Zinc Oxide Nanoparticles Is an Earlier Biomarker for Nanotoxicological Evaluation. ACS Nano 2014, 8, 2562–2574. [Google Scholar] [CrossRef]
- Piccinno, F.; Gottschalk, F.; Seeger, S.; Nowack, B. Industrial production quantities and uses of ten engineered nanomaterials in Europe and the world. J. Nanoparticle Res. 2012, 14, 1–11. [Google Scholar] [CrossRef]
- Cyphert, J.M.; Hargrove, M.M.; Nyska, A.; Schladweiler, M.C.; Kodavanti, U.P.; Gavett, S.H. Long-term toxicity of naturally occurring asbestos in male Fischer 344 rats. J. Toxicol. Environ. Health Part A 2016, 79, 49–60. [Google Scholar] [CrossRef]
- Pastero, L.; Turci, F.; Leinardi, R.; Pavan, C.; Monopoli, M. Synthesis of α-Quartz with Controlled Properties for the Investigation of the Molecular Determinants in Silica Toxicology. Cryst. Growth Des. 2016, 16, 2394–2403. [Google Scholar] [CrossRef]
- Turci, F.; Pavan, C.; Leinardi, R.; Tomatis, M.; Pastero, L.; Garry, D.; Anguissola, S.; Lison, D.; Fubini, B. Revisiting the paradigm of silica pathogenicity with synthetic quartz crystals: The role of crystallinity and surface disorder. Part. Fibre Toxicol. 2015, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, H. A mechanistic review of silica-induced inhalation toxicity. Inhal. Toxicol. 2015, 27, 363–377. [Google Scholar] [CrossRef] [PubMed]
- Echaudhari, N.; Etalwar, P.; Eparimisetty, A.; d’Hellencourt, C.L.; Ravanan, P. A Molecular Web: Endoplasmic Reticulum Stress, Inflammation, and Oxidative Stress. Front. Cell. Neurosci. 2014, 8, 213. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlachogianni, T.; Fiotakis, K.; Loridas, S. Pulmonary Oxidative Stress, Inflammation and Cancer: Respirable Particulate Matter, Fibrous Dusts and Ozone as Major Causes of Lung Carcinogenesis through Reactive Oxygen Species Mechanisms. Int. J. Environ. Res. Public Health 2013, 10, 3886–3907. [Google Scholar] [CrossRef] [PubMed]
- Sushma, S.; Kumar, H.; Ahmad, I.; Dutta, P.K. Evaluation of the DNA damaging potential of indigenous health hazardous quartz nanoparticles on the cultured lung cells. Toxicol. Res. 2017, 6, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, R.J.; Malhotra, J.D. Calcium trafficking integrates endoplasmic reticulum function with mitochondrial bioenergetics. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2014, 1843, 2233–2239. [Google Scholar] [CrossRef]
- Kania, E.; Pająk, B.; Orzechowski, A. Calcium Homeostasis and ER Stress in Control of Autophagy in Cancer Cells. BioMed Res. Int. 2015, 2015, 1–12. [Google Scholar] [CrossRef]
- Rizzuto, R.; De Stefani, D.; Raffaello, A.; Mammucari, C. Mitochondria as sensors and regulators of calcium signalling. Nat. Rev. Mol. Cell Biol. 2012, 13, 566–578. [Google Scholar] [CrossRef]
- Orrenius, S.; Gogvadze, V.; Zhivotovsky, B. Calcium and mitochondria in the regulation of cell death. Biochem. Biophys. Res. Commun. 2015, 460, 72–81. [Google Scholar] [CrossRef]
- Mittal, S.; Pandey, A.K. Cerium Oxide Nanoparticles Induced Toxicity in Human Lung Cells: Role of ROS Mediated DNA Damage and Apoptosis. BioMed Res. Int. 2014, 2014, 1–14. [Google Scholar] [CrossRef]
- Akhtar, M.J.; Ahamed, M.; Khan, M.A.; Alrokayan, S.A.; Ahmad, I.; Kumar, S. Cytotoxicity and apoptosis induction by nanoscale talc particles from two different geographical regions in human lung epithelial cells? Environ. Toxicol. 2014, 29, 394–406. [Google Scholar] [CrossRef] [PubMed]
- Passagne, I.; Morille, M.; Rousset, M.; Pujalté, I.; L’Azou, B. Implication of oxidative stress in size-dependent toxicity of silica nanoparticles in kidney cells. Toxicology 2012, 299, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Napierska, D.; Thomassen, L.C.; Lison, D.; Martens, J.A.; Hoet, P.H.M. The nanosilica hazard: Another variable entity. Part. Fibre Toxicol. 2010, 7, 39. [Google Scholar] [CrossRef] [PubMed]
- Ariano, P.; Zamburlin, A.; Gilardino, R.; Mortera, B.; Onida, M.; Tomatis, M.; Ghiazza, B.; Fubini, D.; Lovisolo, D. Interaction of spherical silica nanoparticleswith neuronal cells: Size dependent toxicity and perturbation of calciumhomeostasis. Small 2011, 7, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Tarantini, R.; Lanceleur, A.; Mourot, M.T.; Lavault, G.; Casterou, G.; Jarry, K.; Hogeveen, V.; Fessard, V. Toxicity, genotoxicity and proinflammatory effects ofamorphous nanosilica in the human intestinal Caco-2 cell line. Toxicol. In Vitro 2015, 29, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Sahu, D.; Vijayaraghavan, R.; Kannan, G.M. Silica nanoparticle induces oxidativestress and provokes inflammation in human lung cells, Experiment. Nanoscience 2015, 10, 983–1000. [Google Scholar]
- Panas, C.; Marquardt, O.; Nalcaci, H.; Bockhorn, W.; Baumann, H.R.; Paur, S.; Mülhopt, S.; Diabaté, C.; Weiss, C. Screening of different metal oxide nanoparticlesreveals selective toxicity and inflammatory potential of silica nanoparticles inlung epithelial cells and macrophages. Nanotoxicology 2013, 7, 259–273. [Google Scholar] [CrossRef]
- Christen, V.; Fent, K. Silica nanoparticles and silver-doped silica nanoparticles induce endoplasmic reticulum stress response and alter cytochromeP4501A activity. Chemosphere 2012, 87, 423–434. [Google Scholar] [CrossRef]
- Christen, V.; Camenzind, M.; Fent, K. Silica nanoparticles induce endoplasmic reticulum stress response, oxidative stress and activate the mitogen-activated protein kinase (MAPK) signaling pathway. Toxicol. Rep. 2014, 1, 1143–1151. [Google Scholar] [CrossRef]
- Hu, H.; Fan, X.; Guo, Q.; Wei, X.; Yang, D.; Zhang, B.; Liu, J.; Wu, Q.; Oh, Y.; Feng, Y.; et al. Silicon dioxide nanoparticles induce insulin resistance through endoplasmic reticulum stress and generation of reactive oxygen species. Part. Fibre Toxicol. 2019, 16, 1–18. [Google Scholar] [CrossRef]
- Ghosh, C.; Nandi, A.; Basu, S. Lipid Nanoparticle-Mediated Induction of Endoplasmic Reticulum Stress in Cancer Cells. ACS Appl. Bio Mater. 2019, 2, 3992–4001. [Google Scholar] [CrossRef]
- Ghalhar, M.G.; Akbarzadeh, A.; Rahmati, M.; Mellatyar, H.; Dariushnejad, H.; Zarghami, N.; Barkhordari, A. Comparison of inhibitory effects of 17-AAG nanoparticles and free 17-AAG on HSP90 gene expression in breast cancer. Asian Pac. J. Cancer Prev. 2014, 15, 7113–7118. [Google Scholar] [CrossRef] [PubMed]
- Jiang, D.; Lynch, C.; Medeiros, B.C.; Liedtke, M.; Bam, R.; Tam, A.B.; Yang, Z.; Alagappan, M.; Abidi, P.; Le, Q.T.; et al. Identification of doxorubicin as an inhibitor of the ire1α-xbp1 axis of the unfolded protein response. Sci. Rep. 2016, 6, 33353. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wang, C.; Li, Z. A new strategy of promoting cisplatin chemotherapeutic efficiency by targeting endoplasmic reticulum stress. Mol. Clin. Oncol. 2014, 2, 3–7. [Google Scholar] [CrossRef]
- Karasawal, T.; Sibrian-Vazquez, M.; Strongin, R.M.; Steyger, P.S. Identification of cisplatin-binding proteins using agarose conjugates of platinum compounds. PLoS ONE 2013, 8, e66220. [Google Scholar]
- Chen, G.-Y.; Yang, H.-J.; Lu, C.-H.; Chao, Y.-C.; Hwang, S.-M.; Chen, C.-L.; Lo, K.-W.; Sung, L.-Y.; Luo, W.-Y.; Tuan, H.-Y.; et al. Simultaneous induction of autophagy and toll-like receptor signaling pathways by graphene oxide. Biomaterials 2012, 33, 6559–6569. [Google Scholar] [CrossRef]
- Jin, P.; Wei, P.; Zhang, Y.; Lin, J.; Sha, R.; Hu, Y.; Zhang, J.; Zhou, W.; Yao, H.; Ren, L.; et al. Autophagy-mediated clearance of ubiquitinated mutant huntingtin by graphene oxide. Nanoscale 2016, 8, 18740–18750. [Google Scholar] [CrossRef]
- Pandey, S.; Nandi, A.; Basu, S.; Ballav, N. Inducing endoplasmic reticulum stress in cancer cells by graphene oxide-based nanoparticles. Nanoscale Adv. 2020. [Google Scholar] [CrossRef]
- Wang, J.; Song, Y.; Zhang, M.; Wu, Z.; Xu, Y.-J.; Lin, J.; Ling, D.; Sheng, Y.; Lu, Y.; Wu, Q.; et al. A liposomal curcumol nanocomposite for magnetic resonance imaging and endoplasmic reticulum stress-mediated chemotherapy of human primary ovarian cancer. J. Mater. Chem. B 2019, 7, 2938–2947. [Google Scholar] [CrossRef]
- Song, Y.; Sheng, Z.; Xu, Y.; Dong, L.; Xu, W.; Li, F.; Wang, J.; Wu, Z.; Yang, Y.; Su, Y.; et al. Magnetic liposomal emodin composite with enhanced killing efficiency against breast cancer. Biomater. Sci. 2019, 7, 867–875. [Google Scholar] [CrossRef]
- Mollereau, B. Establishing Links between Endoplasmic Reticulum-Mediated Hormesis and Cancer. Mol. Cell. Biol. 2013, 33, 2372–2374. [Google Scholar] [CrossRef] [PubMed]
- Foufelle, F.; Fromenty, B. Role of endoplasmic reticulum stress in drug-induced toxicity. Pharmacol. Res. Perspect. 2016, 4, e00211. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Xu, B. Instructed-assembly (IA): A molecular process for controlling cell fate. Bull. Chem. Soc. Jpn. 2018, 91, 900. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.Q.Q.; Wang, H.M.; Du, X.W.; Shi, J.F.; Li, J.; Xu, B. Minimal C-terminal modification boosts peptide self-assembling ability for necroptosis of cancer cells. Chem. Commun. 2016, 52, 6332–6335. [Google Scholar] [CrossRef]
- Tanaka, A.; Fukuoka, Y.; Morimoto, Y.; Honjo, T.; Koda, D.; Goto, M.; Maruyama, T.J. Cancer cell death induced by the intracellular self-assembly of an enzyme-responsive supramolecular gelator. Am. Chem. Soc. 2015, 137, 770. [Google Scholar] [CrossRef]
- Kholodenko, B.N.; Hoek, J.B.; Westerhoff, H.V. Why cytoplasmic signalling proteins should be recruited to cell membranes. Trends Cell Biol. 2000, 10, 173. [Google Scholar] [CrossRef]
- Cai, Y.; Shen, H.; Zhan, J.; Lin, M.; Dai, L.; Ren, C.; Shi, Y.; Liu, J.; Gao, J.; Yang, Z. Supramolecular “Trojan Horse” for nuclear delivery of dual anticancer drugs. J. Am. Chem. Soc. 2017, 139, 2876. [Google Scholar] [CrossRef]
- Wang, H.; Feng, Z.; Wang, Y.; Zhou, R.; Yang, Z.; Xu, B.J. Integrating enzymatic self-assembly and mitochondria targeting for selectively killing cancer cells without acquired drug resistance. Am. Chem. Soc. 2016, 138, 16046. [Google Scholar] [CrossRef]
- Abu Gazia, M.; El-Magd, M.A. Effect of pristine and functionalized multiwalled carbon nanotubes on rat renal cortex. Acta Histochem. 2018, 121, 207–217. [Google Scholar] [CrossRef]
- Guo, Y.-Y.; Zhang, J.; Zheng, Y.-F.; Yang, J.; Zhu, X.-Q. Cytotoxic and genotoxic effects of multi-wall carbon nanotubes on human umbilical vein endothelial cells in vitro. Mutat. Res. Toxicol. Environ. Mutagen. 2011, 721, 184–191. [Google Scholar] [CrossRef]
- Orecna, M.; De Paoli, S.H.; Janouskova, O.; Tegegn, T.Z.; Filipova, M.; Bonevich, J.E.; Holada, K.; Simak, J. Toxicity of carboxylated carbon nanotubes in endothelial cells is attenuated by stimulation of the autophagic flux with the release of nanomaterial in autophagic vesicles. Nanomed. Nanotechnol. Biol. Med. 2014, 10, e939–e948. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Chang, S.; Long, J.; Li, J.; Li, X.; Cao, Y. The toxicity of multi-walled carbon nanotubes (MWCNTs) to human endothelial cells: The influence of diameters of MWCNTs. Food Chem. Toxicol. 2019, 126, 169–177. [Google Scholar] [CrossRef]
- Saleem, J.; Wang, L.; Chen, C. Carbon-Based Nanomaterials for Cancer Therapy via Targeting Tumor Microenvironment. Adv. Health Mater. 2018, 7, e1800525. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shen, W.; Gurunathan, S. Silver Nanoparticle-Mediated Cellular Responses in Various Cell Lines: An in Vitro Model. Int. J. Mol. Sci. 2016, 17, 1603. [Google Scholar] [CrossRef] [PubMed]
- Faedmaleki, F.; Shirazi, F.H.; Salarian, A.-A.; Ashtiani, H.A.; Rastegar, H. Toxicity Effect of Silver Nanoparticles on Mice Liver Primary Cell Culture and HepG2 Cell Line. Iran. J. Pharm. Res. IJPR 2014, 13, 235–242. [Google Scholar]
- Akter, M.; Sikder, T.; Rahman, M.; Ullah, A.A.; Hossain, K.F.B.; Banik, S.; Hosokawa, T.; Saito, T.; Kurasaki, M. A systematic review on silver nanoparticles-induced cytotoxicity: Physicochemical properties and perspectives. J. Adv. Res. 2018, 9, 1–16. [Google Scholar] [CrossRef]
- Bin-Jumah, M.; Al-Abdan, M.; Albasher, G.; Alarifi, S. Effects of Green Silver Nanoparticles on Apoptosis and Oxidative Stress in Normal and Cancerous Human Hepatic Cells in vitro. Int. J. Nanomed. 2020, 15, 1537–1548. [Google Scholar] [CrossRef]
- Hekmat, A.; Saboury, A.A.; Divsalar, A. The Effects of Silver Nanoparticles and Doxorubicin Combination on DNA Structure and Its Antiproliferative Effect Against T47D and MCF7 Cell Lines. J. Biomed. Nanotechnol. 2012, 8, 968–982. [Google Scholar] [CrossRef]
- Gunduz, N.; Ceylan, H.; Guler, M.O.; Tekinay, A.B. Intracellular Accumulation of Gold Nanoparticles Leads to Inhibition of Macropinocytosis to Reduce the Endoplasmic Reticulum Stress. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Rahme, K.; Minassian, G.B.; Sarkis, M.; Nakhl, M.; El Hage, R.; Souaid, E.; Holmes, J.D.; Ghanem, E. Assessment of Charged AuNPs: From Synthesis to Innate Immune Recognition. J. Nanomater. 2018, 2018, 1–12. [Google Scholar] [CrossRef]
- Huai, Y.; Zhang, Y.; Xiong, X.; Das, S.; Bhattacharya, R.; Mukherjee, P. Gold Nanoparticles sensitize pancreatic cancer cells to gemcitabine. Cell Stress 2019, 3, 267–279. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-C.; Yang, Y.-C.; Yang, K.-C.; Shieh, H.-R.; Wang, T.-Y.; Hwu, Y.; Chen, Y.-J. Pegylated Gold Nanoparticles Induce Apoptosis in Human Chronic Myeloid Leukemia Cells. BioMed Res. Int. 2014, 2014, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yasui, H.; Takeuchi, R.; Nagane, M.; Meike, S.; Nakamura, Y.; Yamamori, T.; Ikenaka, Y.; Kon, Y.; Murotani, H.; Oishi, M.; et al. Radiosensitization of tumor cells through endoplasmic reticulum stress induced by PEGylated nanogel containing gold nanoparticles. Cancer Lett. 2014, 347, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Yang, D.; Zhou, X.; Wang, Y.; Tang, S.; Yin, H.; Wang, J.; Chen, R.; Chen, J. Role of Autophagy in Zinc Oxide Nanoparticles-Induced Apoptosis of Mouse LEYDIG Cells. Int. J. Mol. Sci. 2019, 20, 4042. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.T.; Yong, L.Q.; Hande, M.P.; Ong, C.N.; Yu, L.E.; Bay, B.H.; Baeg, G.H. Zinc oxide nanoparticles exhibit cytotoxicity and genotoxicity through oxidative stress responses in human lung fibroblasts and Drosophila melanogaster. Int. J. Nanomed. 2017, 12, 1621–1637. [Google Scholar] [CrossRef] [PubMed]
- An, S.S.A.; Kim, Y.-R.; Park, J.-I.; Lee, E.J.; Park, S.H.; Seong, N.-W.; Kim, J.-H.; Kim, G.-Y.; Meang, E.-H.; Hong, J.-S.; et al. Toxicity of 100 nm zinc oxide nanoparticles: A report of 90-day repeated oral administration in Sprague Dawley rats. Int. J. Nanomed. 2014, 9, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Bose, K.; Lakshminarasimhan, H.; Sundar, K.; Kathiresan, T. Cytotoxic effect of ZnS nanoparticles on primary mouse retinal pigment epithelial cells. Artif. Cells Nanomed. Biotechnol. 2015, 44, 1764–1773. [Google Scholar] [CrossRef]
- Mirshafiee, V.; Sun, B.; Chang, C.H.; Liao, Y.-P.; Jiang, W.; Jiang, J.; Liu, X.; Wang, X.; Xia, T.; Nel, A.E. Toxicological Profiling of Metal Oxide Nanoparticles in Liver Context Reveals Pyroptosis in Kupffer Cells and Macrophages versus Apoptosis in Hepatocytes. ACS Nano 2018, 12, 3836–3852. [Google Scholar] [CrossRef]
- Hamzian, N.; Hashemi, M.; Ghorbani, M.; Toosi, M.H.B.; Ramezani, M. Preparation, Optimization and Toxicity Evaluation of (SPION-PLGA) ±PEG Nanoparticles Loaded with Gemcitabine as a Multifunctional Nanoparticle for Therapeutic and Diagnostic Applications. Iran. J. Pharm. Res. IJPR 2017, 16, 8–21. [Google Scholar]
- Rueda-Romero, C.; Hernández-Pérez, G.; Ramos-Godínez, P.; Vázquez-López, I.; Quintana-Belmares, R.O.; Huerta-García, E.; Stępień, E.; López-Marure, R.; Montiel-Dávalos, A.; Alfaro-Moreno, E. Titanium dioxide nanoparticles induce the expression of early and late receptors for adhesion molecules on monocytes. Part. Fibre Toxicol. 2015, 13, 1–13. [Google Scholar] [CrossRef]
- Binet, S.; Canu, I.G.; Emond, C.; Fessard, V.; Flahaut, E.; Schroeder, H.; Salles, B.; Sorg, O.; Sparfel, L.; Vasseur, P. Le dioxyde de titane sous forme nanoparticulaire. Valeurs toxicologiques de référence. Le dioxyde de titane sous forme nanoparticulaire. Valeurs toxicologiques de référence. Avis de l’Anses. Collective expert appraisal report. ANSES 2019, 1, 1–120. [Google Scholar]
- Tada-Oikawa, S.; Ichihara, G.; Fukatsu, H.; Shimanuki, Y.; Tanaka, N.; Watanabe, E.; Suzuki, Y.; Murakami, M.; Izuoka, K.; Chang, J.; et al. Titanium Dioxide Particle Type and Concentration Influence the Inflammatory Response in Caco-2 Cells. Int. J. Mol. Sci. 2016, 17, 576. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Zhang, Y.; Qin, H.; Liu, K.; Guo, M.; Ge, Y.; Xu, M.; Sun, Y.; Zheng, X. Cytotoxicity of CdTe quantum dots in human umbilical vein endothelial cells: The involvement of cellular uptake and induction of pro-apoptotic endoplasmic reticulum stress. Int. J. Nanomed. 2016, 11, 529–542. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.-C.; Tsai, T.-L.; Su, W.-P.; Hsieh, H.-P.; Yeh, C.-S.; Shieh, D.-B.; Su, W.-C. Pronounced induction of endoplasmic reticulum stress and tumor suppression by surfactant-free poly(lactic-co-glycolic acid) nanoparticles via modulation of the PI3K signaling pathway. Int. J. Nanomed. 2013, 8, 2689–2707. [Google Scholar] [CrossRef] [PubMed]
- Niu, J.; Azfer, A.; Rogers, L.M.; Wang, X.; Kolattukudy, P.E. Cardioprotective effects of cerium oxide nanoparticles in a transgenic murine model of cardiomyopathy. Cardiovasc. Res. 2007, 73, 549–559. [Google Scholar] [CrossRef]
- Tapeinos, C.; Battaglini, M.; Prato, M.; La Rosa, G.; Scarpellini, A.; Ciofani, G. CeO2 Nanoparticles-Loaded pH-Responsive Microparticles with Antitumoral Properties as Therapeutic Modulators for Osteosarcoma. ACS Omega 2018, 3, 8952–8962. [Google Scholar] [CrossRef]
- Yan, M.; Zhang, Z.; Cui, S.; Lei, M.; Zeng, K.; Liao, Y.; Chu, W.; Deng, Y.; Zhao, C. Improvement of pharmacokinetic and antitumor activity of layered double hydroxide nanoparticles by coating with PEGylated phospholipid membrane. Int. J. Nanomed. 2014, 9, 4867. [Google Scholar]
- Meng, J.; Liang, X.; Chen, X.; Zhao, Y. Biological characterizations of [Gd@C82(OH)22]n nanoparticles as fullerene derivatives for cancer therapy. Integr. Biol. 2012, 5, 43–47. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Z.; Gou, Y.; Qin, Y.; Xu, Y.; Liu, J.; Wu, J.-Z. Apoptosis and necrosis induced by novel realgar quantum dots in human endometrial cancer cells via endoplasmic reticulum stress signaling pathway. Int. J. Nanomed. 2015, 10, 5505–5512. [Google Scholar] [CrossRef]
- Wang, Y.; Kaur, G.; Chen, Y.; Santos, A.; Losic, D.; Evdokiou, A. Bioinert Anodic Alumina Nanotubes for Targeting of Endoplasmic Reticulum Stress and Autophagic Signaling: A Combinatorial Nanotube-Based Drug Delivery System for Enhancing Cancer Therapy. ACS Appl. Mater. Interfaces 2015, 7, 27140–27151. [Google Scholar] [CrossRef]
- Wang, Y.; Kaur, G.; Zysk, A.; Liapis, V.; Hay, S.; Santos, A.; Losic, D.; Evdokiou, A. Systematic in vitro nanotoxicity study on anodic alumina nanotubes with engineered aspect ratio: Understanding nanotoxicity by a nanomaterial model. Biomaterials 2015, 46, 117–130. [Google Scholar] [CrossRef]
- Chen, R.; Ling, D.; Zhao, L.; Wang, S.; Liu, Y.; Bai, R.; Baik, S.; Zhao, Y.; Chen, C.; Hyeon, T. Parallel Comparative Studies on Mouse Toxicity of Oxide Nanoparticle- and Gadolinium-Based T1 MRI Contrast Agents. ACS Nano 2015, 9, 12425–12435. [Google Scholar] [CrossRef] [PubMed]
- Chiu, H.-W.; Xia, T.; Lee, Y.-H.; Chen, C.-W.; Tsai, J.-C.; Wang, Y.-J. Cationic polystyrene nanospheres induce autophagic cell death through the induction of endoplasmic reticulum stress. Nanoscale 2015, 7, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Park, M.R.; Gurunathan, S.; Choi, Y.J.; Kwon, D.N.; Han, J.W.; Cho, S.G.; Park, C.; Seo, H.G.; Kim, J.H. Chitosan nanoparticles cause pre-and post-implantation embryo complications in mice. Biol. Reprod. 2013, 88, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Nasery, M.M.; Abadi, B.; Poormoghadam, D.; Zarrabi, A.; Keyhanvar, P.; Khanbabaei, H.; Ashrafizadeh, M.; Mohammadinejad, R.; Tavakol, S.; Sethi, G. Curcumin Delivery Mediated by Bio-Based Nanoparticles: A Review. Molecules 2020, 25, 689. [Google Scholar] [CrossRef] [PubMed]



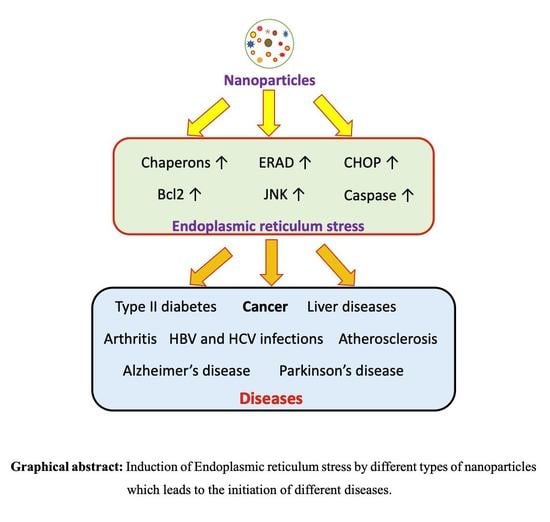
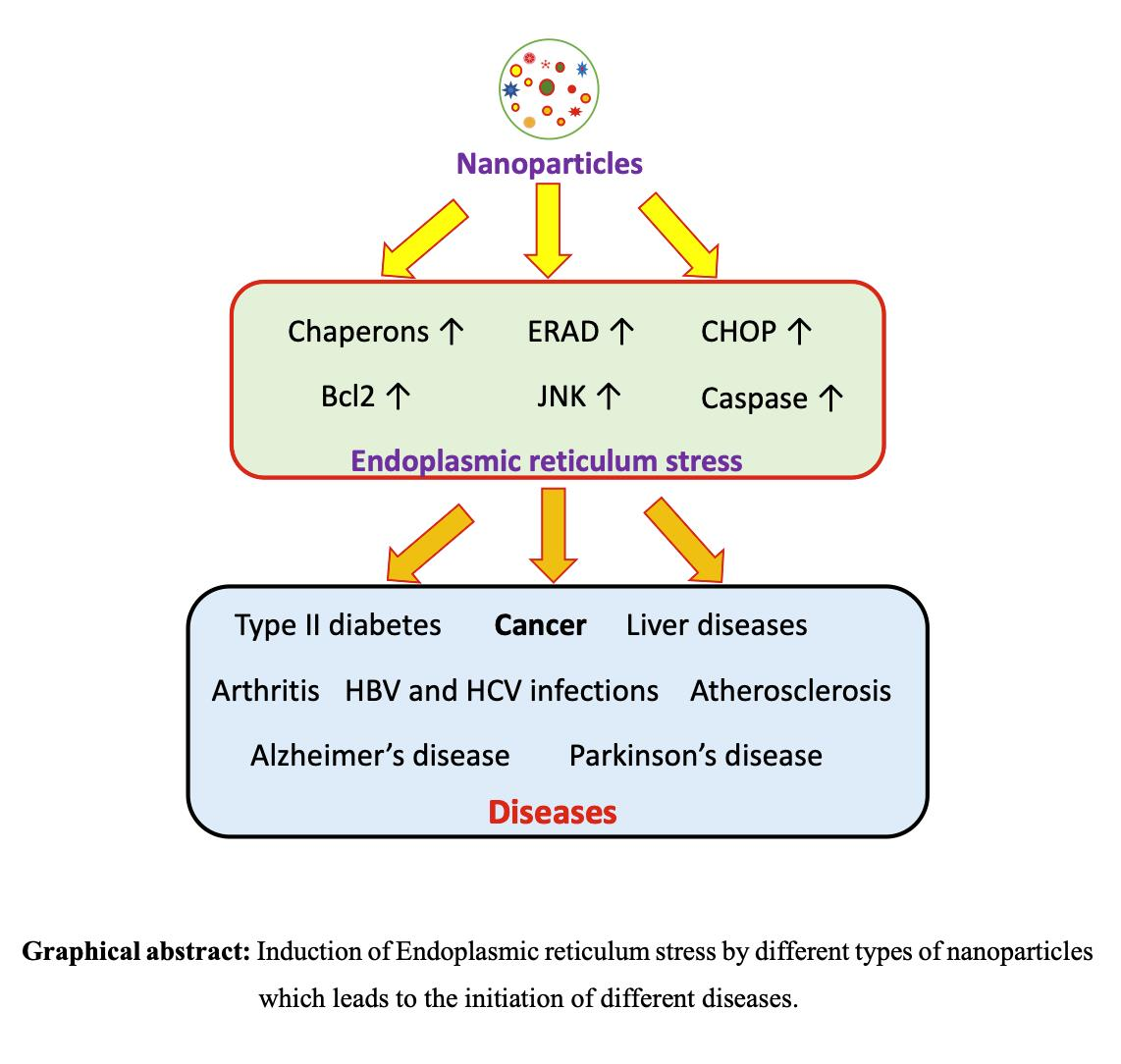{kind=link}
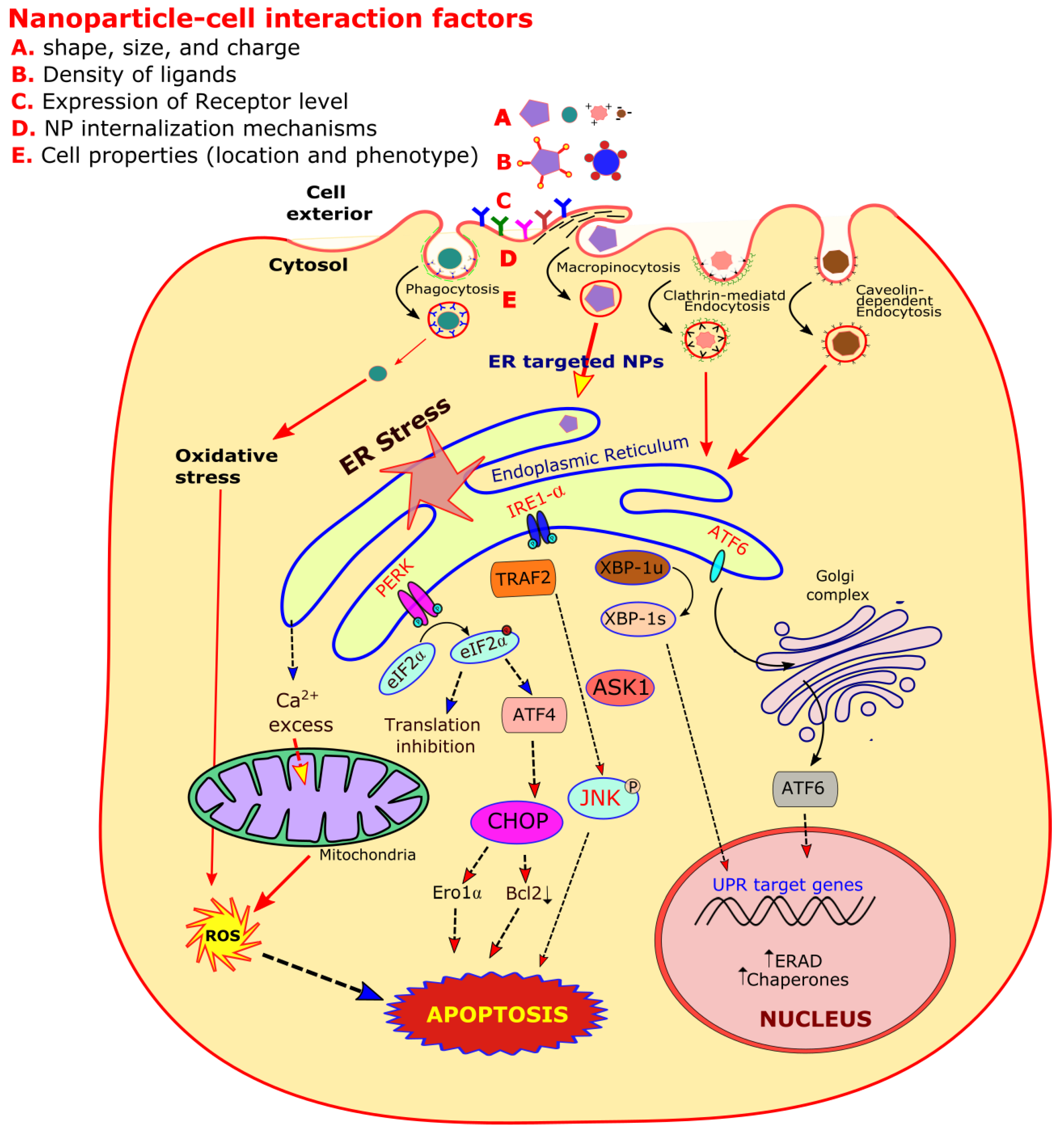{kind=link}
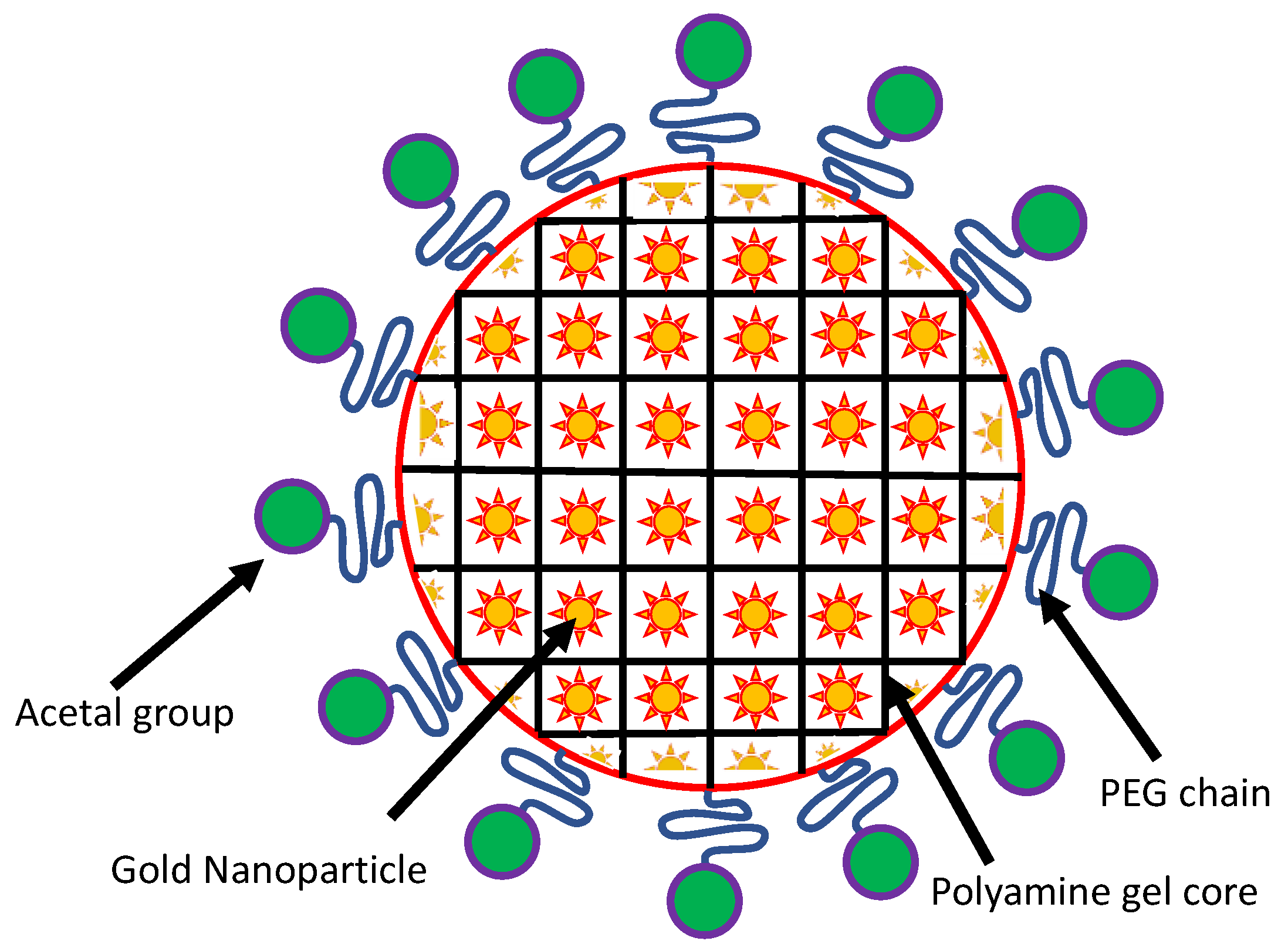{kind=link}
| S. No. | Disease | Role of ER Stress | References |
|---|---|---|---|
| 1. | Type 2 diabetes | ER stress is induced by obesity, gluconeogenesis gets affected by ATF6. The pancreatic β-cell death is induced by hyperglycemia and free fatty acids through CHOP signal | [32] |
| 2. | Parkinson’s disease | Substrate of Parkinson accumulation in ER, leads to ER stress | [33] |
| 3. | Alzheimer’s disease | The CHOP cascade gets activated by mutant presenilin | [34] |
| 4. | Atherosclerosis | Smooth muscle and endothelial cell death is mediated by hyperhomocysteinemia, oxidized phospholipids and cholesterol loading which leads to CHOP cascade. Macrophage death is induced by CHOP cascade which is generated by atherosclerosis related stimuli | [35] |
| 5. | Alcoholic liver disease | The induction of GRP78 and CHOP occurs by alcohol consumption | [36] |
| 6. | Non-alcoholic fatty liver disease | SREBP-1c is induced by ER stress | [37] |
| 7. | HBV and HCV infection | GRP78 and GRP 94 are induced by HBV, while IRE1-XBP1 pathway is suppressed by HCV | [38] |
| 8. | Amyotrophic lateral sclerosis | ER stress is activated by mutant SOD1 aggregates | [39] |
| 9. | Cancer | Different cancers lead to the induction of GRP78 and XBP1 | [40] |
| 10. | Ovarian cancer | The ovarian cancer patients have increased expression of GRP78 | [41] |
| 11. | Liver cancer | In human hepatocellular carcinoma tissues and human hepatocellular carcinoma cells (SMMC7721), the enhanced expression of Grp78 promotes the invasion of hepatocellular carcinoma both in vivo and in vitro | [42] |
| 12. | Prostate cancer | The hormone-resistant prostate cancer cells promote the enhanced expression of Grp78 to the cell surface, which can be further elevated by ER stress in human prostate adenocarcinoma hormone-resistant cells, C4-2B | [43] |
| 13. | Lymphoma | The splicing of XBP1 stimulates tumor growth during hypoxic conditions in such patients | [44] |
| 14. | Breast cancer | In human breast cancer tissue and breast carcinoma cell lines (MDA-MB-231, MCF-7, HCC1500 HS578T), and cells show elevated levels of mRNA and protein Bip/Grp78 | [45] |
| During hypoxia, increased level of ATF4 occurs in MCF7 cell lines | [46] | ||
| Increased expression of unspliced XBP1 mRNA favoring apoptosis of cancer cells and higher levels of spliced XBP1 mRNA increasing cancer survival in human breast cancer tissue | [47] | ||
| The Hormone-resistant breast cancer cells stimulate Grp78 expression on the cell surface, which is further elevated by ER stress in human breast cancer hormone-resistant cells and MCF-7/BUS-10 cell lines | [43] | ||
| 15. | Colorectal cancer | Increased expression of ATF4 occurs during severe hypoxia in HT29 cells | [48] |
| Increased expression of Grp78 on CRC cell surfaces, which promotes CRC cell migration and invasion In human colon carcinoma (SW480, HT29, DLD1, SW620, and Lovo) cell lines | [49] | ||
| 16. | Brain and central nervous system tumor | Enhanced expression of Grp78 is observed in human brain tumor specimens and glioma cell lines (U87, A172, U251, LNZ308, LN-229 and LN-443) | [50] |
| The depletion of XBP-1 dramatically sensitizes U373 cells to viral oncolysis in U373 glioblastoma cells | [51] | ||
| The Inhibition of IRE1α enhances the oncolytic therapy in glioblastoma patient samples | [51] | ||
| 17. | Pancreatic cancer | The expression of PERK supports the proliferation of beta-cell insulinoma and promotes angiogenesis in human tumor xenograft mice | [52] |
| Nanoparticles | Toxicological Effects | Therapeutic Effects |
|---|---|---|
| Aluminum oxide | These NPs used as 1–10 μM in HBMVECs, decreased cell viability, decreased mitochondrial functions, and increased oxidative stress [90] | These NPs were used to penetrated Candida cells, which disrupted the morphological and physiological activity of these cells [91] |
| These NPs 50–80 nm in diameter, were used in mammalian cells EZ4U, caused no significant toxic effect on cell viability [92] | These NPs showed good antibacterial, activity against clinical isolates of P. aeruginosa was significant [93] | |
| The NP were used as 160 nm in size in HMSC, caused decreased cell viability [94] | The NPs were effective against gram-positive and gram-negative bacteria [95] | |
| The NPs were used to check rat blood cells comet micronuclei, caused dose-dependent genotoxicity [96] | The NPs were used as anti-cancer therapy, cytotoxic agents to induce cell death in human prostate cancer cells, and for boosting the efficacy of cancer vaccines [97] | |
| These NPs (50 nm) were used as 0–5000 μg/mL to induce comet DNA damage [98] | The NPs were used as leishmania vaccine to induce autophagy in macrophages and as potent vaccination adjuvant [99] | |
| Copper oxide | These NPs were used in human lung epithelial cells, caused decreased cell viability, increased LDH expression and enhanced lipid peroxidation [100] | The NPs were used in MCF-7 breast cancer cell lines for their cytotoxic effect therapeutics [101] |
| These NPs were used as 0.002–0.2 μg/mL in lung cancer cells, lead to decreased cell viability [102] | These NPs caused skin cancer cells A-375 apoptotic cell death by the activation of caspase-9 [103] | |
| Gold | The NPs caused enhanced lipid peroxidation, oxidative damage and upregulation of stress response genes, and protein expression [104] | The NPs caused the Inhibition of angiogenesis and acted as promising candidates for the drug delivery systems and in cancer therapeutics [105] |
| These NPs induced a significant toxicity, effectively entered the cytoplasm and nucleus, leading to the damage of cellular and nuclear membranes [106] | These NPs were used for the treatment of rheumatic diseases including juvenile arthritis, psoriasis, palindromic rheumatism, and discoid lupus Erythematosus [107] | |
| As 5-nm size, induced the cytotoxicity at 50 mM, whereas no toxicity was observed when used as 15-nm NPs. This shows the size-dependent toxicity of NPs [108] | These NPs increased the apoptosis in B-chronic lymphocytic leukemia (BCLL) treatment-suffering patients [109] | |
| The chitosan-functionalized AuNPs induced the cytotoxicity and pro-inflammatory responses. This indicates the charge-dependent toxicity [110] | The NPs were used as surface coating for different biomedicine applications such as dressing fabrics, implants, glass surfaces [111] | |
| Silica | These NPs used as 10–100 μg/mL in human bronchoalveolar carcinoma cells showed enhanced ROS production, increased LDH expression and higher malondialdehyde formation [112] | As silica-gold nanoshells and gold nanorods were used for tissue stimulating phantoms during photothermal therapy [113] |
| These NPs were used in hepatocellular carcinoma cells (HepG2), showed enhanced ROS production and mitochondrial damage due to increased oxidative stress [114] | The Silica-gold nanoshells were used in human breast carcinoma cells (in vitro) and transmissible venereal tumor (in vivo) as a photothermal therapy agents [115] | |
| Silver | These NPs were used in BRL 3A cell lines and resulted in decreased cell viability, increased LDH expression due to enhanced ROS production [116] | These NPs capped with polyvinyl-pyrrolidone encapsulated in polymer Nanoparticles Human Glioblastoma. Astrocytoma epithelial-like Cell line (U87MG) (in vitro); Swiss mice and severe combined immunodeficiency mice bearing U87MG tumors (in vivo) Therapeutic Evaluation [117] |
| These NPs were used as 0–20 μg/mL in human alveolar cell lines, resulted in decreased cell viability due to increased ROS production [118] | The silver nano-shell with a carbon core were used in prostate adenocarcinoma cell line model as photothermal ablation or radiation enhanced therapy [119] | |
| These NPs (20–40 nm) in size were used in human leukemia cell line WST-1 and resulted in decreased cell viability and the increased expression of LDH [120] | These NPs acted as the excellent candidates for bioimaging and act as good anticancer agents [121] | |
| Zinc oxide | These NPs were used in human colon carcinoma cells, resulted in increased oxidative stress, decreased cell viability and the expression of more inflammatory biomarkers [122] | These NPs when used in murine cell lines showed cytotoxic effects [123] |
| Larger NPs (307–419 nm) in size were used in in human cervix carcinoma cell line (HEp-2), enhanced the DNA damage and decreased the cell viability [124] | These NPs were regarded as a possible treatment for cancer and autoimmune diseases and were found to be involved in specific killing of cancer cells and lead to the activation of human T cells [125] | |
| These NPs resulted in decreased cell viability due to increased DNA damage and increased ROS production and leading to apoptosis [126] | These NPs were used in bioimaging, drug delivery, gene delivery, and as zinc-based biosensors [127] | |
| These NPs were used in human hepatocytes HEK 293 cell line, reduced cell viability, increased mitochondrial damage due to higher oxidative stress [128] | These NPs were used to prevents herpes, by stopping the viral entry and infection [129] | |
| These NPs (<20 nm) were used as 100 μg/mL in human bronchial epithelial cells showed decreased cell viability, LDH release due to enhanced oxidative stress [130] | These NPs were used to prevents the helminth infection as it disrupts the electron transport system and inhibiting the ATP production, so stopping the contractile movement of the parasite [131] | |
| Iron oxide | These NPs were used in murine macrophage cells and resulted in decreased cell viability [132] | These NPs as superparamagnetic NPs were coated with silica-gold nanoshells and used in head and neck cancer cell lines and resulted in overexpression of EGFR and were used for photothermal therapy [133] |
| The NPs as (100–150 nm) in sized used as 0.1 mg/mL in human macrophages resulted in decreased cell viability [134] | These NPs were used in prostate cancer, were magnetic field responsive for thermal ablation [135] | |
| These NPs were used in human hepatocellular carcinoma cells resulted in decreased cell viability [136] | These NPs as aminosilane-coated, were used for thermotherapy during brain tumors [137] | |
| The NPs (20 nm) were used as 0.1 mg/mL in rat mesenchymal stem cells resulted in decreased cell viability [138] | These NPs as starch-coated were magnetically guided for mitoxantrone tumor angiogenesis [139] | |
| Titanium oxide | These NPs were used in mouse models, resulted in enhanced DNA damage and resulted in genotoxicity [140] | These NPs were used in CT26 and LL2 mouse cancer to increase oxidative stress [141] |
| These NPs were used as 10–50 μg/mL in human lung cells resulted in enhanced oxidative stress, more DNA adduct formation and increased cytotoxicity [142] | These NPs were used as efficient drug delivery systems and in photodynamic therapy of tumors [143] |
| Nanoparticle | Physicochemical Properties and Related Studies | Specific Role in ER Stress Induction | Application | Reference |
|---|---|---|---|---|
| AgNPs | Typical size of 120 nm, negatively charged ZFL cells (in vitro); exposure on zebrafish (in vivo) 0.05–0.5 mg/mL for 6–24 h (in vitro); 0.1–5 mg/mL for 24 h (in vivo) | Increase in GRP78, ATF6, and XBP-1s protein expression or mRNA synthesis. | ROS induction, ER stress response, apoptotic and inflammatory pathways activation. | [72] |
| Average size 20 nm, negatively charged 16HBE cells (in vitro); mice (in vivo) 2 μg/cm2 exposure from 4 to 24 h (in vitro); 0.1–0.5 μg/g (in vivo) | Increased p-PERK, XBP-1s, p-IRE1α, CHOP, GRP78, p-eIF2α protein expression or mRNA synthesis | Cellular response on different cell lines, to know the mechanisms of action in various cellular systems, cellular activation of different signaling molecules | [244] | |
| 10, 50 and 100 nm used in HepG2 cells as 1 μg/mL dose for 24 h | Increase in CHOP protein | Can be used as cytotoxic agents on mice liver primary cells and also in human liver HepG2 cells | [245] | |
| 15 nm in size, negatively charged in THP-1 cells as 1–25 μg/mL for 1 or 24 h | Increase in p-PERK protein and ATF6 degradation | Redox active particles can induce toxicity mediated through ROS production and increases oxidative stress | [246] | |
| ≤100 nm size, used in Human Chang liver cells as 4 μg/mL for 3–24 h | Increase in ER tracker staining and protein levels of p-IRE1, p-PERK, ATF6, peIF2α, XBP-1s, GRP78, and CHOP | Increased concentrations of these NPs induce substantial cytotoxicity, DNA damage and apoptosis. | [247] | |
| Size 2 nm to 10 nm; negatively charged used in MCF-7 and T-47D cells | Increase in p-eIF2α, p-PERK, CHOP, p-IRE1α, and ATF4 proteins | Possess anti-cancer activity, DOX + AgNPs can induce conformational changes on DNA | [248] | |
| AuNPs | Size about 12 nm and citrate-capped are negatively charged used in HUVECs as 8 μg/mL from 2 to 35 d | Enhanced XBP-1s mRNA production | Accumulates at steady exposure of lower (non-lethal) dose and causes no measurable cell death while leading to elevated ER stress. | [249] |
| Size 20 to 70 nm and negatively charged used in human neutrophils as 100 μg/mL for 3 h | Increased p-IRE, p-PERK, and ATF6 proteins synthesis | PEG-AuNPs can be efficient drug delivery vehicles, and exhibit least adsorption of proteins and slight size and charge deviation when used in whole blood | [250] | |
| 20 nm in size, citrate coated AsPc1 cells | Enhanced IRE-1α and CHOP proteins synthesis | Sensitization of pancreatic cancer cells by the pre-treatment with these NPs in addition to gemcitabine in colony forming and viability assays | [251] | |
| Size of 1–6 nm and 15–20 nm used in K562 cells | Increased the ER stress related proteins checked by proteomic assay | These NPs can be used to diminish the growth and provoke strong apoptosis in human chronic myeloid leukemia cells | [252] | |
| PEGylated nanogel with AuNPs | Used in SCCVII and A549 cells | Increased GRP78, IRE-1α, p-PERK protein synthesis | Favors the radiosensitization of cells to increase the apoptosis and ER stress provoked DNA repair capacity | [253] |
| ZnO NPs | Size about 100 nm used in HUVEC at a dose of 240 μM for 4–24 h | Augmented CHOP, p-PERK, XBP-1s, p-eIF2α, HSP proteins or mRNA production | Activates the ER stress-responsive pathways | [197] |
| Size between 30 nm to 90 nm, bulk 100–200 nm used in mice at 100 mg/kg/d for 3 d | Increase in eIF2a, PERK, ATF4, JNK, CHOP, GRP94 mRNA in livers | It disrupts seminiferous epithelium of the testis and decreases the sperm density in the epididymis | [254] | |
| The size is about 70 nm; positively charged used in MRC5 cells at 25 and 50 μg/mL for 16 h | Increases CHOP and ERN1 mRNA synthesis | Oxidative stress is promoted, which causes cytotoxicity and genotoxicity in human lung fibroblasts in vitro and in D. melanogaster in vivo | [255] | |
| The size is <100 nm and negatively charged used in mice as gavage for 90 d (200, 400 mg/kg) | It causes the swelling of ER; increases GRP 78/94, XBP-1, and PDI-3 mRNA synthesis, CHOP and p-JNK protein production in liver | The relationship of the dosage and organs affected as pancreas, stomach, eye, and prostate gland | [256] | |
| ZnS NPs | Size between 50 and 100 nm used in mice retinal pigment epithelial cells | Inhibited GRP78 and CHOP protein synthesis | Can be used as anti-age-related macular degeneration | [257] |
| Fe3O4 NPs | Hydrodynamic size about 26 nm, negatively charged and used in RAW 264.7 cells as 6.25–50 μg/mL for 24 h | Increase in CHOP mRNA, CHOP, p-IRE1α, IRE1α protein synthesis | Pyroptosis demonstration and IL-1β synthesis, safety evaluation of metal oxides | [258] |
| 15–20 nm in size, PLGA coated about 300 nm in size used in MCF-7 cells as 100 μg/mL for 24 h. | Disrupts and disperse ER | Gemcitabine loaded NPs demonstrate as multifunctional drag cargo system, can be used during radiosensitization investigations | [259] | |
| TiO2 NPs | Size as P25 (24 nm), and scrolled nanosheets (L/W 178/9), nanoneedles (L/W 45/15), isotropic NPs (29 nm) used in HUVECs cells as 2 μg/cm2 for 1–24 h | Increase in ERdj4, CHOP, HERPUD1 mRNA (scrolled nanosheets | Such NPs can be used to increase the ROS production having a central role in the induction of receptor expression | [260] |
| Size as 19.3 ± 5.4 nm, used in mice, inhaled to 2.5, 5.0 and 10.0 mg/m3 NPs for a span of 28 d | Causes the swelling of ER and increases CHOP, GRP78, and p-IRE1α protein synthesis in lungs | Can be used as toxicological index that acts as a benchmark for assessing the risks to human health | [261] | |
| Hydrodynamic size of about 250 nm, the anatase: rutile ratio of 8:2 used in 16HBE14o-lung cells as 50 and 100 μg/mL for 24 and 48 h | Increases the CHOP, GRP78, IRE-1α, and p-IRE-1α protein synthesis | The Anatase TiO2 NPs induces increased inflammatory responses as compared with other TiO2 particles | [262] | |
| Cadmium telluride (CdTe) quantum dots (QDs) (CdTeQDs) | About 4 nm in size and negative charged used in HUVECs cells as 10 μg/mL for 24 h. | Lead to the dilation of ER and protein synthesis increase of GRP78/95, CHOP, ATF4, p-PERK, peIF2α, and p-JNK. | The toxicity of QDs can act as potential cardiovascular risk factors | [263] |
| Poly [lactic-co-glycolic acid] (PLGA) NPs containing γ-oryzanol | Size about 214.8 nm with negative charge used in obese ob/ob mice | It reduced the CHOP, ERdj4, and XBP-1s mRNA synthesis | Can be used during metabolic diseases treatment | [154] |
| Poly [lactic-co-glycolic acid] (PLGA) NPs containing LY294002 | NPs with an average size of 98.9 ± 2.64 nm and used in H157, H460, H1650, and NL20 cells | It leads to the accumulation in ER; increased GRP78, CHOP, and p-JNK proteins | These NPs act as surfactant-free formulation of PLGA and possesses a promising anticancer activity | [264] |
| CeO2 NPs | Average size of 7 nm used in MCP-1 transgenic mice | It suppresses the GRP78, PDI, and HSP mRNA synthesis | These NPs slow down the advancement of cardiac dysfunction myocardial oxidative stress | [265] |
| Used in H9C2 cells | It reduces the PDI and GRP78 proteins synthesis | These NPs are pH responsive with anti-tumoral activities for osteosarcoma | [266] | |
| PEGylated-Phosphatidyl ethanolamine (PE) micelles | Used in MRC-5, A549, 293T cells for ER dilation | Leads to increased IRE-1α, eIF2α, PERK, ATF4/6, XBP-1s and CHOP proteins synthesis in cancer cells | It enhances the sensitivity of most cancer cells to some chemotherapeutic agents | [267] |
| Gadolinium metallofullerenol [Gd@C82(OH)22]n NPs | Used in MCF-7 and ECV304 cells | It lead to slowed protein processing in ER and also increased the CHOP mRNA synthesis as reported by DNA microarray | These NPs possess high anti-tumor activity but have low toxicity | [268] |
| Realgar QDs | These NPs have an average size of 5.48 nm and used in JEC cells | It induces the dilation of ER and increased CHOP and GRP78 mRNA and proteins synthesis | Can be used effectively against human endometrial cancer cells as it leads to ER stress mediated necrosis and apoptotic cell death | [269] |
| Anodic Alumina Nanotubes (AANTs) loaded with Thapsigargin (TG) | The length is 736 nm ± 460 nm, inner diameter and outer diameter as 33.0 ± 8.0 and 90.0 ± 10.0 nm used in THP-1, HFF, and MDA-MB 231-TXSA cells | It led to increase in IRE1α and GRP78 proteins synthesis and ER tracker staining | It can act as novel biomaterials for clinical cancer therapy as it can act as ER and autophagic delivery systems | [270] |
| Anodic Alumina Nanotubes (AANTs) | It has the aspect ratio of 7.8 (short), 27.7 (medium) and 63.3 (long) used in MDA-MB-231-TXSA and RAW264.7 cells as 100 μg/mL AANTs for 3d. | It led to increased CHOP protein synthesis and ER-tracker staining and decreased IRE1α protein synthesis as reported by long AANT only. | For the advanced drug delivery applications, it has a promising opportunity as it can control the nanotoxicity of high aspect ratio nanomaterials | [271] |
| Extremely small size iron oxide NPs (ESION) and MnONPs | About 3 nm (ESION), and 15 nm (MnONPs) used in mice as 2, 5, 10 μg/g for 1 d | Enhanced the expression of CHOP, HSP, GRP78, XBP-1s mRNA or protein in various organs | NPs exposure causes bodyweight loss, increased NO and MDA levels, inflammatory and hyperplastic changes in the lung homogenates | [272] |
| NH2-labelled Polystyrene (PS) NPs | 60 nm in size, positively charged used in RAW 264.7, BEAS-2B cells as 5–40 μg/mL up to 16 h. | It leads to misfolded protein aggregates; increases ER-tracker staining and IRE1α protein synthesis | These NPs can play an efficient role in autophagy, safe and novel material design and inhibition of the toxicity | [273] |
| Chitosan NPs | Average size of 100 nm used in mouse morula-stage embryos as 100 μg/mL for 24–28 h | It leads to increase in GRP78, CHOP, ATF4, PERK, IRE-1α, protein or mRNA synthesis | These NPs lead to blastocyst complications with no or small cavity | [274] |
| Silica (SiO2) NPs | About 250 nm in size and negatively charged used in Huh7 cells as 0.05–0.5 mg/mL for 4 and 24 h | These NPs increased the GRP78 and XBP-1s mRNA synthesis | These NPs lead to ER stress mediated MAPK pathway, and inflammatory reactions initiation in human hepatoma cells | [197] |
| Polyethyleneimine (PEI) NPs | Used in Neuro2A cells as 3–25 μg/mL for 24 h | It causes increased GRP78, ATF4 and CHOP mRNA synthesis | These NPs cause Neuro2A cells induced cell toxicity in a concentration-dependent manner | [81] |
| Curcumin NPs | Average size of 50 nm and negative charged used in H9C2 cells | These NPs cause suppression of GRP78 and CHOP proteins | These NPs can prevent myocardial injury | [275] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, A.A.; Allemailem, K.S.; Almatroudi, A.; Almatroodi, S.A.; Mahzari, A.; Alsahli, M.A.; Rahmani, A.H. Endoplasmic Reticulum Stress Provocation by Different Nanoparticles: An Innovative Approach to Manage the Cancer and Other Common Diseases. Molecules 2020, 25, 5336. https://doi.org/10.3390/molecules25225336
Khan AA, Allemailem KS, Almatroudi A, Almatroodi SA, Mahzari A, Alsahli MA, Rahmani AH. Endoplasmic Reticulum Stress Provocation by Different Nanoparticles: An Innovative Approach to Manage the Cancer and Other Common Diseases. Molecules. 2020; 25(22):5336. https://doi.org/10.3390/molecules25225336
Chicago/Turabian StyleKhan, Amjad Ali, Khaled S. Allemailem, Ahmad Almatroudi, Saleh A. Almatroodi, Ali Mahzari, Mohammed A. Alsahli, and Arshad Husain Rahmani. 2020. "Endoplasmic Reticulum Stress Provocation by Different Nanoparticles: An Innovative Approach to Manage the Cancer and Other Common Diseases" Molecules 25, no. 22: 5336. https://doi.org/10.3390/molecules25225336
APA StyleKhan, A. A., Allemailem, K. S., Almatroudi, A., Almatroodi, S. A., Mahzari, A., Alsahli, M. A., & Rahmani, A. H. (2020). Endoplasmic Reticulum Stress Provocation by Different Nanoparticles: An Innovative Approach to Manage the Cancer and Other Common Diseases. Molecules, 25(22), 5336. https://doi.org/10.3390/molecules25225336