
Significance of Targeting VEGFR-2 and Cyclin D1 in Luminal-A Breast Cancer


 ,
, 
Abstract

1. Introduction
2. Results
2.1. Characteristics of Clinical Samples and Ethical Clearance
2.2. Kinase Activity in Luminal-A Breast Tumors
2.2.1. Network Analysis
2.2.2. Process Network
2.2.3. GO Processes
2.3. Combination Cytotoxicity and Selectivity Studies
2.4. Clonogenicity of the Three Compounds and their Combinations
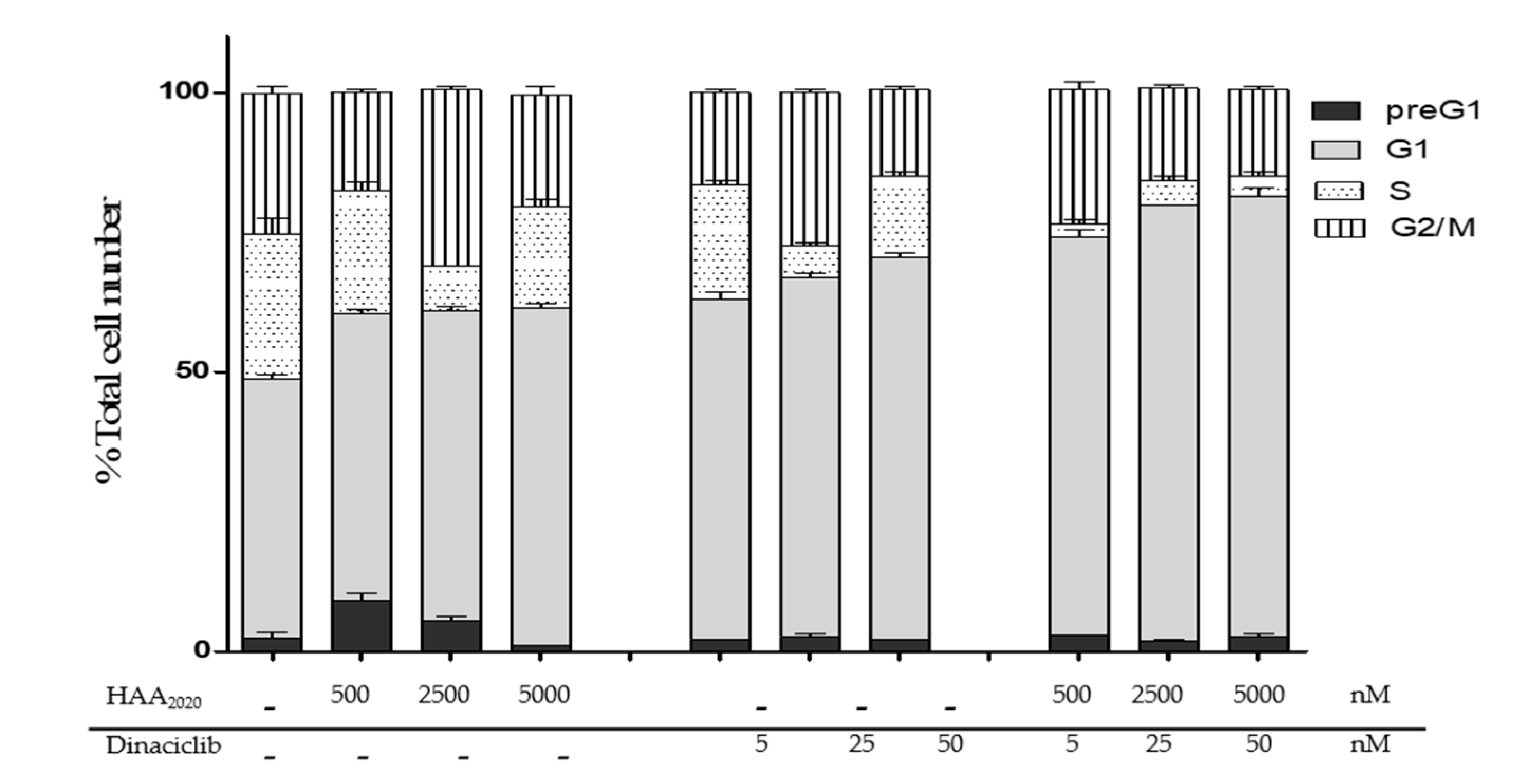
2.5. Cell Cycle Analysis
2.6. Detection of Apoptosis
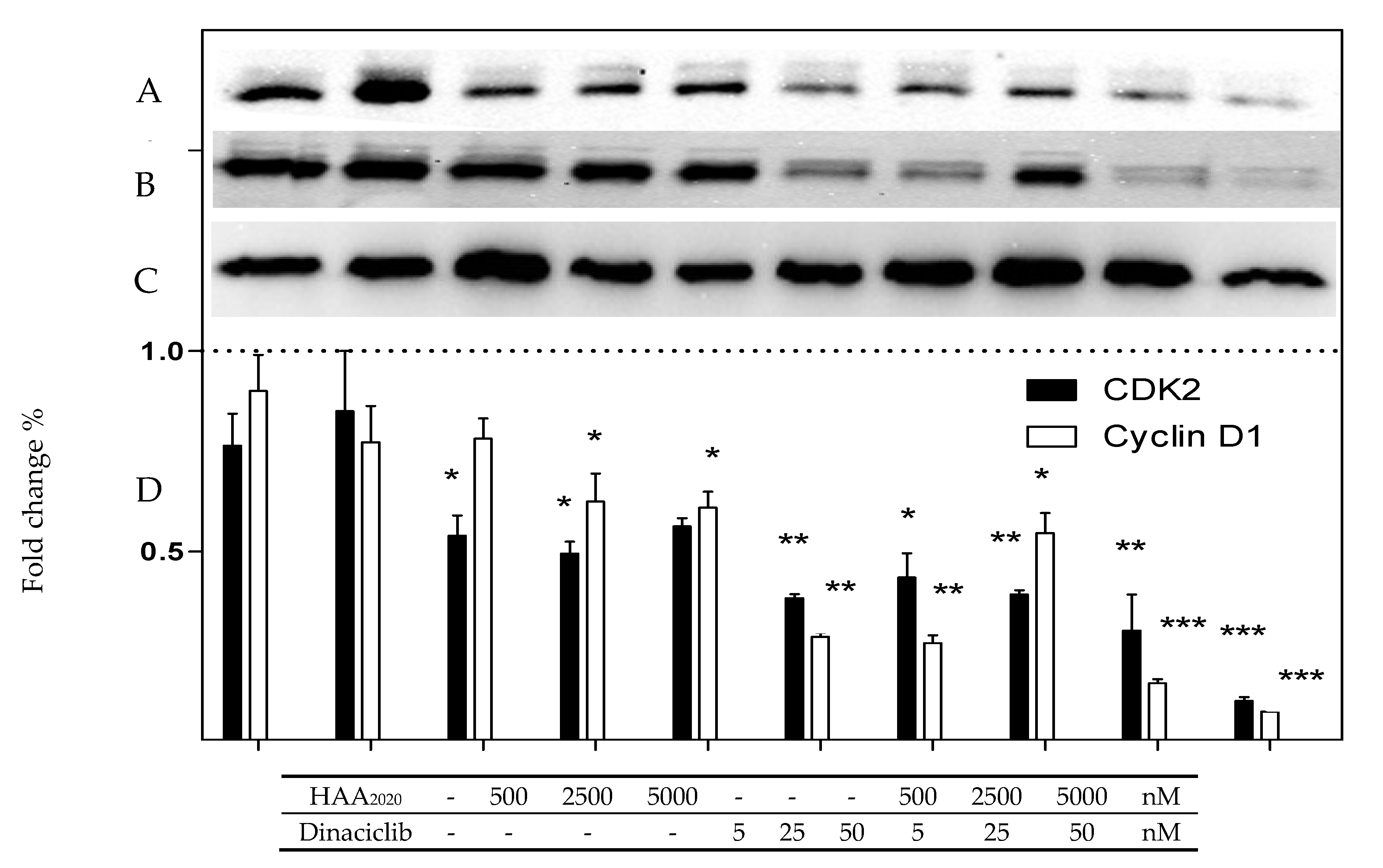
2.7. Western Blotting of CDK2 and Cyclin D 1 Proteins
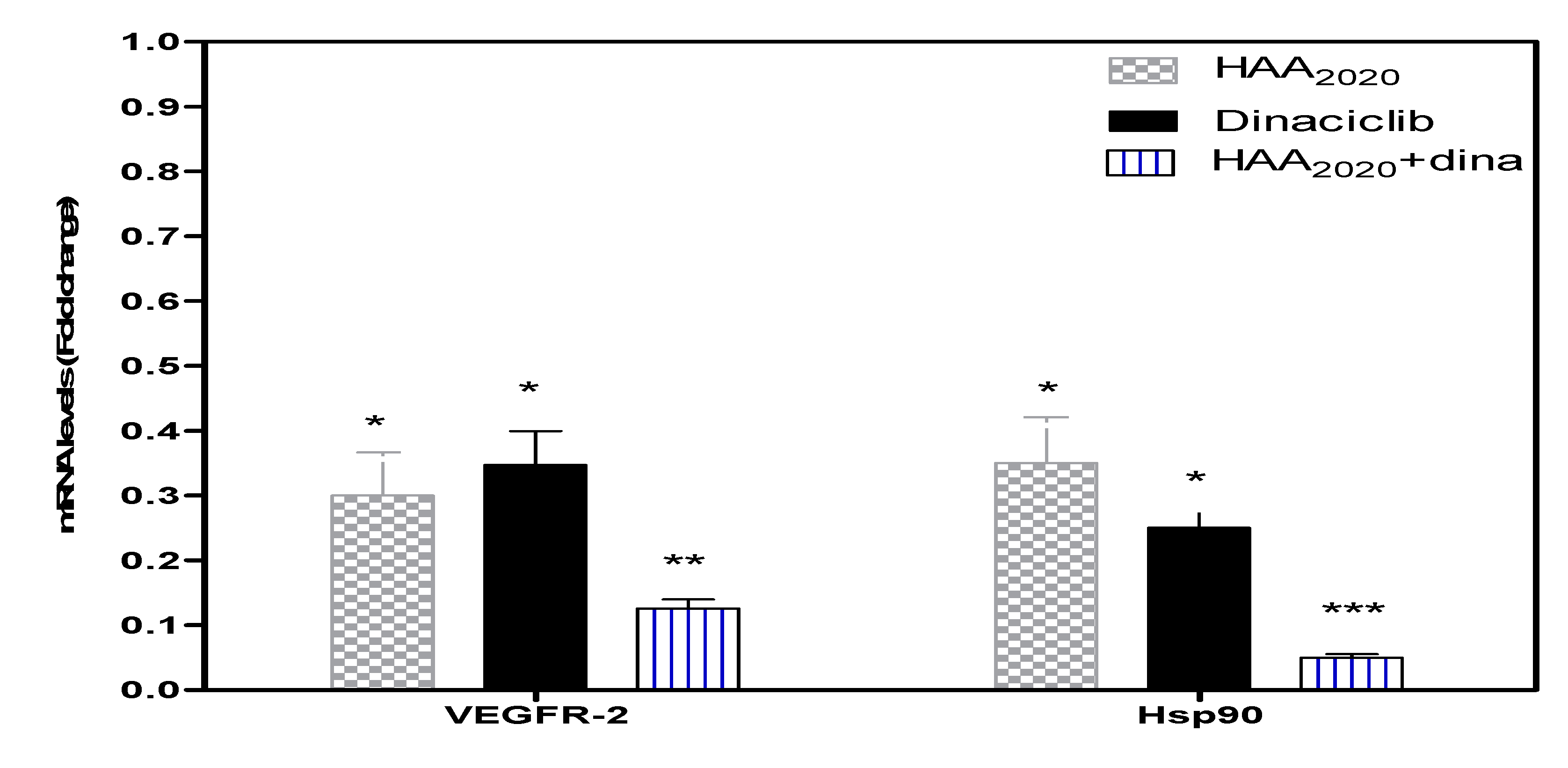
2.8. Real-Time PCR
2.9. Immunofluorescent Imaging
3. Discussion
4. Materials and Methods
4.1. Collection, Preparation of Clinical Samples, and Ethical Clearance
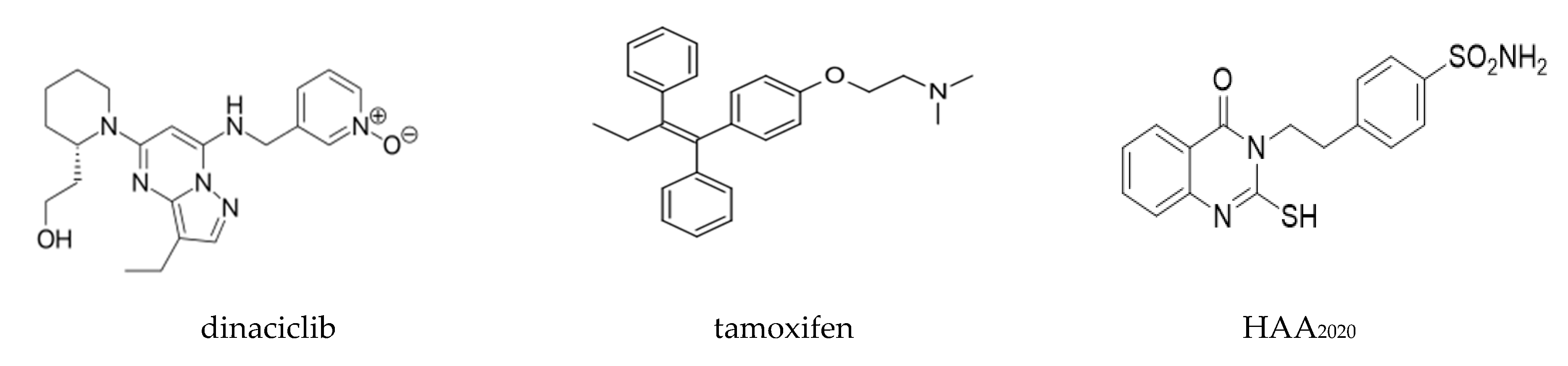
4.2. Compounds and Reagents
4.3. Cell Culture
4.4. PamChip Peptide Microarray-Based Kinase Activity Profiling Arrays
Data Interpretation
4.5. Cytotoxicity and Combination Studies
4.6. Clonogenic Survival Assay
4.7. Cell Cycle Analysis
4.8. Determination of Apoptosis
4.9. Western Blotting
4.10. Real-Time PCR
4.11. Immunofluorescence Staining
4.12. Statistics
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| 17-AAD | 17-N-allylamino-17-demethoxygeldanamycin |
| AIs | Aromatase inhibitors |
| AIB1 | Amplified in breast cancer 1 |
| ATM | Ataxia Telangiectasia Mutated |
| BC | Breast cancer |
| CDK | Cyclin dependent kinases |
| CI | Combination index |
| CREB5 | CAMP Responsive Element Binding Protein 5 |
| CREB3L4 | CAMP Responsive Element Binding Protein 3 Like 4 |
| DMSO | Dimethyl Sulfoxide |
| EFGR | Epidermal growth factor receptor |
| ErbB2 | Erb-B2 Receptor Tyrosine Kinase 2 |
| ER | Estrogen receptor |
| ESR1 | Estrogen receptor 1 |
| FAK1 | Focal adhesion kinase 1 |
| FITC | Fluorescein isothiocyanate |
| FOXM1 | Forkhead box protein M1 |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| HAA2020 | compound 1 (4-(2-(4-Oxo-2-thioxo-1,4-dihydroquinazolin-3(2H)-yl)ethyl) benzene sulfonamide) |
| Her2 | Human epidermal growth factor receptor 2 |
| Hsp90 | Heat shock protein 90 |
| Hsp90i | Heat shock protein 90 inhibitors |
| HR | Hormone receptor |
| IGFBP2 | Insulin-like growth factor (IGF) binding protein 2 |
| MAPK | Mitogen activated protein kinase |
| MRAS | Muscle RAS oncogene |
| PA | Pathway analysis |
| PCNA | Proliferating cell nuclear antigen |
| PI | Propidium iodide |
| PI3K/AKT | Phosphoinositide-3-kinases/Protein kinase B |
| PIK3C2G | Phosphatidylinositol-4-Phosphate 3 Kinase Catalytic Subunit Type 2 Gamma |
| PKC | Protein kinase C |
| PLC gamma 1 | Phospholipase C, gamma 1 |
| RhoA | Ras homolog family member A |
| RoCK-1 | Rho Associated Coiled-Coil Containing Protein Kinase 1 |
| PR | Progestrone receptor |
| RT-PCR | Reverse transcription polymerase chain reaction |
| SERDs | Selective estrogen receptor degraders |
| SERMs | Selective estrogen receptor modulators |
| SI | Selectivity index |
| SRC | Proto-oncogene tyrosine-protein kinase |
| TKI | Tyrosine kinase inhibitors |
| TRAM-1 | Translocation Associated Membrane Protein 1 |
| VEGF | Vascular endothelial growth factor |
| VEGFR | Vascular endothelial growth factor receptor |
| VEGFR-2 | Vascular endothelial growth factor receptor-2 |
References
- Bray, F.; Me, J.F.; Soerjomataram, I.; Siegel, R.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Al-Shahrani, Z.; Al-Rawaji, A.; Al-Madouj, A.N.; Hayder, M.S.; Al-Zahrani, A.; Al-Mutlaq, H.; Bazarbashi, S.M. Cancer Incidence Report Saudi Arabia 2014. National Health Information Center: Riyadh, Saudi Arabia, 2017. Available online: https://nhic.gov.sa/eServices/Documents/2014.pdf (accessed on 1 September 2020).
- American Cancer Society. Breast Cancer Facts & Figures 2019–2020; American Cancer Society: Atlanta, GA, USA, 2019; pp. 1–44. [Google Scholar]
- Alnegheimish, N.A.; Alshatwi, R.A.; Alhefdhi, R.M.; Arafah, M.M.; AlRikabi, A.C.; Husain, S. Molecular subtypes of breast carcinoma in Saudi Arabia. Saudi Med. J. 2016, 37, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Shawarby, M.; Al-Tamimi, D.; Ahmed, A. Molecular classification of breast cancer: An overview with emphasis on ethnic variations and future perspectives. Saudi J. Med. Med. Sci. 2013, 1, 14. [Google Scholar] [CrossRef]
- Mestres, J.A.; Imolins, A.B.; Martínez, L.C.; López-Muñiz, J.I.C.; Gil, E.C.; Ferré, A.D.J.; Berrón, S.D.B.; Pérez, Y.F.; Mata, J.G.; Palomo, A.G.; et al. Defining the optimal sequence for the systemic treatment of metastatic breast cancer. Clin. Transl. Oncol. 2016, 19, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, K.E.H.; Elamin, A. Adherence to endocrine therapy and its relation to disease-free survival among breast cancer patients visiting an out-patient clinic at Khartoum Oncology Hospital, Sudan. J. Evaluation Clin. Pr. 2020. [Google Scholar] [CrossRef]
- Shagufta; Ahmad, I. Tamoxifen a pioneering drug: An update on the therapeutic potential of tamoxifen derivatives. Eur. J. Med. Chem. 2018, 143, 515–531. [Google Scholar] [CrossRef] [PubMed]
- Ximenez, J.P.B.; De Andrade, J.M.; Marques, M.P.; Coelho, E.B.; Suarez-Kurtz, G.; Lanchote, V.L. Hormonal status affects plasma exposure of tamoxifen and its main metabolites in tamoxifen-treated breast cancer patients. BMC Pharmacol. Toxicol. 2019, 20, 1–10. [Google Scholar] [CrossRef]
- Clemons, M.; Danson, S.; Howell, A. Tamoxifen (‘Nolvadex’): A review. Cancer Treat. Rev. 2002, 28, 165–180. [Google Scholar] [CrossRef]
- Chang, M. Tamoxifen resistance in breast cancer. Biomol. Ther. 2012, 20, 256–267. [Google Scholar] [CrossRef]
- Haque, M.; Desai, K.V. Pathways to Endocrine Therapy Resistance in Breast Cancer. Front. Endocrinol. 2019, 10, 573. [Google Scholar] [CrossRef]
- Munzone, E.; Curigliano, G.; Rocca, A.; Bonizzi, G.; Renne, G.; Goldhirsch, A.; Nole, F. Reverting estrogen-receptor-negative phenotype in HER-2-overexpressing advanced breast cancer patients exposed to trastuzumab plus chemotherapy. Breast Cancer Res. 2006, 8, R1–R4. [Google Scholar] [CrossRef]
- Fan, P.; Wang, J.; Santen, R.J.; Yue, W. Long-term Treatment with Tamoxifen Facilitates Translocation of Estrogen Receptor α out of the Nucleus and Enhances its Interaction with EGFR in MCF-7 Breast Cancer Cells. Cancer Res. 2007, 67, 1352–1360. [Google Scholar] [CrossRef] [PubMed]
- García-Campos, M.A.; Espinal-Enríquez, J.; Hernández-Lemus, E. Pathway Analysis: State of the Art. Front. Physiol. 2015, 6, 383. [Google Scholar] [CrossRef]
- Vishnubalaji, R.; Nair, V.S.; Ouararhni, K.; Elkord, E.; Alajez, N.M. Integrated Transcriptome and Pathway Analyses Revealed Multiple Activated Pathways in Breast Cancer. Front. Oncol. 2019, 9, 910. [Google Scholar] [CrossRef] [PubMed]
- Karim, S.; Merdad, A.; Schulten, H.-J.; Jayapal, M.; Dallol, A.; Buhmeida, A.; Al-Thubaity, F.; Mirza, Z.; Gari, M.A.; Chaudhary, A.G.; et al. Low expression of leptin and its association with breast cancer: A transcriptomic study. Oncol. Rep. 2016, 36, 43–48. [Google Scholar] [CrossRef]
- Merdad, A.; Karim, S.; Schulten, H.-J.; Jayapal, M.; Dallol, A.; Buhmeida, A.; Al-Thubaity, F.; A GariI, M.; Chaudhary, A.G.; Abuzenadah, A.; et al. Transcriptomics profiling study of breast cancer from Kingdom of Saudi Arabia revealed altered expression of Adiponectin and Fatty Acid Binding Protein4: Is lipid metabolism associated with breast cancer? BMC Genom. 2015, 16, S11. [Google Scholar] [CrossRef]
- Hilhorst, M.H.; Houkes, L.; Korsten, H.; Mommersteeg, M.; Trapman, J.; Ruijtenbeek, R. Abstract 4046: Direct detection of AKT/PKB activity in a Pten knock out mouse model using dynamic peptide microarrays. In Cellular and Molecular Biology; American Association for Cancer Research (AACR): Philadelphia, PA, USA, 2011; Volume 71, p. 4046. [Google Scholar]
- Chirumamilla, C.S.; Fazil, M.H.U.T.; Perez-Novo, C.; Rangarajan, S.; De Wijn, R.; Ramireddy, P.; Verma, N.K.; Berghe, W.V. Profiling Activity of Cellular Kinases in Migrating T-Cells. Adv. Str. Safety Stu. 2019, 99–113. [Google Scholar] [CrossRef]
- Hilhorst, R.; Houkes, L.; Mommersteeg, M.; Musch, J.; Berg, A.V.D.; Ruijtenbeek, R. Peptide Microarrays for Profiling of Serine/Threonine Kinase Activity of Recombinant Kinases and Lysates of Cells and Tissue Samples. In Advanced Structural Safety Studies; Springer Science and Business Media LLC: New York, NY, USA, 2013; Volume 977, pp. 259–271. [Google Scholar]
- Alkahtani, H.M.; Abdalla, A.N.; Obaidullah, A.J.; Alanazi, M.M.; Almehizia, A.A.; Alanazi, M.G.; Ahmed, A.Y.; Alwassil, O.I.; Darwish, H.W.; Abdel-Aziz, A.A.-M.; et al. Synthesis, cytotoxic evaluation, and molecular docking studies of novel quinazoline derivatives with benzenesulfonamide and anilide tails: Dual inhibitors of EGFR/HER2. Bioorg. Chem. 2020, 95, 103461. [Google Scholar] [CrossRef]
- Abdalla, A.N.; Abdallah, M.E.; Aslam, A.; Bader, A.; Vassallo, A.; De Tommasi, N.; Almalki, W.H.; Gouda, A.M.; Mukhtar, M.H.; El-Readi, M.Z.; et al. Synergistic Anti Leukemia Effect of a Novel Hsp90 and a Pan Cyclin Dependent Kinase Inhibitors. Molecules 2020, 25, 2220. [Google Scholar] [CrossRef]
- Comunanza, V.; Bussolino, F. Therapy for Cancer: Strategy of Combining Anti-Angiogenic and Target Therapies. Front. Cell Dev. Biol. 2017, 5, 101. [Google Scholar] [CrossRef]
- Barnabas, O.; Wang, H.; Gao, X.-M. Role of estrogen in angiogenesis in cardiovascular diseases. J. Geriatr. Cardiol. 2013, 10, 377–382. [Google Scholar] [PubMed]
- Zhao, R.; Leung, E.; Gruner, S.; Schapira, M.; Houry, W.A. Tamoxifen Enhances the Hsp90 Molecular Chaperone ATPase Activity. PLoS ONE 2010, 5, e9934. [Google Scholar] [CrossRef]
- Zagouri, F.; Bournakis, E.; Koutsoukos, K.; Papadimitriou, M. Heat Shock Protein 90 (Hsp90) Expression and Breast Cancer. Pharmaceuticals 2012, 5, 1008–1020. [Google Scholar] [CrossRef] [PubMed]
- Yamaki, H.; Nakajima, M.; Shimotohno, K.W.; Tanaka, N. Molecular basis for the actions of Hsp90 inhibitors and cancer therapy. J. Antibiot. 2011, 64, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Prince, T.; Sun, L.; Matts, R.L. Cdk2: A Genuine Protein Kinase Client of Hsp90 and Cdc37. Biochemistry 2005, 44, 15287–15295. [Google Scholar] [CrossRef] [PubMed]
- Zagouri, F.; Sergentanis, T.N.; Chrysikos, D.; Papadimitriou, M.; Dimopoulos, M.-A.; Psaltopoulou, T. Hsp90 inhibitors in breast cancer: A systematic review. Breast 2013, 22, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Lu, M.; Yao, M.; Zhu, W. Effects of treatment with an Hsp90 inhibitor in tumors based on 15 phase II clinical trials. Mol. Clin. Oncol. 2016, 5, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Bedin, M.; Gaben, A.-M.; Mester, J. Geldanamycin, an inhibitor of the chaperone activity of HSP90, induces MAPK-independent cell cycle arrest. Int. J. Cancer 2004, 109, 643–652. [Google Scholar] [CrossRef] [PubMed]
- Shuai, Z.; El-Deiry, W.S. Dual Inhibition of Cdk and Hsp90 Destabilize Hif1alpha and Synergistically Induces Cancer Cell Death. 2018. Available online: https://uspto.report/patent/app/20180243306 (accessed on 1 September 2020).
- Jin, H.; Dan, H.-G.; Rao, G.-W. Research progress in quinazoline derivatives as multi-target tyrosine kinase inhibitors. Heterocycl. Commun. 2018, 24, 1–10. [Google Scholar] [CrossRef]
- Park, H.; Kim, Y.-J.; Hahn, J.-S. A novel class of Hsp90 inhibitors isolated by structure-based virtual screening. Bioorganic Med. Chem. Lett. 2007, 17, 6345–6349. [Google Scholar] [CrossRef]
- Abdalla, A.N.; Abdallah, M.E.; Idris, O.F.; Alkahtani, H.M.; Abdel-Aziz, A.-M.; El-Azab, A.S. Mechanistic and Combination Studies of a Novel Dual Her2/EGFR Inhibitor in Breast Cancer Cells. Proceedings of 17th Annual Congress of International Drug Discovery Science and Technology-2019, Kyoto, Japan, 25–27 July 2019. [Google Scholar]
- Nemenqani, D.M. Expression of cyclin D1 in breast carcinoma and its relation to other prognostic markers in Saudi female patients. Int. J. Adv. Res. 2015, 3, 990–996. [Google Scholar]
- Ding, L.; Cao, J.; Lin, W.; Chen, H.; Xiong, X.; Ao, H.; Yu, M.; Lin, J.; Cui, Q. The Roles of Cyclin-Dependent Kinases in Cell-Cycle Progression and Therapeutic Strategies in Human Breast Cancer. Int. J. Mol. Sci. 2020, 21, 1960. [Google Scholar] [CrossRef] [PubMed]
- Criscitiello, C.; Viale, G.; Esposito, A.; Curigliano, G. Dinaciclib for the treatment of breast cancer. Expert Opin. Investig. Drugs 2014, 23, 1305–1312. [Google Scholar] [CrossRef]
- Mita, M.M.; Joy, A.A.; Mita, A.; Sankhala, K.; Jou, Y.-M.; Zhang, D.; Statkevich, P.; Zhu, Y.; Yao, S.-L.; Small, K.; et al. Randomized Phase II Trial of the Cyclin-Dependent Kinase Inhibitor Dinaciclib (MK-7965) Versus Capecitabine in Patients With Advanced Breast Cancer. Clin. Breast Cancer 2014, 14, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Lee, N.V.; Hu, W.; Xu, M.; Ferre, R.A.; Lam, H.; Bergqvist, S.; Solowiej, J.; Diehl, W.; He, Y.-A.; et al. Spectrum and Degree of CDK Drug Interactions Predicts Clinical Performance. Mol. Cancer Ther. 2016, 15, 2273–2281. [Google Scholar] [CrossRef]
- Jane, E.P.; Premkumar, D.R.; Cavaleri, J.M.; Sutera, P.A.; Rajasekar, T.; Pollack, I.F. Dinaciclib, a Cyclin-Dependent Kinase Inhibitor Promotes Proteasomal Degradation of Mcl-1 and Enhances ABT-737-Mediated Cell Death in Malignant Human Glioma Cell Lines. J. Pharmacol. Exp. Ther. 2015, 356, 354–365. [Google Scholar] [CrossRef]
- Deep, G.; Agarwal, R. New combination therapies with cell-cycle agents. Curr. Opin. Investig. Drugs (Lond. Engl. 2000) 2008, 9, 591–604. [Google Scholar]
- Dhanjaland, P.; Fry, J.R. Determinants of MTT reduction in rat hepatocytes. Biomarkers 1997, 2, 111–116. [Google Scholar] [CrossRef]
- Bruggisser, R.; Von Daeniken, K.; Jundt, G.; Schaffner, W.; Tullberg-Reinert, H. Interference of Plant Extracts, Phytoestrogens and Antioxidants with the MTT Tetrazolium Assay. Planta Medica 2002, 68, 445–448. [Google Scholar] [CrossRef]
- Ostrikov, K.; Cheng, H.; Bai, Z.; Li, J. Breast Cancer Cell Line Classification and Its Relevance with Breast Tumor Subtyping. J. Cancer 2017, 8, 3131–3141. [Google Scholar] [CrossRef]
- Comşa, Ş.; Cimpean, A.M.; Raica, M. The Story of MCF-7 Breast Cancer Cell Line: 40 years of Experience in Research. Anticancer Res. 2015, 35, 3147–3154. [Google Scholar] [PubMed]
- Subik, K.; Lee, J.-F.; Baxter, L.; Strzepek, T.; Costello, D.; Crowley, P.; Xing, L.; Hung, M.-C.; Bonfiglio, T.; Hicks, D.G.; et al. The Expression Patterns of ER, PR, HER2, CK5/6, EGFR, Ki-67 and AR by Immunohistochemical Analysis in Breast Cancer Cell Lines. Breast Cancer Basic Clin. Res. 2010, 4, 35–41. [Google Scholar] [CrossRef]
- Sharma, P.; Alsharif, S.; Bursch, K.; Parvathaneni, S.; Anastasakis, D.G.; Chahine, J.; Fallatah, A.; Nicolas, K.; Sharma, S.; Hafner, M.; et al. Keratin 19 regulates cell cycle pathway and sensitivity of breast cancer cells to CDK inhibitors. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Crouch, B.T.; Murphy, H.; Belonwu, S.; Martinez, A.F.; Gallagher, J.; Hall, A.; Soo, M.S.; Lee, M.; Hughes, P.; Haystead, T.; et al. Leveraging ectopic Hsp90 expression to assay the presence of tumor cells and aggressive tumor phenotypes in breast specimens. Sci. Rep. 2017, 7, 17487. [Google Scholar] [CrossRef] [PubMed]
- Shawky, A.M.; Abourehab, M.A.; Abdalla, A.N.; Gouda, A.M. Optimization of pyrrolizine-based Schiff bases with 4-thiazolidinone motif: Design, synthesis and investigation of cytotoxicity and anti-inflammatory potency. Eur. J. Med. Chem. 2020, 185, 111780. [Google Scholar] [CrossRef] [PubMed]
- Makeen, H.A.; Alhazmi, H.A.; Khalid, A.; Al Bratty, M.; Syame, S.M.; Abdalla, A.N.; Homeida, H.E.; Sultana, S.; Ahsan, W. Phytochemical, antimicrobial and cytotoxicity screening of ethanol extract of Acacia ehrenbergiana Hayne grown in Jazan Region of Saudi Arabia. Trop. J. Pharm. Res. 2020, 19, 313–321. [Google Scholar] [CrossRef]
- Attalah, K.M.; Abdalla, A.N.; Aslam, A.; Ahmed, M.; Abourehab, M.A.; ElSawy, N.A.; Gouda, A.M. Ethyl benzoate bearing pyrrolizine/indolizine moieties: Design, synthesis and biological evaluation of anti-inflammatory and cytotoxic activities. Bioorg. Chem. 2020, 94, 103371. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Abdalla, A.N.; Shaheen, U.; Abdallah, Q.M.A.; Flamini, G.; Bkhaitan, M.M.; Abdelhady, M.I.S.; Ascrizzi, R.; Bader, A. Proapoptotic Activity of Achillea membranacea Essential Oil and Its Major Constituent 1,8-Cineole against A2780 Ovarian Cancer Cells. Molecules 2020, 25, 1582. [Google Scholar] [CrossRef]
- Guo, S.; Colbert, L.S.; Fuller, M.; Zhang, Y.; Gonzalez-Perez, R.R. Vascular endothelial growth factor receptor-2 in breast cancer. Biochim. Biophys. Acta (BBA)-Bioenerg. 2010, 1806, 108–121. [Google Scholar] [CrossRef]
- Zhang, Q.; Lu, S.; Li, T.; Yu, L.; Zhang, Y.; Zeng, H.; Qian, X.; Bi, J.; Lin, Y. ACE2 inhibits breast cancer angiogenesis via suppressing the VEGFa/VEGFR2/ERK pathway. J. Exp Clin Cancer Res. 2019, 38, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ceci, C.; Atzori, M.G.; Lacal, P.M.; Graziani, G. Role of VEGFs/VEGFR-1 Signaling and Its Inhibition in Modulating Tumor Invasion: Experimental Evidence in Different Metastatic Cancer Models. Int. J. Mol. Sci. 2020, 21, 1388. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Rafii, S.; Witte, L. Vegfr-1 Antibodies to Treat Breast Cancer. U.S. Patent 2004/0241160 A1, 2 December 2004. [Google Scholar]
- Abdallah, M.E.; El-Readi, M.Z.; Althubiti, M.A.; Almaimani, R.A.; Ismail, A.M.; Idris, S.; Refaat, B.; Almalki, W.H.; Babakr, A.T.; Mukhtar, M.H.; et al. Tamoxifen and the PI3K Inhibitor: LY294002 Synergistically Induce Apoptosis and Cell Cycle Arrest in Breast Cancer MCF-7 Cells. Molecules 2020, 25, 3355. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age | n | Tumor size (cm) | n |
| 40–49 | 6 | 0–1.9 | - |
| 50–59 | 2 | 2–3.9 | 9 |
| 60–69 | 3 | 4–5.9 | 2 |
| 70–79 | 1 | 6–7.9 | 1 |
| gender | n | Position | n |
| female | 12 | right | 6 |
| male | 0 | left | 6 |
| Grade | n | Histology | n |
| II | 8 | ductal | 10 |
| III | 4 | lobular | 2 |
| ER% | n | PR% | n |
| 1–19 | - | 1–19 | 7 |
| 20–39 | 1 | 20–39 | 1 |
| 40–59 | - | 40–59 | - |
| 60–79 | 6 | 60–79 | 1 |
| 80–99 | 5 | 80–99 | 3 |
| Ki-67% | n | ||
| 1–19 | 9 | ||
| 20–39 | 1 | ||
| 40–59 | 2 | ||
| 60–79 | - | ||
| 80–99 | - |
| No | Network Name | Processes | Size | Target | Pathways | p-Value | zScore | gScore |
|---|---|---|---|---|---|---|---|---|
| 1 | VEGFR-2, cRaf-1, FAK1, c-Src, VEGFR-3 | Transmembrane receptor protein tyrosine kinase signalling pathway (77.6%), enzyme linked receptor protein signalling pathway (81.6%), locomotion (85.7%), cell morphogenesis (73.5%), cell projection morphogenesis (67.3%). | 52 | 16 | 56 | 4.65 × 10−36 | 70.29 | 140.29 |
| 2 | TrkA, LAT, Syk, c-Src, Fer | Peptidyl-tyrosine phosphorylation (58.1%), peptidyl-tyrosine modification (58.1%), transmembrane receptor protein tyrosine kinase signalling pathway (65.1%), positive regulation of ERK1 and ERK2 cascade (55.8%), regulation of ERk1 and ERK2 cascade (58.1%). | 53 | 29 | 3 | 1.25 × 10−74 | 128.86 | 132.61 |
| 3 | Ephrin-A receptor 3, YES, Ephrin-A receptor 2, Fer, p130 | Transmembrane receptor protein tyrosine kinase signalling pathway (50.0%), regulation of phosphorylation (68.8%), chemotaxis (47.9%), taxis (47.9%), regulation of protein phosphorylation (64.6%). | 50 | 20 | 5 | 2.28 × 10−47 | 88.80 | 95.05 |
| Drug(s) nM | MCF7 | MRC5 | ||||
|---|---|---|---|---|---|---|
| HAA2020 | Tamoxifen | Dinaciclib | IC50 | *CI | IC50 | **SI |
| 125 | 480 ± 30 | 15221 ± 169 | 32 | |||
| 250 | ||||||
| 500 | - | |||||
| 2500 | ||||||
| 5000 | ||||||
| 187.5 | 738 ± 54 | 18450 ± 213 | 25 | |||
| 375 | ||||||
| 750 | - | |||||
| 3750 | ||||||
| 7500 | ||||||
| 1.25 | 4 ± 0 | 250 ± 14 | 62 | |||
| 2.5 | ||||||
| 5 | - | |||||
| 25 | ||||||
| 50 | ||||||
| HAA2020: Tam (1:1.5) | 207 ± 50 | 4558 ± 150 | 22 | |||
| 500 | 750 | 0.964 | ||||
| 2500 | 3750 | 0.823 | ||||
| 5000 | 7500 | 0.811 | ||||
| HAA2020: Din (100:1) | 1 ± 0 | 105 ± 11 | 105 | |||
| 500 | 5 | 0.463 | ||||
| 2500 | 25 | 0.301 | ||||
| 5000 | 50 | 0.260 | ||||
| Tam: Dina (150:1) | 830 ± 91 | 16600 ± 390 | 20 | |||
| 750 | 5 | >1 | ||||
| 3750 | 25 | 0.954 | ||||
| 7500 | 50 | 0.901 | ||||
| HAA2020: Tam: Dina (100:150:1) | 3 ± 0 | 153 ± 29 | 51 | |||
| 500 | 750 | 5 | 0.810 | |||
| 2500 | 3750 | 25 | 0.714 | |||
| 5000 | 7500 | 50 | 0.680 | |||
| Gene | Sequence |
|---|---|
| Hsp90 | F: TTGGTTACTTCCCCGTGCTG R: GCCTTTTGCCGTAGGGTTTC |
| VEGFR-2 | F: TGATACTGGAGCCTACAAGTGCTT R: CCTGTAATCTTGAACGTAGACATAAATGA |
| GAPDH | F: AGGTCGGTGTGAACGGATTTG R: TGTAGACCATGTAGTTGAGGTCA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdalla, A.N.; Qattan, A.; Malki, W.H.; Shahid, I.; Hossain, M.A.; Ahmed, M. Significance of Targeting VEGFR-2 and Cyclin D1 in Luminal-A Breast Cancer. Molecules 2020, 25, 4606. https://doi.org/10.3390/molecules25204606
Abdalla AN, Qattan A, Malki WH, Shahid I, Hossain MA, Ahmed M. Significance of Targeting VEGFR-2 and Cyclin D1 in Luminal-A Breast Cancer. Molecules. 2020; 25(20):4606. https://doi.org/10.3390/molecules25204606
Chicago/Turabian StyleAbdalla, Ashraf N., Amal Qattan, Waleed H. Malki, Imran Shahid, Mohammad Akbar Hossain, and Muhammad Ahmed. 2020. "Significance of Targeting VEGFR-2 and Cyclin D1 in Luminal-A Breast Cancer" Molecules 25, no. 20: 4606. https://doi.org/10.3390/molecules25204606
APA StyleAbdalla, A. N., Qattan, A., Malki, W. H., Shahid, I., Hossain, M. A., & Ahmed, M. (2020). Significance of Targeting VEGFR-2 and Cyclin D1 in Luminal-A Breast Cancer. Molecules, 25(20), 4606. https://doi.org/10.3390/molecules25204606