Chemotaxonomic Study of Bostrychia spp. (Ceramiales, Rhodophyta) Based on Their Mycosporine-Like Amino Acid Content


Abstract
:1. Introduction
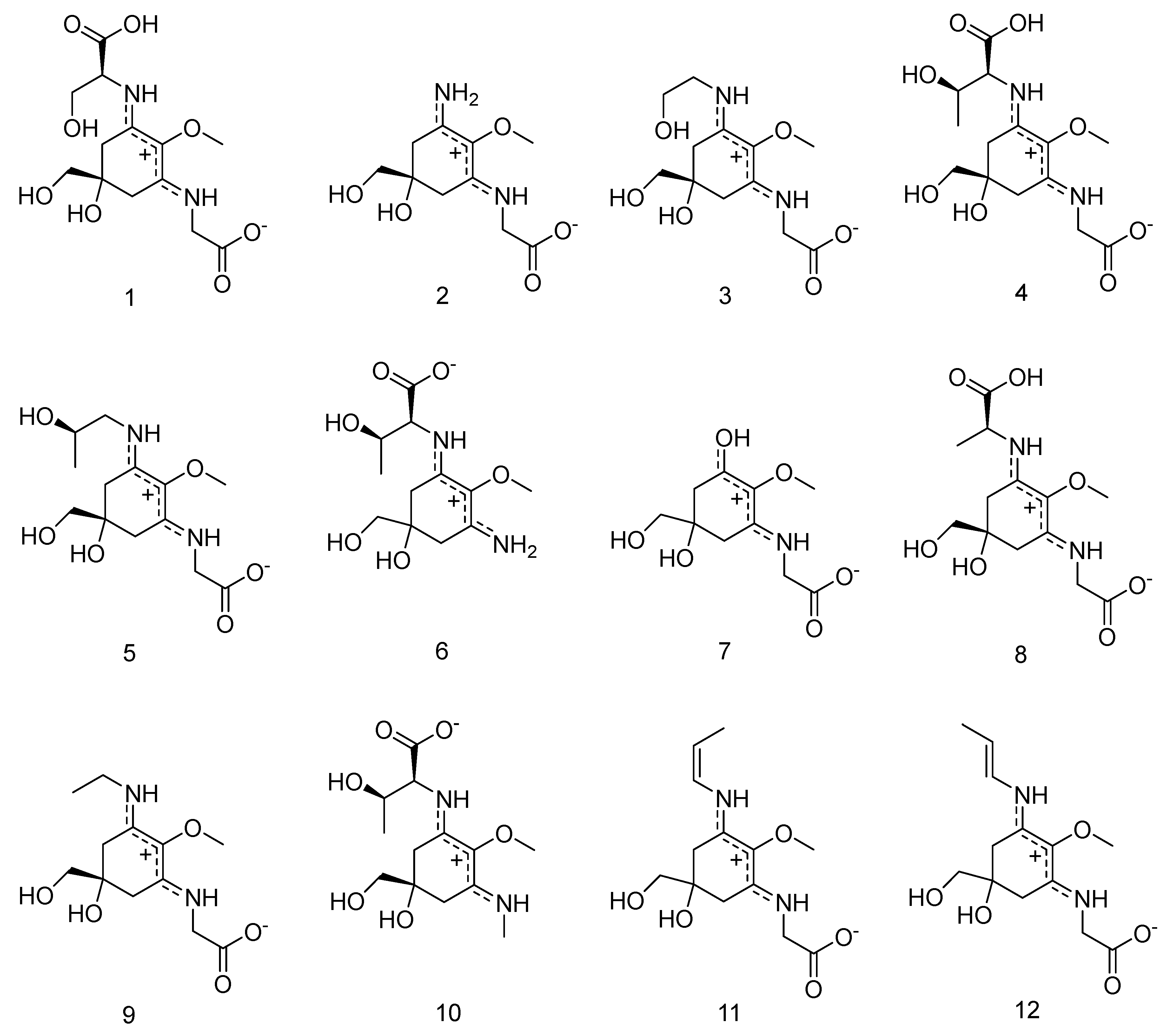
2. Results
2.1. Method Development
2.1.1. Sample Preparation
2.1.2. HPLC-DAD
2.2. Method Validation
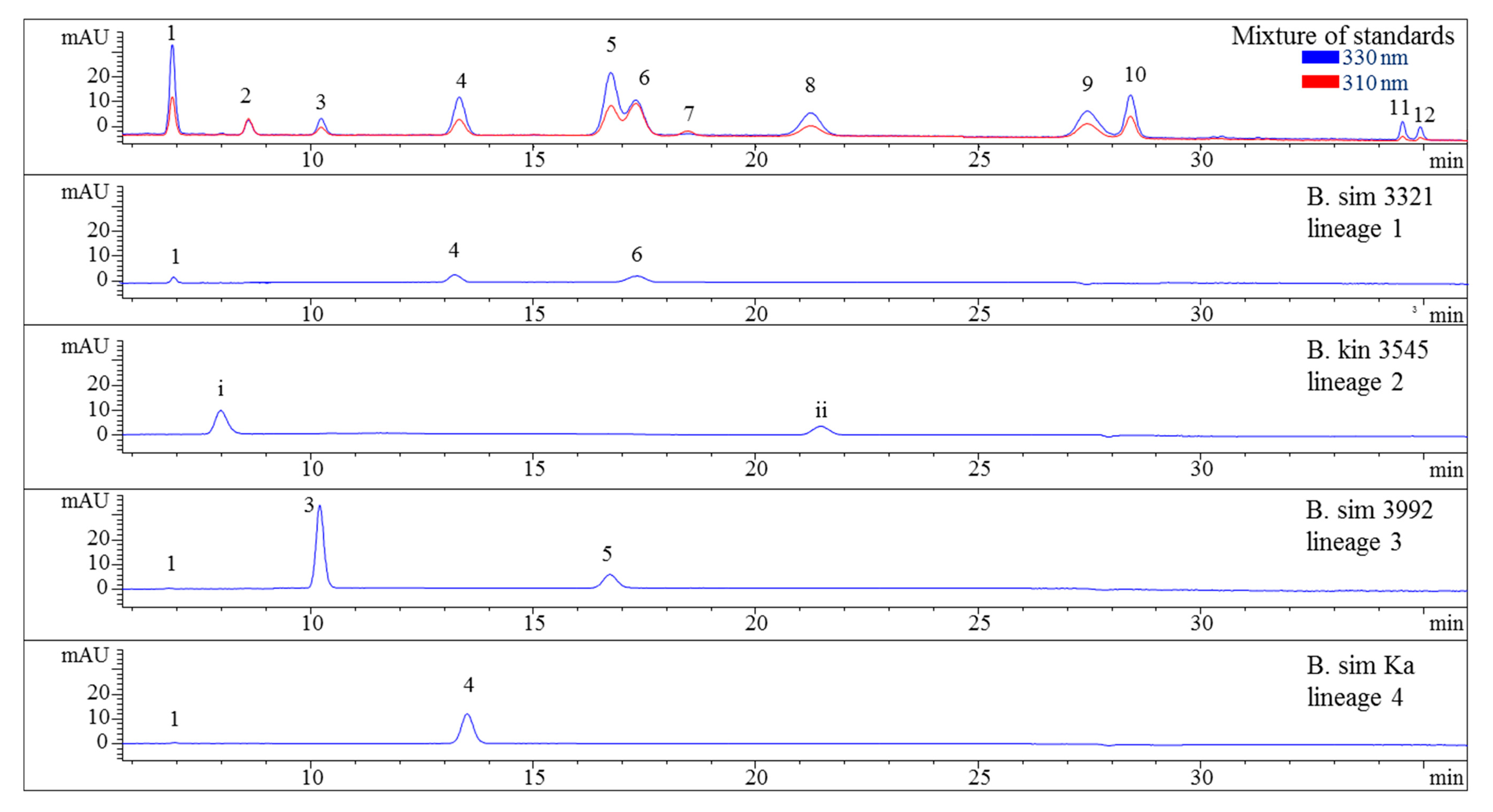
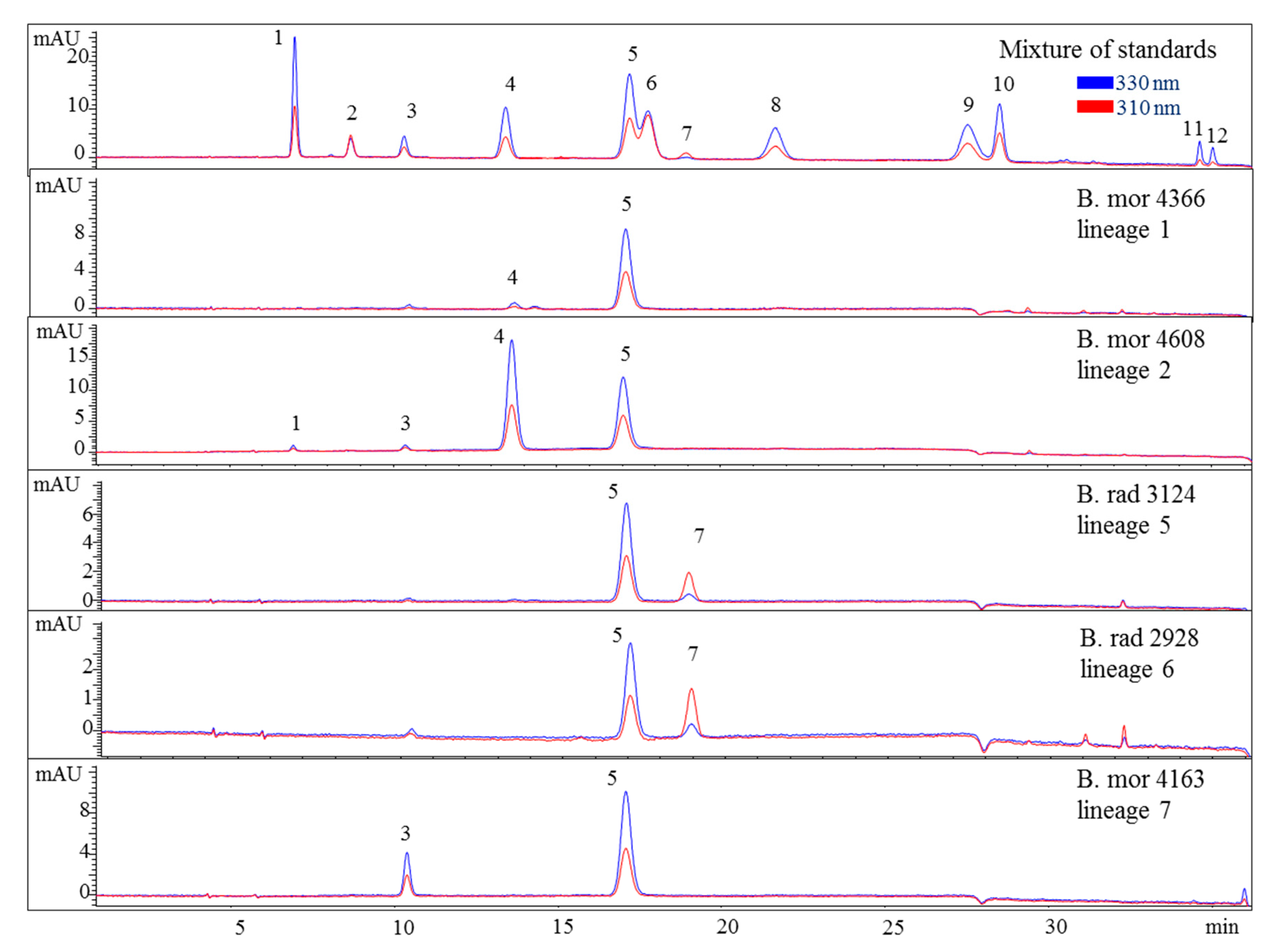
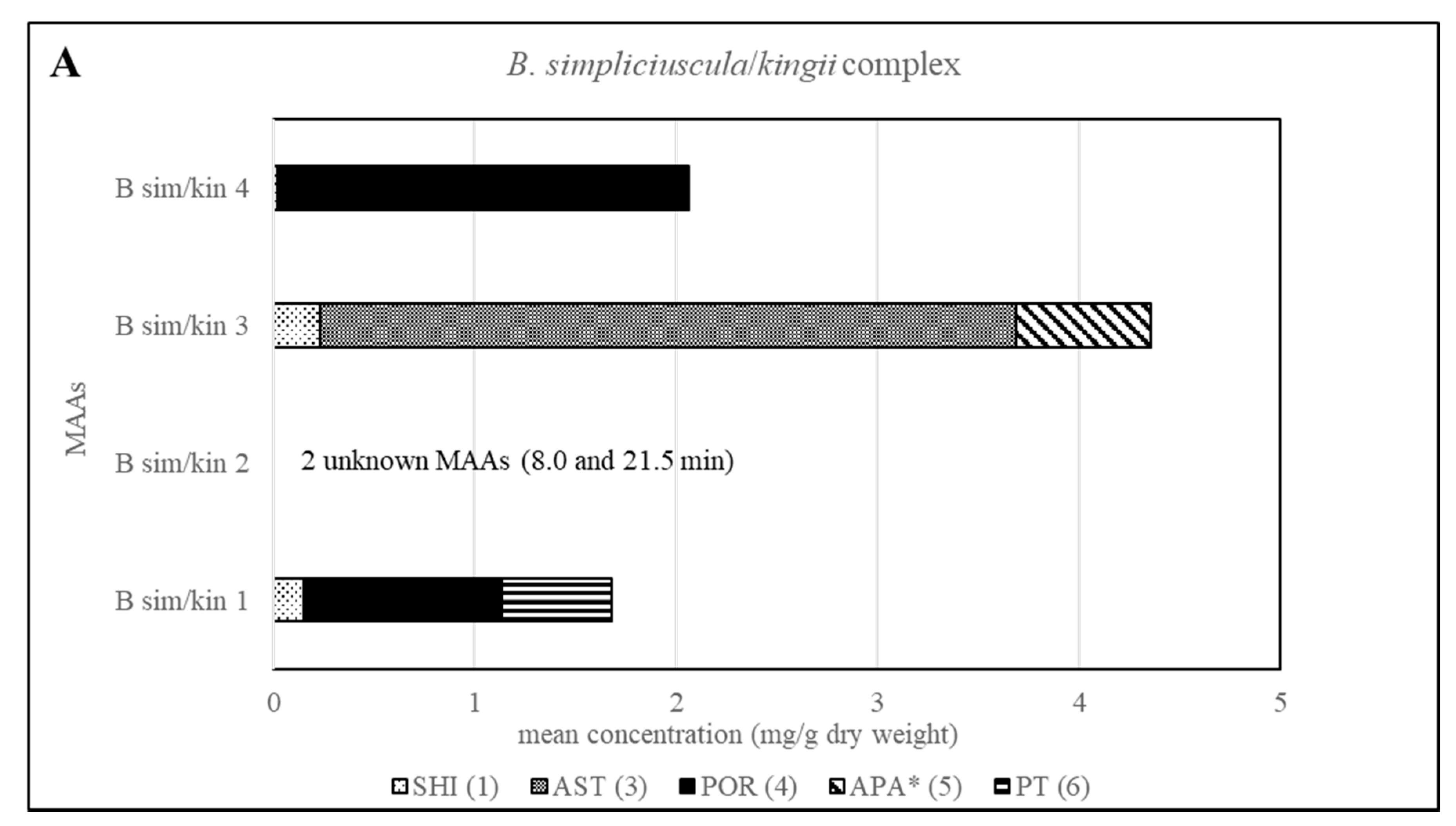
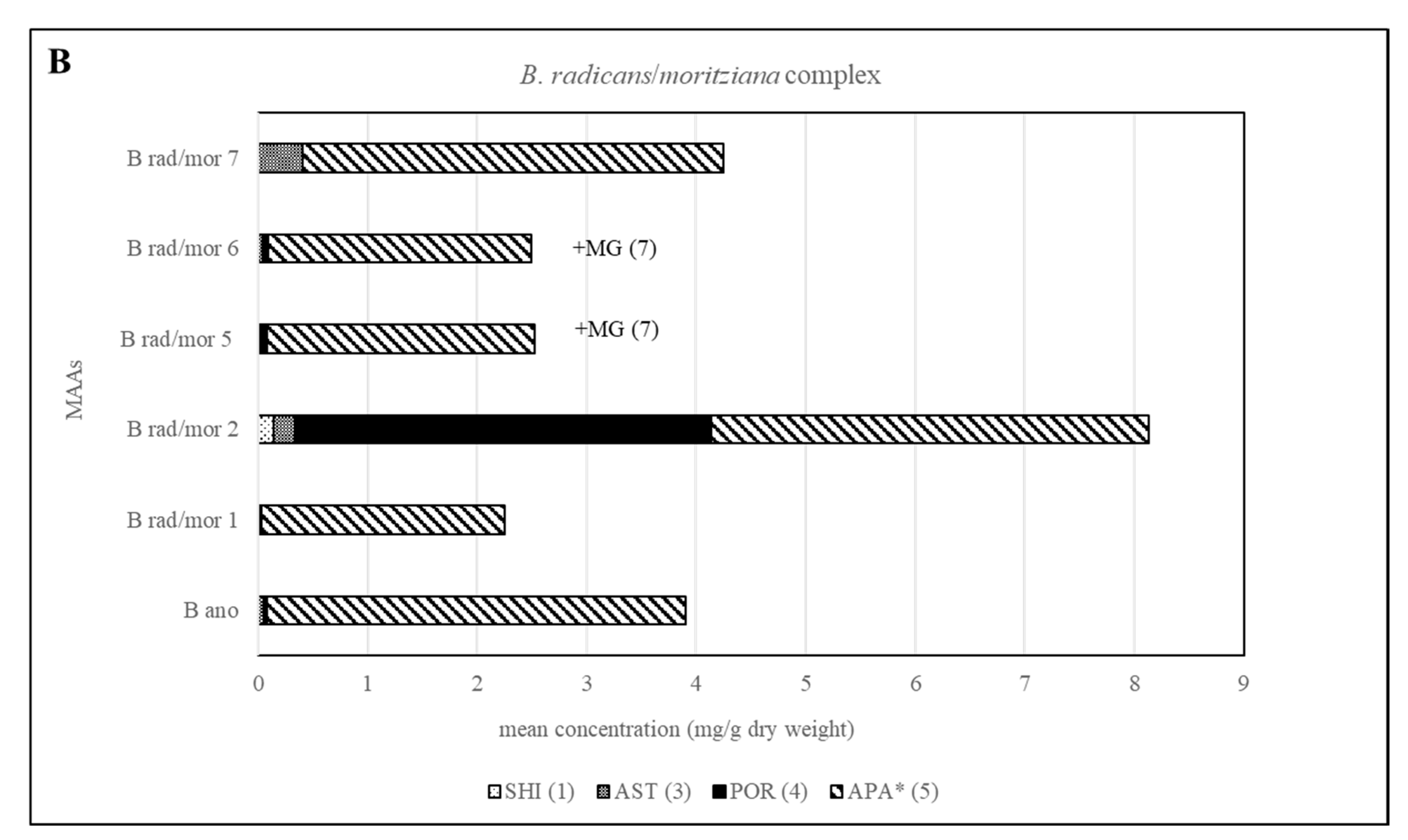
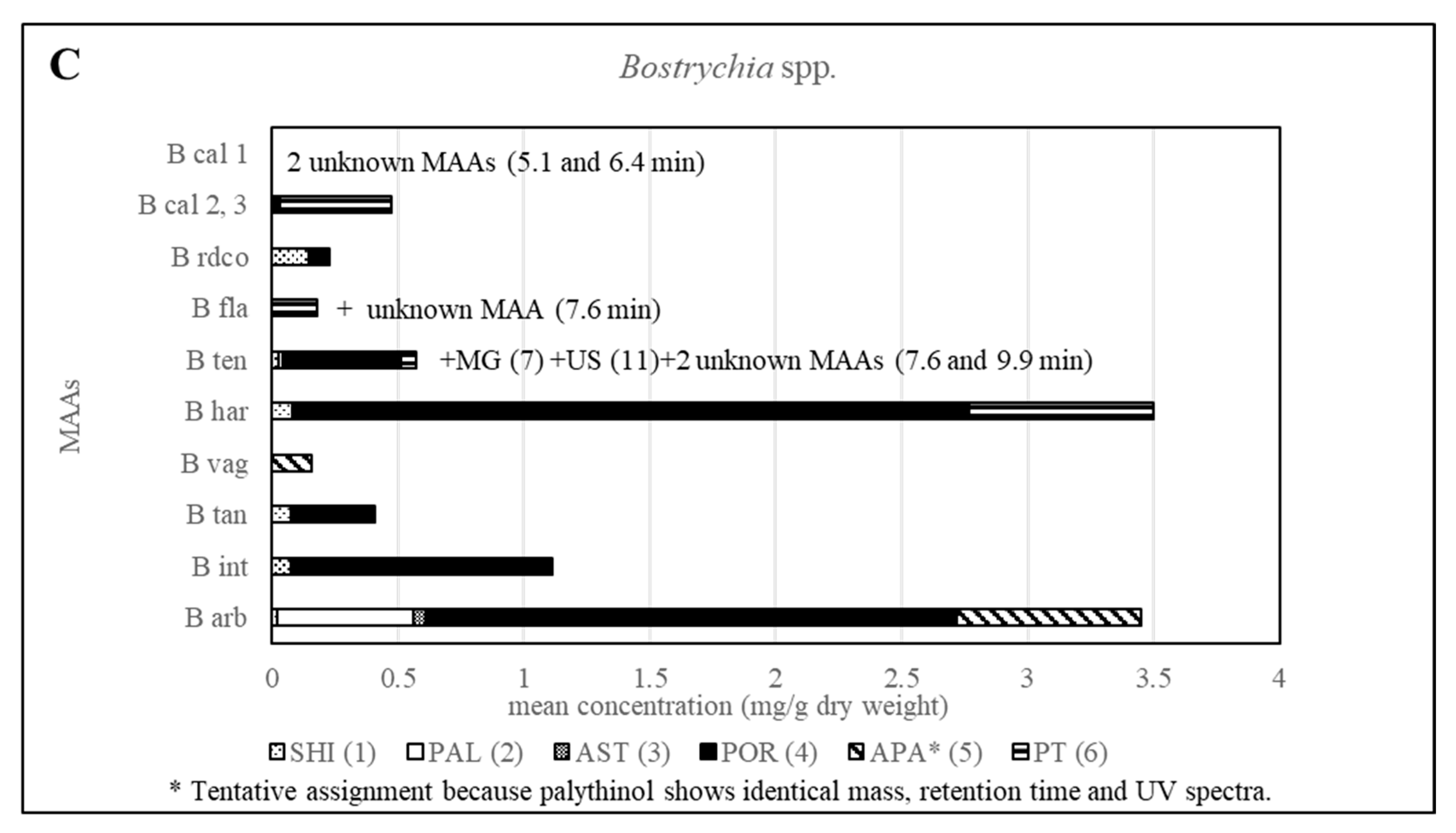
2.3. Sample Analysis
2.4. LC-MS Based Analyte Identification
3. Discussion
4. Experimental
4.1. Biological Material
4.2. Chemicals
4.3. Analyte Isolation
4.4. HPLC Sample Preparation for Quantitation Purposes
4.5. General Experimental Procedures
4.5.1. HPLC-DAD Analysis
4.5.2. HPLC-DAD-MS Analysis
4.6. Calibration and Method Validation
4.6.1. Linearity, Limit of Detection (LOD) and Limit of Quantification (LOQ)
4.6.2. Precision and Accuracy
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Saengkaew, J.; Muangmai, N.; Zuccarello, G. Cryptic diversity of the mangrove-associated alga Bostrychia (Rhodomelaceae, Rhodophyta) from Thailand. Bot. Mar. 2016, 59, 363–371. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase; World-Wide Electronic Publication, National University of Ireland: Galway, Ireland, 2019. Available online: http://www.algaebase.org (accessed on 11 July 2019).
- King, R.; Puttock, C. Morphological and taxonomy of Bostrychia and Stictosiphonia (Rhodomelaceae/Rhodophyta). Aust. Syst. Bot. 1989, 2, 1–73. [Google Scholar] [CrossRef]
- Khin Gyi, K.; Htun, U.S. The genus Bostrychia Montagne (Ceramiales, Rhodophyta) in Setse and Kyaikkhami coastal areas. Mawlamyine Univ. Res. J. 2012, 4, 1–17. [Google Scholar]
- Zuccarello, G.C.; West, J.A.; Kamiya, M. Non-monophyly of Bostrychia simpliciuscula (Ceramiales, Rhodophyta): Multiple species with very similar morphologies, a revised taxonomy of cryptic species: Polyphyletic Bostrychia species. Phycol. Res. 2018, 66, 100–107. [Google Scholar] [CrossRef]
- Zuccarello, G.; West, J.; Goer, S. Diversity of the Bostrychia radicans/Bostrychia moritziana species complex (Rhodomelaceae, Rhodophyta) in the mangroves of New Caledonia. Cryptogam. Algol. 2006, 27, 245–254. [Google Scholar]
- Zuccarello, G.; West, J.; King, R. Evolutionary divergence in the Bostrychia moritziana/B. radicans complex (Rhodomelaceae, Rhodophyta): Molecular and hybridization data. Phycologia 1999, 38, 234–244. [Google Scholar] [CrossRef]
- West, J.; Kamiya, M.; Goer, S.; Karsten, U.; Zuccarello, G. Observations on some mangrove-associated algae from the western Pacific (Guam, Chuuk, Kosrae, and Pohnpei). Algae 2013, 28, 241–266. [Google Scholar] [CrossRef] [Green Version]
- West, J.; Goer, S.; Zuccarello, G. Monosiphonous growth and cell-death in an unusual Bostrychia (Rhodomelaceae, Rhodophyta): B. anomala sp. nov. Algae 2013, 28, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Carreto, J.; Carignan, M.; Montoya, N. A high-resolution reverse-phase liquid chromatography method for the analysis of mycosporine-like amino acids (MAAs) in marine organisms. Mar. Biol. 2005, 146, 237–252. [Google Scholar] [CrossRef]
- Hartmann, A.; Becker, K.; Karsten, U.; Remias, D.; Ganzera, M. Analysis of Mycosporine-Like Amino Acids in Selected Algae and Cyanobacteria by Hydrophilic Interaction Liquid Chromatography and a Novel MAA from the Red Alga Catenella repens. Mar. Drugs 2015, 13, 6291–6305. [Google Scholar] [CrossRef] [PubMed]
- Orfanoudaki, M.; Hartmann, A.; Karsten, U.; Ganzera, M. Chemical profiling of mycosporine-like amino acids in twenty-three red algal species. J. Phycol. 2019, 55, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Orfanoudaki, M.; Hartmann, A.; Ngoc, H.N.; Gelbrich, T.; West, J.; Karsten, U.; Ganzera, M. Mycosporine-like amino acids, brominated and sulphated phenols: Suitable chemotaxonomic markers for the reassessment of classification of Bostrychia calliptera (Ceramiales, Rhodophyta). Phytochemistry 2020, 174, 112344. [Google Scholar] [CrossRef] [PubMed]
- Carreto, J.I.; Carignan, M.O. Mycosporine-like amino acids: Relevant secondary metabolites. Chemical and ecological aspects. Mar. Drugs 2011, 9, 387–446. [Google Scholar] [CrossRef] [PubMed]
- ICH official Web Site: ICH. Available online: https://www.ich.org/ (accessed on 14 July 2020).







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Calibration Data | ||||||
| Substance | Regression Equation a | Coefficient of Determination | Linearity b (μg/mL) | LOD (μg/mL) | LOQ (μg/mL) | |
| 1 | y = 69632x + 2.8305 | R² = 0.9997 | 0.0153–7.8125 | 0.0020 | 0.0060 | |
| 4 | y = 68147x + 2.6536 | R² = 0.9998 | 0.0557–7.125 | 0.0094 | 0.0284 | |
| 5 | y = 45708x + 4.9671 | R² = 0.9988 | 0.0669–34.250 | 0.0118 | 0.0359 | |
| 8 | y = 51522x + 2.3346 | R² = 0.9999 | 0.0605–7.750 | 0.0101 | 0.0306 | |
| 10 | y = 32772x + 4.1515 | R² = 0.9998 | 0.1621–41.5 | 0.0189 | 0.0571 | |
| Accuracy and Precision | ||||||
| Precision | Accuracy e | |||||
| Substance | Intra-day c | Inter-day d | Substance | Low | Medium | High |
| 1 | 3.94 | 5.15 | 1 | 95.11 ± 1.33 | 92.70 ± 0.66 | 92.59 ± 0.32 |
| 2 | 3.67 | 3.55 | 4 | 97.93 ± 0.33 | 97.04 ± 0.56 | 97.53 ± 0.28 |
| 3 | 3.54 | 4.98 | 5 | 97.45 ± 1.57 | 95.78 ± 0.24 | 95.67 ± 0.80 |
| 4 | 3.76 | 3.67 | 8 | 97.89 ± 4.85 | 95.08 ± 0.64 | 95.29 ± 0.85 |
| 5 | 3.87 | 4.06 | 10 | 96.22 ± 4.68 | 95.53 ± 1.79 | 96.64 ± 2.22 |
| Sample | SHI (1) | PAL (2) | AST (3) | POR (4) | APA* (5) | PT (6) | MG (7) | US (11) | MAA at 7.6 min | MAA at 9.9 min | MAA at 8.0 min | MAA at 21.5 min | MAAs at 5.1 and 6.4 min |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| B cal 1 | Present | ||||||||||||
| B cal 2,3 | 0.03 (±0.05) | 0.44 (±0.41) | |||||||||||
| B fla | 0.18 (±0.22) | Sometimes Present | |||||||||||
| B har | 0.08 | 2.689 | 0.731 | ||||||||||
| B ten | 0.03 (±0.12) | 0.01 (±0.08) | 0.01 (±0.08) | 0.46 (±0.46) | 0.06 (±0.07) | Sometimes Present | Sometimes Present | Present | Sometimes Present | ||||
| B sim 1 | 0.15 (±0.08) | 0.98 (±0.49) | 0.55 (±0.29) | ||||||||||
| B sim 4 | 0.02 (±0.01) | 2.04 (±1.07) | |||||||||||
| B int | 0.07 (±0.08) | 1.04 (±0.26) | |||||||||||
| B tan | 0.07 (±0.04) | 0.34 (±0.11) | |||||||||||
| B rdco | 0.15 (±0.21) | 0.08 (±0.02) | |||||||||||
| B arb | 0.01 | 0.54 | 0.05 | 2.11 | 0.74 | ||||||||
| B ano | 0.37 (±0.42) | 0.11 (±0.22) | 3.46 (±1.15) | ||||||||||
| B rad/mor 1 | 0.03 (±0.04) | 2.22 (±0.88) | |||||||||||
| B rad/mor 2 | 0.14 (±0.18) | 0.19 (±0.14) | 3.80 (±2.34) | 4.00 (±1.49) | |||||||||
| B rad/mor 5 | 0.004 (±0.01) | 0.08 (±0.15) | 2.44 (±1.35) | Present | |||||||||
| B rad/mor 6 | 0.04 (±0.08) | 0.05 (±0.1) | 2.40 (±0.87) | Present | |||||||||
| B rad/mor 7 | 0.41 (±0.39) | 3.84 (±0.46) | |||||||||||
| B vag | 0.15 | ||||||||||||
| B sim 3 | 0.23 (±0.23) | 3.45 (±2.95) | 0.67 (±0.95) | ||||||||||
| B kin (B sim 2) | Present | Present |
| Retention Time | λmax (nm) | Mass (Da) |
|---|---|---|
| 7.6 | 320 | 274 |
| 8.0 | 334 | 403 |
| 9.9 | 318 | 294 |
| 21.5 | 308 | 316 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orfanoudaki, M.; Hartmann, A.; Kamiya, M.; West, J.; Ganzera, M. Chemotaxonomic Study of Bostrychia spp. (Ceramiales, Rhodophyta) Based on Their Mycosporine-Like Amino Acid Content. Molecules 2020, 25, 3273. https://doi.org/10.3390/molecules25143273
Orfanoudaki M, Hartmann A, Kamiya M, West J, Ganzera M. Chemotaxonomic Study of Bostrychia spp. (Ceramiales, Rhodophyta) Based on Their Mycosporine-Like Amino Acid Content. Molecules. 2020; 25(14):3273. https://doi.org/10.3390/molecules25143273
Chicago/Turabian StyleOrfanoudaki, Maria, Anja Hartmann, Mitsunobu Kamiya, John West, and Markus Ganzera. 2020. "Chemotaxonomic Study of Bostrychia spp. (Ceramiales, Rhodophyta) Based on Their Mycosporine-Like Amino Acid Content" Molecules 25, no. 14: 3273. https://doi.org/10.3390/molecules25143273
APA StyleOrfanoudaki, M., Hartmann, A., Kamiya, M., West, J., & Ganzera, M. (2020). Chemotaxonomic Study of Bostrychia spp. (Ceramiales, Rhodophyta) Based on Their Mycosporine-Like Amino Acid Content. Molecules, 25(14), 3273. https://doi.org/10.3390/molecules25143273