
P-MAPA and Interleukin-12 Reduce Cell Migration/Invasion and Attenuate the Toll-Like Receptor-Mediated Inflammatory Response in Ovarian Cancer SKOV-3 Cells: A Preliminary Study


 , , ,
, , ,  and
and 
Abstract
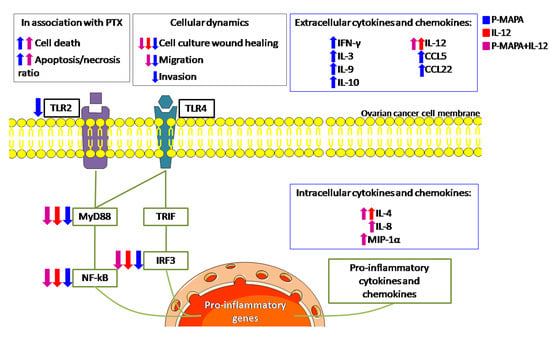
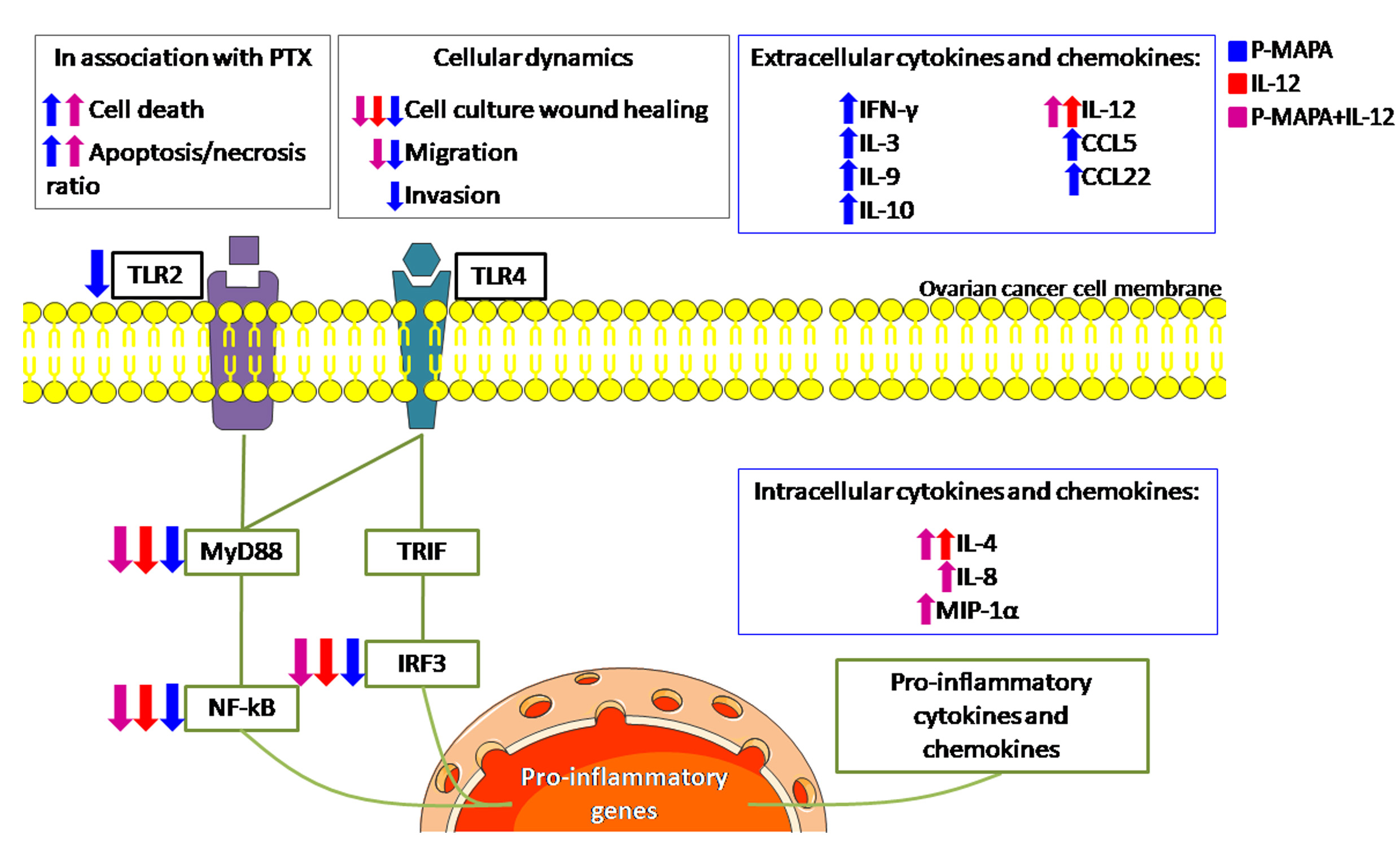{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. P-MAPA and IL-12 Reduce Cell Viability and Induce Apoptosis in the Presence of PTX in SKOV-3 Cells
2.2. Combination of P-MAPA with IL-12 Is Essential to Reduce Cell Migration Whereas P-MAPA Alone Decreased The Invasive Potential of SKOV-3 Cells
2.3. Immunotherapy with P-MAPA and IL-12 Significantly Reduced the TLR-Mediated Downstream Molecules Involved in the Inflammatory Process of SKOV-3 Cells
2.4. P-MAPA Stimulates the Secretion of Pro- and Anti-Inflammatory Molecules, Whereas Its Association with IL-12 Induced the Synthesis of Inflammatory Cytokines in SKOV-3 Cells
3. Discussion
4. Materials and Methods
4.1. Cell Line and Cell Culture
4.2. Treatments with P-MAPA and IL-12
4.3. Cell Cytotoxicity (MTT Assay)
4.4. Apoptosis Rate by Annexin V-FITC/PI Staining
4.5. Cell Cycle Determination by PI Staining
4.6. Wound-Healing Assay
4.7. Cell Migration Using Transwell Insert
4.8. Invasion Assay
4.9. Immunofluorescence Assay
4.10. Western Blot Analysis
4.11. Cytokine and Chemokine Assay
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BSA | Bovine serum albumin |
| CEEA | Ethical Committee of the Institute of Bioscience/UNESP |
| CEMIB | Multidisciplinary Center for Biological Investigation |
| DAB | Diaminobenzidine |
| DAPI | 6-diamidino-2-phenylindole |
| DCs | Dendritic cells |
| FITC | Fluorescein Isothiocyanate |
| HRP-conjugated | Horseradish peroxidase-conjugated |
| H | Hematoxylin |
| IFN | Interferon |
| IFN-γ | Interferon gamma |
| IL-6 | Interleukin 6 |
| IL-8 | Interleukin 8 |
| IRF3 | Interferon regulatory factor 3 |
| MyD88 | Myeloid differentiation factor 88 |
| NF-kB p65 | Nuclear factor kappa B subunit p65 |
| NK | natural killer cells |
| OC | ovarian cancer |
| PBS | phosphate-buffered saline |
| P-MAPA | Protein aggregate magnesium-ammonium phospholinoleate-palmitoleate anhydride |
| PTX | paclitaxel |
| RIPA | Radioimmunoprecipitation assay buffer |
| SDS-PAGE | Sodium dodecyl sulphate-polyacrylamide gel electrophoresis |
| TBS-T | Tris-Buffered Saline plus Tween 20 |
| CD4 + T | CD4-positive T cells |
| CD8 + T | CD8-positive T cells |
| Th1 | T helper 1 |
| TLR (s) | Toll-like receptor (s) |
| TLR2 | Toll-like receptor 2 |
| TLR4 | Toll-like receptor 4 |
| TRIF | TIR domain-containing adaptor inducing interferon-beta |
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Fallows, S.; Price, J.; Atkinson, R.J.; Johnston, P.G.; Hickey, I.; Russell, S.E. P53 mutation does not affect prognosis in ovarian epithelial malignancies. J. Pathol. 2001, 194, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Cannistra, S.A. Cancer of the ovary. N. Engl. J. Med. 2004, 351, 2519–2565. [Google Scholar] [CrossRef] [PubMed]
- Chuffa, L.G.; Fioruci-Fontanelli, B.A.; Mendes, L.O.; Fávaro, W.J.; Pinheiro, P.F.; Martinez, M.; Martinez, F.E. Characterization of chemically induced ovarian carcinomas in an ethanol-preferring rat model: Influence of long-term melatonin treatment. PLoS ONE 2013, 8, e81676. [Google Scholar] [CrossRef] [PubMed]
- Chuffa, L.G.; Lupi-Júnior, L.A.; Costa, A.B.; Amorim, J.P.; Seiva, F.R. The role of sex hormones and steroid receptors on female reproductive cancers. Steroids 2017, 118, 93–108. [Google Scholar] [CrossRef]
- Ebell, M.H.; Culp, M.B.; Radke, T.J. A systematic review of symptoms for the diagnosis of ovarian cancer. Am. J. Prev. Med. 2016, 50, 384–394. [Google Scholar] [CrossRef]
- Kelly, M.G.; Alvero, A.B.; Chen, R.; Silasi, D.A.; Abrahams, V.M.; Chan, S.; Visintin, I.; Rutherford, T.; Mor, G. TLR-4 signaling promotes tumor growth and paclitaxel chemoresistance in ovarian cancer. Cancer Res. 2006, 66, 3859–3868. [Google Scholar] [CrossRef]
- Bronte, G.; Cicero, G.; Sortino, G.; Pernice, G.; Catarella, M.T.; D’Alia, P.; Cusenza, S.; Lo Dico, S.; Bronte, E.; Sprini, D.; et al. Immunotherapy for recurrent ovarian cancer: A further piece of the puzzle or a striking strategy? Expert Opin. Biol. Ther. 2014, 14, 103–114. [Google Scholar] [CrossRef][Green Version]
- Fávaro, W.J.; Nunes, O.S.; Seiva, F.R.; Nunes, I.S.; Woolhiser, L.K.; Durán, N.; Lenaerts, A.J. Effects of P-MAPA immunomodulator on Toll-like receptors and p53: Potential therapeutic strategies for infectious diseases and cancer. Infect. Agent. Cancer 2012, 7, 1–15. [Google Scholar]
- Garcia, P.V.; Seiva, F.R.; Carniato, A.P.; de Mello Júnior, W.; Duran, N.; Macedo, A.M.; de Oliveira, A.G.; Romih, R.; de Oliveira, A.G.; Romih, R.; et al. Increased toll-like receptors and p53 levels regulate apoptosis and angiogenesis in non-muscle invasive bladder cancer: Mechanism of action of P-MAPA biological response modifier. BMC Cancer 2016, 16, 422. [Google Scholar] [CrossRef]
- de Almeida Chuffa, L.G.; de Moura Ferreira, G.; Lupi, L.A.; da Silva Nunes, I.; Fávaro, W.J. P-MAPA immunotherapy potentiates the effect of cisplatin on serous ovarian carcinoma through targeting TLR4 signaling. J. Ovarian Res. 2018, 11, 8. [Google Scholar] [CrossRef]
- Chen, R.; Alvero, A.B.; Silasi, D.A.; Steffensen, K.D.; Mor, G. Cancers take their Toll–the function and regulation of Toll-like receptors in cancer cells. Oncogene 2008, 27, 225–233. [Google Scholar] [CrossRef]
- Szajnik, M.; Szczepanski, M.J.; Czystowska, M.; Elishaev, E.; Mandapathil, M.; Nowak-Markwitz, E.; Spaczynski, M.; Whiteside, T.L. TLR4 signaling induced by lipopolysaccharide or paclitaxel regulates tumor survival and chemoresistance in ovarian cancer. Oncogene 2009, 28, 4353–4363. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.C.; Su, Q.B.; Wu, F.X.; Zhang, X.L.; Liu, P.S. Role of TLR4 for paclitaxel chemotherapy in human epithelial ovarian cancer cells. Eur. J. Clin. Investig. 2009, 39, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Trinchieri, G. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat. Rev. Immunol. 2003, 3, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Cohen, C.A.; Shea, A.A.; Heffron, C.L.; Schmelz, E.M.; Roberts, P.C. Interleukin-12 Immunomodulation Delays the Onset of Lethal Peritoneal Disease of Ovarian Cancer. J. Interferon Cytokine Res. 2016, 36, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.P.; Trinchieri, G. Interleukin-12 in anti-tumor immunity and immunotherapy. Cytokine Growth Factor Rev. 2002, 13, 155–168. [Google Scholar] [CrossRef]
- Hurteau, J.A.; Blessing, J.A.; DeCesare, S.L.; Creasman, W.T. Evaluation of recombinanthuman interleukin-12 in patients with recurrent or refractory ovarian cancer: Agynecologic oncology group study. Gynecol. Oncol. 2001, 82, 7–10. [Google Scholar] [CrossRef]
- Lenzi, R.; Edwards, R.; June, C.; Seiden, M.V.; Garcia, M.E.; Rosenblum, M.; Freedman, R.S. Phase II study of intraperitoneal recombinant interleukin-12 (rhIL-12) in patients with peritoneal carcinomatosis (residual disease <1 cm) associated with ovarian cancer or primary peritoneal carcinoma. J. Transl. Med. 2007, 5, 66. [Google Scholar]
- Präbst, K.; Engelhardt, H.; Ringgeler, S.; Hübner, H. Cell Viability Assays: Methods and Protocols. Methods Mol. Biol. 2017, 1601, 117. [Google Scholar]
- Ahn, H.J.; Kim, Y.S.; Kim, J.U.; Han, S.M.; Shin, J.W.; Yang, H.O. Mechanism of taxol-inducedapoptosis in human SKOV3 ovarian carcinoma cells. J. Cell Biochem. 2004, 91, 1043–1052. [Google Scholar] [CrossRef]
- Wang, Y.J.; Fletcher, R.; Yu, J.; Zhang, L. Immunogenic effects of chemotherapy-induced tumor cell death. Genes Dis. 2018, 5, 194–203. [Google Scholar] [CrossRef]
- Chuffa, L.G.; Fioruci-Fontanelli, B.A.; Mendes, L.O.; Ferreira Seiva, F.R.; Martinez, M.; Fávaro, W.J.; Domeniconi, R.F.; Pinheiro, P.F.; Delazari Dos Santos, L.; Martinez, F.E. Melatonin attenuates the TLR4-mediated inflammatory response through MyD88- and TRIF-dependent signaling pathways in an in vivo model of ovarian cancer. BMC Cancer 2015, 15, 34. [Google Scholar] [CrossRef]
- Lavoue, V.; Thedrez, A.; Leveque, J.; Foucher, F.; Henno, S.; Jauffret, V.; Belaud-Rotureau, M.A.; Catros, V.; Cabillic, F. Immunity of human epithelial ovarian carcinoma: The paradigm of immunesuppression in cancer. J. Transl. Med. 2013, 11, 1–12. [Google Scholar] [CrossRef]
- Jouhi, L.; Koljonen, V.; Böhling, T.; Haglund, C.; Hagström, J. The expression of toll-like receptors 2, 4, 5, 7 and 9 in Merkel cell carcinoma. Anticancer Res. 2015, 35, 1843–1849. [Google Scholar]
- Zhu, G.; Gui, Z. Effect of silkworm peptide on inducting M1 type polarization andTh1 activation via TLR2-induced MyD88-dependent pathway. Food Sci. Nutr. 2019, 7, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- d’Adhemar, C.J.; Spillane, C.D.; Gallagher, M.F.; Bates, M.; Costello, K.M.; Barry-O’Crowley, J.; Haley, K.; Kernan, N.; Murphy, C.; Smyth, P.C.; et al. The MyD88+ phenotype is an adverse prognostic factor in epithelial ovarian cancer. PLoS ONE 2014, 9, e100816. [Google Scholar] [CrossRef]
- Kim, K.H.; Jo, M.S.; Suh, D.S.; Yoon, M.S.; Shin, D.H.; Lee, J.H.; Choi, K.U. Expression and significance of the TLR4/MyD88 signaling pathway in ovarian epithelial cancers. World J. Surg. Oncol. 2012, 10, 193. [Google Scholar] [CrossRef] [PubMed]
- Gaikwad, S.M.; Thakur, B.; Sakpal, A.; Singh, R.K.; Ray, P. Differential activation of NF-κB signaling is associated with platinum and taxane resistance in MyD88deficient epithelial ovarian cancer cells. Int. J. Biochem. Cell Biol. 2015, 61, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Xiang, F.; Wu, R.; Xu, J.; Ni, Z.; Jiang, J.; Kang, X. MiRNA-149 modulates chemosensitivity of ovarian cancer A2780 cells to paclitaxel by targeting MyD88. J. Ovarian Res. 2015, 8, 48. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Block, M.S.; Vierkant, R.A.; Fogarty, Z.C.; Winham, S.J.; Visscher, D.W.; Kalli, K.R.; Wang, C.; Goode, E.L. The inflammatory microenvironment in epithelial ovarian cancer: A role for TLR4 and MyD88 and related proteins. Tumour Biol. 2016, 37, 13279–13286. [Google Scholar] [CrossRef] [PubMed]
- Meng, Y.; Hu, J.; Chen, Y.; Yu, T.; Hu, L. Silencing MARCH1 suppresses proliferation, migration and invasion of ovarian cancer SKOV3 cells via downregulation of NF-κB and Wnt/β-catenin pathways. Oncol. Rep. 2016, 36, 2463–2470. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Sato, S.; Hemmi, H.; Hoshino, K.; Kaisho, T.; Sanjo, H.; Takeuchi, O.; Sugiyama, M.; Okabe, M.; Takeda, K.; et al. Role of adaptor TRIF in the MyD88-independent toll-like receptor signaling pathway. Science 2003, 301, 640–643. [Google Scholar] [CrossRef] [PubMed]
- Ullah, M.O.; Sweet, M.J.; Mansell, A.; Kellie, S.; Kobe, B. TRIF-dependent TLR signaling, its functions in host defense and inflammation, and its potential as a therapeutic target. J. Leukoc. Biol. 2016, 100, 27–45. [Google Scholar] [CrossRef] [PubMed]
- Muccioli, M.; Sprague, L.; Nandigam, H.; Pate, M.; Benencia, F. Toll-like receptors as novel therapeutic targets for ovarian cancer. ISRN Oncol. 2012, 2012, 642141. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, K.; Yajima, A.; Terashima, Y.; Nozawa, S.; Taketani, Y.; Yakushiji, M.; Noda, K. Phase II clinical study on the effects of recombinant human interleukin-3 onthrombocytopenia after chemotherapy for advanced ovarian cancer. SDZ ILE 964[IL-3] Study Group. J. Immunother. 1999, 22, 539–545. [Google Scholar] [CrossRef]
- Veldhuis, G.J.; Willemse, P.H.; van Gameren, M.M.; Aalders, J.G.; Mulder, N.H.; Mull, B.; Biesma, B.; deVries, E.G. Recombinant human interleukin-3 to dose-intensify carboplatin and cyclophosphamide chemotherapy in epithelial ovarian cancer: A phase I trial. J. Clin. Oncol. 1995, 13, 733–740. [Google Scholar] [CrossRef]
- Rivera-Vargas, T.; Humblin, E.; Végran, F.; Ghiringhelli, F.; Apetoh, L. Th9 cells inanti-tumor immunity. Semin. Immunopathol. 2017, 39, 39–46. [Google Scholar] [CrossRef]
- Park, J.; Li, H.; Zhang, M.; Lu, Y.; Hong, B.; Zheng, Y.; He, J.; Yang, J.; Qian, J.; Yi, Q. Murine Th9 cells promote the survival of myeloid dendritic cells in cancerimmunotherapy. Cancer Immunol. Immunother. 2014, 63, 835–845. [Google Scholar] [CrossRef]
- Singh, M.; Loftus, T.; Webb, E.; Benencia, F. Minireview: Regulatory T Cells and Ovarian Cancer. Immunol. Investig. 2016, 45, 712–720. [Google Scholar] [CrossRef]
- Qu, Q.X.; Xie, F.; Huang, Q.; Zhang, X.G. Membranous and cytoplasmic expression ofPD-L1 in ovarian cancer cells. Cell. Physiol. Biochem. 2017, 43, 1893–1906. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, F.; Bajetto, A.; Florio, T. Role of chemokine network in the development and progression of ovarian cancer: A potential novel pharmacological target. J. Oncol. 2010, 2010, 426956. [Google Scholar] [CrossRef] [PubMed]
- Zsiros, E.; Duttagupta, P.; Dangaj, D.; Li, H.; Frank, R.; Garrabrant, T.; Hagemann, I.S.; Levine, B.L.; June, C.H.; Zhang, L.; et al. The ovarian cancer chemokine landscape isconducive to homing of vaccine-primed and CD3/CD28-costimulated T cells prepared for adoptive therapy. Clin. Cancer Res. 2015, 21, 2840–2850. [Google Scholar] [CrossRef]
- Vulcano, M.; Albanesi, C.; Stoppacciaro, A.; Bagnati, R.; D’Amico, G.; Struyf, S.; Transidico, P.; Bonecchi, R.; Del Prete, A.; Allavena, P.; et al. Dendritic cells as a major source of macrophage-derived chemokine/CCL22 in vitro and in vivo. Eur. J. Immunol. 2001, 31, 812–822. [Google Scholar] [CrossRef]
- Fialová, A.; Partlová, S.; Sojka, L.; Hromádková, H.; Brtnický, T.; Fučíková, J.; Kocián, P.; Rob, L.; Bartůňková, J.; Spíšek, R. Dynamics of T-cell infiltration during thecourse of ovarian cancer: The gradual shift from a Th17 effector cell response toa predominant infiltration by regulatory T-cells. Int. J. Cancer 2013, 132, 1070–1079. [Google Scholar]
- Giuntoli, R.L., 2nd; Webb, T.J.; Zoso, A.; Rogers, O.; Diaz-Montes, T.P.; Bristow, R.E.; Oelke, M. Ovarian cancer-associated ascites demonstrates altered immune environment: Implications for antitumor immunity. Anticancer Res. 2009, 29, 2875–2884. [Google Scholar]
- Lee, L.F.; Hellendall, R.P.; Wang, Y.; Haskill, J.S.; Mukaida, N.; Matsushima, K.; Ting, J.P. IL-8 reduced tumorigenicity of human ovarian cancer in vivo due to neutrophil infiltration. J. Immunol. 2000, 164, 2769–2775. [Google Scholar] [CrossRef]
- Colvin, E.K. Tumor-associated macrophages contribute to tumor progression in ovarian cancer. Front. Oncol. 2014, 4, 137. [Google Scholar] [CrossRef]
- Tino, A.B.; Chitcholtan, K.; Sykes, P.H.; Garrill, A. Resveratrol and acetyl-resveratrol modulate activity of VEGF and IL-8 in ovarian cancer cell aggregates via attenuation of the NF-κB protein. J. Ovarian Res. 2016, 9, 84. [Google Scholar] [CrossRef]
- Milliken, D.; Scotton, C.; Raju, S.; Balkwill, F.; Wilson, J. Analysis of chemokinesand chemokine receptor expression in ovarian cancer ascites. Clin. Cancer Res. 2002, 8, 1108–1114. [Google Scholar]
- Soria, G.; Ben-Baruch, A. The inflammatory chemokines CCL2 and CCL5 in breastcancer. Cancer Lett. 2008, 267, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Ito, T.; Oyama, T.; Kitagawa, T.; Yamori, T.; Fujiwara, H.; Matsuda, H. The direct effect of IL-12 on tumor cells: IL-12 acts directly on tumor cells to activate NF-kB and enhance IFN-g-mediated STAT1 phosphorylation. Biochem. Biophys. Res. Commun. 2001, 280, 503–5121. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lupi, L.A.; Delella, F.K.; Cucielo, M.S.; Romagnoli, G.G.; Kaneno, R.; Nunes, I.d.S.; Domeniconi, R.F.; Martinez, M.; Martinez, F.E.; Fávaro, W.J.; et al. P-MAPA and Interleukin-12 Reduce Cell Migration/Invasion and Attenuate the Toll-Like Receptor-Mediated Inflammatory Response in Ovarian Cancer SKOV-3 Cells: A Preliminary Study. Molecules 2020, 25, 5. https://doi.org/10.3390/molecules25010005
Lupi LA, Delella FK, Cucielo MS, Romagnoli GG, Kaneno R, Nunes IdS, Domeniconi RF, Martinez M, Martinez FE, Fávaro WJ, et al. P-MAPA and Interleukin-12 Reduce Cell Migration/Invasion and Attenuate the Toll-Like Receptor-Mediated Inflammatory Response in Ovarian Cancer SKOV-3 Cells: A Preliminary Study. Molecules. 2020; 25(1):5. https://doi.org/10.3390/molecules25010005
Chicago/Turabian StyleLupi, Luiz Antonio, Flávia Karina Delella, Maira Smaniotto Cucielo, Graziela Gorete Romagnoli, Ramon Kaneno, Iseu da Silva Nunes, Raquel Fantin Domeniconi, Marcelo Martinez, Francisco Eduardo Martinez, Wagner José Fávaro, and et al. 2020. "P-MAPA and Interleukin-12 Reduce Cell Migration/Invasion and Attenuate the Toll-Like Receptor-Mediated Inflammatory Response in Ovarian Cancer SKOV-3 Cells: A Preliminary Study" Molecules 25, no. 1: 5. https://doi.org/10.3390/molecules25010005
APA StyleLupi, L. A., Delella, F. K., Cucielo, M. S., Romagnoli, G. G., Kaneno, R., Nunes, I. d. S., Domeniconi, R. F., Martinez, M., Martinez, F. E., Fávaro, W. J., & Chuffa, L. G. d. A. (2020). P-MAPA and Interleukin-12 Reduce Cell Migration/Invasion and Attenuate the Toll-Like Receptor-Mediated Inflammatory Response in Ovarian Cancer SKOV-3 Cells: A Preliminary Study. Molecules, 25(1), 5. https://doi.org/10.3390/molecules25010005