Abstract
Background: Suicide behavior (SB) has been highly associated with the response to stress and the hypothalamic–pituitary–adrenal (HPA) axis. The aim of this study was to summarize the results obtained in genetic studies that analyzed the HPA axis—stress pathway and SB through a systematic review. Methods: We performed an online search in PubMed, EBSCO, Web of Science, Scopus, and PsycoInfo databases up to May 2021. We followed the PRISMA guidelines for systematic reviews. We included case-control and expression studies that provided data on mRNA expression and single-nucleotide polymorphisms of genes associated with SB. Results: A total of 21,926 individuals participated across 41 studies (not repeats); 34 studies provided data on single-nucleotide polymorphisms in 21,284 participants and 11 studies reported data on mRNA expression in 1034 participants. Ten genes were identified: FKBP5, CRH, CRHBP, CRHR1, CRHR2, NR3C1, NR3C2, SKA2, MC2R, and POMC. Conclusions: Our findings suggest that key stress pathway genes are significantly associated with SB and show potential as biomarkers for SB.
1. Introduction
Suicide is the act of intentionally ending one’s own life, in which the nonfatal suicidal thoughts and behaviors (hereafter called “suicide behaviors”) are classified specifically into three categories: suicide ideation (SI), suicide plan (SP), and suicide attempt (SA). Suicide ideation and attempts can have negative consequences and are strongly predictive of deaths by suicide [1,2].
Globally, there are approximately 11.4 suicides per 100,000 people, and the suicidality rates are high among those with psychiatric disorders such as depression and anxiety. Moreover, higher rates of SI and SA are observed among females, but higher rates of suicide deaths are observed in males [3,4].
The interaction of internal and external stressors with the psychopathological and cognitive traits form a diathesis for suicide risk [5]. The stress-diathesis model depicts suicide behavior (SB) as a consequence of an interaction between acute stressors and a set of suicide-related traits; this interaction moderates the likelihood of SB in response to stressors [6,7].
The response to stress involves a more immediate short-term noradrenergic system response, as well as a more enduring hypothalamic–pituitary–adrenal (HPA) axis response [8]. The HPA system is activated and glucocorticoids (GCs) are released into the systemic blood flow reaching every organ of the body. GCs exert their effects through the mineralocorticoid receptor (MR) and the glucocorticoid receptor (GR), both of which are nuclear receptors [9,10]. When a stressor (either physiological or psychological) is encountered, the hypothalamus releases corticotrophin-releasing hormone (CRH) and vasopressin; when these hormones reach the anterior pituitary, they stimulate the corticotropic cells to release adrenocorticotropic hormone (ACTH). Blood-borne ACTH circulates out of the central nervous system and reaches the adrenal glands above the kidneys, to upregulate the production of GCs, including cortisol (Figure 1).
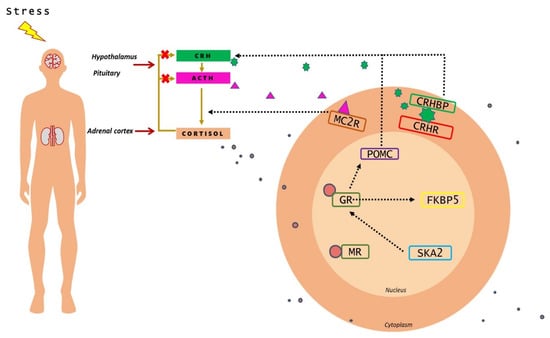
Figure 1.
Representative image of HPA axis and the stress pathway genes implicated in suicide behavior. Upon perception of stress, CRH is released from the hypothalamus, which promotes the synthesis and release of ACTH from the pituitary. ACTH, in turn, increases the release of cortisol from the adrenal glands. Through negative feedback, cortisol inhibits the hormone release from the hypothalamus and anterior pituitary glands. Inside the cell, cortisol causes the activation of molecular mechanisms that regulate the impact of the HPA axis.
The ACTH derives from the cleavage of the precursor hormone pro-opiomelanocortin (POMC) by prohormone convertase enzymes. ACTH activates the production and release of cortisol from the zona fasciculata of the adrenal cortex via the melanocortin receptor MC2R [11]. The CRH is regarded as the principal mediator of the stress response in the brain, and its actions are moderated by a high-affinity binding protein (CRHBP) that modulates CRH-mediated activation of CRH receptors in brain and periphery (Figure 1). CRHR1 is the key receptor for CRH-mediated ACTH release in pituitary response to stress. CRHR1 plays a critical role in the acute phase of stress-induced HPA response and CRHR2 is involved in the recovery phase [12,13].
Following stress, cortisol binds to brain tissue with affinity to GRs (encoded by NR3C1 gene) or MRs (encoded by NR3C2 gene). MRs are occupied under basal glucocorticoid conditions, whereas GR occupancy is increased as cortisol levels rise following stress. Intracellularly, GR binds to cochaperone FKBP5, an important functional regulator of GR sensitivity; when it is bound to the receptor complex, cortisol binds with lower affinity and nuclear translocation of the receptor is less efficient. FKBP5 mRNA and protein expression are induced by GR activation; high intracellular levels of FKBP5 lower the binding affinity of GR for glucocorticoids leading to GR resistance [14,15]. The SKA2 protein has been implicated in enabling GR nuclear transactivation [16] (Figure 1), therefore impairing the negative feedback of HPA-axis.
Several studies have demonstrated that stress and a dysregulated HPA axis activity are important additional risk factors of SB [17,18]. Alterations in stress-induced regulation via any genetic factor could have important influences on SB. Some stress pathways genes that have been studied in SB include NR3C1, [19,20]; FKBP5 [21,22]; CRHR1 [23,24]; and SKA2 gene [25,26]. Candidate gene studies have shown differentially expressed mRNA patterns and single-nucleotide polymorphisms (SNPs) in Caucasian and African American populations associated with SB [27,28].
Thus, a deep understanding of genetic associations underpinning SB is of paramount importance in developing effective treatment interventions. There is increasing evidence that genes regulating HPA axis have effects on SB.
The dysregulation of HPA axis activity is associated with other systems implicated in suicide, including opioids, serotonin, glutamate systems, lipid status, inflammatory pathways, and neurogenesis; therefore, elucidating the dysregulation of the stress system could help us understand the importance of HPA axis in SB [29,30]. To test this idea, we identified and summarized studies that examined stress pathways genes associated with SB through a systematic review. We included case-control and genetic expression studies that provided data on mRNA expression and single-nucleotide polymorphisms; then, we described the relationship between these genes and SB.
2. Methods
2.1. Search Strategy
A search for studies that investigated the association between HPA axis—stress pathway genes and SB was conducted up to May 2021 following the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines (Supplementary Table S1). The systematic search was performed using five online databases: PubMed, EBSCO, Science Direct, PsychInfo, and Scopus in order to find and include the most pertinent literature.
Keywords included in the literature search were: (1) for SB: suicide, suicide plan, suicide ideation, suicide attempt; (2) for stress: HPA, cortisol, stress pathway; and (3) for genetic influences: gene, genotype, SNP, polymorphism, gene expression, and candidate gene. The scope of the online search was further expanded by assessing bibliographic references of the eligible full text articles in order to detect other relevant studies.
2.2. Inclusion Criteria and Data Extraction
The studies were initially retrieved as title and abstract and screened for eligibility. To be selected, the articles had to fulfill the inclusion criteria: (1) original article, (2) peer-reviewed research, (3) articles published between 2001 and 2021, (4) case-control and expression studies that provided data on mRNA expression and single-nucleotide polymorphisms of stress pathway genes associated with SB, and (5) to be written in English.
After removing duplicates and scanning titles and abstracts, articles that met the inclusion criteria were reviewed. The following data were extracted from each eligible article: authors, year of publication, studied population (number of participants, ethnicity, diagnostic), tissue source, gene name, polymorphism, gene expression, and main findings of the study. Two authors (YHD and TBGC) conducted all screening analysis and data extraction.
2.3. Quality Assessment of Primary Studies
The methodological quality of the included studies was evaluated using the Newcastle–Ottawa scale (NOS). The NOS scale gives scores that range from zero to nine, giving a point to each accomplished item and categorizing the studies as high quality (score 7–9), moderate quality (score 4–6), or poor quality (score 0–3). The tool assesses the studies based on three dimensions: selection, compatibility, exposure, or outcome. Authors rated the article independently and discussed the ratings.
2.4. Data Synthesis
Significant information from the studies included was carefully organized; the phenotypic outcomes that were considered as SB in this systematic review were: suicide ideation (SI), suicide attempt (SA), suicide plan (SP), and completed suicide (CS). The most promising genes were extracted from the results and the main findings from the texts; tables summarized the study characteristics.
3. Results
3.1. Study Selection
Figure 2 highlights the identification and selection process following the PRISMA statement. The search in PubMed, EBSCO, Science Direct, PsychInfo, and Scopus databases resulted in a total of 176 identified articles, and 103 articles remained after removal of duplicate records. Then, 62 unrelated articles were excluded; finally, 41 articles were accepted for the systematic review based on our inclusion and exclusion criteria. The analysis outcomes of the selected publications are shown in Table 1 and Table 2.
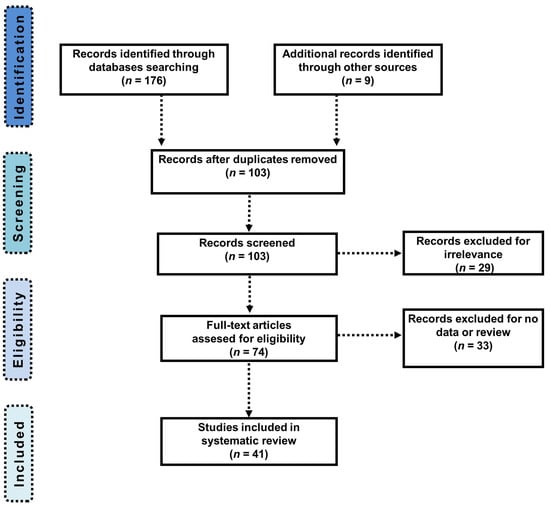
Figure 2.
PRISMA flow chart presenting the articles identification and selection processes.

Table 1.
Detailed analysis of the selected publications regarding the association between single-nucleotide polymorphisms (SNP) in HPA genes and the pathogenesis of suicide behavior.
Table 2.
Characteristics of the included publications that evaluated HPA genes expression (mRNA) in the pathogenesis of suicide behavior.
3.2. Studies Caracteristics
A total of 21,926 individuals (repeated individuals were excluded) participated across the 41 studies. Thirty-three studies included a control group and 8 studies only evaluated cases. In the majority of studies, controls were described as healthy individuals. The sample sizes ranged from 7 to 3623. All the studies were conducted between 2001 and 2021.
The main psychiatric disorders present in individuals with SB were major depression disorder, bipolar disorder, and substance use dependence. Tissue sources utilized for genotyping or genetic expression analyses were blood, brain, saliva, and buccal cells. The methods used for measuring trauma exposure also differed across studies, including the childhood trauma questionnaire (CTQ), early life events scale (ELES), life events checklist (LEC), and the adverse childhood experiences questionnaire (ACE). A comprehensive description of the studies characteristics is presented in Table 1 and Table 2. Finally, the quality assessment using NOS scale revealed a mean score of 7.17 (ranging from 6 to 9) for SB studies (Table 3).
Table 3.
NOS scores of 41 studies included in the systematic review.
3.3. Phenotypes and Genes
Ten genes were identified: FKBP5, CRH, CRHBP, CRHR1, CRHR2, NR3C1, NR3C2, SKA2, MC2R, and POMC. Of the included studies that analyzed SNPs in SB, SA was the phenotype most frequently evaluated, followed by SI and CS. In total, 264 DNA SNPs comprised in 10 different genes were analyzed across the studies included in this review (Table 1).
We observed an upregulation of CRH and SKA2 genes; however, findings on mRNA expression were not consistent across studies, as some studies indicated a downregulation of SKA2 or did not find important changes. The NR3C1, FKBP5, CRH1, and SKA2 genes were the most frequently studied in expression studies (Table 2). Finally, CS and SI were the phenotypes most evaluated in the studies that analyzed mRNA/gene expression levels.
3.4. Synthesis of Results
3.4.1. FKBP5 Gene
Eighteen studies [19,21,22,28,31,32,33,34,35,36,37,38,39,40,41,42,43,44] analyzed the association between FKBP5 SNPs and SB phenotypes comprising 4239 cases and 9646 controls. The first study conducted in 2007 by Papiol et al. [31] highlighted a significant association between the rs1360780 SNP and SA. Significant associations were also identified between rs3800373, rs3777747, rs2766533, rs4713902, rs9470080, rs1043805, and rs9296158 SNPs and SB in other studies. Additionally, rs3800373 was significantly associated with stress exposure.
On the other hand, mRNA expression levels and SB were analyzed in five studies [14,41,56,57,58] including 204 cases and 145 controls. The FKBP5 gene was downregulated in samples of brain and blood, as well as mRNA which was consistently correlated with heroin, painkillers, and ecstasy use [58].
3.4.2. CRH Gene
Four studies [27,28,38,45] examined the CRH SNPs and suicide attempt, including 1395 cases and 1496 (Table 1). All the studies reported that SNPs in this gene were not significantly associated with SA.
Only two studies [55,60] analyzed the expression levels of CRH gene in CS including 29 cases and 19 controls. Both studies demonstrated that CRH gene was upregulated in brain tissue of CS compared with brain tissue from controls.
3.4.3. CRHBP Gene
Five studies [19,27,28,31,38] investigated the CRHBP SNPs, each with significant findings (1016 cases and 1592 controls). In 2010, De Luca et al. [27] observed that the heterozygous genotype of rs1875999 was significantly associated with SA and risk of SA. Additionally, Roy et al. [28] found that rs6453267, rs7728378, and rs10474485 showed a nominally significant interaction with the continuous CTQ score to predict SA. No changes in the CRHBP gene expression between cases and controls were observed [55,56,60].
3.4.4. CRHR1 Gene
Sixteen studies [19,23,24,27,28,31,38,42,44,45,46,47,48,49,50,51] evaluated the association between the CRHR1 SNPs and SB phenotypes in 3718 cases and 4539 controls. Significant associations were identified between s7209436, rs110402, rs16940665, rs4792887, rs12936511, rs1396862, rs878886, and rs242948 SNPs and SB.
In a study conducted by Pawlak et al. [49], the rs16940665 polymorphism was associated with males who had attempted suicide and had major depression disorder. Ludwig et al. [51] indicated that there was a significant gene-environment-interactions for rs7209436 and rs110402 SNPs, reflecting the impact of childhood trauma and CRHR1 polymorphisms on previous SA.
Four studies [55,56,59,60] analyzed the CRHR1 gene expression levels in CS and SI (52 cases and 46 controls). A CRHR1 downregulation was observed in only one study associated with CS [60].
3.4.5. CRHR2 Gene
Seven studies [19,23,27,28,31,38,47] evaluated the association between CRHR2 SNPs and SB in 1407 cases and 1775 controls. Allele G carriers of rs2270007 showed a worse overall response to citalopram in follow-up time and showed a 2.93 increased risk of nonresponding to citalopram at week 4 of treatment. Additionally, sexual abuse in childhood and childhood emotional neglect interacted with the rs255098 to modulate adult decision making in SA [31,47].
Three studies [55,59,60] reported no changes in the mRNA levels of this gene in CS (38 cases and 26 controls).
3.4.6. NR3C1 Gene
Eight studies [19,20,27,36,38,41,44,48] (783 cases and 2570 controls) analyzed the association between NR3C1 SNPs and SB phenotypes. The first one was conducted in 2010 by De Luca et al. [27] and highlighted a significant association between rs6196 SNP and SA. Other associations were identified between rs9324924, rs2963155, and rs41423247 SNPs and SB in other studies.
mRNA expression levels and SB were analyzed in eight studies [14,41,53,54,55,56,57,58] including 186 cases and 198 controls. NR3C1 gene was observed to be downregulated in samples of brain and blood. mRNA was significantly reduced in individuals who CS, many of whom had history of childhood abuse in comparison with non-abused CS [53]. Cortisol levels were associated with mRNA [57] and with the expression levels of rs10052957, rs6190 and rs41423247 SNPs [54].
3.4.7. NR3C2 Gene
Two studies [19,36] examined the NR3C2 SNPs and SB in 348 cases and 228 controls. All studies reported that SNPs in this gene were not significantly associated with SB. No data were reported for expression.
3.4.8. SKA2 Gene
Four studies [25,41,44,52] examined the association between SKA2 SNPs and SB in 1675 cases and 1320 controls. rs8082544 and rs7502947 showed an association with CS [41] as well as significant interactions for SKA2 3′-UTR DNA methylation, while the rs7208505 SNP was associated with SI and SA [25].
Four studies [26,41,57,58] analyzed SKA2 gene expression levels and CS in 179 cases and 193 controls. Data showed a downregulation of SKA2 gene; however, findings in mRNA expression were not consistent across studies, as some studies indicated an upregulation of SKA2 or no changes [57,58]. No associations with trauma exposure and concentrations of cortisol were indicated.
3.4.9. MC2R Gene
Three studies [19,27,38] evaluated the association between the MC2R SNPs and SB phenotypes in 841 cases and 807 controls. All studies reported that the SNPs in this gene were not significantly associated with SB. No data were reported for expression.
3.4.10. POMC Gene
Two studies [19,38] investigated the POMC SNPs (in 760 cases and 657 controls) in association with SB. Both studies reported that this gene was not associated with SB and no data were reported for expression.
4. Discussion
This systematic review aimed to summarize the findings of genetic variants that have been associated with SB. We reviewed 41 publications that gathered 10 promising genes associated with SB: FKBP5, CRH, CRHBP, CRHR1, CRHR2, NR3C1, NR3C2, MC2R, SKA2, and POMC.
4.1. Main Findings
The study of polymorphisms may contribute, at least in part, to explain the alterations observed in SB; additionally, different polymorphisms could alter the genes expression levels and HPA activity in response to stress [14,56]. Our results are in agreement with studies that utilize others approximation. A recent study using a network meta-analysis observed that FKBP5 gene in union with other mediators could increase the risk of suicide behavior [61]. Additionally, studies suggest that these mediators could be childhood victimization [21]. They found that FKBP5, CRHBP, and childhood victimization could increase the risk for suicide behavior. Additionally, several studies indicated that genetic and epigenetic variations in different regions of FKBP5 gene may contribute, at least in part, to the FKBP5 alterations observed in SB. Then, the positive evidence in the literature and our results in the present systematic review suggest a possible role of FKBP5 gene in suicidal behavior.
Second, we found that other genes such as the CRH family (CRH, CRHR1, and CRHR2 genes) and CRHBP gene (an antagonist of the stress hormone CRH) showed conflicting results between SNPs and mRNA expression levels. As an example, in the frontopolar cortex, mRNA for CRHR1, but not CRHR2 receptors were reduced in brains of individuals who died by suicide, possibly secondary to high levels of CRH activity [60]. This could be partially explained by ethnic discrepancies or studies with small sample sizes observed in the studies.
Third, our findings suggest that GR (encoded by NR3C1) might underlie a contribution of HPA axis to SB phenotypes. Functional polymorphisms within the NR3C1 gene may impact its gene expression [54]; moreover, mRNA was positively and moderately correlated with hair cortisol concentrations and also negatively correlated with childhood abuse [57]. However, we observed that NR3C2 gene (mineralocorticoid receptor) did not play a role in SB [19].
Fourth, no significant associations between MC2R and POMC genes with SB were reported. Alternatively, polymorphisms in these genes might be in high linkage disequilibrium with the causative variants. Studies have shown that epigenetics, especially DNA methylation, play an important role in the occurrence, development, and progression of psychiatric disorders. In addition, research on epigenetics proves that environmental factors are also closely related to the occurrence of diseases [62,63]. Nonetheless, literature on these genes is extremely poor, and, therefore, further research is required to confirm or reject the hypothesis of their non-association with SB.
Fith, while most of the articles examined focused on one or a few candidate genes, SB is a complex and polygenic disease with each genetic variant likely to be contributing a small percentage to disease. Then, studies the GWAS studies that analyze the specially the genes implicated in the HPA axis are necessary.
Finally, we observed a variation across studies in terms of psychiatric disorders and exposure to traumatic events. Individuals with serious mental illnesses (e.g., schizophrenia, bipolar disorder, major depressive disorder) have significantly higher suicide rates than the general population; additionally, the heterogeneity of the findings could indicate that the presence of a mental illness as well as the expression of genetic and environmental effects (traumatic events) could contribute to different phenotypes. This also highlights the importance of conducting psychiatric diagnostic stratified studies.
4.2. HPA Axis and Suicide Behavior
A dysfunction of the hypothalamic-pituitary-adrenal (HPA) axis is considered a possible pathogenic background of suicide. Because some polymorphisms regulate the genic expression levels that lead to GR resistance and impaired negative feedback, we could speculate that some alleles cause a slower return to baseline of stress-induced cortisol levels, increasing the risk for psychiatric disorders such as SB. As gene expression is responsive to cortisol, genetic modifications that alter this interaction could modulate the effects of environmental stressors on HPA axis [28,44].
Altered mechanisms may exert deleterious effects on the development of brain structures implicated in suicide behavior. In both of these contexts, genes may contribute to alter neurobiological functions, and a maladaptive prolonged stress response may render individuals more vulnerable to suicide [18]. The specific pattern of this intracellular crosstalk may vary across tissues and may contribute to the pleiotropic consequences of HPA axis dysregulation in suicide [64,65]. Therefore, elucidating the molecular underpinnings of this variability is of great relevance for developing individualized prevention strategies and treatments for individuals with SB. Finally, drugs targeting the function of HPA axis genes may potentially serve to prevent negative long-term effects of stress.
4.3. Strengths and Limitations
This is the first systematic review to explore the association between stress pathways (particularly the HPA axis) genes and SB. While some methodological weaknesses were observed, most studies were well designed and conducted according to the NOS scale. Nonetheless, this systematic review has some limitations. Findings within this review were at times conflicting. Incongruities may be partly explained due to the differences in methodological aspects such the participant characteristics. For example, the presence of a psychiatric disorder, current use of medication, and differences in the racial/ethnic component may affect the susceptibility to SB. Suicide is a complex disease involved in the regulation of a series of genetic factors besides HPA axis genes. As a multifactorial disease, the risk of developing it is closely related to various elements, and not just a single factor. Second, exposition to adversities during childhood influence the development of SB; however, several studies not taken this characteristic into consideration. Third, there is a lack of endophenotype data that may help to understand the association between genes and SB. Another drawback was that several studies examined a small sample population, and many did not establish statistical significance due to this. Finally, we cited articles written in English only, thus we could have missed important articles in other languages.
4.4. Future Directions
Future research studies should focus on the simultaneous analysis of the widest possible range of genes and their interactions. It is important to consider epigenetic variation of gene activity that can occur as a reaction to external factors. Populations should be divided by sex, as SB is different between females and males. Further still, more extensive explorations of the candidate genes highlighted in this review should provide further insight into the pathogenesis of suicide behavior.
5. Conclusions
This review identified and systematically compiled key stress pathways (particularly the HPA axis) genes that are significantly associated with SB. In total, 10 genes that predicted suicide risk were identified. The outcomes of this review could help to further illuminate the genetic basis of suicide behavior. Further research into this field is definitely necessary to achieve a better understanding of the pathogenesis of SB phenotypes.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/genes12101608/s1, Table S1: PRISMA checklist.
Author Contributions
Conceptualization: Y.H.-D. and A.D.G.-M.; data curation: C.A.T.-Z. and H.N.; investigation: Y.H.-D. and T.B.G.-C.; methodology: I.E.J.-R.; writing–original draft: Y.H.-D. and A.D.G.-M.; writing—review and editing: C.A.T.-Z. and M.L.L.-N. All authors have read and agreed to the published version of the manuscript.
Funding
No funding source.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare that they have no competing interests.
References
- Nock, M.K.; Borges, G.; Bromet, E.J.; Cha, C.B.; Kessler, R.C.; Lee, S. Suicide and Suicidal Behavior. Epidemiol. Rev. 2008, 30, 133–154. [Google Scholar] [CrossRef] [PubMed]
- Silverman, M.M.; Berman, A.L.; Sanddal, N.D.; O’Carroll, P.W.; Joiner, T.E. Rebuilding the Tower of Babel: A Revised Nomenclature for the Study of Suicide and Suicidal Behaviors Part 2: Suicide-Related Ideations, Communications, and Behaviors. Suicide Life-Threat. Behav. 2007, 37, 264–277. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, S. Epidemiology of Suicide and the Psychiatric Perspective. Int. J. Environ. Res. Public Health 2018, 15, 1425. [Google Scholar] [CrossRef] [PubMed]
- Turecki, G.; Brent, D.A. Suicide and suicidal behaviour. Lancet 2016, 387, 1227–1239. [Google Scholar] [CrossRef]
- Van Heeringen, K. Frontiers in Neuroscience. Stress–Diathesis Model of Suicidal Behavior. In The Neurobiological Basis of Suicide; Dwivedi, Y., Ed.; CRC Press: Boca Raton, FL, USA; Taylor & Francis: Boca Raton, FL, USA, 2012. Available online: https://pubmed.ncbi.nlm.nih.gov/23035289/ (accessed on 15 July 2021).
- Mann, J.J.; Rizk, M.M. A Brain-Centric Model of Suicidal Behavior. Am. J. Psychiatry 2020, 177, 902–916. [Google Scholar] [CrossRef] [PubMed]
- Baud, P. Personality traits as intermediary phenotypes in suicidal behavior: Genetic issues. Am. J. Med. Genet. Part C Semin. Med. Genet. 2005, 133C, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the Hypothalamic-Pituitary-Adrenocortical Stress Response. Compr. Physiol. 2016, 6, 603–621. [Google Scholar] [CrossRef] [PubMed]
- De Kloet, E.R.; Joels, M.; Holsboer, F. Stress and the brain: From adaptation to disease. Nat. Rev. Neurosci. 2005, 6, 463–475. [Google Scholar] [CrossRef]
- Kloet, E.R. Hormones and the Stressed Brain. Ann. N.Y. Acad. Sci. 2004, 1018, 1–15. [Google Scholar] [CrossRef]
- Lim, C.T.; Khoo, B.; Feingold, K.R.; Anawalt, B.; Boyce, A.; Chrousos, G.; de Herder, W.W.; Dhatariya, K.; Dungan, K.; Grossman, A.; et al. Normal Physiology of ACTH and GH Release in the Hypothalamus and Anterior Pituitary in Man. In Endotext; MDText.com, Inc.: South Dartmouth, MA, USA, 2020. Available online: https://pubmed.ncbi.nlm.nih.gov/25905340/ (accessed on 21 May 2021).
- Reul, J.M.M.; Holsboer, F. On the role of corticotropin-releasing hormone receptors in anxiety and depression. Dialog Clin. Neurosci. 2002, 4, 31–46. [Google Scholar] [CrossRef]
- Ketchesin, K.; Stinnett, G.S.; Seasholtz, A.F. Corticotropin-releasing hormone-binding protein and stress: From invertebrates to humans. Stress 2017, 20, 449–464. [Google Scholar] [CrossRef]
- Pérez-Ortiz, J.M.; García-Gutiérrez, M.S.; Navarrete, F.; Giner, S.; Manzanares, J. Gene and protein alterations of FKBP5 and glucocorticoid receptor in the amygdala of suicide victims. Psychoneuroendocrinology 2013, 38, 1251–1258. [Google Scholar] [CrossRef]
- Fries, G.R.; Gassen, N.; Schmidt, U.; Rein, T. The FKBP51-Glucocorticoid Receptor Balance in Stress-Related Mental Disorders. Curr. Mol. Pharmacol. 2015, 9, 126–140. [Google Scholar] [CrossRef]
- Rice, L.; Waters, E.C.; Eccles, J.; Garside, H.; Sommer, P.; Kay, P.; Blackhall, F.H.; Zeef, L.; Telfer, B.; Stratford, I.; et al. Identification and functional analysis of SKA2 interaction with the glucocorticoid receptor. J. Endocrinol. 2008, 198, 499–509. [Google Scholar] [CrossRef]
- O’Connor, D.B.; Gartland, N.; O’Connor, R.C. Stress, cortisol and suicide risk. Dev. Psychopathol. 2020, 152, 101–130. [Google Scholar] [CrossRef]
- Berardelli, I.; Serafini, G.; Cortese, N.; Fiaschè, F.; O’Connor, R.C.; Pompili, M. The Involvement of Hypothalamus–Pituitary–Adrenal (HPA) Axis in Suicide Risk. Brain Sci. 2020, 10, 653. [Google Scholar] [CrossRef]
- Segura, A.G.; Mitjans, M.; Jimenez, E.; Fatjó-Vilas, M.; Ruiz, V.; Saiz, P.; García-Portilla, M.; Gonzalez-Blanco, L.; Bobes, J.; Vieta, E.; et al. Association of childhood trauma and genetic variability of CRH-BP and FKBP5 genes with suicidal behavior in bipolar patients. J. Affect. Disord. 2019, 255, 15–22. [Google Scholar] [CrossRef]
- Park, S.; Hong, J.P.; Lee, J.-K.; Park, Y.-M.; Park, Y.; Jeon, J.; Ahn, M.H.; Yoon, S.C. Associations between the neuron-specific glucocorticoid receptor (NR3C1) Bcl-1 polymorphisms and suicide in cancer patients within the first year of diagnosis. Behav. Brain Funct. 2016, 12, 22. [Google Scholar] [CrossRef]
- Berent, D.; Szymańska, B.; Kulczycka-Wojdala, D.; Macander, M.; Pawłowska, Z.; Wojnar, M. The role of childhood adversities, FKBP5, BDNF, NRN1, and generalized self-efficacy in suicide attempts in alcohol-dependent patients. Pharmacol. Rep. Pr. 2020, 72, 730–743. [Google Scholar] [CrossRef]
- Hernández-Díaz, Y.; Genis-Mendoza, A.; Fresán, A.; González-Castro, T.; Tovilla-Zárate, C.; Juárez-Rojop, I.; López-Narváez, M.; Martínez-Magaña, J.; Nicolini, H. Knowledge, Emotions and Stressors in Front-Line Healthcare Workers during the COVID-19 Outbreak in Mexico. Int. J. Environ. Res. Public Heal. 2021, 18, 5622. [Google Scholar] [CrossRef]
- Sanabrais-Jiménez, M.A.; Sotelo-Ramirez, C.E.; Ordoñez-Martinez, B.; Jiménez-Pavón, J.; Ahumada-Curiel, G.; Piana-Diaz, S.; Flores-Flores, G.; Flores-Ramos, M.; Jiménez-Anguiano, A.; Camarena, B. Effect of CRHR1 and CRHR2 gene polymorphisms and childhood trauma in suicide attempt. J. Neural Transm. 2019, 126, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Wasserman, D.; Wasserman, J.; Rozanov, V.; Sokolowski, M. Depression in suicidal males: Genetic risk variants in theCRHR1gene. Genes Brain Behav. 2009, 8, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Kaminsky, A.Z.; Wilcox, H.; Eaton, W.W.; Van Eck, K.; Kilaru, V.; Jovanovic, T.; Klengel, T.; Bradley, B.P.; Binder, E.B.; Ressler, K.J.; et al. Epigenetic and genetic variation at SKA2 predict suicidal behavior and post-traumatic stress disorder. Transl. Psychiatry 2015, 5, e627. [Google Scholar] [CrossRef] [PubMed]
- Pandey, G.N.; Rizavi, H.S.; Zhang, H.; Bhaumik, R.; Ren, X. The Expression of the Suicide-Associated Gene SKA2 Is Decreased in the Prefrontal Cortex of Suicide Victims but Not of Nonsuicidal Patients. Int. J. Neuropsychopharmacol. 2016, 19. [Google Scholar] [CrossRef]
- De Luca, V.; Tharmalingam, S.; Zai, C.; Potapova, N.; Strauss, J.; Vincent, J.; Kennedy, J. Association of HPA axis genes with suicidal behaviour in schizophrenia. J. Psychopharmacol. 2008, 24, 677–682. [Google Scholar] [CrossRef]
- Roy, A.; Hodgkinson, C.A.; DeLuca, V.; Goldman, D.; Enoch, M.-A. Two HPA axis genes, CRHBP and FKBP5, interact with childhood trauma to increase the risk for suicidal behavior. J. Psychiatr. Res. 2012, 46, 72–79. [Google Scholar] [CrossRef]
- Pjevac, M.; Pregel, J.P. Neurobiology of suicidal behaviour. Psychiatr. Danub. 2012, 24, 336–341. [Google Scholar]
- Duval, F.; Mokrani, M.-C.; Monreal, J.; Weiss, T.; Fattah, S.; Hamel, B.; Macher, J.-P. Interaction between the serotonergic system and HPA and HPT axes in patients with major depression: Implications for pathogenesis of suicidal behavior. Dialog Clin. Neurosci. 2002, 4, 417. [Google Scholar] [CrossRef]
- Papiol, S.; Arias, B.; Gastó, C.; Gutiérrez, B.; Catalán, R.; Fañanás, L. Genetic variability at HPA axis in major depression and clinical response to antidepressant treatment. J. Affect. Disord. 2007, 104, 83–90. [Google Scholar] [CrossRef]
- Willour, V.L.; Chen, H.; Toolan, J.; Belmonte, P.; Cutler, D.J.; Goes, F.S.; Zandi, P.P.; Lee, R.; MacKinnon, D.F.; Mondimore, F.M.; et al. Family-based association of FKBP5 in bipolar disorder. Mol. Psychiatry 2008, 14, 261–268. [Google Scholar] [CrossRef]
- Brent, D.; Melhem, N.; Ferrell, R.; Emslie, G.; Wagner, K.D.; Ryan, N.; Vitiello, B.; Birmaher, B.; Mayes, T.; Zelazny, J.; et al. Association of FKBP5 Polymorphisms with Suicidal Events in the Treatment of Resistant Depression in Adolescents (TORDIA) Study. Am. J. Psychiatry 2010, 167, 190–197. [Google Scholar] [CrossRef]
- Roy, A.; Gorodetsky, E.; Yuan, Q.; Goldman, D.; Enoch, M.A. Interaction of FKBP5, a stress-related gene, with childhood trauma increases the risk for attempting suicide. Neuropsychopharmacoly 2010, 35, 1674–1683. [Google Scholar] [CrossRef]
- Perroud, N.; Bondolfi, G.; Uher, R.; Gex-Fabry, M.; Aubry, J.-M.; Bertschy, G.; Malafosse, A.; Kosel, M. Clinical and genetic correlates of suicidal ideation during antidepressant treatment in a depressed outpatient sample. Pharmacogenomics 2011, 12, 365–377. [Google Scholar] [CrossRef]
- Supriyanto, I.; Sasada, T.; Fukutake, M.; Asano, M.; Ueno, Y.; Nagasaki, Y.; Shirakawa, O.; Hishimoto, A. Association of FKBP5 gene haplotypes with completed suicide in the Japanese population. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 252–256. [Google Scholar] [CrossRef]
- Leszczyńska-Rodziewicz, A.; Szczepankiewicz, A.; Skibińska, M.; Pawlak, J.; Dmitrzak-Węglarz, M.; Hauser, J.; Narozna, B. Possible association between haplotypes of the FKBP5 gene and suicidal bipolar disorder, but not with melancholic depression and psychotic features, in the course of bipolar disorder. Neuropsychiatr. Dis. Treat. 2014, 10, 243–248. [Google Scholar] [CrossRef][Green Version]
- Breen, M.E.; Seifuddin, F.; Zandi, P.P.; Potash, J.B.; Willour, V.L. Investigating the role of early childhood abuse and HPA axis genes in suicide attempters with bipolar disorder. Psychiatr. Genet. 2015, 25, 106–111. [Google Scholar] [CrossRef]
- Fudalej, S.; Kopera, M.; Wołyńczyk-Gmaj, D.; Fudalej, M.; Krajewski, P.; Wasilewska, K.; Szymański, K.; Chojnicka, I.; Podgórska, A.; Wojnar, M.; et al. Association between FKBP5 Functional Polymorphisms and Completed Suicide. Neuropsychobiology 2015, 72, 126–131. [Google Scholar] [CrossRef]
- Breen, M.E.; Gaynor, S.C.; Monson, E.T.; De Klerk, K.; Parsons, M.G.; Braun, T.A.; DeLuca, A.P.; Zandi, P.P.; Potash, J.B.; Willour, V.L. Targeted Sequencing of FKBP5 in Suicide Attempters with Bipolar Disorder. PLoS ONE 2016, 11, e0169158. [Google Scholar] [CrossRef]
- Yin, H.; Galfalvy, H.; Pantazatos, S.P.; Huang, Y.-Y.; Rosoklija, G.B.; Dwork, A.J.; Burke, A.; Arango, V.; Oquendo, M.A.; Mann, J.J. Glucocorticoid Receptor-Related Genes: Genotype and Brain Gene Expression Relationships to Suicide and Major Depressive Disorder. Depress. Anxiety 2016, 33, 531–540. [Google Scholar] [CrossRef]
- Mirkovic, B.; Cohen, D.; Laurent, C.; Lasfar, M.; Marguet, C.; Gerardin, P. A case-control association study of 12 candidate genes and attempted suicide in French adolescents. Int. J. Adolesc. Med. Health 2017, 32. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, X.-Z.; Benedek, D.M.; Fullerton, C.S.; Forsten, R.D.; Naifeh, J.; Li, X.; Wynn, G.; Russell, D.W.; Ursano, R.J. Genetic predictor of current suicidal ideation in US service members deployed to Iraq and Afghanistan. J. Psychiatr. Res. 2019, 113, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Nobile, B.; Ramoz, N.; Jaussent, I.; Dubois, J.; Guillaume, S.; Gorwood, P.; Courtet, P. Polymorphisms of stress pathway genes and emergence of suicidal ideation at antidepressant treatment onset. Transl. Psychiatry 2020, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wasserman, D.; Sokolowski, M.; Rozanov, V.; Wasserman, J. The CRHR1 gene: A marker for suicidality in depressed males exposed to low stress. Genes Brain Behav. 2007, 7, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Ben-Efraim, Y.J.; Wasserman, D.; Sokolowski, M. Gene-environment interactions between CRHR1 variants and physical assault in suicide attempts. Genes Brain Behav. 2011, 10, 663–672. [Google Scholar] [CrossRef]
- Guillaume, S.; Perroud, N.; Jollant, F.; Jaussent, I.; Olié, E.; Malafosse, A.; Courtet, P. HPA axis genes may modulate the effect of childhood adversities on decision-making in suicide attempters. J. Psychiatr. Res. 2013, 47, 259–265. [Google Scholar] [CrossRef]
- Leszczynska-Rodziewicz, A.; Maciukiewicz, M.; Szczepankiewicz, A.; Pogłodziński, A.; Hauser, J. Association between OPCRIT dimensions and polymorphisms of HPA axis genes in bipolar disorder. J. Affect. Disord. 2013, 151, 744–747. [Google Scholar] [CrossRef]
- Pawlak, J.; Dmitrzak-Weglarz, M.; Wiłkość-Dębczyńska, M.; Szczepankiewicz, A.; Leszczynska-Rodziewicz, A.; Zaremba, D.; Kapelski, P.; Rajewska-Rager, A.; Hauser, J. Suicide behavior as a quantitative trait and its genetic background. J. Affect. Disord. 2016, 206, 241–250. [Google Scholar] [CrossRef]
- Bastos, C.R.; Gazal, M.; Quevedo, L.D.A.; Costa, J.L.; Wiener, C.D.; Jansen, K.; de Mola, C.L.; Oses, J.P.; Souza, L.D.; Portela, L.V.; et al. Polymorphism in CRHR1 gene affects the IL-1β levels in suicidal attempters. J. Psychiatr. Res. 2017, 86, 34–38. [Google Scholar] [CrossRef]
- Ludwig, B.; Kienesberger, K.; Carlberg, L.; Swoboda, P.; Bernegger, A.; Koller, R.; Wang, Q.; Inaner, M.; Zotter, M.; Kapusta, N.D.; et al. Influence of CRHR1 Polymorphisms and Childhood Abuse on Suicide Attempts in Affective Disorders: A GxE Approach. Front. Psychiatry 2018, 9, 165. [Google Scholar] [CrossRef]
- Sadeh, N.; Wolf, E.; Logue, M.W.; Hayes, J.P.; Stone, A.; Griffin, L.M.; Schichman, S.A.; Miller, M. Epigenetic Variation at Ska2 Predicts Suicide Phenotypes and Internalizing Psychopathology. Depress. Anxiety 2016, 33, 308–315. [Google Scholar] [CrossRef]
- McGowan, O.P.; Sasaki, A.; D’Alessio, A.C.; Dymov, S.; Labonté, B.; Szyf, M.; Turecki, G.; Meaney, M.J. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat. Neurosci. 2009, 12, 342–348. [Google Scholar] [CrossRef]
- Sinclair, D.; Fullerton, J.; Webster, M.J.; Weickert, C.S. Glucocorticoid Receptor 1B and 1C mRNA Transcript Alterations in Schizophrenia and Bipolar Disorder, and Their Possible Regulation by GR Gene Variants. PLoS ONE 2012, 7, e31720. [Google Scholar] [CrossRef]
- Zhao, J.; Qi, X.-R.; Gao, S.-F.; Lu, J.; van Wamelen, D.; Kamphuis, W.; Bao, A.-M.; Swaab, D. Different stress-related gene expression in depression and suicide. J. Psychiatr. Res. 2015, 68, 176–185. [Google Scholar] [CrossRef]
- Roy, B.; Shelton, R.C.; Dwivedi, Y. DNA methylation and expression of stress related genes in PBMC of MDD patients with and without serious suicidal ideation. J. Psychiatr. Res. 2017, 89, 115–124. [Google Scholar] [CrossRef]
- Melhem, N.M.; Munroe, S.; Marsland, A.; Gray, K.; Brent, D.; Porta, G.; Douaihy, A.; Laudenslager, M.; DePietro, F.; Diler, R.S.; et al. Blunted HPA axis activity prior to suicide attempt and increased inflammation in attempters. Psychoneuroendocrinology 2017, 77, 284–294. [Google Scholar] [CrossRef]
- Chang, H.B.; Munroe, S.; Gray, K.; Porta, G.; Douaihy, A.; Marsland, A.; Brent, D.; Melhem, N.M. The role of substance use, smoking, and inflammation in risk for suicidal behavior. J. Affect. Disord. 2018, 243, 33–41. [Google Scholar] [CrossRef]
- Hiroi, N.; Wong, M.-L.; Licinio, J.; Park, C.; Young, M.; Gold, P.W.; Chrousos, G.P.; Bornstein, S.R. Expression of corticotropin releasing hormone receptors type I and type II mRNA in suicide victims and controls. Mol. Psychiatry 2001, 6, 540–546. [Google Scholar] [CrossRef]
- Merali, Z.; Du, L.; Hrdina, P.; Palkovits, M.; Faludi, G.; Poulter, M.O.; Anisman, H. Dysregulation in the Suicide Brain: mRNA Expression of Corticotropin-Releasing Hormone Receptors and GABAA Receptor Subunits in Frontal Cortical Brain Region. J. Neurosci. 2004, 24, 1478–1485. [Google Scholar] [CrossRef]
- Thomas, N.; Armstrong, C.W.; Hudaib, A.R.; Kulkarni, J.; Gurvich, C. A network meta-analysis of stress mediators in suicide behaviour. Front. Neuroendocrinol. 2021, 63, 100946. [Google Scholar] [CrossRef]
- Zheng, D.; Bi, X.; Zhang, T.; Han, C.; Ma, T.; Wang, L.; Sun, M.; Cui, K.; Yang, L.; Liu, L. Epigenetic Alterations of the Promoter Region of the POMC Gene in Adolescent Depressive Disorder Patients with Nonsuicidal Self-Injury Behaviors. Psychol. Res. Behav. Manag. 2020, 13, 997–1008. [Google Scholar] [CrossRef]
- Groh, A.; Rhein, M.; Buchholz, V.; Burkert, A.; Huber, C.G.; Lang, U.E.; Borgwardt, S.; Heberlein, A.; Muschler, M.A.; Hillemacher, T.; et al. Epigenetic Effects of Intravenous Diacetylmorphine on the Methylation of POMC and NR3C1. Neuropsychobiology 2017, 75, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Pandey, G.N.; Rizavi, H.S.; Ren, X.; Dwivedi, Y.; Palkovits, M. Region-specific alterations in glucocorticoid receptor expression in the postmortem brain of teenage suicide victims. Psychoneuroendocrinology 2013, 38, 2628–2639. [Google Scholar] [CrossRef] [PubMed]
- Braquehais, M.D.; Picouto, M.D.; Casas, M.; Sher, L. Hypothalamic-pituitary-adrenal axis dysfunction as a neurobiological correlate of emotion dysregulation in adolescent suicide. World J. Pediatr. 2012, 8, 197–206. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).