
Discogenic Low Back Pain: Anatomy, Pathophysiology and Treatments of Intervertebral Disc Degeneration

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
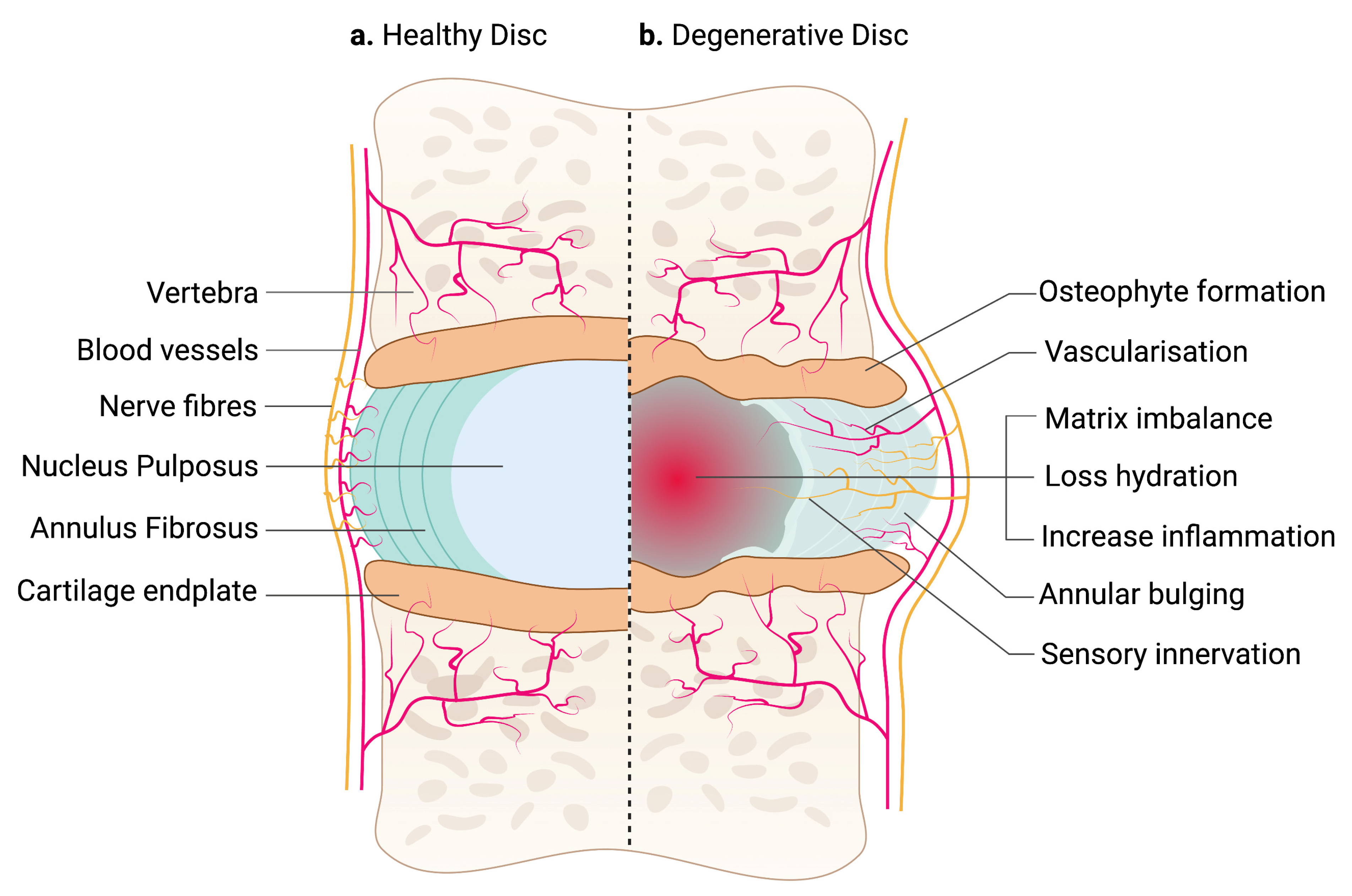
2. Anatomy of Intervertebral Disc
2.1. Annulus Fibrosus
2.2. Nucleus Pulposus
2.3. Cartilage End Plate
2.4. Blood and Nerve Supplies
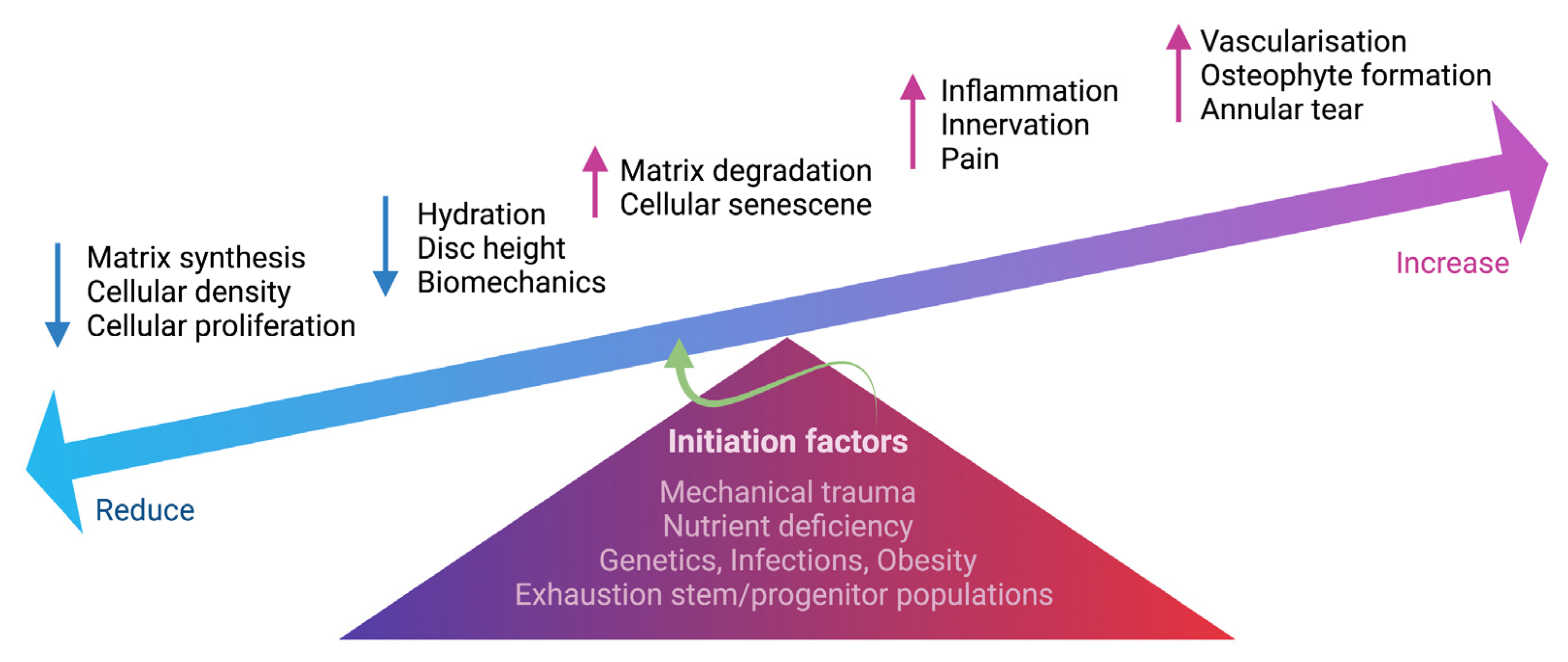
3. Pathophysiology of Intervertebral Disc Degeneration
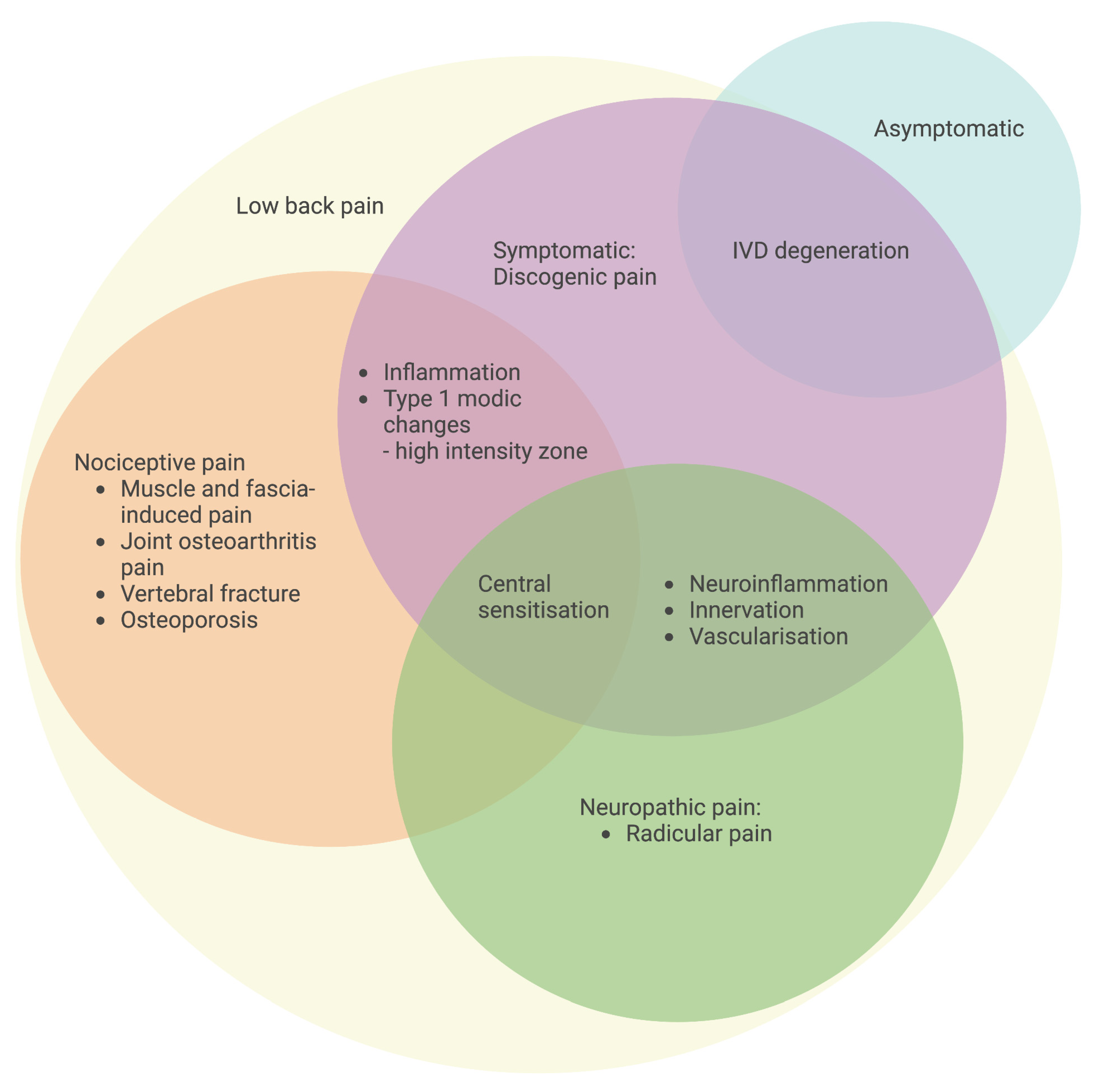
4. Discogenic Low Back Pain
5. Mechanisms of Intervertebral Disc Degeneration Underlying Discogenic Low Back Pain
5.1. Neuroinflammation-Induced Innervation of Nociceptive Fibers in the Discs
5.2. Neuroinflammation-Induced Nociception
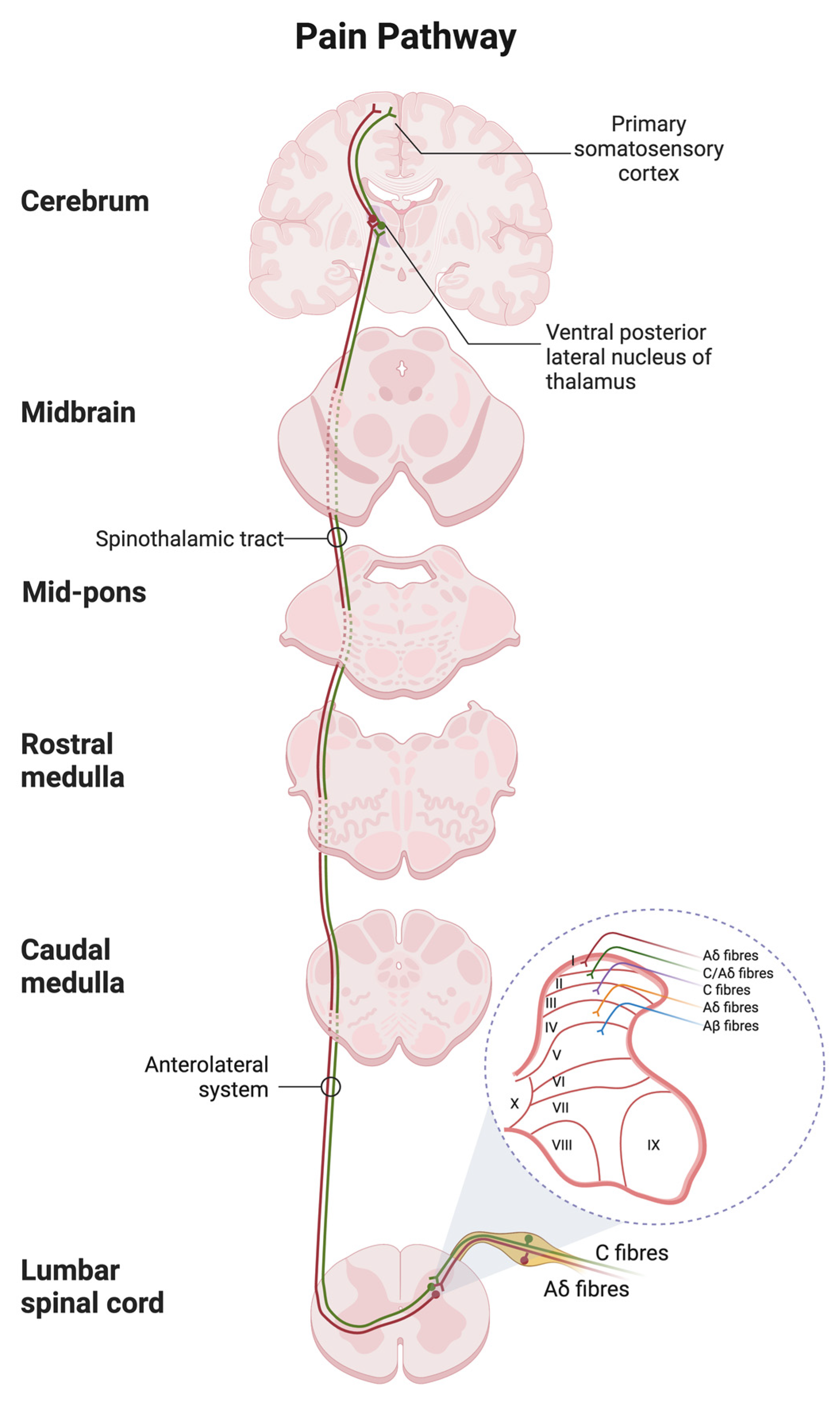
6. Pain Pathway
6.1. Peripheral Sensitisation
6.2. Central Sensitisation
6.3. Ascending and Descending Tracts
7. Current Treatments
8. Clinical Perspective and Future Treatments
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collaborators, G.B.D.; Murray, C.; Aravkin, A.; Zheng, P.; Vos, T.; Lim, S.; Furtado, J.; Ghafourifard, M.; Abbas, K.; Abbasi, M.; et al. Global Burden of 369 Diseases and Injuries in 204 Countries and Territories, 1990–2019: A Systematic Analysis for the Global Burden of Disease Study 2019. Lancet 2020, 396, 1135–1159. [Google Scholar] [CrossRef]
- Wu, A.; March, L.; Zheng, X.; Huang, J.; Wang, X.; Zhao, J.; Blyth, F.M.; Smith, E.; Buchbinder, R.; Hoy, D. Global Low Back Pain Prevalence and Years Lived with Disability from 1990 to 2017: Estimates from the Global Burden of Disease Study 2017. Ann. Transl. Med. 2020, 8, 299. [Google Scholar] [CrossRef] [PubMed]
- Abidin, N.Z.; Rohani, J.M.; Nordin, A.N.; Zein, R.M.; Anak Ayak, A.S. Financial Impact and Causes of Chronic Musculoskeletal Disease Cases in Malaysia Based on Social Security Organization of Malaysia Claims Record. Int. J. Eng. Technol. 2018, 7, 23–27. [Google Scholar] [CrossRef][Green Version]
- Hall, J.A.; Jowett, S.; Lewis, M.; Oppong, R.; Konstantinou, K. The STarT Back Stratified Care Model for Nonspecific Low Back Pain: A Model-based Evaluation of Long-term Cost-effectiveness. Pain 2021, 162, 702–710. [Google Scholar] [CrossRef]
- Belitskaya-Levy, I.; David Clark, J.; Shih, M.C.; Bair, M.J. Treatment Preferences for Chronic Low Back Pain: Views of Veterans and Their Providers. J. Pain Res. 2021, 14, 161–171. [Google Scholar] [CrossRef]
- Montgomery, W.; Sato, M.; Nagasaka, Y.; Vietri, J. The Economic and Humanistic Costs of Chronic Lower Back Pain in Japan. Clin. Outcomes Res. 2017, 9, 361–371. [Google Scholar] [CrossRef]
- Malik, M.H.A.; Singh, D.; Mansor, M.; Mohamed Kamil, O.I.; Yin Choy, C.; Cardosa, M.S.; Hasnan, N.; Vijayan, R. The Malaysian Low Back Pain Guideline; Malaysian Association for the Study of Pain and Spine Society: Kuala Lumpur, Malaysia, 2014; Volume 1. [Google Scholar]
- Peng, B.-G. Pathophysiology, Diagnosis, and Treatment of Discogenic Low Back Pain. World J. Orthop. 2013, 4, 42–52. [Google Scholar] [CrossRef]
- Mohd Isa, I.L.; Mokhtar, S.A.; Abbah, S.A.; Fauzi, M.B.; Devitt, A.; Pandit, A. Intervertebral Disc Degeneration: Biomaterials and Tissue Engineering Strategies Towards Precision Medicine. Adv. Healthc. Mater. 2022, 11, e2102530. [Google Scholar] [CrossRef]
- Prithvi, R.P. Intervertebral Disc: Anatomy-Physiology-Pathophysiology-Treatment. Pain Pract. 2008, 8, 18–44. [Google Scholar] [CrossRef]
- Sakai, D.; Nakamura, Y.; Nakai, T.; Mishima, T.; Kato, S.; Grad, S.; Alini, M.; Risbud, M.V.; Chan, D.; Cheah, K.S.E.; et al. Exhaustion of Nucleus Pulposus Progenitor Cells with Ageing and Degeneration of the Intervertebral Disc. Nat. Commun. 2012, 3, 1264. [Google Scholar] [CrossRef]
- Bermudez-Lekerika, P.; Crump, K.B.; Tseranidou, S.; Nüesch, A.; Kanelis, E.; Alminnawi, A.; Baumgartner, L.; Muñoz-Moya, E.; Compte, R.; Gualdi, F.; et al. Immuno-Modulatory Effects of Intervertebral Disc Cells. Front. Cell Dev. Biol. 2022, 10, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Lyu, F.J.; Cui, H.; Pan, H.; MC Cheung, K.; Cao, X.; Iatridis, J.C.; Zheng, Z. Painful Intervertebral Disc Degeneration and Inflammation: From Laboratory Evidence to Clinical Interventions. Bone Res. 2021, 9, 7. [Google Scholar] [CrossRef] [PubMed]
- Lyu, F.J.; Cheung, K.M.; Zheng, Z.; Wang, H.; Sakai, D.; Leung, V.Y. IVD Progenitor Cells: A New Horizon for Understanding Disc Homeostasis and Repair. Nat. Rev. Rheumatol. 2019, 15, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Mayer, J.E.; Iatridis, J.C.; Chan, D.; Qureshi, S.A.; Gottesman, O.; Hecht, A.C. Genetic Polymorphisms Associated with Intervertebral Disc Degeneration. Spine J. 2013, 13, 299–317. [Google Scholar] [CrossRef]
- Shiri, R.; Karppinen, J.; Leino-Arjas, P.; Solovieva, S.; Viikari-Juntura, E. The Association between Smoking and Low Back Pain: A Meta-analysis. Am. J. Med. 2010, 123, 87.e7–87.e35. [Google Scholar] [CrossRef]
- Shiri, R.; Karppinen, J.; Leino-Arjas, P.; Solovieva, S.; Viikari-Juntura, E. The Association Between Obesity and Low Back Pain: A meta-analysis. Am. J. Epidemiol. 2010, 171, 135–154. [Google Scholar] [CrossRef]
- Moore, K.L.; DalleyI, A.F.; Agur, A.M.R. Clinially Oriented Anatomy; Lippincott Williams & Wilkins: Baltimore, MA, USA; Philadelphia, PA, USA, 2014; ISBN 9781451119459. [Google Scholar]
- Newell, N.; Little, J.P.; Christou, A.; Adams, M.A.; Adam, C.J.; Masouros, S.D. Biomechanics of the Human Intervertebral Disc: A Review of Testing Techniques and Results. J. Mech. Behav. Biomed. Mater. 2017, 69, 420–434. [Google Scholar] [CrossRef]
- Shapiro, I.M.; Risbud, M.V. Introduction to the Structure, Function, and Comparative Anatomy of the Vertebrae and the Intervertebral Disc. In The Intervertebral Disc; Springer: Wien, Austria; Heidelberg, Germany; New York, NY, USA; Dordrecht, The Netherlands; London, UK, 2014; pp. 3–429. ISBN 9783030209254. [Google Scholar]
- Molladavoodi, S.; McMorran, J.; Gregory, D. Mechanobiology of Annulus Fibrosus and Nucleus Pulposus Cells in Intervertebral Discs. Cell Tissue Res. 2020, 379, 429–444. [Google Scholar] [CrossRef]
- Stein, D.; Assaf, Y.; Dar, G.; Cohen, H.; Slon, V.; Kedar, E.; Medlej, B.; Abbas, J.; Hay, O.; Barazany, D.; et al. 3D Virtual Reconstruction and Quantitative Assessment of the Human Intervertebral Disc’s Annulus Fibrosus: A DTI Tractography Study. Sci. Rep. 2021, 11, 6815. [Google Scholar] [CrossRef]
- Torre, O.M.; Mroz, V.; Bartelstein, M.K.; Huang, A.H.; Iatridis, J.C. Annulus Fibrosus Cell Phenotypes in Homeostasis and Injury: Implications for Regenerative Strategies. Ann. N. Y. Acad. Sci. 2019, 1442, 61–78. [Google Scholar] [CrossRef]
- Van Den Akker, G.G.H.; Surtel, D.A.M.; Cremers, A.; Richardson, S.M.; Hoyland, J.A.; Van Rhijn, L.W.; Voncken, J.W.; Welting, T.J.M. Novel Immortal Cell Lines Support Cellular Heterogeneity in the Human Annulus Fibrosus. PLoS ONE 2016, 11, e0144497. [Google Scholar] [CrossRef] [PubMed]
- Lv, F.; Leung, V.Y.L.; Huang, S.; Huang, Y.; Sun, Y.; Cheung, K.M.C. In Search of Nucleus Pulposus-specific Molecular Markers. Rheumatology 2014, 53, 600–610. [Google Scholar] [CrossRef] [PubMed]
- Manira, M.; Shamsul, B.S.; Aminuddin, B.S.; Ruszymah, B.H.I. Cultivation of Intervertebral Disc Cells in Medium Fortified with Growth Factors Improved In Vitro Chondrogenesis. Biomed. Res. 2011, 22, 435–441. [Google Scholar]
- Tang, X.; Jing, L.; Chen, J. Changes in the Molecular Phenotype of Nucleus Pulposus Cells with Intervertebral Disc Aging. PLoS ONE 2012, 7, e52020. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.M.; Ludwinski, F.E.; Gnanalingham, K.K.; Atkinson, R.A.; Freemont, A.J.; Hoyland, J.A. Notochordal and Nucleus Pulposus Marker Expression is Maintained by Sub-populations of Adult Human Nucleus Pulposus Cells through Aging and Degeneration. Sci. Rep. 2017, 7, 1501. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, S.; Pinelli, R.; Pricop, P.; Albert, T.J.; Dahia, C.L. Chondrocyte-like Nested Cells in the Aged Intervertebral Disc are Late-stage Nucleus Pulposus Cells. Aging Cell 2019, 18, e13006. [Google Scholar] [CrossRef]
- Moon, S.M.; Yoder, J.H.; Wright, A.C.; Smith, L.J.; Vresilovic, E.J.; Elliott, D.M. Evaluation of Intervertebral Disc Cartilaginous Endplate Structure using Magnetic Resonance Imaging. Eur. Spine J. 2013, 22, 1820–1828. [Google Scholar] [CrossRef]
- Rodrigues, S.A.; Wade, K.R.; Thambyah, A.; Broom, N.D. Micromechanics of Annulus-end Plate Integration in the Intervertebral Disc. Spine J. 2012, 12, 143–150. [Google Scholar] [CrossRef]
- Lakstins, K.; Arnold, L.; Gunsch, G.; Flanigan, D.; Khan, S.; Gadde, N.; Jones, B.; Agarwal, G.; Purmessur, D. Characterization of the Human Intervertebral Disc Cartilage Endplate at the Molecular, Cell, and Tissue levels. J. Orthop. Res. 2021, 39, 1898–1907. [Google Scholar] [CrossRef]
- Standring, S. Gray’s Anatomy: The Anatomical Basis of Clinical Practice; Churchill Livingstone Elsevier: London, UK, 2020; ISBN 9781118840931. [Google Scholar]
- Shirazi-Adl, A.; Taheri, M.; Urban, J.P.G. Analysis of Cell Viability in Intervertebral Disc: Effect of Endplate Permeability on Cell Population. J. Biomech. 2010, 43, 1330–1336. [Google Scholar] [CrossRef]
- Groh, A.M.R.; Fournier, D.E.; Battié, M.C.; Séguin, C.A. Innervation of the Human Intervertebral Disc: A Scoping Review. Pain Med. 2021, 22, 1281–1304. [Google Scholar] [CrossRef] [PubMed]
- Aoyama, R.; Yamane, J.; Ninomiya, K.; Takahashi, Y.; Kitamura, K.; Nori, S.; Suzuki, S.; Shiraishi, T. Disc Height Narrowing Could Not Stabilize the Mobility at the Level of Cervical Spondylolisthesis: A Retrospective Study of 83 Patients with Cervical Single-Level Spondylolisthesis. Asian Spine J. 2022, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Park, D.K.; Shah, J.; Phillips, F.M. The Biomechanics and Biology of the Spinal Degenerative Cascade. Semin. Spine Surg. 2005, 17, 128–136. [Google Scholar] [CrossRef]
- Tang, S.; Richards, J.; Khan, S.; Hoyland, J.; Gallego-Perez, D.; Higuita-Castro, N.; Walter, B.; Purmessur, D. Nonviral Transfection With Brachyury Reprograms Human Intervertebral Disc Cells to a Pro-Anabolic Anti-Catabolic/Inflammatory Phenotype: A Proof of Concept Study. J. Orthop. Res. 2019, 37, 2389–2400. [Google Scholar] [CrossRef]
- Sheyn, D.; Ben-David, S.; Tawackoli, W.; Zhou, Z.; Salehi, K.; Bez, M.; de Mel, S.; Chan, V.; Roth, J.; Avalos, P.; et al. Human iPSCs Can Be Differentiated Into Notochordal Cells That Reduce Intervertebral Disc Degeneration in A Porcine Model. Theranostics 2019, 9, 7506–7524. [Google Scholar] [CrossRef]
- Sha’Ban, M.; Yoon, S.J.; Ko, Y.K.; Ha, H.J.; Kim, S.H.; So, J.W.; Idrus, R.B.H.; Khang, G. Fibrin Promotes Proliferation and Matrix Production of Intervertebral Disc Cells Cultured in Three-dimensional Poly(lactic-co-glycolic acid) Scaffold. J. Biomater. Sci. Polym. Ed. 2008, 19, 1219–1237. [Google Scholar] [CrossRef]
- Khan, A.N.; Jacobsen, H.E.; Khan, J.; Filippi, C.G.; Levine, M.; Lehman, R.A.; Riew, K.D.; Lenke, L.G.; Chahine, N.O. Inflammatory Biomarkers of Low Back Pain and Disc Degeneration: A Review. Ann. N. Y. Acad. Sci. 2017, 1410, 68–84. [Google Scholar] [CrossRef]
- Binch, A.L.A.; Fitzgerald, J.C.; Growney, E.A.; Barry, F. Cell-based Strategies for IVD Repair: Clinical Progress and Translational Obstacles. Nat. Rev. Rheumatol. 2021, 17, 158–175. [Google Scholar] [CrossRef]
- Kazezian, Z.; Gawri, R.; Haglund, L.; Ouellet, J.; Mwale, F.; Tarrant, F.; O’Gaora, P.; Pandit, A.; Alini, M.; Grad, S. Gene Expression Profiling Identifies Interferon Signalling Molecules and IGFBP3 in Human Degenerative Annulus Fibrosus. Sci. Rep. 2015, 5, 15662. [Google Scholar] [CrossRef]
- Hiyama, A.; Sakai, D.; Tanaka, M.; Arai, F.; Nakajima, D.; Abe, K.; Mochida, J. The Relationship Between the Wnt/β-catenin and TGF-β/BMP Signals in the Intervertebral Disc Cell. J. Cell. Physiol. 2011, 226, 1139–1148. [Google Scholar] [CrossRef]
- Riester, S.M.; Lin, Y.; Wang, W.; Cong, L.; Mohamed Ali, A.M.; Peck, S.H.; Smith, L.J.; Currier, B.L.; Clark, M.; Huddleston, P.; et al. RNA Sequencing Identifies Gene Regulatory Networks Controlling Extracellular Matrix Synthesis in Intervertebral Disk Tissues. J. Orthop. Res. 2018, 36, 1356–1369. [Google Scholar] [CrossRef] [PubMed]
- Navone, S.E.; Marfia, G.; Giannoni, A.; Beretta, M.; Guarnaccia, L.; Gualtierotti, R.; Nicoli, D.; Rampini, P.; Campanella, R. Inflammatory Mediators and Signalling Pathways Controlling Intervertebral Disc Degeneration. Histol. Histopathol. 2017, 32, 523–542. [Google Scholar] [CrossRef] [PubMed]
- Risbud, M.V.; Shapiro, I.M. Role of Cytokines in Intervertebral Disc Degeneration: Pain and Disc Content. Nat. Rev. Rheumatol. 2014, 10, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Hiyama, A.; Suyama, K.; Sakai, D.; Tanaka, M.; Watanabe, M. Correlational Analysis of Chemokine and Inflammatory Cytokine Expression in the Intervertebral Disc and Blood in Patients with Lumbar Disc Disease. J. Orthop. Res. 2021, 40, 1213–1222. [Google Scholar] [CrossRef] [PubMed]
- Mohd Isa, I.L.; Günay, B.; Joyce, K.; Pandit, A. Tissue Engineering: Biomaterials for Disc Repair. Curr. Mol. Biol. Rep. 2018, 4, 161–172. [Google Scholar] [CrossRef]
- Wang, J.; Markova, D.; Anderson, D.G.; Zheng, Z.; Shapiro, I.M.; Risbud, M.V. TNF-α and IL-1β Promote A Disintegrin-like and Metalloprotease with Thrombospondin Type I Motif-5-mediated Aggrecan Degradation Through Syndecan-4 in Intervertebral disc. J. Biol. Chem. 2011, 286, 39738–39749. [Google Scholar] [CrossRef]
- Wang, Y.; Che, M.; Xin, J.; Zheng, Z.; Li, J.; Zhang, S. The Role of IL-1β and TNF-α in Intervertebral Disc Degeneration. Biomed. Pharmacother. 2020, 131, 110660. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.; Yang, H.; Li, J.; Cai, Q.; Shapiro, I.M.; Risbud, M. V Tumor Necrosis Factor-α- and Interleukin-1β-Dependent Matrix Metalloproteinase-3 Expression in Nucleus Pulposus Cells Requires Cooperative Signaling via Syndecan 4 and Mitogen-activated Protein Kinase-NF-κB Axis: Implications in Inflammatory Disc Disease. Am. J. Pathol. 2014, 184, 2560–2572. [Google Scholar] [CrossRef]
- Joyce, K.; Mohd Isa, I.L.; Krouwels, A.; Creemers, L.B.; Devitt, A.; Pandit, A. The Role of Altered Glycosylation in Human Nucleus Pulposus Cells in Inflammation and Degeneration. Eur. Cells Mater. 2021, 40, 401–420. [Google Scholar] [CrossRef]
- Shankar, H.; Scarlett, J.A.; Abram, S.E. Anatomy and Pathophysiology of Intervertebral Disc Disease. Tech. Reg. Anesth. Pain Manag. 2009, 13, 67–75. [Google Scholar] [CrossRef]
- Stefanakis, M.; Al-Abbasi, M.; Harding, I.; Pollintine, P.; Dolan, P.; Tarlton, J.; Adams, M.A. Annulus Fissures are Mechanically and Chemically Conducive to the Ingrowth of Nerves and Blood Vessels. Spine 2012, 37, 1883–1891. [Google Scholar] [CrossRef] [PubMed]
- Peng, B.; Wu, W.; Hou, S.; Li, P.; Zhang, C.; Yang, Y. The Pathogenesis of Discogenic Low Back Pain. J. Bone Jt. Surg.-Ser. B 2005, 87, 62–67. [Google Scholar] [CrossRef]
- Hsieh, A.; Yoon, S.T. Update on the Pathophysiology of Degenerative Disc Disease and New Developments in Treatment Strategies. Open Access J. Sport. Med. 2010, 1, 191. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Song, J.Y.; Baek, M.; Jung, H.-Y.; Kang, H.; Han, I.B.; Kwon, Y.D.; Shin, D.E. Interleukin-1β Induces Angiogenesis and Innervation in Human Intervertebral Disc Degeneration. J. Orthop. Res. 2011, 29, 265–269. [Google Scholar] [CrossRef]
- Purmessur, D.; Freemont, A.J.; Hoyland, J.A. Expression and Regulation of Neurotrophins in the Nondegenerate and Degenerate Human Intervertebral Disc. Arthritis Res. Ther. 2008, 10, R99. [Google Scholar] [CrossRef]
- Mohd Isa, I.L.; Srivastava, A.; Tiernan, D.; Owens, P.; Rooney, P.; Dockery, P.; Pandit, A. Hyaluronic Acid Based Hydrogels Attenuate Inflammatory Receptors and Neurotrophins in Interleukin-1β Induced Inflammation Model of Nucleus Pulposus Cells. Biomacromolecules 2015, 16, 1714–1725. [Google Scholar] [CrossRef]
- Freemont, A.J.; Peacock, T.E.; Goupille, P.; Hoyland, J.A.; Brien, J.O.; Jayson, M.I. V Early Report Nerve Ingrowth into Diseased Intervertebral Disc in Chronic Back Pain. Lancet 1997, 350, 178–181. [Google Scholar] [CrossRef]
- Freemont, A.J.; Watkins, A.; Le Maitre, C.; Baird, P.; Jeziorska, M.; Knight, M.T.N.; Ross, E.R.S.; O’Brien, J.P.; Hoyland, J.A. Nerve Growth Factor Expression and Innervation of the Painful Intervertebral Disc. J. Pathol. 2002, 197, 286–292. [Google Scholar] [CrossRef]
- Coppes, M.H.; Marani, E.; Thomeer, R.T.W.M.; Groen, G.J. Innervation of Painful Lumbar Discs. Spine 1997, 22, 2342–2350. [Google Scholar] [CrossRef]
- Thompson, J.P.; Pearce, R.H.; Schechter, M.T.; Adams, M.E.; Tsang, I.K.; Bishop, P.B. Preliminary Evaluation of a Scheme for Grading the Gross Morphology of the Human Intervertebral Disc. Spine 1990, 15, 411–415. [Google Scholar] [CrossRef]
- Rutges, J.P.H.J.; Duit, R.A.; Kummer, J.A.; Bekkers, J.E.J.; Oner, F.C.; Castelein, R.M.; Dhert, W.J.A.; Creemers, L.B. A Validated New Histological Classification for Intervertebral Disc Degeneration. Osteoarthr. Cartil. 2013, 21, 2039–2047. [Google Scholar] [CrossRef] [PubMed]
- Le Maitre, C.L.; Dahia, C.L.; Giers, M.; Illien-Junger, S.; Cicione, C.; Samartzis, D.; Vadala, G.; Fields, A.; Lotz, J. Development of a Standardized Histopathology Scoring System for Human Intervertebral Disc Degeneration: An Orthopaedic Research Society Spine Section Initiative. JOR Spine 2021, 4, e1167. [Google Scholar] [CrossRef] [PubMed]
- Pfirrmann, C.W.A.; Metzdorf, A.; Zanetti, M.; Hodler, J.; Boos, N. Magnetic Resonance Classification of Lumbar Intervertebral Disc Degeneration. Spine 2001, 26, 1873–1878. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K.; Yamazaki, M.; Kang, J.D.; Risbud, M.V.; Cho, S.K.; Qureshi, S.A.; Hecht, A.C.; Iatridis, J.C. Discogenic Back Pain: Literature Review of Definition, Diagnosis, and Treatment. JBMR Plus 2019, 3, e10180. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Aoki, Y.; Ohtori, S. Resolving Discogenic Pain. Eur. Spine J. 2008, 17 (Suppl. S4), S428. [Google Scholar] [CrossRef]
- Chou, D.; Samartzis, D.; Bellabarba, C.; Patel, A.; Luk, K.D.K.; Kisser, J.M.S.; Skelly, A.C. Degenerative Magnetic Resonance Imaging Changes in Patients with Chronic Low Back Pain. Spine 2011, 36, S43–S53. [Google Scholar] [CrossRef]
- Von Forell, G.A.; Stephens, T.K.; Samartzis, D.; Bowden, A.E. Low Back Pain: A Biomechanical Rationale Based on “Patterns” of Disc Degeneration. Spine 2015, 40, 1165–1172. [Google Scholar] [CrossRef]
- Kim, H.S.; Wu, P.H.; Jang, I.T. Lumbar Degenerative Disease Part 1: Anatomy and Pathophysiology of Intervertebral Discogenic Pain and Radiofrequency Ablation of Basivertebral and Sinuvertebral Nerve Treatment for Chronic Discogenic Back Pain: A Prospective Case Series and Review. Int. J. Mol. Sci. 2020, 21, 1483. [Google Scholar] [CrossRef]
- Nakamura, S.-I.; Takahashi, K.; Takahashi, Y.; Yamagata, M.; Moriya, H. The Afferent Pathways of Discogenic Low-back Pain. J. Bone Jt. Surg. 1996, 78, 606. [Google Scholar] [CrossRef]
- Inoue, M.; Mohd Isa, I.L.; Orita, S.; Suzuki, M.; Inage, K.; Shiga, Y.; Norimoto, M.; Umimura, T.; Sakai, T.; Eguchi, Y.; et al. An Injectable Hyaluronic Acid Hydrogel Promotes Intervertebral Disc Repair in a Rabbit Model. Spine 2021, 46, E810–E816. [Google Scholar] [CrossRef]
- Basbaum, A.I.; Bautista, D.M.; Scherrer, G.; Julius, D. Cellular and Molecular Mechanisms of Pain. Cell 2010, 139, 267–284. [Google Scholar] [CrossRef] [PubMed]
- Snider, W.D.; McMahon, S.B. Tackling Pain at the Source: New Ideas About Nociceptors. Neuron 1998, 20, 629–632. [Google Scholar] [CrossRef] [PubMed]
- Aoki, Y.; Ohtori, S.; Takahashi, K.; Ino, H.; Takahashi, Y.; Chiba, T.; Moriya, H. Innervation of the Lumbar Intervertebral Disc by Nerve Growth Factor-dependent Neurons Related to Inflammatory Pain. Spine 2004, 29, 1077–1081. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.F.; Hukkanen, M.V.; McCarthy, I.D.; Redfern, D.R.; Batten, J.J.; Crock, H.V.; Hughes, S.P.; Polak, J.M. Sensory and Sympathetic Innervation of the Vertebral Endplate in Patients with Degenerative Disc Disease. J. Bone Jt. Surg. 1997, 79, 147–153. [Google Scholar] [CrossRef]
- García-Cosamalón, J.; del Valle, M.E.; Calavia, M.G.; García-Suárez, O.; López-Muñiz, A.; Otero, J.; Vega, J.A. Intervertebral Disc, Sensory Nerves and Neurotrophins: Who is Who in Discogenic Pain? J. Anat. 2010, 217, 1–15. [Google Scholar] [CrossRef]
- Bach, F.C.; Poramba-Liyanage, D.W.; Riemers, F.M.; Guicheux, J.; Camus, A.; Iatridis, J.C.; Chan, D.; Ito, K.; Le Maitre, C.L.; Tryfonidou, M.A. Notochordal Cell-Based Treatment Strategies and Their Potential in Intervertebral Disc Regeneration. Front. Cell Dev. Biol. 2022, 9, 3645. [Google Scholar] [CrossRef]
- Purmessur, D.; Guterl, C.C.; Cho, S.K.; Cornejo, M.C.; Lam, Y.W.; Ballif, B.A.; Laudier, D.M.; Iatridis, J.C. Dynamic Pressurization Induces Transition of Notochordal Cells to a Mature Phenotype While Retaining Production of Important Patterning Ligands From Development. Arthritis Res. Ther. 2013, 15, R122. [Google Scholar] [CrossRef]
- Richardson, S.M.; Purmessur, D.; Baird, P.; Probyn, B.; Freemont, A.J.; Hoyland, J. Degenerate Human Nucleus Pulposus Cells Promote Neurite Outgrowth in Neural Cells. PLoS ONE 2012, 7, e47735. [Google Scholar] [CrossRef]
- Ji, R.R.; Samad, T.A.; Jin, S.X.; Schmoll, R.; Woolf, C.J. p38 MAPK Activation by NGF in Primary Sensory Neurons after Inflammation Increases TRPV1 Levels and Maintains Heat Hyperalgesia. Neuron 2002, 36, 57–68. [Google Scholar] [CrossRef]
- Zhang, X.; Huang, J.; McNaughton, P.A. NGF Rapidly Increases Membrane Expression of TRPV1 heat-gated Ion Channels. EMBO J. 2005, 24, 4211–4223. [Google Scholar] [CrossRef]
- Mohd Isa, I.L.; Abbah, S.A.; Kilcoyne, M.; Sakai, D.; Dockery, P.; Finn, D.P.; Pandit, A. Implantation of Hyaluronic Acid Hydrogel Prevents the Pain Phenotype in a Rat Model of Intervertebral Disc Injury. Sci. Adv. 2018, 4, eaaq0597. [Google Scholar] [CrossRef]
- Byron, K.L.; Overland, C.; Vanderah, T.W.; Spencer, R.H. Peripheral Mechanisms of Pain and Analgesia. Brain Res. Rev. 2010, 60, 90–113. [Google Scholar] [CrossRef]
- Mannion, R.J.; Costigan, M.; Decosterd, I.; Amaya, F.; Ma, Q.P.; Holstege, J.C.; Ji, R.R.; Acheson, A.; Lindsay, R.M.; Wilkinson, G.A.; et al. Neurotrophins: Peripherally and Centrally Acting Modulators of Tactile Stimulus-induced Inflammatory Pain Hypersensitivity. Proc. Natl. Acad. Sci. USA 1999, 96, 9385–9390. [Google Scholar] [CrossRef] [PubMed]
- Kepler, C.K.; Markova, D.Z.; Hilibrand, A.S.; Vaccaro, A.R.; Risbud, M.V.; Albert, T.J.; Anderson, D.G. Substance P Stimulates Production of Inflammatory Cytokines in Human Disc Cells. Spine 2013, 38, E1291–E1299. [Google Scholar] [CrossRef] [PubMed]
- Neumann, S.; Doubell, T.P.; Leslie, T.; Woolf, C.J. Inflammatory Pain Hypersensitivity Mediated by Phenotypic Switch in Myelinated Primary Sensory Neurons. Nature 1996, 384, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Sakuma, Y.; Suzuki, M.; Orita, S.; Yamauchi, K.; Inoue, G.; Aoki, Y.; Ishikawa, T.; Miyagi, M.; Kamoda, H.; et al. Evaluation of Pain Behavior and Calcitonin Gene-Related Peptide Immunoreactive Sensory Nerve Fibers in the Spinal Dorsal Horn After Sciatic Nerve Compression and Application of Nucleus Pulposus in Rats. Spine 2014, 39, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, Y.; Cheng, C.-C.; Danielson, K.G.; Mochida, J.; Albert, T.J.; Shapiro, I.M.; Risbud, M. V Expression of Acid-sensing Ion Channel 3 (ASIC3) in Nucleus Pulposus Cells of the Intervertebral Disc is Regulated by p75NTR and ERK Signaling. J. Bone Miner. Res. 2007, 22, 1996–2006. [Google Scholar] [CrossRef]
- Park, S.Y.; Choi, J.Y.; Kim, R.U.; Lee, Y.S.; Cho, H.J.; Kim, D.S. Downregulation of Voltage-gated Potassium Channel Alpha Gene Expression by Axotomy and Neurotrophins in Rat Dorsal Root Ganglia. Mol. Cells 2003, 16, 256–259. [Google Scholar]
- Steeds, C.E. The Anatomy and Physiology of Pain. Basic Sci. 2016, 34, 55–59. [Google Scholar] [CrossRef]
- Todd, A.J. Neuronal Circuitry for Pain Processing in the Dorsal Horn. Nat. Rev. Neurosci. 2010, 11, 823–836. [Google Scholar] [CrossRef]
- Millan, M.I. The Induction of Pain: An integrative Review. Prog. Neurobiol. 1999, 57, 1–164. [Google Scholar] [CrossRef] [PubMed]
- Ohtori, S.; Takahashi, K.; Ino, H.; Chiba, T.; Yamagata, M.; Sameda, H.; Moriya, H. Up-regulation of Substance P and NMDA Receptor mRNA in Dorsal Horn and Preganglionic Sympathetic Neurons During Adjuvant-induced Noxious Stimulation in Rats. Ann. Anat. 2002, 184, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Merighi, A.; Salio, C.; Ghirri, A.; Lossi, L.; Ferrini, F.; Betelli, C.; Bardoni, R. BDNF as a Pain Modulator. Prog. Neurobiol. 2008, 85, 297–317. [Google Scholar] [CrossRef] [PubMed]
- Hunt, S.P.; Pini, A.; Evan, G. Induction of c-fos-like Protein in Spinal Cord Neurons Following Sensory Stimulation. Nature 1987, 328, 632–634. [Google Scholar] [CrossRef]
- Millan, M.J. Descending Control of Pain. Prog. Neurobiol. 2002, 66, 355–474. [Google Scholar] [CrossRef]
- North American Spine Society. Evidence-Based Clinical Guidelines for Multidisciplinary Spine Care: Diagnosis & Treatment of Low Back Pain; North American Spine Society: Burr Ridge, IL, USA, 2020; ISBN 978-1-929988-65-5. [Google Scholar]
- Forssell, M.Z. The Back School. Spine 1981, 6, 104–106. [Google Scholar] [CrossRef]
- Wu, P.H.; Kim, H.S.; Jang, I.T. Intervertebral Disc Diseases Part 2: A Review of the Current Diagnostic and Treatment Strategies for Intervertebral Disc Disease. Int. J. Mol. Sci. 2020, 21, 2135. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohd Isa, I.L.; Teoh, S.L.; Mohd Nor, N.H.; Mokhtar, S.A. Discogenic Low Back Pain: Anatomy, Pathophysiology and Treatments of Intervertebral Disc Degeneration. Int. J. Mol. Sci. 2023, 24, 208. https://doi.org/10.3390/ijms24010208
Mohd Isa IL, Teoh SL, Mohd Nor NH, Mokhtar SA. Discogenic Low Back Pain: Anatomy, Pathophysiology and Treatments of Intervertebral Disc Degeneration. International Journal of Molecular Sciences. 2023; 24(1):208. https://doi.org/10.3390/ijms24010208
Chicago/Turabian StyleMohd Isa, Isma Liza, Seong Lin Teoh, Nurul Huda Mohd Nor, and Sabarul Afian Mokhtar. 2023. "Discogenic Low Back Pain: Anatomy, Pathophysiology and Treatments of Intervertebral Disc Degeneration" International Journal of Molecular Sciences 24, no. 1: 208. https://doi.org/10.3390/ijms24010208
APA StyleMohd Isa, I. L., Teoh, S. L., Mohd Nor, N. H., & Mokhtar, S. A. (2023). Discogenic Low Back Pain: Anatomy, Pathophysiology and Treatments of Intervertebral Disc Degeneration. International Journal of Molecular Sciences, 24(1), 208. https://doi.org/10.3390/ijms24010208