
Effects of the Treatment with Flavonoids on Metabolic Syndrome Components in Humans: A Systematic Review Focusing on Mechanisms of Action

 ,
,  ,
, 
Abstract
:1. Introduction
2. Methods
2.1. Systematic Review Reporting and Protocol Registration
2.2. Search Strategy
2.3. Inclusion Criteria
2.4. Data Extraction
2.5. Analysis of Risk of Bias
3. Results
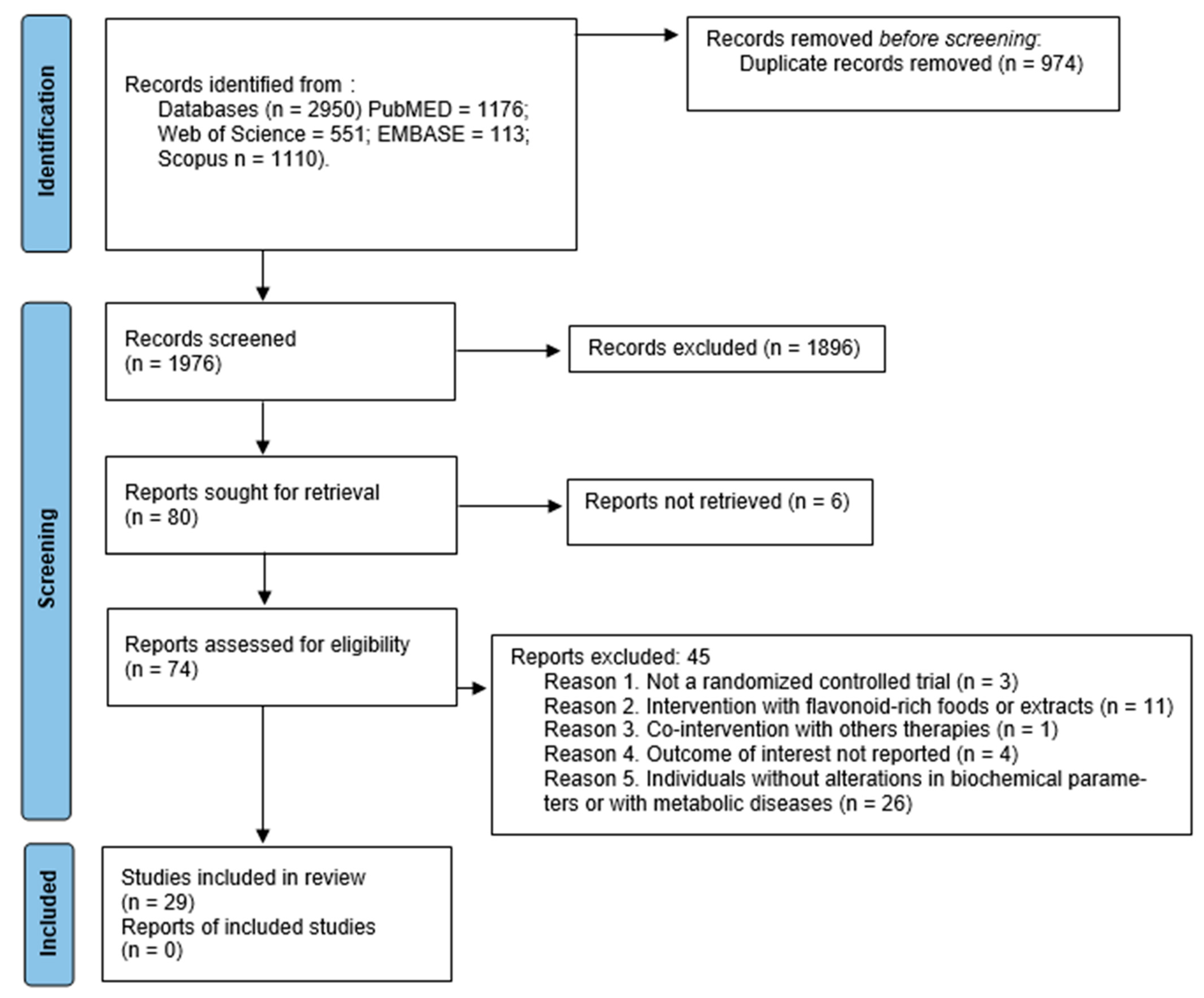
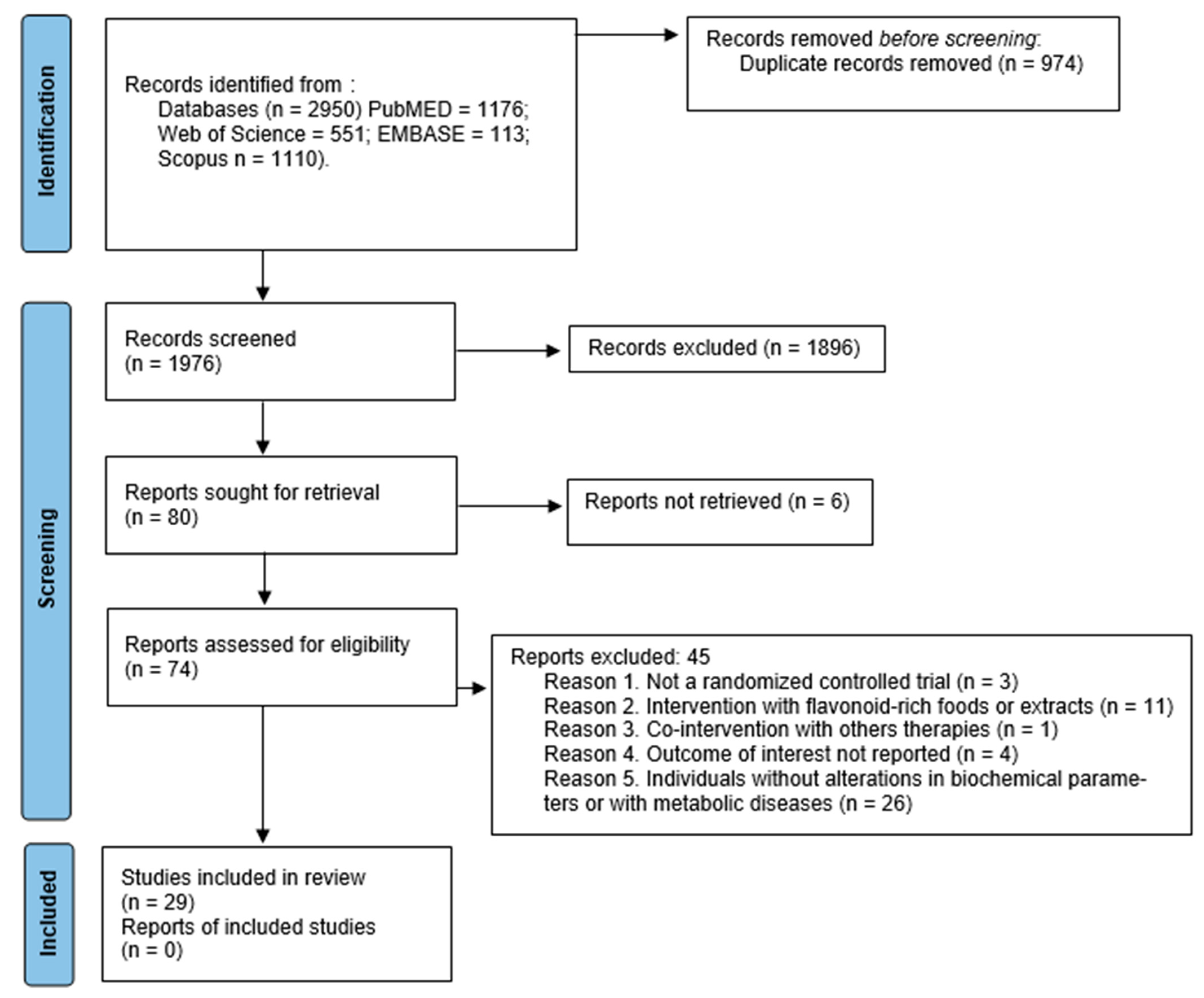
3.1. Search Results
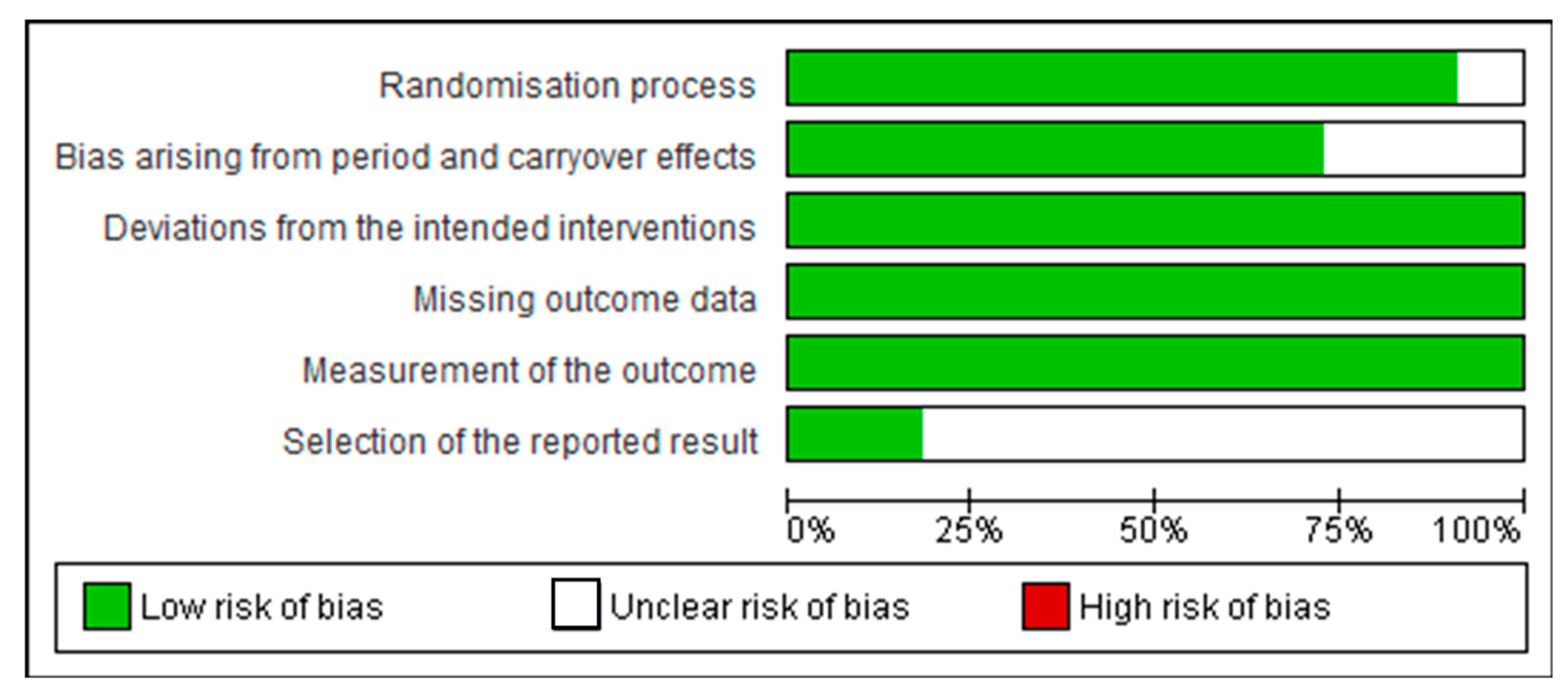
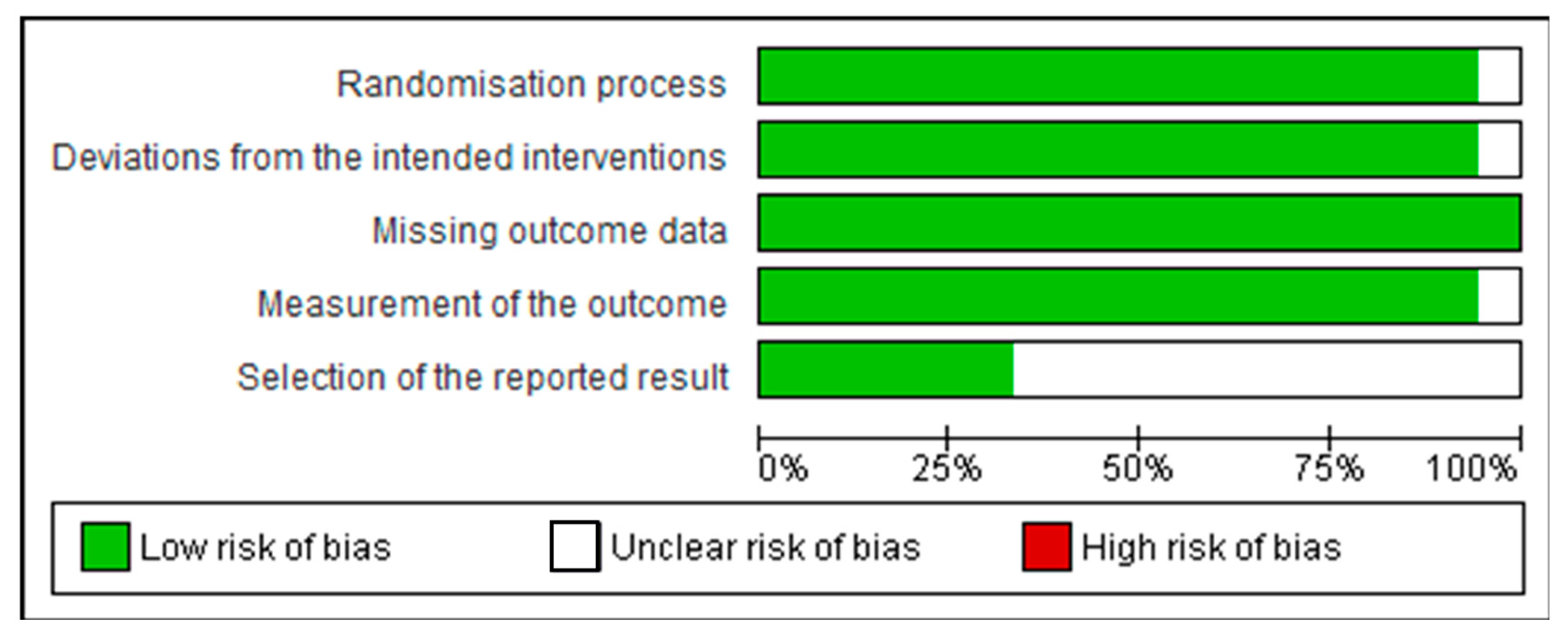
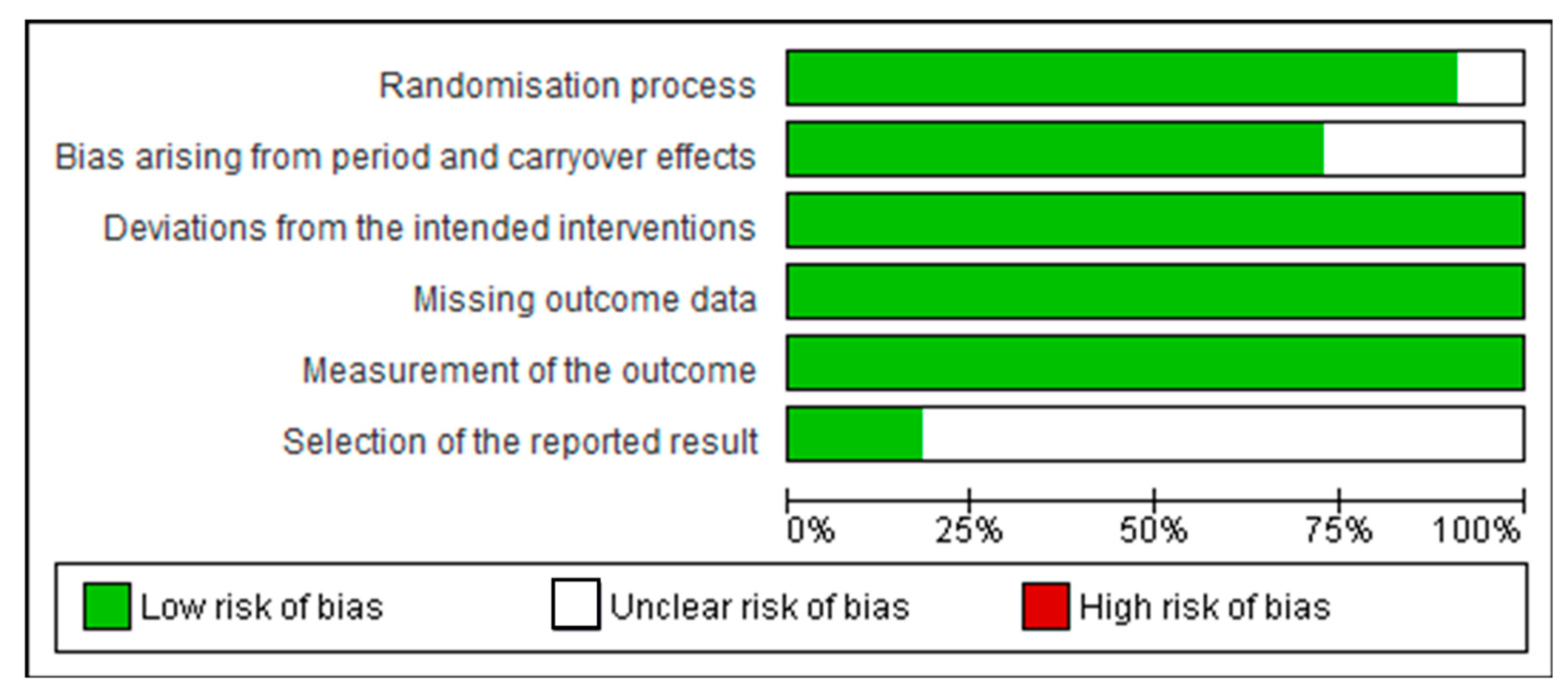
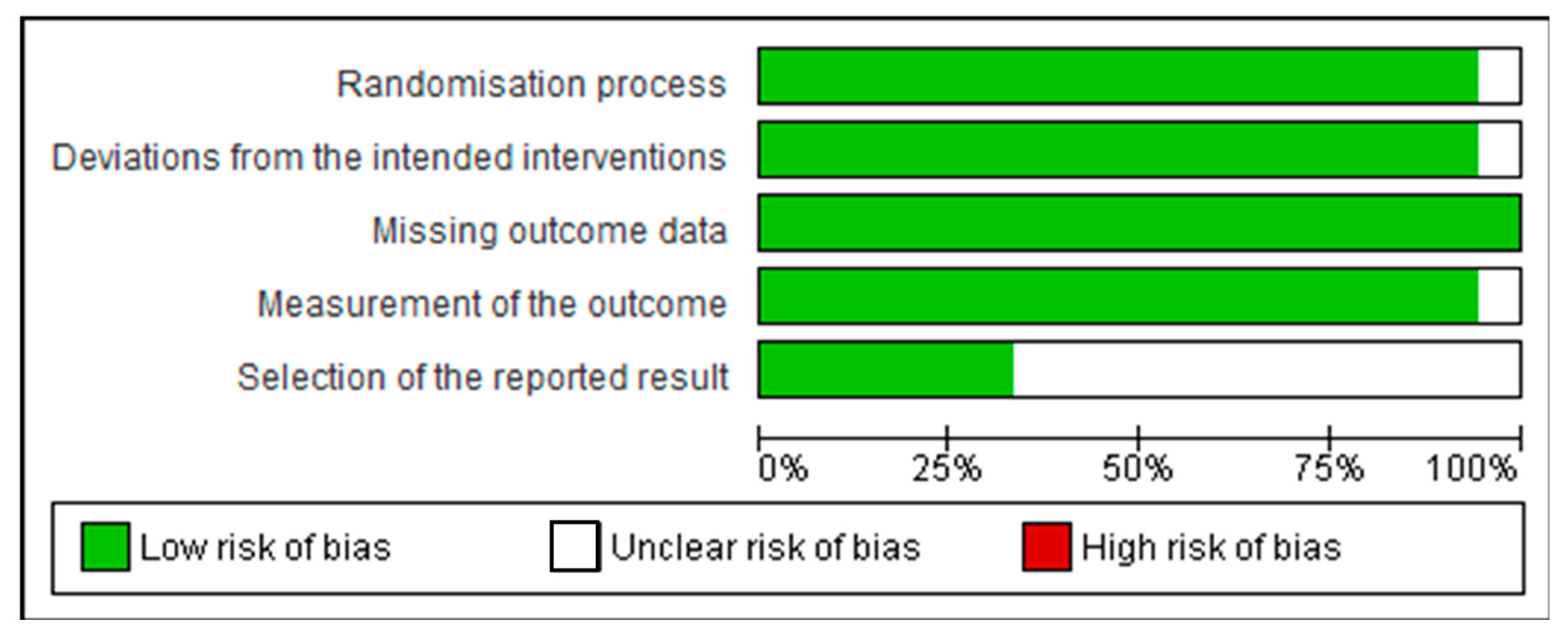
3.2. Assessment of Quality of Studies
3.3. Clinical and Methodological Characteristics
3.4. Intervention
3.5. Outcomes
3.5.1. Anthocyanin-Associated Outcomes
3.5.2. Hesperidin-Associated Outcomes
3.5.3. Quercetin-Associated Outcomes
3.5.4. Epicatechin-Associated Outcomes
3.5.5. Epigallocatechin Gallate-Associated Outcomes
3.5.6. Genistein-Associated Outcomes
3.5.7. Theaflavins and Catechin-Associated Outcomes
3.5.8. Eriocitrin, Hesperidin, Naringin, and Didymin-Associated Outcomes
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Grundy, S.M.; Cleeman, J.I.; Daniels, S.R.; Donato, K.A.; Eckel, R.H.; Franklin, B.A.; Gordon, D.J.; Krauss, R.M.; Savage, P.J.; Smith, S.C.; et al. Diagnosis and Management of the Metabolic Syndrome: An American Heart Association/National Heart, Lung, and Blood Institute Scientific Statement. Circulation 2005, 112, 2735–2752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberti, K.G.M.M.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.C.; James, W.P.T.; Loria, C.M.; Smith, S.C. Harmonizing the Metabolic Syndrome: A Joint Interim Statement of the International Diabetes Federation Task Force on Epidemiology and Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International Atherosclerosis Society; and International Association for the Study of Obesity. Circulation 2009, 120, 1640–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranasinghe, P.; Mathangasinghe, Y.; Jayawardena, R.; Hills, A.P.; Misra, A. Prevalence and Trends of Metabolic Syndrome among Adults in the Asia-Pacific Region: A Systematic Review. BMC Public Health 2017, 17, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amiot, M.J.; Riva, C.; Vinet, A. Effects of Dietary Polyphenols on Metabolic Syndrome Features in Humans: A Systematic Review. Obes. Rev. 2016, 17, 573–586. [Google Scholar] [CrossRef] [PubMed]
- do Vale Moreira, N.C.; Hussain, A.; Bhowmik, B.; Mdala, I.; Siddiquee, T.; Fernandes, V.O.; Montenegro Júnior, R.M.; Meyer, H.E. Prevalence of Metabolic Syndrome by Different Definitions, and Its Association with Type 2 Diabetes, Pre-Diabetes, and Cardiovascular Disease Risk in Brazil. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 1217–1224. [Google Scholar] [CrossRef] [PubMed]
- Després, J.P.; Lemieux, I. Abdominal Obesity and Metabolic Syndrome. Nature 2006, 444, 881–887. [Google Scholar] [CrossRef]
- World Health Organization. WHO Reveals Leading Causes of Death and Disability Worldwide: 2000–2019. Available online: https://www.who.int/news/item/09-12-2020-who-reveals-leading-causes-of-death-and-disability-worldwide-2000-2019 (accessed on 23 July 2022).
- Paley, C.A.; Johnson, M.I. Abdominal Obesity and Metabolic Syndrome: Exercise as Medicine? BMC Sports Sci. Med. Rehabil. 2018, 10, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-González, M.A.; Salas-Salvadó, J.; Estruch, R.; Corella, D.; Fitó, M.; Ros, E. Benefits of the Mediterranean Diet: Insights From the PREDIMED Study. Prog. Cardiovasc. Dis. 2015, 58, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Finicelli, M.; Squillaro, T.; Di Cristo, F.; Di Salle, A.; Melone, M.A.B.; Galderisi, U.; Peluso, G. Metabolic Syndrome, Mediterranean Diet, and Polyphenols: Evidence and Perspectives. J. Cell. Physiol. 2019, 234, 5807–5826. [Google Scholar] [CrossRef]
- Sohrab, G.; Ebrahimof, S.; Hosseinpour-Niazi, S.; Yuzbashian, E.; Mirmiran, P.; Azizi, F. Association of Dietary Intakes of Total Polyphenol and Its Subclasses with the Risk of Metabolic Syndrome: Tehran Lipid and Glucose Study. Metab. Syndr. Relat. Disord. 2018, 16, 274–281. [Google Scholar] [CrossRef]
- Lecour, S.; Lamont, K.T. Natural Polyphenols and Cardioprotection. Mini-Rev. Med. Chem. 2012, 11, 1191–1199. [Google Scholar] [CrossRef]
- Singla, R.K.; Dubey, A.K.; Garg, A.; Sharma, R.K.; Fiorino, M.; Ameen, S.M.; Haddad, M.A.; Al-Hiary, M. Natural Polyphenols: Chemical Classification, Definition of Classes, Subcategories, and Structures. J. AOAC Int. 2019, 102, 1397–1400. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macready, A.L.; Kennedy, O.B.; Ellis, J.A.; Williams, C.M.; Spencer, J.P.E.; Butler, L.T. Flavonoids and Cognitive Function: A Review of Human Randomized Controlled Trial Studies and Recommendations for Future Studies. Genes Nutr. 2009, 4, 227–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Lee, I.M.; Zhang, S.M.; Blumberg, J.B.; Buring, J.E.; Sesso, H.D. Dietary Intake of Selected Flavonols, Flavones, and Flavonoid-Rich Foods and Risk of Cancer in Middle-Aged and Older Women. Am. J. Clin. Nutr. 2009, 89, 905–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cassidy, A.; O’Reilly, É.J.; Kay, C.; Sampson, L.; Franz, M.; Forman, J.P.; Curhan, G.; Rimm, E.B. Habitual Intake of Flavonoid Subclasses and Incident Hypertension in Adults. Am. J. Clin. Nutr. 2011, 93, 338–347. [Google Scholar] [CrossRef] [Green Version]
- Hertog, M.; Kromhout, D.; Aravanis, C.; Blackburn, H.; Buzina, R.; Fidanza, F.; Giampaoli, S.; Jansen, A.; Menotti, A.; Nedeljkovic, S.; et al. Flavonoid Intake and Long-Term Risk in the Seven Countries Study. Arch. Intern Med. 1995, 155, 381–386. [Google Scholar] [CrossRef]
- Mink, P.J.; Scrafford, C.G.; Barraj, L.M.; Harnack, L.; Hong, C.P.; Nettleton, J.A.; Jacobs, D.R. Flavonoid Intake and Cardiovascular Disease Mortality: A Prospective Study in Postmenopausal Women. Am. J. Clin. Nutr. 2007, 85, 895–909. [Google Scholar] [CrossRef] [Green Version]
- Akbari, M.; Tamtaji, O.R.; Lankarani, K.B.; Tabrizi, R.; Dadgostar, E.; Haghighat, N.; Kolahdooz, F.; Ghaderi, A.; Mansournia, M.A.; Asemi, Z. The Effects of Resveratrol on Lipid Profiles and Liver Enzymes in Patients with Metabolic Syndrome and Related Disorders: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Lipids Health Dis. 2020, 19, 25. [Google Scholar] [CrossRef] [Green Version]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Reprint-preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. Phys. Ther. 2009, 89, 873–880. [Google Scholar] [CrossRef]
- Sterne, J.A.C.; Savović, J.; Page, M.J.; Elbers, R.G.; Blencowe, N.S.; Boutron, I.; Cates, C.J.; Cheng, H.Y.; Corbett, M.S.; Eldridge, S.M.; et al. RoB 2: A Revised Tool for Assessing Risk of Bias in Randomised Trials. BMJ 2019, 366, l4898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Yang, L.; Ling, W.; Qiu, Y.; Liu, Y.; Wang, L.; Yang, J.; Wang, C.; Ma, J. Anthocyanins Increase Serum Adiponectin in Newly Diagnosed Diabetes but Not in Prediabetes: A Randomized Controlled Trial. Nutr. Metab. 2020, 17, 78. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Xia, M.; Yang, Y.; Liu, F.; Li, Z.; Hao, Y.; Mi, M.; Jin, T.; Ling, W. Purified Anthocyanin Supplementation Improves Endothelial Function via NO-CGMP Activation in Hypercholesterolemic Individuals. Clin. Chem. 2011, 57, 1524–1533. [Google Scholar] [CrossRef] [Green Version]
- Pfeuffer, M.; Auinger, A.; Bley, U.; Kraus-Stojanowic, I.; Laue, C.; Winkler, P.; Rüfer, C.E.; Frank, J.; Bösch-Saadatmandi, C.; Rimbach, G.; et al. Effect of Quercetin on Traits of the Metabolic Syndrome, Endothelial Function and Inflammation in Men with Different APOE Isoforms. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 403–409. [Google Scholar] [CrossRef]
- Xu, Z.; Xie, J.; Zhang, H.; Pang, J.; Li, Q.; Wang, X.; Xu, H.; Sun, X.; Zhao, H.; Yang, Y.; et al. Anthocyanin Supplementation at Different Doses Improves Cholesterol Efflux Capacity in Subjects with Dyslipidemia—A Randomized Controlled Trial. Eur. J. Clin. Nutr. 2021, 75, 345–354. [Google Scholar] [CrossRef]
- Zhao, Y.; Xu, H.; Tian, Z.; Wang, X.; Xu, L.; Li, K.; Gao, X.; Fan, D.; Ma, X.; Ling, W.; et al. Dose-Dependent Reductions in Plasma Ceramides after Anthocyanin Supplementation Are Associated with Improvements in Plasma Lipids and Cholesterol Efflux Capacity in Dyslipidemia: A Randomized Controlled Trial. Clin. Nutr. 2021, 40, 1871–1878. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Xia, M.; Ma, J.; Hao, Y.; Liu, J.; Mou, H.; Cao, L.; Ling, W. Anthocyanin Supplementation Improves Serum LDL- and HDL-Cholesterol Concentrations Associated with the Inhibition of Cholesteryl Ester Transfer Protein in Dyslipidemic Subjects. Am. J. Clin. Nutr. 2009, 90, 485–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodgson, J.M.; Puddey, I.B.; Beilin, L.J.; Mori, T.A.; Croft, K.D. Supplementation with Isoflavonoid Phytoestrogens Does Not Alter Serum Lipid Concentrations: A Randomized Controlled Trial in Humans. J. Nutr. 1998, 128, 728–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trautwein, E.A.; Du, Y.; Meynen, E.; Yan, X.; Wen, Y.; Wang, H.; Molhuizen, H.O.F. Purified Black Tea Theaflavins and Theaflavins/Catechin Supplements Did Not Affect Serum Lipids in Healthy Individuals with Mildly to Moderately Elevated Cholesterol Concentrations. Eur. J. Nutr. 2010, 49, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Ling, W.; Guo, H.; Song, F.; Ye, Q.; Zou, T.; Li, D.; Zhang, Y.; Li, G.; Xiao, Y.; et al. Anti-Inflammatory Effect of Purified Dietary Anthocyanin in Adults with Hypercholesterolemia: A Randomized Controlled Trial. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Shu, F.; Zeng, Y.; Meng, X.; Wang, B.; Diao, L.; Wang, L.; Wan, J.; Zhu, J.; Wang, J.; et al. Daidzein Supplementation Decreases Serum Triglyceride and Uric Acid Concentrations in Hypercholesterolemic Adults with the Effect on Triglycerides Being Greater in Those with the GA Compared with the GG Genotype of ESR-β RsaI. J. Nutr. 2014, 144, 49–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhu, Y.; Song, F.; Yao, Y.; Ya, F.; Li, D.; Ling, W.; Yang, Y. Effects of Purified Anthocyanin Supplementation on Platelet Chemokines in Hypocholesterolemic Individuals: A Randomized Controlled Trial. Nutr. Metab. 2016, 13, 86. [Google Scholar] [CrossRef] [Green Version]
- Ohara, T.; Muroyama, K.; Yamamoto, Y.; Murosaki, S. Oral Intake of a Combination of Glucosyl Hesperidin and Caffeine Elicits an Anti-Obesity Effect in Healthy, Moderately Obese Subjects: A Randomized Double-Blind Placebo-Controlled Trial. Nutr. J. 2016, 15, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassellund, S.S.; Flaa, A.; Sandvik, L.; Kjeldsen, S.E.; Rostrup, M. Effects of Anthocyanins on Blood Pressure and Stress Reactivity: A Double-Blind Randomized Placebo-Controlled Crossover Study. J. Hum. Hypertens. 2012, 26, 396–404. [Google Scholar] [CrossRef] [PubMed]
- Hassellund, S.S.; Flaa, A.; Kjeldsen, S.E.; Seljeflot, I.; Karlsen, A.; Erlund, I.; Rostrup, M. Effects of Anthocyanins on Cardiovascular Risk Factors and Inflammation in Pre-Hypertensive Men: A Double-Blind Randomized Placebo-Controlled Crossover Study. J. Hum. Hypertens. 2013, 27, 100–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salden, B.N.; Troost, F.J.; De Groot, E.; Stevens, Y.R.; Garcés-Rimón, M.; Possemiers, S.; Winkens, B.; Masclee, A.A. Randomized Clinical Trial on the Efficacy of Hesperidin 2S on Validated Cardiovascular Biomarkers in Healthy Overweight Individuals. Am. J. Clin. Nutr. 2016, 104, 1523–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dower, J.I.; Geleijnse, J.M.; Gijsbers, L.; Zock, P.L.; Kromhout, D.; Hollman, P.C.H. Effects of the Pure Flavonoids Epicatechin and Quercetin on Vascular Function and Cardiometabolic Health: A Randomized, Double-Blind, Placebo-Controlled, Crossover Trial. Am. J. Clin. Nutr. 2015, 101, 914–921. [Google Scholar] [CrossRef] [PubMed]
- Esser, D.; Geleijnse, J.M.; Matualatupauw, J.C.; Dower, J.I.; Kromhout, D.; Hollman, P.C.H.; Afman, L.A. Pure Flavonoid Epicatechin and Whole Genome Gene Expression Profiles in Circulating Immune Cells in Adults with Elevated Blood Pressure: A Randomised Double-Blind, Placebo-Controlled, Crossover Trial. PLoS ONE 2018, 13, e0194229. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, C.B.; Ramos, F.M.; Manthey, J.A.; Cesar, T.B. Effectiveness of Eriomin® in Managing Hyperglycemia and Reversal of Prediabetes Condition: A Double-Blind, Randomized, Controlled Study. Phyther. Res. 2019, 33, 1921–1933. [Google Scholar] [CrossRef] [Green Version]
- Morand, C.; Dubray, C.; Milenkovic, D.; Lioger, D.; Martin, J.F.; Scalbert, A.; Mazur, A. Hesperidin Contributes to the Vascular Protective Effects of Orange Juice: A Randomized Crossover Study in Healthy Volunteers. Am. J. Clin. Nutr. 2011, 93, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Egert, S.; Bosy-Westphal, A.; Seiberl, J.; Kuerbitz, C.; Settler, U.; Plachta-Danielzik, S.; Wagner, A.E.; Frank, J.; Schrezenmeir, J.; Rimbach, G.; et al. Quercetin Reduces Systolic Blood Pressure and Plasma Oxidised Low-Density Lipoprotein Concentrations in Overweight Subjects with a High-Cardiovascular Disease Risk Phenotype: A Double-Blinded, Placebo-Controlled Cross-over Study. Br. J. Nutr. 2009, 102, 1065–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirch, N.; Berk, L.; Liegl, Y.; Adelsbach, M.; Zimmermann, B.F.; Stehle, P.; Stoffel-Wagner, B.; Ludwig, N.; Schieber, A.; Helfrich, H.-P.; et al. A Nutritive Dose of Pure (-)-Epicatechin Does Not Beneficially Affect Increased Cardiometabolic Risk Factors in Overweight-to-Obese Adults—A Randomized, Placebo-Controlled, Double-Blind Crossover Study. Am. J. Clin. Nutr. 2018, 107, 948–956. [Google Scholar] [CrossRef]
- Chatree, S.; Sitticharoon, C.; Maikaew, P.; Pongwattanapakin, K.; Keadkraichaiwat, I.; Churintaraphan, M.; Sripong, C.; Sririwichitchai, R.; Tapechum, S. Epigallocatechin Gallate Decreases Plasma Triglyceride, Blood Pressure, and Serum Kisspeptin in Obese Human Subjects. Exp. Biol. Med. 2021, 246, 163–176. [Google Scholar] [CrossRef]
- Brown, A.L.; Lane, J.; Coverly, J.; Stocks, J.; Jackson, S.; Stephen, A.; Bluck, L.; Coward, A.; Hendrickx, H. Effects of Dietary Supplementation with the Green Tea Polyphenol Epigallocatechin-3-Gallate on Insulin Resistance and Associated Metabolic Risk Factors: Randomized Controlled Trial. Br. J. Nutr. 2009, 101, 886–894. [Google Scholar] [CrossRef] [PubMed]
- Guevara-Cruz, M.; Godinez-Salas, E.T.; Sanchez-Tapia, M.; Torres-Villalobos, G.; Pichardo-Ontiveros, E.; Guizar-Heredia, R.; Arteaga-Sanchez, L.; Gamba, G.; Mojica-Espinosa, R.; Schcolnik-Cabrera, A.; et al. Genistein Stimulates Insulin Sensitivity through Gut Microbiota Reshaping and Skeletal Muscle AMPK Activation in Obese Subjects. BMJ Open Diabetes Res. Care 2020, 8, e000948. [Google Scholar] [CrossRef] [Green Version]
- Rizza, S.; Muniyappa, R.; Iantorno, M.; Kim, J.; Chen, H.; Pullikotil, P.; Senese, N.; Tesauro, M.; Lauro, D.; Cardillo, C.; et al. Citrus Polyphenol Hesperidin Stimulates Production of Nitric Oxide in Endothelial Cells While Improving Endothelial Function and Reducing Inflammatory Markers in Patients with Metabolic Syndrome. J. Clin. Endocrinol. Metab. 2011, 96, E782–E792. [Google Scholar] [CrossRef]
- Yari, Z.; Cheraghpour, M.; Hekmatdoost, A. Flaxseed and/or Hesperidin Supplementation in Metabolic Syndrome: An Open-Labeled Randomized Controlled Trial. Eur. J. Nutr. 2021, 60, 287–298. [Google Scholar] [CrossRef]
- Yari, Z.; Movahedian, M.; Imani, H.; Alavian, S.M.; Hedayati, M.; Hekmatdoost, A. The Effect of Hesperidin Supplementation on Metabolic Profiles in Patients with Metabolic Syndrome: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Eur. J. Nutr. 2020, 59, 2569–2577. [Google Scholar] [CrossRef]
- Egert, S.; Boesch-Saadatmandi, C.; Wolffram, S.; Rimbach, G.; Müller, M.J. Serum Lipid and Blood Pressure Responses to Quercetin Vary in Overweight Patients by Apolipoprotein E Genotype. J. Nutr. 2010, 140, 278–284. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Huang, X.; Zhang, Y.; Wang, Y.; Liu, Y.; Sun, R.; Xia, M. Anthocyanin Supplementation Improves HDL-Associated Paraoxonase 1 Activity and Enhances Cholesterol Efflux Capacity in Subjects With Hypercholesterolemia. J. Clin. Endocrinol. Metab. 2014, 99, 561–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, K.; Pederick, W.; Singh, I.; Santhakumar, A.B. Anthocyanin Supplementation in Alleviating Thrombogenesis in Overweight and Obese Population: A Randomized, Double-Blind, Placebo-Controlled Study. J. Funct. Foods 2017, 32, 131–138. [Google Scholar] [CrossRef]
- Assmann, G.; Schulte, H. Relation of High-Density Lipoprotein Cholesterol and Triglycerides to Incidence of Atherosclerotic Coronary Artery Disease (the PROCAM Experience). Am. J. Cardiol. 1992, 70, 733–737. [Google Scholar] [CrossRef]
- Lewis, G.F.; Rader, D.J. New Insights into the Regulation of HDL Metabolism and Reverse Cholesterol Transport. Circ. Res. 2005, 96, 1221–1232. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, S.J.; Nelson, A.J. HDL and Cardiovascular Disease. Pathology 2019, 51, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Fielding, C.J.; Fielding, P.E. Molecular Physiology of Reverse Cholesterol Transport. J. Lipid Res. 1995, 36, 211–228. [Google Scholar] [CrossRef]
- Barter, P.J.; Nicholls, S.; Rye, K.A.; Anantharamaiah, G.M.; Navab, M.; Fogelman, A.M. Antiinflammatory Properties of HDL. Circ. Res. 2004, 95, 764–772. [Google Scholar] [CrossRef] [PubMed]
- deGoma, E.M.; DeGoma, R.L.; Rader, D.J. Beyond High-Density Lipoprotein Cholesterol Levels. J. Am. Coll. Cardiol. 2008, 51, 2199–2211. [Google Scholar] [CrossRef] [Green Version]
- Durrington, P.N.; Mackness, B.; Mackness, M.I. Paraoxonase and Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 473–480. [Google Scholar] [CrossRef] [Green Version]
- Brown, B.G.; Stukovsky, K.H.; Zhao, X.Q. Simultaneous Low-Density Lipoprotein-C Lowering and High-Density Lipoprotein-C Elevation for Optimum Cardiovascular Disease Prevention with Various Drug Classes, and Their Combinations: A Meta-Analysis of 23 Randomized Lipid Trials. Curr. Opin. Lipidol. 2006, 17, 631–636. [Google Scholar] [CrossRef]
- Brousseau, M.E.; Schaefer, E.J.; Wolfe, M.L.; Bloedon, L.T.; Digenio, A.G.; Clark, R.W.; Mancuso, J.P.; Rader, D.J. Effects of an Inhibitor of Cholesteryl Ester Transfer Protein on HDL Cholesterol. N. Engl. J. Med. 2004, 350, 1505–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rader, D.J. Illuminating HDL—Is It Still a Viable Therapeutic Target? N. Engl. J. Med. 2007, 357, 2180–2183. [Google Scholar] [CrossRef] [PubMed]
- Khera, A.V.; Cuchel, M.; de la Llera-Moya, M.; Rodrigues, A.; Burke, M.F.; Jafri, K.; French, B.C.; Phillips, J.A.; Mucksavage, M.L.; Wilensky, R.L.; et al. Cholesterol Efflux Capacity, High-Density Lipoprotein Function, and Atherosclerosis. N. Engl. J. Med. 2011, 364, 127–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, K.C.; Wild, S.H.; Byrne, C.D. Controlling for Apolipoprotein A-I Concentrations Changes the Inverse Direction of the Relationship between High HDL-C Concentration and a Measure of Pre-Clinical Atherosclerosis. Atherosclerosis 2013, 231, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Assini, J.M.; Mulvihill, E.E.; Huff, M.W. Citrus Flavonoids and Lipid Metabolism. Curr. Opin. Lipidol. 2013, 24, 34–40. [Google Scholar] [CrossRef]
- Duval, C.; Müller, M.; Kersten, S. PPARα and Dyslipidemia. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2007, 1771, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Lee, M.K.; Park, Y.B.; Kang, M.A.; Choi, M.S. Effect of Citrus Flavonoids on Lipid Metabolism and Glucose-Regulating Enzyme MRNA Levels in Type-2 Diabetic Mice. Int. J. Biochem. Cell Biol. 2006, 38, 1134–1145. [Google Scholar] [CrossRef]
- Akiyama, S.; Katsumata, S.I.; Suzuki, K.; Nakaya, Y.; Ishimi, Y.; Uehara, M. Hypoglycemic and Hypolipidemic Effects of Hesperidin and Cyclodextrin-Clathrated Hesperetin in Goto-Kakizaki Rats with Type 2 Diabetes. Biosci. Biotechnol. Biochem. 2009, 73, 2779–2782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorinstein, S.; Caspi, A.; Libman, I.; Leontowicz, H.; Leontowicz, M.; Tashma, Z.; Katrich, E.; Jastrzebski, Z.; Trakhtenberg, S. Bioactivity of Beer and Its Influence on Human Metabolism. Int. J. Food Sci. Nutr. 2007, 58, 94–107. [Google Scholar] [CrossRef] [PubMed]
- Ohara, T.; Muroyama, K.; Yamamoto, Y.; Murosaki, S. A Combination of Glucosyl Hesperidin and Caffeine Exhibits an Anti-Obesity Effect by Inhibition of Hepatic Lipogenesis in Mice. Phyther. Res. 2015, 29, 310–316. [Google Scholar] [CrossRef] [PubMed]
- Stocker, R.; Keaney, J.F. Role of Oxidative Modifications in Atherosclerosis. Physiol. Rev. 2004, 84, 1381–1478. [Google Scholar] [CrossRef] [PubMed]
- Holvoet, P.; Kritchevsky, S.B.; Tracy, R.P.; Mertens, A.; Rubin, S.M.; Butler, J.; Goodpaster, B.; Harris, T.B. The Metabolic Syndrome, Circulating Oxidized LDL, and Risk of Myocardial Infarction in Well-Functioning Elderly People in the Health, Aging, and Body Composition Cohort. Diabetes 2004, 53, 1068–1073. [Google Scholar] [CrossRef] [Green Version]
- Wai, M.L.; Proudfoot, J.M.; Mckinley, A.J.; Needs, P.W.; Kroon, P.A.; Hodgson, J.M.; Croft, K.D. Quercetin and Its in Vivo Metabolites Inhibit Neutrophil-Mediated Low-Density Lipoprotein Oxidation. J. Agric. Food Chem. 2008, 56, 3609–3615. [Google Scholar] [CrossRef]
- Ikeda, I.; Tsuda, K.; Suzuki, Y.; Kobayashi, M.; Unno, T.; Tomoyori, H.; Goto, H.; Kawata, Y.; Imaizumi, K.; Nozawa, A.; et al. Tea Catechins with a Galloyl Moiety Suppress Postprandial Hypertriacylglycerolemia by Delaying Lymphatic Transport of Dietary Fat in Rats. J. Nutr. 2005, 135, 155–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Stillemark-Billton, P.; Beck, C.; Boström, P.; Andersson, L.; Rutberg, M.; Ericsson, J.; Magnusson, B.; Marchesan, D.; Ljungberg, A.; et al. Epigallocatechin Gallate Increases the Formation of Cytosolic Lipid Droplets and Decreases the Secretion of ApoB-100 VLDL. J. Lipid Res. 2006, 47, 67–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franklin, S.S. Systolic Blood Pressure: It’s Time to Take Control. Am. J. Hypertens. 2004, 17, S49–S54. [Google Scholar] [CrossRef] [Green Version]
- Law, M.R.; Morris, J.K.; Wald, N.J. Use of Blood Pressure Lowering Drugs in the Prevention of Cardiovascular Disease: Meta-Analysis of 147 Randomised Trials in the Context of Expectations from Prospective Epidemiological Studies. BMJ 2009, 338, b1665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grassi, D.; Desideri, G.; Croce, G.; Tiberti, S.; Aggio, A.; Ferri, C. Flavonoids, Vascular Function and Cardiovascular Protection. Curr. Pharm. Des. 2009, 15, 1072–1084. [Google Scholar] [CrossRef]
- Fraga, C.G.; Actis-Goretta, L.; Ottaviani, J.I.; Carrasquedo, F.; Lotito, S.B.; Lazarus, S.; Schmitz, H.H.; Keen, C.L. Regular Consumption of a Flavanol-Rich Chocolate Can Improve Oxidant Stress in Young Soccer Players. Clin. Dev. Immunol. 2005, 12, 11–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, M.; Suzuki, A.; Jokura, H.; Yamamoto, N.; Hase, T. Glucosyl Hesperidin Prevents Endothelial Dysfunction and Oxidative Stress in Spontaneously Hypertensive Rats. Nutrition 2008, 24, 470–476. [Google Scholar] [CrossRef]
- Turner, J.; Belch, J.J.F.; Khan, F. Current Concepts in Assessment of Microvascular Endothelial Function Using Laser Doppler Imaging and Iontophoresis. Trends Cardiovasc. Med. 2008, 18, 109–116. [Google Scholar] [CrossRef]
- Lorenz, M.; Wessler, S.; Follmann, E.; Michaelis, W.; Düsterhöft, T.; Baumann, G.; Stangl, K.; Stangl, V. A Constituent of Green Tea, Epigallocatechin-3-Gallate, Activates Endothelial Nitric Oxide Synthase by a Phosphatidylinositol-3-OH-Kinase-, CAMP-Dependent Protein Kinase-, and Akt-Dependent Pathway and Leads to Endothelial-Dependent Vasorelaxation. J. Biol. Chem. 2004, 279, 6190–6195. [Google Scholar] [CrossRef] [Green Version]
- Antonello, M.; Montemurro, D.; Bolognesi, M.; Dipascoli, M.; Piva, A.; Grego, F.; Sticchi, D.; Giuliani, L.; Garbisa, S.; Rossi, G. Prevention of Hypertension, Cardiovascular Damage and Endothelial Dysfunction with Green Tea Extracts. Am. J. Hypertens. 2007, 20, 1321–1328. [Google Scholar] [CrossRef] [Green Version]
- Mead, E.J.; Maguire, J.J.; Kuc, R.E.; Davenport, A.P. Kisspeptins Are Novel Potent Vasoconstrictors in Humans, with a Discrete Localization of Their Receptor, G Protein-Coupled Receptor 54, to Atherosclerosis-Prone Vessels. Endocrinology 2007, 148, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, I.; Smillie, S.J.; Bodkin, J.V.; Fernandes, E.; O’Byrne, K.T.; Brain, S.D. The Vasoactive Potential of Kisspeptin-10 in the Peripheral Vasculature. PLoS ONE 2011, 6, e14671. [Google Scholar] [CrossRef] [PubMed]
- Grassi, D.; Desideri, G.; Necozione, S.; Lippi, C.; Casale, R.; Properzi, G.; Blumberg, J.B.; Ferri, C. Blood Pressure Is Reduced and Insulin Sensitivity Increased in Glucose-Intolerant, Hypertensive Subjects after 15 Days of Consuming High-Polyphenol Dark Chocolate. J. Nutr. 2008, 138, 1671–1676. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Lee, M.K.; Jeong, K.S.; Choi, M.S. The Hypoglycemic Effects of Hesperidin and Naringin Are Partly Mediated by Hepatic Glucose-Regulating Enzymes in C57BL/KsJ-Db/Db Mice. J. Nutr. 2004, 134, 2499–2503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, O.M.; Mahmoud, A.M.; Abdel-Moneim, A.; Ashour, M.B. Antidiabetic Effects of Hesperidin and Naringin in Type 2 Diabetic Rats. Diabetol. Croat. 2012, 41, 53–67. [Google Scholar]
- Zhang, W.Y.; Lee, J.J.; Kim, Y.; Kim, I.S.; Han, J.H.; Lee, S.G.; Ahn, M.J.; Jung, S.H.; Myung, C.S. Effect of Eriodictyol on Glucose Uptake and Insulin Resistance in Vitro. J. Agric. Food Chem. 2012, 60, 7652–7658. [Google Scholar] [CrossRef]
- Bonora, E.; Kiechl, S.; Willeit, J.; Oberhollenzer, F.; Egger, G.; Meigs, J.B.; Bonadonna, R.C.; Muggeo, M. Insulin Resistance as Estimated by Homeostasis Model Assessment Predicts Incident Symptomatic Cardiovascular Disease in Caucasian Subjects from the General Population: The Bruneck Study. Diabetes Care 2007, 30, 318–324. [Google Scholar] [CrossRef] [Green Version]
- Katz, A.; Nambi, S.S.; Mather, K.; Baron, A.D.; Follmann, D.A.; Sullivan, G.; Quon, M.J. Quantitative Insulin Sensitivity Check Index: A Simple, Accurate Method for Assessing Insulin Sensitivity in Humans. J. Clin. Endocrinol. Metab. 2000, 85, 2402–2410. [Google Scholar] [CrossRef] [PubMed]
- Granado-Serrano, A.B.; Martín, M.A.; Haegeman, G.; Goya, L.; Bravo, L.; Ramos, S. Epicatechin Induces NF-ΚB, Activator Protein-1 (AP-1) and Nuclear Transcription Factor Erythroid 2p45-Related Factor-2 (Nrf2) via Phosphatidylinositol-3-Kinase/Protein Kinase B (PI3K/AKT) and Extracellular Regulated Kinase (ERK) Signalling in HepG2 Cells. Br. J. Nutr. 2010, 103, 168–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.L.; Ye, F.; Li, J.; Zhu, L.Y.; Feng, G.; Chang, X.Y.; Sun, K. Impaired Secretion of Glucagon-like Peptide 1 during Oral Glucose Tolerance Test in Patients with Newly Diagnosed Type 2 Diabetes Mellitus. Saudi Med. J. 2016, 37, 48–54. [Google Scholar] [CrossRef]
- Gastaldelli, A.; Gaggini, M.; DeFronzo, R. Glucose Kinetics: An Update and Novel Insights into Its Regulation by Glucagon and GLP-1. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 300–309. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia Muciniphila in Overweight and Obese Human Volunteers: A Proof-of-Concept Exploratory Study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.Y.; Park, J.S.; Noh, S.Y.; Rhee, E.J.; Sung, K.C.; Kim, B.S.; Kang, J.H.; Kim, S.W.; Lee, M.H.; Park, J.R. C-Reactive Protein Concentrations Are Related to Insulin Resistance and Metabolic Syndrome as Defined by the ATP III Report. Int. J. Cardiol. 2004, 97, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Ridker, P.M. Clinical Application of C-Reactive Protein for Cardiovascular Disease Detection and Prevention. Circulation 2003, 107, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Chu, W.-M. Tumor Necrosis Factor. Cancer Lett. 2013, 328, 222–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarro, J.F.; Mora, C. Diabetes, Inflammation, Proinflammatory Cytokines, and Diabetic Nephropathy. Sci. World J. 2006, 6, 712843. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K. Anti-Inflammatory Effects of Eriodictyol in Lipopolysaccharidestimulated Raw 264.7 Murine Macrophages. Arch. Pharm. Res. 2011, 34, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Gamo, K.; Miyachi, H.; Nakamura, K.; Matsuura, N. Hesperetin Glucuronides Induce Adipocyte Differentiation via Activation and Expression of Peroxisome Proliferator-Activated Receptor-γ. Biosci. Biotechnol. Biochem. 2014, 78, 1052–1059. [Google Scholar] [CrossRef]
- Francis, G.A.; Annicotte, J.S.; Auwerx, J. PPAR-α Effects on the Heart and Other Vascular Tissues. Am. J. Physiol. Heart. Circ. Physiol. 2003, 285, H1–H9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rufino, A.T.; Costa, V.M.; Carvalho, F.; Fernandes, E. Flavonoids as Antiobesity Agents: A Review. Med. Res. Rev. 2021, 41, 556–585. [Google Scholar] [CrossRef] [PubMed]
- Rohde, J.; Jacobsen, C.; Kromann-Andersen, H. Toxic Hepatitis Triggered by Green Tea. Ugeskr. Laeger 2011, 173, 205–206. [Google Scholar] [PubMed]
- Teschke, R.; Xuan, T.D. Suspected Herb Induced Liver Injury by Green Tea Extracts: Critical Review and Case Analysis Applying RUCAM for Causality Assessment. Jpn. J. Gastroenterol. Hepatol. 2019, 6, 1–16. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Authors | Type | Population | Gender | Age |
|---|---|---|---|---|
| Xu et al., 2021 [27] | Paralell | 40 mg (n = 45), 80 mg (n = 42), 320 mg (n = 43), placebo (n = 46) | 176 total, 26.1% males and 73.9% females | 57.41 ± 7.95 |
| Zhu et al., 2011 [25] | Crossover | 320 mg (n = 75), placebo (n = 75) | 146 total, 47.8% males and 52.2% females | 40–65 |
| Zhu et al., 2014 [52] | Paralell | anthocyanins (n = 61), placebo (n = 61) | 122 total, 42% males and 58% females | 55.1 ± 5.4 |
| Zhao et al., 2021 [28] | Paralell | 40 mg (n = 45), 80 mg (n = 42), 320 mg (n = 43), placebo (n = 46) | 169 total, 26.1% males and 73.9% females | 57.45 ± 0.86 |
| Thompson et al., 2017 [53] | Paralell | Antocyanins (n = 13), placebo (n = 13) | 26 total, 34.6% males and 65.4% females | 39 ± 11 |
| Hassellund et al., 2013 [37] | Crossover | Anthocyanins (n = 14), placebo (n = 13) | 27 males | 41 ± 3 |
| Hassellund et al., 2012 [36] | Crossover | Anthocyanins (n = 14), placebo (n = 13) | 27 males | 35–51 |
| Qin, et al., 2009 [29] | Paralell | Anthocynins (n = 60), placebo (n = 60) | 120 total, 35% males and 65% females | 55.1 ± 5.4 placebo, 55.3 ± 5.0 anthocyanin |
| Zhu et al., 2013 [32] | Paralell | Anthocyanins (n = 73), placebo (n = 73) | 146 total, 41.8% males and 58.2% females | 40–65 |
| Yang et al., 2020 [24] | Paralell | Anthocyanins prediabetes (n = 40), placebo prediabetes (n = 36) | 138 total, 33.75% males and 66.25% females | 61.2 ± 6.9 placebo, 60.8 ± 7.9 anthocyanin |
| Zhang et al., 2016 [34] | Paralell | Anthocyanins (n = 73), placebo (n = 73) | 146 total, 41.8% males and 58.2% females | 40–65 |
| Rizza et al., 2011 [48] | Crossover | Hesperidin (n = 12), placebo (n = 12) | 24 total, 62.5% males and 37.5% females | 51.66 ± 1.52 |
| Yari et al., 2020 [50] | Paralell | Hesperidin (n = 25), placebo (n = 24) | 49 total, 51% males and 49% females | 45.19 ± 11.11 |
| Morand et al., 2011 [42] | Crossover | Control drink + hesperidin (n = 8), control drink + placebo (n = 8) | 24 males | 56 ± 1 |
| Salden et al., 2016 [38] | Paralell | Hesperidin (n = 34), placebo (n = 34) | 68 total, 42.6% males and 57.4% females | 53 ± 14 |
| Yari et al., 2021 [49] | Paralell | Placebo (n = 22), Hesperidin (n = 22) | 44 total, 47.8% males and 52.2% females | 46.41 ± 11.10 control, 45.82 ± 11.69 hesperidin |
| Ohara et al., 2016 [35] | Paralell | Placebo (n = 15), G-Hesperidin (n = 15) | 74 total, 50% males and 50% females | 49.12 ± 1.24 |
| Egert et al., 2009 [43] | Crossover | Quercetin (n = 93), placebo (n = 93) | 93 total, 45.1% males and 54.9% females | 45.1 ± 10.53 |
| Egert et al., 2010 [51] | Crossover | apoE3 Quercetin (n = 86), E4 Quercetin (86), placebo (n = 86) | 93 total, 45.1% males and 54.9% females | 45.5 ± 9.45 |
| Pfeuffer et al., 2013 [26] | Crossover | apoE3/3 (n = 19), apoE4 (n = 30) | 49 total, 51% males and 49% females | 59.4 ± 0.9 |
| Dower et al., 2015 [39] | Crossover | Epicatechin (n = 11), Quercetin (n = 14), placebo (n = 12) | 37 total, 67.6% males and 32.4% females | 66.4 ± 7.9 |
| Esser et al., 2018 [40] | Crossover | Epicatechin (n = 32), placebo (n = 32) | 32 total, 62.5% males and 37.5% females | 65.8 ± 7.9 |
| Kirch et al., 2018 [44] | Crossover | Epicatechin (n = 24), placebo (n = 23) | 47 total, 53.2% males and 46.8% females | 36 ± 12 males, 35 ±16 females |
| Chatree et al., 2021 [45] | Paralell | EGCG (n = 15), placebo (n = 15) | 30, gender no specified | older than 18 years |
| Brown et al., 2009 [46] | Paralell | EGCG (n = 46), placebo (n = 42) | 88 males | 50.57 ± 56.48 placebo 52.15 ± 6.43 EGCG |
| Guevara-Cruz et al., 2020 [47] | Paralell | Genistein (n = 22), placebo (n = 23) | 45, gender no specified | 43.0 ± 2.28 placebo 42.6 ± 1.9 Genistein |
| Hodgson et al., 1998 [30] | Paralell | Isoflavonoid group (n = 30), placebo group (n = 29) | 59 total, 78% males and 22% females | 57.0 placebo and 54.3 Isoflav. |
| Trautwein et al., 2010 [31] | Paralell | Theaflavins (n = 34), Theaflavins + Catechin (n = 31) and placebo (n = 34) | 99 total, 65.7% males and 34.3% females | 48.1 ± 6.1 |
| Ribeiro et al., 2019 [41] | Paralell | 200 mg (n = 26), 400 mg (n = 27), 800 mg (n = 25), placebo (n = 25) | 103 total, 46.7% males and 52.4% females | 49 ± 10 |
| Authors | Condition | Flavonoid | Dosis | Duration |
|---|---|---|---|---|
| Xu et al., 2021 [27] | Dyslipidemia | Anthocyanin | 40, 80 or 320 mg/day | 12 weeks |
| Zhu et al., 2011 [25] | Hypercholesterolemia | Anthocyanin | 320 mg/day | 12 weeks |
| Zhu et al., 2014 [52] | Hypercholesterolemia | Anthocyanin | 320 mg/day | 24 weeks |
| Zhao et al., 2021 [28] | Dyslipidemia | Anthocyanin | 40, 80 or 320 mg/day | 12 weeks |
| Thompson et al., 2017 [53] | Pro-trombotic overweight and obese individuals | Anthocyanin | 320 mg/day | 4 weeks |
| Hassellund et al., 2013 [37] | Prehypertension | Anthocyanin | 640 mg/day | 4 weeks |
| Hassellund et al., 2012 [36] | Prehypertension | Anthocyanin | 640 mg/day | 4 weeks |
| Qin, et al., 2009 [29] | Dyslipidemia | Anthocyanin | 320 mg/day | 12 weeks |
| Zhu et al., 2013 [32] | Hypercholesterolemia | Anthocyanin | 320 mg/day | 24 weeks |
| Yang et al., 2020 [24] | Prediabetes | Anthocyanin | 320 mg/day | 12 weeks |
| Zhang et al., 2016 [34] | Hypercholesterolemia | Anthocyanin | 320 mg/day | 24 weeks |
| Rizza et al., 2011 [48] | Metabolic Syndrome | Hesperidin | 500 mg/day | 3 weeks |
| Yari et al., 2020 [50] | Metabolic Syndrome | Hesperidin | 1000 mg/day | 12 weeks |
| Morand et al., 2011 [42] | Obesisty | Hesperidin | 146 mg/day | 4 weeks |
| Salden et al., 2016 [38] | Overweight prehypertensive | Hesperidin | 450 mg/day | 6 weeks |
| Yari et al., 2021 [49] | Metabolic Syndrome | Hesperidin | 1000 mg/day | 12 weeks |
| Ohara et al., 2016 [35] | Overweight and hypertriglyceridemia | Glucosyl hesperidin | 500 mg/day | 12 weeks |
| Egert et al., 2009 [43] | Overweight or obesity | Quercetin | 150 mg/day | 6 weeks |
| Egert et al., 2010 [51] | Metabolic Syndrome | Quercetin | 150 mg/day | 6 weeks |
| Pfeuffer et al., 2013 [26] | APOE genotype | Quercetin | 150 mg/day | 8 weeks |
| Dower et al., 2015 [39] | Prehypertension | (-)-Epicatechin or Quercetin | epicatechin (100 mg/day) or quercetin-3-glucoside (160 mg/day) | 4 weeks |
| Esser et al., 2018 [40] | Prehypertension | (-)-Epicatechin | 100 mg/day | 4 weeks |
| Kirch et al., 2018 [44] | Overweight or obesity and MetS | (-)-Epicatechin | 25 mg/day | 2 weeks |
| Chatree et al., 2021 [45] | Obesity | Epigallocatechin gallate | 300 mg/day | 8 weeks |
| Brown et al., 2009 [46] | Overweight or obesity | Epigallocatechin gallate | 800 mg/day | 8 weeks |
| Guevara-Cruz et al., 2020 [47] | Obesity and insulin resistance | Genistein | 50 mg/day | 8 weeks |
| Hodgson et al., 1998 [30] | Predyslipidemia | Isoflavonoids, mainly genistein | 55 mg/day | 8 weeks |
| Trautwein et al., 2010 [31] | Dyslipidemia | Theaflavin and catechin | 75 mg theaflavins and 149.4 mg catechins | 11 weeks |
| Ribeiro et al., 2019 [41] | Prediabetes | 70% eriocitrin, 5% hesperidin, 4% naringin, and 1% didymin | 200, 400 or 800 mg/day | 12 weeks |
| Flavonoid | BW | BMI | WC | SBP | DBP | LDL-c | HDL-c | TG | TC | BG | IR |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Anthocyanin | |||||||||||
| Qin et al., 2009 [29] | = | = | = | = | = | ↓ | ↑ | = | = | = | NA |
| Hassellund et al., 2012 [36] | NA | NA | NA | = | = | NA | NA | NA | NA | NA | NA |
| Hassellund et al., 2013 [37] | NA | NA | NA | NA | NA | = | ↑ | = | = | ↑ | = |
| Zhu et al., 2011 [25] | = | = | = | = | = | ↓ | ↑ | = | = | = | = |
| Zhu et al., 2013 [32] | NA | NA | NA | NA | NA | ↓ | ↑ | = | = | NA | NA |
| Zhu et al., 2014 [52] | NA | NA | NA | NA | NA | ↓ | ↑ | = | = | = | = |
| Zhang et al., 2016 [34] | NA | NA | NA | NA | NA | ↓ | ↑ | = | = | NA | NA |
| Thompson et al., 2017 [53] | = | = | = | = | = | = | = | = | NA | = | NA |
| Yang et al., 2020 [24] | NA | NA | NA | NA | NA | = | = | = | = | = | = |
| Xu et al., 2021 [27] | = | = | = | = | = | = | ↑ | = | = | = | = |
| Zhao et al., 2021 [28] | = | = | = | = | = | = | ↑ | = | ↓ | = | = |
| Hesperidin | |||||||||||
| Morand et al., 2011 [42] | = | = | NA | = | ↓ | = | = | = | = | = | = |
| Rizza et al., 2011 [48] | = | = | = | = | = | = | ↑ | = | ↓ | = | ↓ |
| Salden et al., 2016 [38] | = | = | = | ↓ | ↓ | = | = | = | = | = | = |
| Ohara et al., 2016 [35] | = | = | = | NA | NA | ↓ | = | = | ↓ | NA | NA |
| Yari et al., 2020 [50] | = | = | = | ↓ | = | ↓ | = | ↓ | ↓ | ↓ | ↓ |
| Yari et al., 2021 [49] | = | = | = | ↓ | ↓ | NA | = | ↓ | NA | = | = |
| Quercetin | |||||||||||
| Egert et al., 2009 [43] | = | = | = | ↓ | = | = | ↓ | = | = | = | NA |
| Egert et al., 2010 [51] | = | = | = | ↓ | = | ↓ | ↓ | = | = | = | NA |
| Pfeuffer et al., 2013 [26] | = | = | ↓ | ↓ | = | = | ↑ | ↓ | = | = | = |
| Dower et al., 2015 [39] | = | = | NA | = | = | = | = | = | = | = | = |
| Epicatechin | |||||||||||
| Dower et al., 2015 [39] | = | = | NA | = | = | = | = | = | = | = | ↓ |
| Esser et al., 2018 [40] | = | = | NA | = | = | = | = | = | = | ↓ | ↓ |
| Kirch et al., 2018 [44] | = | = | = | = | = | = | = | = | = | = | = |
| Epigallocatechin G. | |||||||||||
| Brown et al., 2009 [46] | = | = | = | = | ↓ | = | = | = | = | = | = |
| Chatree et al., 2021 [45] | = | = | = | ↓ | ↓ | = | = | ↓ | = | = | = |
| Genistein | |||||||||||
| Hodgson et al., 1998 [30] | = | NA | NA | NA | NA | = | = | = | = | NA | NA |
| Guevara-Cruz et al., 2020 [47] | = | = | = | = | = | = | = | = | = | = | ↓ |
| Theaflavins and Catechin | |||||||||||
| Trautwein et al., 2010 [31] | = | = | NA | NA | NA | = | = | = | = | = | NA |
| Eriocitrin, Hesperidin, Naringin, and Didymin | |||||||||||
| Ribeiro et al., 2019 [41] | = | = | NA | ↓ | = | = | = | = | = | ↓ | ↓ |
| Flavonoid | TNF-α and hs-CRP | Others Markers |
|---|---|---|
| Anthocyanin | ||
| Qin et al., 2009 [29] | NA | ↓ mass and activity CETP, and ↑ cellular cholesterol efflux |
| Hassellund et al., 2012 [36] | NA | No changes in levels of renin, aldosterone, or angiotensin-converting enzyme |
| Hassellund et al., 2013 [37] | No changes in TNF-α and hs-CRP | ↑ von Willebrand factor |
| Zhu et al., 2011 [25] | NA | ↓ FMD, cGMP and adhesion molecules, such as sVCAM-1 |
| Zhu et al., 2013 [32] | No changes in TNF-α, ↓ hs-CRP | ↓ sVCAM-1 and IL-1β |
| Zhu et al., 2014 [52] | NA | ↑ cholesterol efflux capacity and HDL-PON1 activity |
| Zhang et al., 2016 [34] | No changes in TNF-α, ↓ hs-CRP | ↓ IL-1β and sP-selectin |
| Thompson et al., 2017 [53] | No changes in hs-CRP | ↓ (ADP)-induced platelet activation, PAC-1 and p-selectin |
| Yang et al., 2020 [24] | NA | No changes in adiponectin levels |
| Xu et al., 2021 [27] | NA | ↑ cholesterol efflux capacity and ApoA-I |
| Zhao et al., 2021 [28] | NA | ↑ cholesterol efflux capacity, ApoA-I and Apo B |
| Hesperidin | ||
| Morand et al., 2011 [42] | No changes in hs-CRP | ↑ endothelium-dependent vasodilation and ↓ interleukin-6 |
| Rizza et al., 2011 [48] | ↓ hs-CRP | ↑ FMD, ↓ apoB, and ↓ sE-selectin |
| Salden et al., 2016 [38] | NA | No changes in FMD, ↓ adhesion molecules such as sVCAM-1 and sICAM-1 |
| Ohara et al., 2016 [35] | NA | No changes in abdominal fat area (visceral and subcutaneous) |
| Yari et al., 2020 [50] | ↓ TNF-α and hs-CRP | NA |
| Yari et al., 2021 [49] | NA | NA |
| Quercetin | ||
| Egert et al., 2009 [43] | No changes in TNF-α and hs-CRP | ↓ ox-LDL, ↓ pulse pressure |
| Egert et al., 2010 [51] | ↓ TNF-α, no changes in hs-CRP | ↓ ox-LDL |
| Pfeuffer et al., 2013 [26] | ↑TNF-α, no changes in hs-CRP | No changes in s-E-Selectin, s-VCAM, s-ICAM, ox-LDL, and hs-CRP |
| Dower et al., 2015 [39] | NA | No changes in FMD or EID |
| Epicatechin | ||
| Dower et al., 2015 [39] | NA | No changes in FMD or EID |
| Esser et al., 2018 [40] | ↓ TNF upstream regulator | ↓ IL8-CXCR1/2 and AMB2_neutrophil genes |
| Kirch et al., 2018 [44] | NA | No changes in ox-LDL |
| Epigallocatechin G. | ||
| Brown et al., 2009 [46] | NA | ↑ mood (hedonic tone) |
| Chatree et al., 2021 [45] | NA | ↓ serum kisspeptin |
| Genistein | ||
| Hodgson et al., 1998 [30] | NA | No changes in lipoprotein (a) concentrations |
| Guevara-Cruz et al., 2020 [47] | ↓ hs-CRP | ↓ metabolic endotoxemia, ↑ skeletal muscle fatty acid oxidation |
| Theaflavins and Catechin | ||
| Trautwein et al., 2010 [31] | NA | NA |
| Eriocitrin, Hesperidin, Naringin, and Didymin | ||
| Ribeiro et al., 2019 [41] | ↓ TNF-α and hs-CRP | ↓ C-peptide and Interleukin-6, ↑ antioxidant capacity |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gouveia, H.J.C.B.; Urquiza-Martínez, M.V.; Manhães-de-Castro, R.; Costa-de-Santana, B.J.R.; Villarreal, J.P.; Mercado-Camargo, R.; Torner, L.; de Souza Aquino, J.; Toscano, A.E.; Guzmán-Quevedo, O. Effects of the Treatment with Flavonoids on Metabolic Syndrome Components in Humans: A Systematic Review Focusing on Mechanisms of Action. Int. J. Mol. Sci. 2022, 23, 8344. https://doi.org/10.3390/ijms23158344
Gouveia HJCB, Urquiza-Martínez MV, Manhães-de-Castro R, Costa-de-Santana BJR, Villarreal JP, Mercado-Camargo R, Torner L, de Souza Aquino J, Toscano AE, Guzmán-Quevedo O. Effects of the Treatment with Flavonoids on Metabolic Syndrome Components in Humans: A Systematic Review Focusing on Mechanisms of Action. International Journal of Molecular Sciences. 2022; 23(15):8344. https://doi.org/10.3390/ijms23158344
Chicago/Turabian StyleGouveia, Henrique J. C. B., Mercedes V. Urquiza-Martínez, Raul Manhães-de-Castro, Bárbara J. R. Costa-de-Santana, José Pérez Villarreal, Rosalío Mercado-Camargo, Luz Torner, Jailane de Souza Aquino, Ana E. Toscano, and Omar Guzmán-Quevedo. 2022. "Effects of the Treatment with Flavonoids on Metabolic Syndrome Components in Humans: A Systematic Review Focusing on Mechanisms of Action" International Journal of Molecular Sciences 23, no. 15: 8344. https://doi.org/10.3390/ijms23158344
APA StyleGouveia, H. J. C. B., Urquiza-Martínez, M. V., Manhães-de-Castro, R., Costa-de-Santana, B. J. R., Villarreal, J. P., Mercado-Camargo, R., Torner, L., de Souza Aquino, J., Toscano, A. E., & Guzmán-Quevedo, O. (2022). Effects of the Treatment with Flavonoids on Metabolic Syndrome Components in Humans: A Systematic Review Focusing on Mechanisms of Action. International Journal of Molecular Sciences, 23(15), 8344. https://doi.org/10.3390/ijms23158344