Sex-Specific Effects of Early Life Stress on Brain Mitochondrial Function, Monoamine Levels and Neuroinflammation

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Maternal Separation Procedure
2.2. CCO Quantitative Histochemistry
2.3. Determination of Brain Monoaminergic Activity
2.4. Real-Time PCR Analysis of mRNA Cytokine Expression
2.5. Statistical Analysis
3. Results
3.1. CCO Activity
3.1.1. Main Effects of Sex and Rearing
3.1.2. Interaction between Sex and Rearing
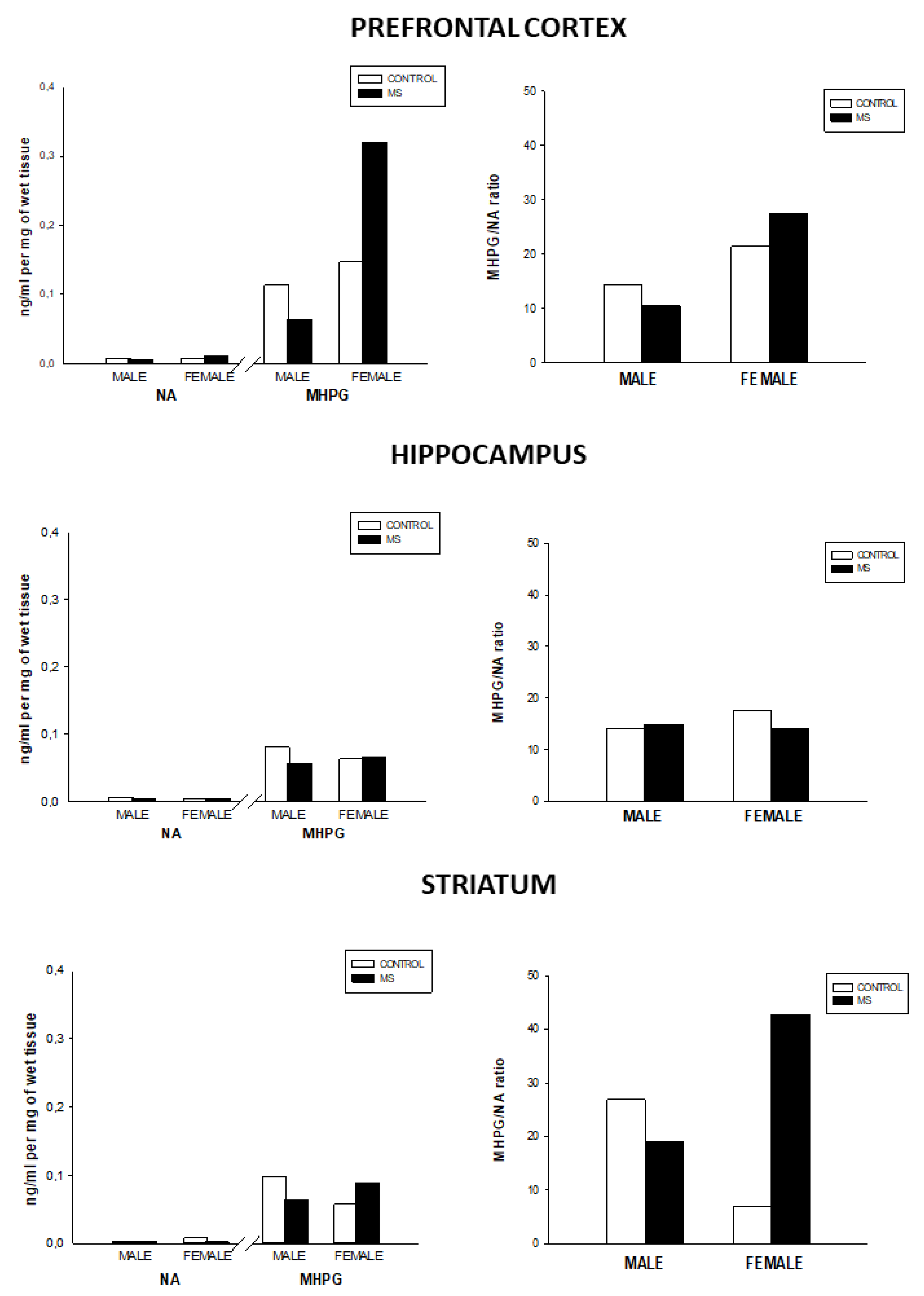
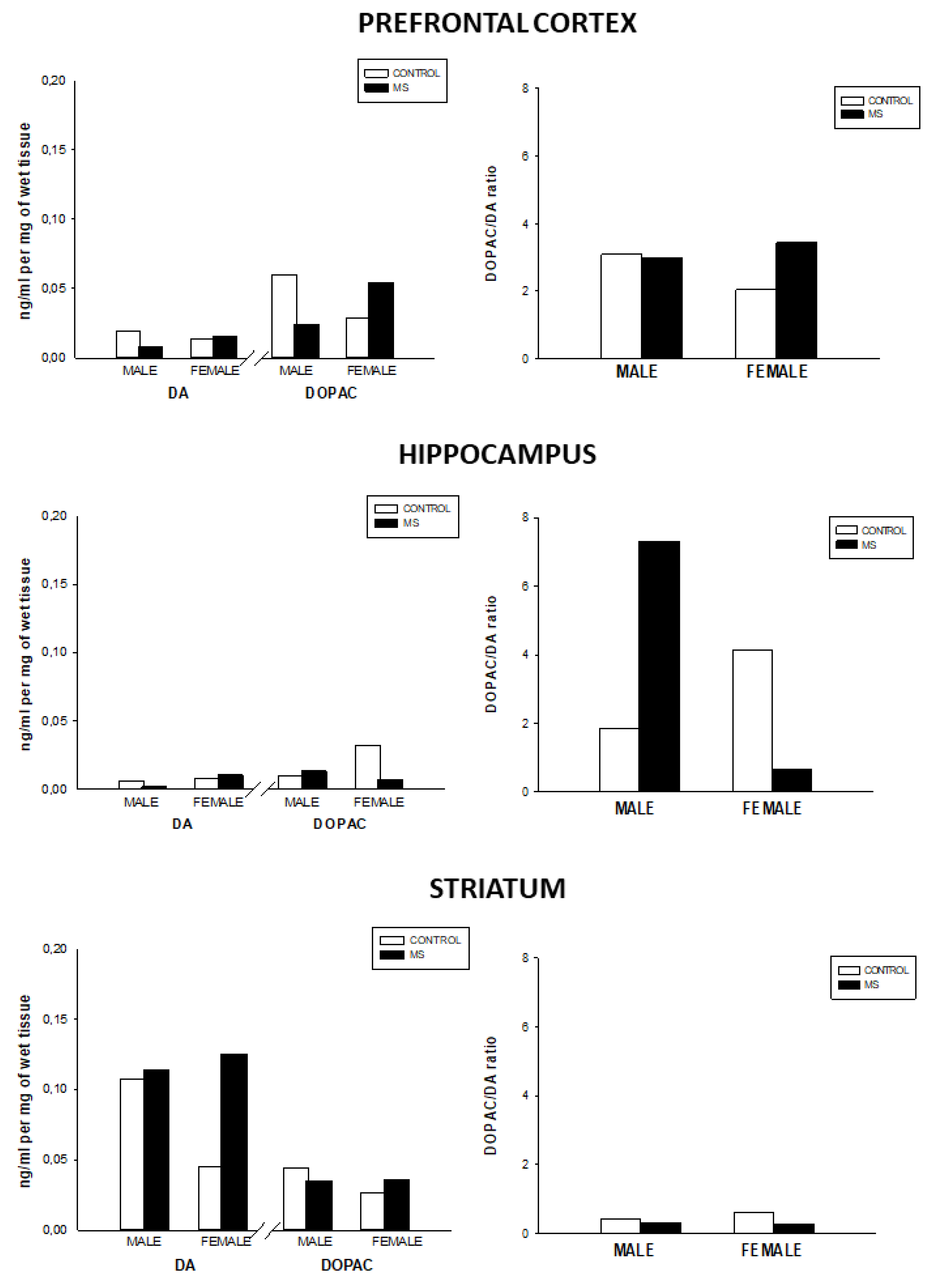
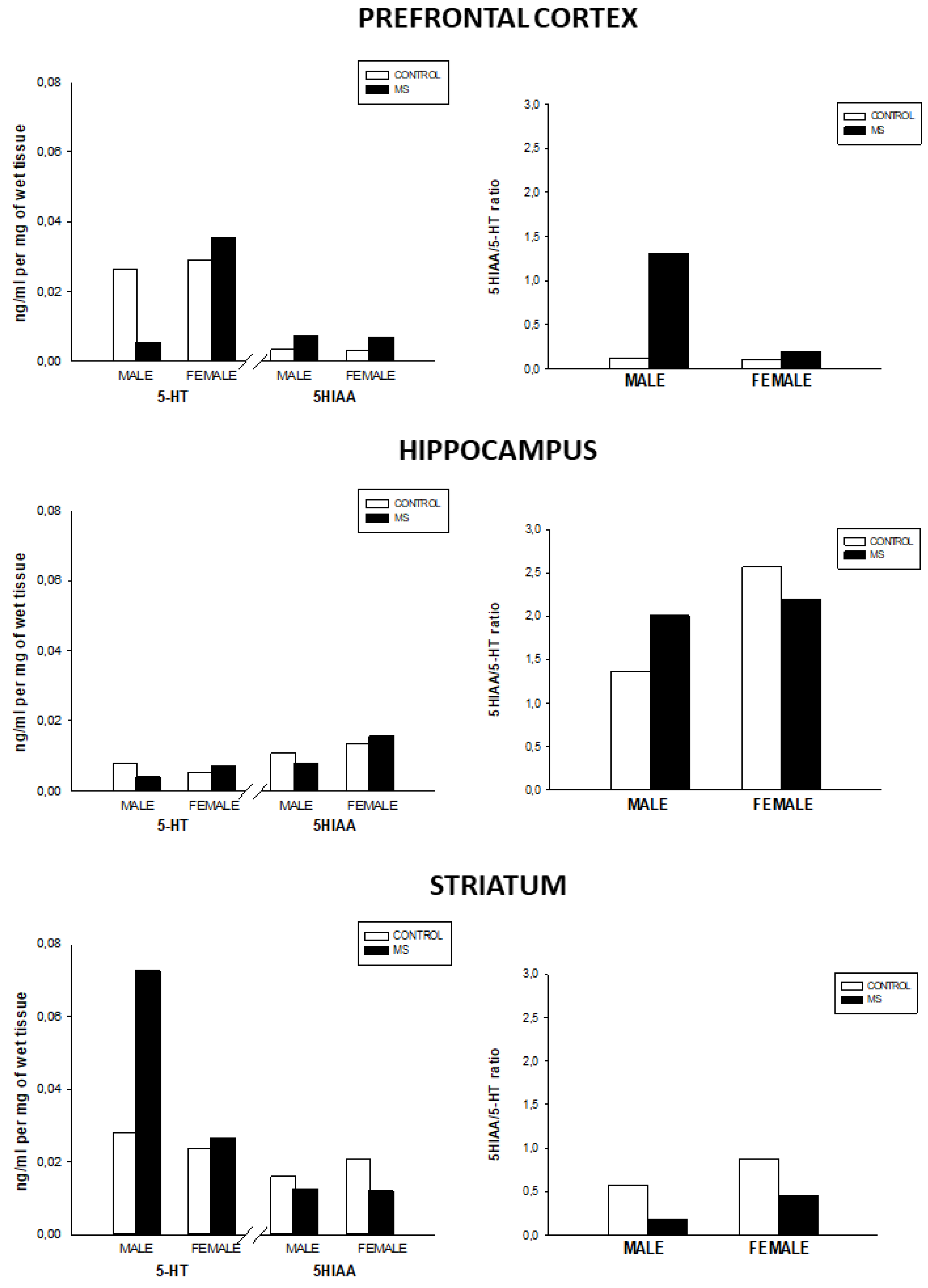
3.2. Effect of Sex and Rearing on the Concentrations of Monoamines Levels and Turnover in Selected Brain Regions
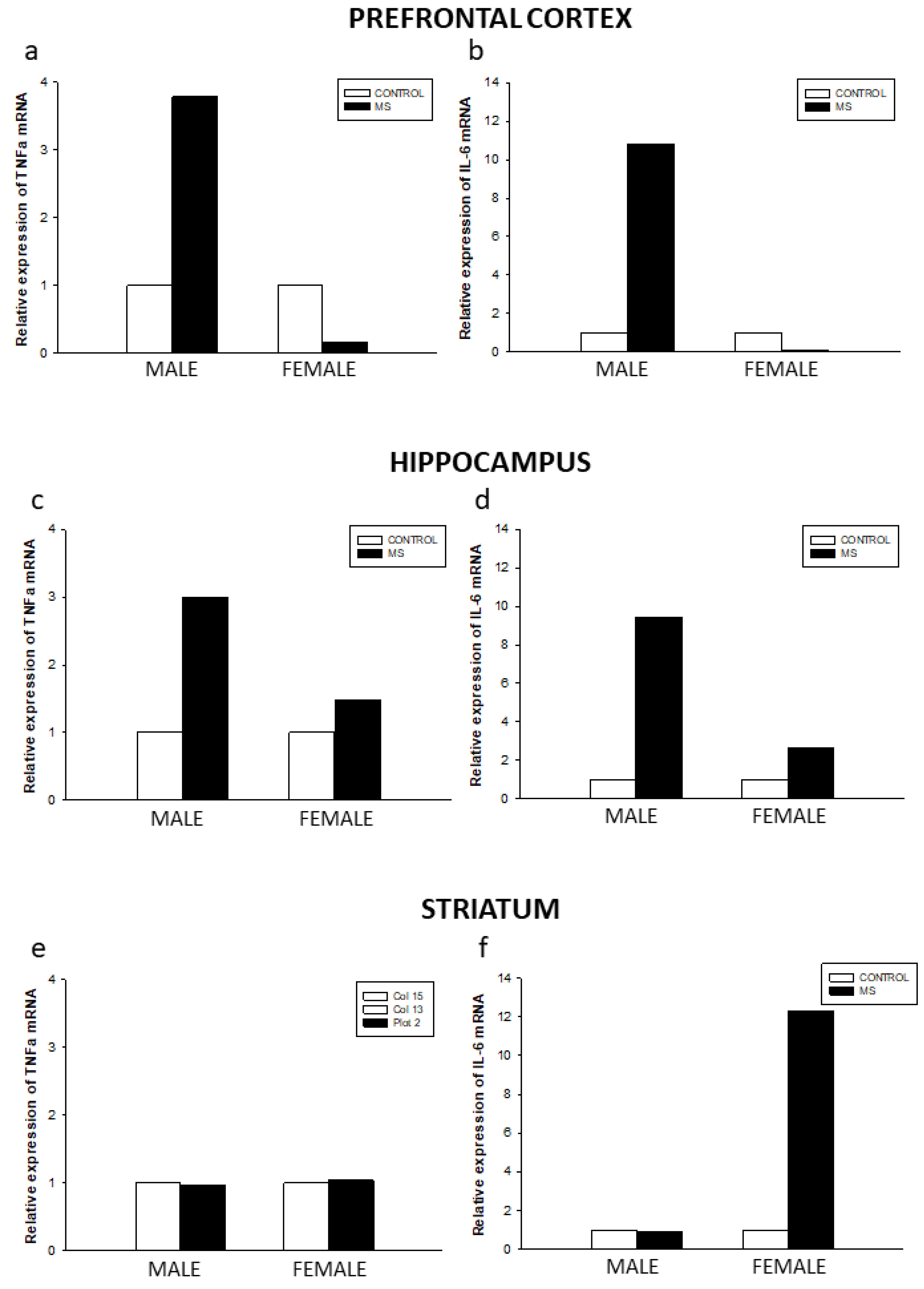
3.3. Effect of Sex and Rearing on Cytokine mRNA Levels in Selected Brain Regions
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Nemeroff, C.B. Paradise Lost: The Neurobiological and Clinical Consequences of Child Abuse and Neglect. Neuron 2016, 89, 892–909. [Google Scholar] [CrossRef]
- Johnson, S.B.; Riley, A.W.; Granger, D.A.; Riis, J. The science of Early Life Toxic Stress for Pediatric Practice and Advocacy. Pediatrics 2013, 131, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.E.; Chen, E.; Parker, K.J. Psychological Stress in Childhood and Susceptibility to the Chronic Diseases of Aging: Moving Toward a Model of Behavioral and Biological Mechanisms. Psychol. Bull. 2011, 137, 959–997. [Google Scholar] [CrossRef] [PubMed]
- Nusslock, R.; Miller, G.E. Early-Life Adversity and Physical and Emotional Health across the Lifespan: A Neuroimmune Network Hypothesis. Biol. Psychiatry 2016, 80, 23–32. [Google Scholar] [CrossRef]
- Sale, A. A Systematic Look at Environmental Modulation and Its Impact in Brain Development. Trends Neurosci. 2018, 41, 4–17. [Google Scholar] [CrossRef]
- Réus, G.Z.; Fernandes, G.C.; de Moura, A.B.; Silva, R.H.; Darabas, A.C.; de Souza, T.G.; Abelaira, H.M.; Carneiro, C.; Wendhausen, D.; Michels, M.; et al. Early Life Experience Contributes to the Developmental Programming of Depressive-Like Behaviour, Neuroinflammation and Oxidative Stress. J. Psychiatr. Res. 2017, 95, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Daskalakis, N.P.; Oitzl, M.S.; Schächinger, H.; Champagne, D.L.; de Kloet, E.R. Testing the Cumulative Stress and Mismatch Hypotheses of Psychopathology in a Rat Model of Early-Life Adversity. Physiol. Behav. 2012, 106, 707–721. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.E.; Chen, E. Harsh Family Climate in Early Life Presages the Emergence of a Proinflammatory Phenotype in Adolescence. Psychol. Sci. J. Am. Psychol. Soc. 2010, 21, 848–856. [Google Scholar] [CrossRef]
- Lin, J.E.; Neylan, T.C.; Epel, E.; O’Donovan, A. Associations of Childhood Adversity and Adulthood Trauma with C-Reactive Protein: A cross-Sectional Population-Based Study. Brain Behav. Immun. 2016, 53, 105–112. [Google Scholar] [CrossRef]
- Johnson, J.D.; Barnard, D.F.; Kulp, A.C.; Mehta, D.M. Neuroendocrine Regulation of Brain Cytokines after Psychological Stress. J. Endocr. Soc. 2019, 3, 1302–1320. [Google Scholar] [CrossRef]
- Blaisdell, K.N.; Imhof, A.M.; Fisher, P.A. Early Adversity, Child Neglect, and Stress Neurobiology: From Observations of Impact to Empirical Evaluations of Mechanisms. Int. J. Dev. Neurosci. 2019, 78, 139–146. [Google Scholar] [CrossRef]
- Ohta, K.I.; Miki, T.; Warita, K.; Suzuki, S.; Kusaka, T.; Yakura, T.; Liu, J.Q.; Tamai, M.; Takeuchi, Y. Prolonged Maternal Separation Disturbs the Serotonergic System during Early Brain Development. Int. J. Dev. Neurosci. 2014, 33, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Récamier-Carballo, S.; Estrada-Camarena, E.; López-Rubalcava, C. Maternal Separation Induces Long-Term Effects on Monoamines and Brain-Derived Neurotrophic Factor Levels on the Frontal Cortex, Amygdala, and Hippocampus: Differential Effects after a Stress Challenge. Behav. Pharmacol. 2017, 28, 545–557. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, S.E.; Quadros, I.M.H.; Machado, R.B.; Suchecki, D. Sex-Dependent Effects of Maternal Separation on Plasma Corticosterone and Brain Monoamines in Response to Chronic Ethanol Administration. Neuroscience 2013, 253, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Dayer, A. Serotonin-Related Pathways and Developmental Plasticity: Relevance for Psychiatric Disorders. Dialogues Clin. Neurosci. 2014, 16, 29–41. [Google Scholar]
- Picard, M.; McEwen, B.S.; Epel, E.S.; Sandi, C. An Energetic View of Stress: Focus on Mitochondria. Front. Neuroendocrinol. 2018, 49, 72–85. [Google Scholar] [CrossRef]
- Picard, M.; McEwen, B.S. Psychological Stress and Mitochondria. Psychosom. Med. 2018, 80, 126–140. [Google Scholar] [CrossRef]
- Picard, M.; Juster, R.P.; McEwen, B.S. Mitochondrial Allostatic Load Puts the “Gluc” back in Glucocorticoids. Nat. Rev. Endocrinol. 2014, 10, 303–310. [Google Scholar] [CrossRef]
- Manoli, I.; Alesci, S.; Blackman, M.R.; Su, Y.A.; Rennert, O.M.; Chrousos, G.P. Mitochondria as Key Components of the Stress Response. Trends Endocrinol. Metab. 2007, 18, 190–198. [Google Scholar] [CrossRef]
- Daniels, T.E.; Olsen, E.M.; Tyrka, A.R. Stress and Psychiatric Disorders: The Role of Mitochondria. Annu. Rev. Clin. Psychol. 2020, 16, 165–186. [Google Scholar] [CrossRef]
- Magistretti, P.J.; Allaman, I. A Cellular Perspective on Brain Energy Metabolism and Functional Imaging. Neuron 2015, 86, 883–901. [Google Scholar] [CrossRef] [PubMed]
- Meyer, T.; Wirtz, P.H. Mechanisms of Mitochondrial Redox Signaling in Psychosocial Stress-Responsive Systems: New Insights into an Old Story. Antioxid. Redox Signal. 2018, 28, 760–772. [Google Scholar] [CrossRef]
- McCoy, C.R.; Sabbagh, M.N.; Huaman, J.P.; Pickrell, A.M.; Clinton, S.M. Oxidative metabolism alterations in the emotional brain of anxiety-prone rats. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 95, 109706. [Google Scholar] [CrossRef] [PubMed]
- Harro, J.; Kanarik, M.; Matrov, D.; Panksepp, J. Mapping Patterns of Depression-Related Brain Regions with Cytochrome Oxidase Histochemistry: Relevance of Animal Affective Systems to Human Disorders, with a Focus on Resilience to Adverse events. Neurosci. Biobehav. Rev. 2011, 35, 1876–1889. [Google Scholar] [CrossRef]
- Kanarik, M.; Alttoa, A.; Matrov, D.; Kõiv, K.; Sharp, T.; Panksepp, J.; Harro, J. Brain Responses to Chronic Social Defeat Stress: Effects on Regional Oxidative Metabolism as a Function of a Hedonic Trait, and Gene Expression in Susceptible and Resilient Rats. Eur. Neuropsychopharmacol. 2011, 21, 92–107. [Google Scholar] [CrossRef] [PubMed]
- Ventura-Clapier, R.; Moulin, M.; Piquereau, J.; Lemaire, C.; Mericskay, M.; Veksler, V.; Garnier, A. Mitochondria: A Central Target for Sex Differences in Pathologies. Clin. Sci. 2017, 131, 803–822. [Google Scholar] [CrossRef]
- Torrens-Mas, M.; Pons, D.G.; Sastre-Serra, J.; Oliver, J.; Roca, P. Sexual Hormones Regulate the Redox Status and Mitochondrial Function in the Brain. Pathological implications. Redox Biol. 2020, 31, 101505. [Google Scholar] [CrossRef] [PubMed]
- Gaignard, P.; Liere, P.; Thérond, P.; Schumacher, M.; Slama, A.; Guennoun, R. Role of Sex Hormones on Brain Mitochondrial Function, with Special Reference to Aging and Neurodegenerative Diseases. Front. Aging Neurosci. 2017, 9, 406. [Google Scholar] [CrossRef]
- Bekhbat, M.; Neigh, G.N. Sex Differences in the Neuro-Immune Consequences of Stress: Focus on Depression and Anxiety. Brain Behav. Immun. 2018, 67, 1–12. [Google Scholar] [CrossRef]
- Barnard, D.F.; Gabella, K.M.; Kulp, A.C.; Parker, A.D.; Dugan, P.B.; Johnson, J.D. Sex Differences in the Regulation of Brain IL-1β in Response to Chronic Stress. Psychoneuroendocrinology 2019, 103, 203–211. [Google Scholar] [CrossRef]
- Spivey, J.M.; Colorado, R.A.; Conejo-Jimenez, N.; Gonzalez-Pardo, H.; Gonzalez-Lima, F. Juvenile Male Rats Display Lower Cortical Metabolic Capacity than Females. Neurosci. Lett. 2008, 440, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Spivey, J.M.; Padilla, E.; Shumake, J.D.; Gonzalez-Lima, F. Effects of Maternal Separation, Early Handling, and Gonadal Sex on Regional Metabolic Capacity of the Preweanling Rat Brain. Brain Res. 2011, 1367, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, C.F.; Odebode, G.; Naidu, L.; Koban, M. Early Life Stress Alters Adult Inflammatory Responses in a Mouse Model for Depression. Ann. Psychiatry Ment. Health 2017, 5, 1095. [Google Scholar] [PubMed]
- González-Pardo, H.; Arias, J.L.; Vallejo, G.; Conejo, N.M. Environmental Enrichment Effects after Early Stress on Behavior and Functional Brain Networks in Adult Rats. PLoS ONE 2019, 14, e0226377. [Google Scholar] [CrossRef]
- Banqueri, M.; Méndez, M.; Arias, J.L. Why Are Maternally Separated Females Inflexible? Brain Activity Pattern of COx and c-Fos. Neurobiol. Learn. Mem. 2018, 155, 30–41. [Google Scholar] [CrossRef]
- Vivinetto, A.L.; Suárez, M.M.; Rivarola, M.A. Neurobiological Effects of Neonatal Maternal Separation and Post-Weaning Environmental Enrichment. Behav. Brain Res. 2013, 240, 110–118. [Google Scholar] [CrossRef]
- Dandi, Ε.; Kalamari, A.; Touloumi, O.; Lagoudaki, R.; Nousiopoulou, E.; Simeonidou, C.; Spandou, E.; Tata, D.A. Beneficial Effects of Environmental Enrichment on Behavior, Stress Reactivity and Synaptophysin/BDNF Expression in Hippocampus Following Early Life Stress. Int. J. Dev. Neurosci. 2018, 67, 19–32. [Google Scholar] [CrossRef]
- Li, M.; Xue, X.; Shao, S.; Shao, F.; Wang, W. Cognitive, Emotional and Neurochemical Effects of Repeated Maternal Separation in Adolescent Rats. Brain Res. 2013, 1518, 82–90. [Google Scholar] [CrossRef]
- González-Pardo, H.; Arias, J.L.; Vallejo, G.; Conejo, N.M. Influence of Environmental Enrichment on the Volume of Brain Regions Sensitive to Early Life Stress by Maternal Separation in Rats. Psicothema 2019, 31, 46–52. [Google Scholar] [CrossRef]
- Wong-Riley, M.T.T. Cytochrome Oxidase: An Endogenous Metabolic Marker for Neuronal Activity. Trends Neurosci. 1989, 12, 94–101. [Google Scholar] [CrossRef]
- Spivey, J.M.; Shumake, J.; Colorado, R.A.; Conejo-Jimenez, N.; Gonzalez-Pardo, H.; Gonzalez-Lima, F. Adolescent Female Rats Are More Resistant than Males to the Effects of Early Stress on Prefrontal Cortex and Impulsive Behavior. Dev. Psychobiol. 2009, 51, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Lima, F.; Cada, A. Cytochrome Oxidase Activity in the Auditory System of the Mouse: A Qualitative and Quantitative Histochemical Study. Neuroscience 1994, 63, 559–578. [Google Scholar] [CrossRef]
- Shumake, J.; Conejo-Jimenez, N.; Gonzalez-Pardo, H.; Gonzalez-Lima, F. Brain Differences in Newborn Rats Predisposed to Helpless and Depressive Behavior. Brain Res. 2004, 1030, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates: Hard Cover Edition; Elsevier Science: New York, NY, USA, 2013; ISBN 9780124157521. [Google Scholar]
- Spijker, S. Dissection of Rodent Brain Regions. Neuromethods 2011, 57, 13–26. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Harro, J.; Kanarik, M.; Kaart, T.; Matrov, D.; Kõiv, K.; Mällo, T.; Del Río, J.; Tordera, R.M.; Ramirez, M.J. Revealing the Cerebral Regions and Networks Mediating Vulnerability to Depression: Oxidative Metabolism Mapping of Rat Brain. Behav. Brain Res. 2014, 267, 83–94. [Google Scholar] [CrossRef]
- Hollis, F.; Van Der Kooij, M.A.; Zanoletti, O.; Lozano, L.; Cantó, C.; Sandi, C. Mitochondrial Function in the Brain Links Anxiety with Social Subordination. Proc. Natl. Acad. Sci. USA 2015, 112, 15486–15491. [Google Scholar] [CrossRef]
- Filiou, M.D.; Sandi, C. Anxiety and Brain Mitochondria: A Bidirectional Crosstalk. Trends Neurosci. 2019, 42, 573–588. [Google Scholar] [CrossRef]
- Mällo, T.; Matrov, D.; Kõiv, K.; Harro, J. Effect of Chronic Stress on Behavior and Cerebral Oxidative Metabolism in Rats with High or Low Positive Affect. Neuroscience 2009, 164, 963–974. [Google Scholar] [CrossRef]
- Shumake, J.; Poremba, A.; Edwards, E.; Gonzalez-Lima, F. Congenital Helpless Rats as a Genetic Model for Cortex Metabolism in Depression. Neuroreport 2000, 11, 3793–3798. [Google Scholar] [CrossRef]
- Holper, L.; Lan, M.J.; Brown, P.J.; Sublette, E.M.; Burke, A.; Mann, J.J. Brain Cytochrome-C-Oxidase as a Marker of Mitochondrial Function: A Pilot Study in Major Depression Using NIRS. Depress. Anxiety 2019, 36, 766–779. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Huang, S.; Cao, J.; Chen, T.; Zhu, P.; Zhu, R.; Su, P.; Ruan, D. The Timing of Maternal Separation Affects Morris Water Maze Performance and Long-Term Potentiation in Male Rats. Dev. Psychobiol. 2014, 56, 1102–1109. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Cao, K.; Lin, H.; Cui, S.; Shen, C.; Wen, W.; Mo, H.; Dong, Z.; Bai, S.; Yang, L.; et al. Early-Life Stress Induces Depression-Like Behavior and Synaptic-Plasticity Changes in a Maternal Separation Rat Model: Gender Difference and Metabolomics Study. Front. Pharmacol. 2020, 11, 102. [Google Scholar] [CrossRef] [PubMed]
- Maghami, S.; Zardooz, H.; Khodagholi, F.; Binayi, F.; Saber, R.R.; Hedayati, M.; Sahraei, H.; Ansar, M.A. Maternal Separation Blunted Spatial Memory Formation Independent of Peripheral and Hippocampal Insulin Content in Young Adult Male Rats. PLoS ONE 2018, 13, e0204731. [Google Scholar] [CrossRef]
- Hoffmann, A.; Spengler, D. The Mitochondrion as Potential Interface in Early-Life Stress Brain Programming. Front. Behav. Neurosci. 2018, 12, 306. [Google Scholar] [CrossRef]
- Hamon, M.; Blier, P. Monoamine Neurocircuitry in Depression and Strategies for New Treatments. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2013, 45, 54–63. [Google Scholar] [CrossRef]
- Altemus, M.; Sarvaiya, N.; Neill Epperson, C. Sex Differences in Anxiety and Depression Clinical Perspectives. Front. Neuroendocrinol. 2014, 35, 320–330. [Google Scholar] [CrossRef]
- Arborelius, L.; Owens, M.J.; Plotsky, P.M.; Nemeroff, C.B. The Role of Corticotropin-Releasing Factor in Depression and Anxiety Disorders. J. Endocrinol. 1999, 160, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Arborelius, L.; Eklund, M.B. Both Long and Brief Maternal Separation Produces Persistent Changes in Tissue Levels of Brain Monoamines in Middle-Aged Female Rats. Neuroscience 2007, 145, 738–750. [Google Scholar] [CrossRef] [PubMed]
- Leonard, B.E.; Myint, A. The psychoneuroimmunology of depression. Hum. Psychopharmacol. 2009, 24, 165–175. [Google Scholar] [CrossRef] [PubMed]
- van Bodegom, M.; Homberg, J.R.; Henckens, M.J.A.G. Modulation of the Hypothalamic-Pituitary-Adrenal Axis by Early Life Stress Exposure. Front. Cell. Neurosci. 2017, 11, 87. [Google Scholar] [CrossRef]
- Handa, R.J.; Weiser, M.J. Gonadal Steroid Hormones and the Hypothalamo-Pituitary-Adrenal Axis. Front. Neuroendocrinol. 2014, 35, 197–220. [Google Scholar] [CrossRef]
- Panagiotakopoulos, L.; Neigh, G.N. Development of the HPA axis: Where and When Do Sex Differences Manifest? Front. Neuroendocrinol. 2014, 35, 285–302. [Google Scholar] [CrossRef]
- Heck, A.L.; Handa, R.J. Sex Differences in the Hypothalamic–Pituitary–Adrenal Axis’ Response to Stress: An Important Role for Gonadal Hormones. Neuropsychopharmacology 2019, 44, 45–58. [Google Scholar] [CrossRef] [PubMed]
- Llorente, R.; O’Shea, E.; Gutierrez-Lopez, M.D.; Llorente-Berzal, A.; Colado, M.I.; Viveros, M.P. Sex-dependent maternal deprivation effects on brain monoamine content in adolescent rats. Neurosci. Lett. 2010, 479, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Bowman, R.E.; Beck, K.D.; Luine, V.N. Chronic Stress Effects on Memory: Sex Differences in Performance and Monoaminergic Activity. Horm. Behav. 2003, 43, 48–59. [Google Scholar] [CrossRef]
- Matthews, K.; Dalley, J.W.; Matthews, C.; Tsai, T.H.; Robbins, T.W. Periodic Maternal Separation of Neonatal Rats Produces Region- and Gender-Specific Effects on Biogenic Amine Content in Postmortem Adult Brain. Synapse 2001, 40, 1–10. [Google Scholar] [CrossRef]
- Daniels, W.M.U.; Pietersen, C.Y.; Carstens, M.E.; Stein, D.J. Maternal Separation in Rats Leads to Anxiety-Like Behavior and a Blunted ACTH Response and Altered Neurotransmitter Levels in Response to a Subsequent Stressor. Metab. Brain Dis. 2004, 19, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.; Courtiol, E.; Castellanos, F.X.; Teixeira, C.M. Abnormal Serotonin Levels during Perinatal Development Lead to Behavioral Deficits in Adulthood. Front. Behav. Neurosci. 2018, 12, 114. [Google Scholar] [CrossRef] [PubMed]
- Sachs, B.D.; Rodriguiz, R.M.; Siesser, W.B.; Kenan, A.; Royer, E.L.; Jacobsen, J.P.R.; Wetsel, W.C.; Caron, M.G. The Effects of Brain Serotonin Deficiency on Behavioural Disinhibition and Anxiety-Like Behaviour Following Mild Early Life Stress. Int. J. Neuropsychopharmacol. 2013, 16, 2081–2094. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Dong, X.; Wang, Y.; Liu, M.; Sun, A.; Li, N.; Lin, Y.; Geng, Z.; Jin, Y.; Li, X. Adolescent Escitalopram Prevents the Effects of Maternal Separation on Depression- and Anxiety-Like Behaviours and Regulates the Levels of Inflammatory Cytokines in Adult Male Mice. Int. J. Dev. Neurosci. 2017, 62, 37–45. [Google Scholar] [CrossRef]
- Honeycutt, J.A.; Demaestri, C.; Peterzell, S.; Silveri, M.M.; Cai, X.; Kulkarni, P.; Cunningham, M.G.; Ferris, C.F.; Brenhouse, H.C. Altered Corticolimbic Connectivity Reveals Sex-Specific Adolescent Outcomes in a Rat Model of Early Life Adversity. eLife 2020, 9, e52651. [Google Scholar] [CrossRef] [PubMed]
- Roque, A.; Ochoa-Zarzosa, A.; Torner, L. Maternal Separation Activates Microglial Cells and Induces an Inflammatory Response in the Hippocampus of Male Rat Pups, Independently of Hypothalamic and Peripheral Cytokine Levels. Brain Behav. Immun. 2016, 55, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Bekhbat, M.; Howell, P.A.; Rowson, S.A.; Kelly, S.D.; Tansey, M.G.; Neigh, G.N. Chronic Adolescent Stress Sex-Specifically Alters Central and Peripheral Neuro-Immune Reactivity in Rats. Brain Behav. Immun. 2019, 76, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Leonard, B.E. HPA and immune axes in stress: Involvement of the Serotonergic System. Neuroimmunomodulation 2007, 13, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Bollinger, J.L.; Bergeon Burns, C.M.; Wellman, C.L. Differential Effects of Stress on Microglial Cell Activation in Male and Female Medial Prefrontal Cortex. Brain Behav. Immun. 2016, 52, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Pyter, L.M.; Kelly, S.D.; Harrell, C.S.; Neigh, G.N. Sex Differences in the Effects of Adolescent Stress on Adult Brain Inflammatory Markers in Rats. Brain Behav. Immun. 2013, 30, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Bollinger, J.L.; Collins, K.E.; Patel, R.; Wellman, C.L. Behavioral Stress Alters Corticolimbic Microglia in a Sex- and Brain Region-Specific Manner. PLoS ONE 2017, 12, 0187631. [Google Scholar] [CrossRef]
- Członkowska, A.; Ciesielska, A.; Gromadzka, G.; Kurkowska-Jastrzebska, I. Gender Differences in Neurological Disease: Role of Estrogens and Cytokines. Endocrine 2006, 29, 243–256. [Google Scholar] [CrossRef]
- Sukoff Rizzo, S.J.; Neal, S.J.; Hughes, Z.A.; Beyna, M.; Rosenzweig-Lipson, S.; Moss, S.J.; Brandon, N.J. Evidence for Sustained Elevation of IL-6 in the CNS as a Key Contributor of Depressive-Like Phenotypes. Transl. Psychiatry 2012, 2, e199. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, X.; Zhou, Y.; Zheng, Q.; Chen, Z.; Zhang, H.; Sun, Z.; Xu, G.; Hu, G. Hypothalamus-Pituitary-Adrenal Axis Imbalance and Inflammation Contribute to Sex Differences in Separation- and Restraint-Induced Depression. Horm. Behav. 2020, 122, 104741. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Group | Maternal Separation Group | Effects (P=) | |||||
|---|---|---|---|---|---|---|---|
| Male | Female | Male | Female | Sex | Separation | Sex × Rearing | |
| Cingulate cortex (Cgl) | 31.3 ± 1.1 | 41.7 ±1.0 | 26.7 ± 0.9 | 29.7 ± 0.9 | <0.001 | <0.001 | <0.001 |
| Prelimbic cortex (PrL) | 32.1 ± 0.9 | 40.3 ± 0.8 | 27.2 ± 0.9 | 28.9 ± 0.7 | <0.001 | <0.001 | 0.001 |
| Infralimbic cortex (IL) | 30.6 ± 1.9 | 43.1 ± 0.6 | 25.8 ± 1.8 | 28.4 ± 0.5 | <0.001 | <0.001 | <0.002 |
| Doral striatum (dCPu) | 27.3 ± 0.6 | 30.2 ± 0.4 | 29.3 ± 1.7 | 32.5 ± 0.6 | <0.007 | 0.055 | 0.902 |
| Accumbens nucleus, core (AcbC) | 33.5 ± 1.4 | 42.4 ± 0.5 | 33.9 ± 1.3 | 43.7 ± 0.7 | <0.001 | 0.430 | 0.689 |
| Accumbens nucleaus, shell (AcbSh) | 38.8 ± 0.5 | 52.1 ± 0.6 | 36.7 ± 0.9 | 45.1 ± 09 | <0.001 | <0.091 | <0.007 |
| Dorsal field CA1 of the hippocampus (dCA1) | 22.1 ± 0.4 | 26.7 ± 0.7 | 22.2 ± 0.7 | 24.5 ± 0.2 | <0.001 | <0.091 | 0.076 |
| Dorsal field CA3 of the hippocampus (dCA3) | 24.7 ± 0.9 | 29. 4 ± 0.7 | 21.7 ± 09 | 22.8 ± 0.8 | 0.003 | <0.001 | 0.504 |
| Dorsal dentate gyrus (dDG) | 32.4 ± 0.5 | 40.9 ± 1.3 | 27.0± 0.9 | 27.3 ±1.1 | <0.001 | <0.001 | <0.001 |
| Ventral field CA1of the hippocampus (vCA1) | 23.7 ± 0.2 | 31.6 ± 1.0 | 22.2 ± 0.6 | 32.2± 0.6 | <0.001 | 0.488 | 0.142 |
| Ventral field CA3 of the hippocampus (vCA3) | 22.8 ± 0.9 | 33.5 ± 1.1 | 20.2 ± 0.8 | 40.7 ± 1.4 | <0.001 | 0.998 | 0.101 |
| Ventral dentare gyrus (vDG) | 24.05 ± 0.4 | 30.9 ± 1.2 | 21.6 ± 0.8 | 30.1 ± 1. | <0.001 | 0.054 | 0.247 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Pardo, H.; Arias, J.L.; Gómez-Lázaro, E.; López Taboada, I.; Conejo, N.M. Sex-Specific Effects of Early Life Stress on Brain Mitochondrial Function, Monoamine Levels and Neuroinflammation. Brain Sci. 2020, 10, 447. https://doi.org/10.3390/brainsci10070447
González-Pardo H, Arias JL, Gómez-Lázaro E, López Taboada I, Conejo NM. Sex-Specific Effects of Early Life Stress on Brain Mitochondrial Function, Monoamine Levels and Neuroinflammation. Brain Sciences. 2020; 10(7):447. https://doi.org/10.3390/brainsci10070447
Chicago/Turabian StyleGonzález-Pardo, Héctor, Jorge L. Arias, Eneritz Gómez-Lázaro, Isabel López Taboada, and Nélida M. Conejo. 2020. "Sex-Specific Effects of Early Life Stress on Brain Mitochondrial Function, Monoamine Levels and Neuroinflammation" Brain Sciences 10, no. 7: 447. https://doi.org/10.3390/brainsci10070447
APA StyleGonzález-Pardo, H., Arias, J. L., Gómez-Lázaro, E., López Taboada, I., & Conejo, N. M. (2020). Sex-Specific Effects of Early Life Stress on Brain Mitochondrial Function, Monoamine Levels and Neuroinflammation. Brain Sciences, 10(7), 447. https://doi.org/10.3390/brainsci10070447