Abstract
Non-tuberculous mycobacteria (NTM) represent a diverse group of mycobacterial species known for causing opportunistic infections, especially in individuals with underlying health conditions. Unlike Mycobacterium tuberculosis (Mtb), the causative agent of tuberculosis, NTM species exhibit different pathogenic characteristics and drug resistance mechanisms, making them increasingly relevant in clinical settings. PtkA is a crucial protein tyrosine kinase that regulates bacterial growth, stress response, and virulence by phosphorylating various substrates in Mtb. Understanding whether PtkA homologs exist in NTM could provide insights into their virulence and resistance mechanisms. In silico approaches, which utilize computational tools for sequence alignment, structure prediction, and functional annotation, offer a powerful means to identify homologous proteins across different species. In this article, we have employed tools like BLAST (Basic Local Alignment Search Tool), protein structure databases, and the NTM database to identify PtkA homologs in NTM genomes, providing a foundation for further studies.
1. Introduction
Based on sequence homology, PtkA belongs to the haloacid dehalogenase (HAD) superfamily, which includes various phosphotransferases, phosphatases, ATPases, and phosphoglucomutases, in addition to dehalogenases [1]. Biochemical analysis, however, confirms that PtkA functions as a bona fide protein tyrosine kinase [2], making it the first known kinase of its kind within the HAD family. Tyrosine phosphorylation has been observed in pathogenic mycobacteria but not in non-pathogenic strains, indicating a potential role in virulence [3]. Phosphorylation at Ser/Thr/Tyr residue plays a crucial role in various bacterial cellular processes, including virulence and survival, as demonstrated in species like E. coli, H. pylori, B. subtilis, S. coelicolor, M. pneumoniae, S. pneumoniae, K. pneumoniae, L. lactis, Campylobacter jejuni, and Pseudomonas species [4]. The mycobacterial cell wall, which is unique in composition, is essential for its pathogenicity. While tyrosine phosphorylation is vital for maintaining cell wall integrity in other bacteria, it remains understudied in mycobacteria [5,6,7].
In Streptococcus pneumoniae, capsular polysaccharide (CPS) production, critical for virulence, is regulated by the tyrosine phosphorylation of the CpsD protein, which is mediated by the CPS biosynthesis proteins CpsB, CpsC, and CpsD [8]. CpsD is an autophosphorylating protein-tyrosine kinase, and CpsC is required for its tyrosine phosphorylation [9]. Similarly, the Caulobacter crescentus divL gene encodes a unique bacterial tyrosine kinase, which is essential for cell viability and division [10]. PtkA is predicted to localize in the mycobacterial cell wall or outer membrane [11]. ptkA-deficient mycobacteria show the downregulation of Wag31 (identical to Ag 84; Rv2145c) and Ag 85c (Rv0129c) [12,13]. Wag31 of Mycobacterium tuberculosis belongs to the DivIVA family of proteins known to regulate cell morphology in Gram-positive bacteria. It is phosphorylated by the serine/threonine kinase PknA and regulates polar cell wall synthesis [14]. Ag85c is an essential component for mycobacterial cell walls [12]. This suggests that tyrosine phosphorylation by PtkA plays an important role in maintaining cell integrity in mycobacterial species.
2. Results and Discussion
2.1. Phylogenetic Analysis of Tyrosine Kinase (PtkA) Within the MTB Complex
A phylogenetic tree was constructed to determine the evolutionary relationship of the identified homologs with the known PtkA. The clustering of NTM homologs with the PtkA of M. tuberculosis highlights that these kinases likely evolved from a common ancestor. The conservation of crucial functional motifs in the NTM homologs suggests that they might play a similar role in regulating cellular functions, which could influence their pathogenicity. PtkA full-length protein is present in M. tuberculosis, M. bovis, M. marinum, and M. smegmatis. The dendrogram (Figure 1A) shows the phylogenesis of tyrosine kinase present within the MTB complex, including M. africanum, M. microti, M. bovis, M. tuberculosis, M. marinum, M. smegmatis, and M. leprae. The PTKs (Rv2232 and MS4308 and putative tyrosine kinase MS5955) and PTPs in pathogenic (Rv2234, Rv0153c) and non-pathogenic (MS4309 and MS0100) mycobacteria were aligned, and their phylogenesis is shown in (Figure 1B).
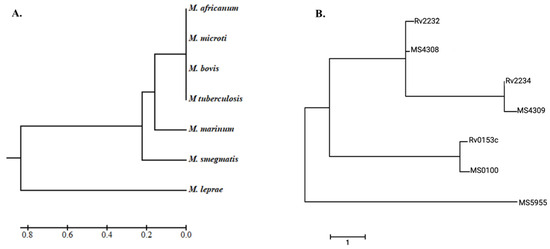
Figure 1.
Phylogenetic analysis. (A) Evolutionary analysis of Mtb species and (B) comparison of kinase domain proteins to NTM proteins. These images were generated using MEGA 7.1 software.
2.2. Homology Modeling of Protein Tyrosine Kinases (Rv2232, MS4308 and MS5955)
As reported earlier, PtpA is the substrate of PtkA [2]. Based on PtkA-PtpA operon conservation theory, we have targeted the PtpA orthologs present in M. smegmatis (MS) as the substrate of MS tyrosine kinase. The alpha fold (AF) models, Rv2232 (pTM = 0.72) and MS5955 (pTM = 0.70), have an averagely low prediction quality score, while MS0100, MS4308, and MS4309 have a good prediction quality score that indicates accurately predicted structures through the AF algorithm. The 3D structures of Rv2232 (PtkA) [AF Model], Rv2234 (PtpA) [PDB ID: 1U2P], and Rv0153c (PtpB) [PDB ID: 1YWF] identified a similar template for MS4309 and MS0100 for protein–protein interaction (Figure 2). Based on the bioinformatic prediction, we have analyzed the interaction of mycobacterial tyrosine kinases with mycobacterial tyrosine phosphatase (Table 1 and Table 2).
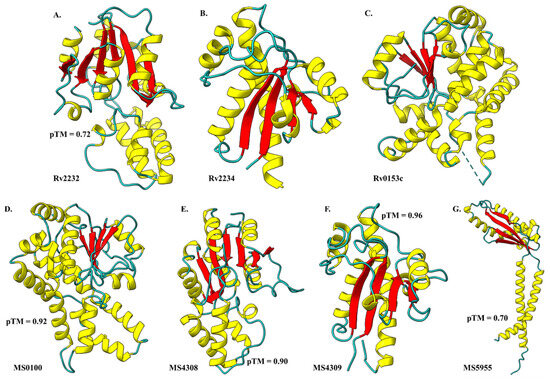
Figure 2.
3D structures of Mtb and NTM species having similar kinase domains. (A) Rv2232 model, crystallographic structures; (B) Rv2234 and (C) Rv0153c, Alpha fold predicted models; (D) MS0100, (E) MS4308, (F) MS4309, and (G) MS5955. Secondary structures are represented as helices in yellow, sheets in red, and coils/loops in Cyan using ChimeraX.

Table 1.
In silico studies for protein–protein interactions.
Table 2.
Bioinformatic prediction of mycobacterial tyrosine kinases interaction with mycobacterial tyrosine phosphatase. InterProScan online server used for predicting interaction probabilities.
2.3. Evolution of Tyrosine Kinase Within Bacterial Kingdom
We retrieved protein sequences of protein tyrosine kinase from different organisms mentioned by Joseph D. Chao et al., 2014 [4] from the UniProt database (http://www.uniprot.org). All collected protein sequences were aligned by the multiple sequence alignment method using ClustalW available in MEGA v11. Consensus sequences were predicted in Rv2232 by aligned sequences with MS4308 and MS5955 kinase protein sequences using the ENDscript/ESPript online web server (http://espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi accessed on 1 August 2024).
2.4. Divergence of PtkA from Bacterial Tyrosine (BY) Kinase
Prokaryotes have been found to possess a diverse array of protein kinases, including histidine and aspartic acid kinases, serine/threonine kinases, and more recently, tyrosine and arginine kinases [4,15]. Interestingly, bacterial tyrosine kinases have evolved independently, bearing no structural resemblance to their eukaryotic counterparts. These unique bacterial kinases utilize the ATP/GTP-binding Walker motif to catalyze both autophosphorylation and substrate phosphorylation on tyrosine residues [15,16]. Tyrosine phosphorylation has emerged as a critical regulatory mechanism in bacterial physiology, influencing processes such as exopolysaccharide production, virulence, stress response, and DNA metabolism [17,18,19]. Phylogenetic analysis of the kinase domains of known protein-tyrosine kinases (PPTKs) shows their clustering into three distinct families. The catalytic domain of BY-kinases contains variants of the Walker A (P-loop) and Walker B motifs, which are essential for nucleotide binding and hydrolysis, respectively. In addition, a unique motif, termed Walker A’, is located between the Walker A and Walker B motifs. This Walker A’ motif is also present in certain P-loop nucleotide kinases and other ATPases [15,16].
2.5. PtkA-PtpA Operon
The PtkA-PtpA operon is conserved across various species within the Actinomycetes order, including genera such as Rhodococcus, Corynebacterium, Gordonia, Amycolicicoccus, and Mycobacterium, although protein homology for PtkA ranges from 39% to 76% identity [4]. Given the conservation of this operon, it is suggested that PtkA homologs function as protein tyrosine kinases. Within the Mycobacterium tuberculosis (Mtb) complex, this operon is highly conserved, displaying 100% protein identity, with PtpA playing a known role in virulence [2]. A search in the KEGG Sequence Similarity Database identified 1772 orthologs of PtkA, with a minimum Smith-Waterman (SW) score of 150, using the best alignment FastA algorithm. The PtpA operon, which encodes PtkA, was also conserved in 35 of the 123 Actinomycetes species [4].
2.6. Functional Prediction and Implications
The identification of these homologs paves the way for further experimental studies. PtkA is known to phosphorylate several mycobacterial proteins [2,12], as well as host proteins during infection, playing key roles in maintaining bacterial physiology and virulence [20]. However, PtkA remains relatively underexplored, and much more needs to be uncovered about its functions. Tyrosine kinases in non-tuberculous mycobacteria (NTMs) may be involved in processes analogous to those observed in M. tuberculosis, such as modulating host immune responses, regulating stress responses within the intracellular environment, and potentially contributing to mechanisms of drug resistance.
3. Methods
3.1. Sequence Homology Search
The protein sequence of ptkA from Mycobacterium tuberculosis was retrieved from the Mycobrowser database (https://mycobrowser.epfl.ch/, accessed on 1 December 2023) using the sequence ID: Rv2232. This sequence was then used as a query to identify homologs in non-tuberculous mycobacteria (NTM) species. The Basic Local Alignment Search Tool (BLAST) [21], available through the National Center for Biotechnology Information (NCBI) (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 1 March 2024), was employed to search for homologous sequences within the RCSB Protein Data Bank (https://www.rcsb.org/, accessed on 1 March 2024). Homologous sequences were selected based on their sequence similarity and identity. Additionally, the ptkA sequence was compared against the NTM genome database (https://ngdc.cncb.ac.cn/ntmdb/, accessed on 1 March 2024) to further identify homologous sequences based on sequence similarity. A high-scoring match indicates the presence of a potential homolog.
3.2. Phylogenetic Analysis
The identified sequences were subsequently subjected to phylogenetic analysis to explore evolutionary relationships between PtkA in Mtb and its homologs across various NTM species. Phylogenetic analysis was performed using the Molecular Evolutionary Genetics Analysis (MEGA v11) software [22]. The PtkA and homologous protein sequences were aligned using ClustalW [23]. Following alignment, a phylogenetic tree was constructed using the Maximum Likelihood method, with the Kimura 2-parameter in the substitution model followed by 1000 bootstrap replicates set to provide statistical confidence for the tree’s nodes. The Newick output file is used for further visual analysis.
3.3. Functional Domain Conservation
Conserved functional domains, such as kinase domains, were analyzed using tools like InterProScan and Pfam to confirm that the identified proteins share structural and functional similarities with Mtb’s PtkA. Conserved functional domains, such as kinase domains, were analyzed using domain prediction tools such as SMART (https://smart.embl-heidelberg.de/, accessed on 25 June 2024) and InterPro (https://www.ebi.ac.uk/interpro/search/sequence/, accessed on 25 June 2024), which detect conserved protein domains based on hidden Markov models (HMMs) methods. The aligned sequences from ClustalW were used as input for predicting the domains.
3.4. Homology Modeling
The complete structure of Mtb’s PtkA is not available in the protein databank, except for partial structures. Therefore, the prediction method was applied using AlphaFold v3 (https://alphafoldserver.com, accessed on 1 September 2024). All identified NTM homologous proteins were also remodeled using AlphaFold v3. The modeled structures were visualized using ChimeraX v1.8 [24].
4. Conclusions
The in-silico identification of PtkA homologs in NTMs provides foundational knowledge for exploring their biological significance. Understanding the function of these kinases can aid in designing novel therapeutic strategies targeting NTMs, which are increasingly recognized for causing pulmonary diseases, especially in immunocompromised patients. Further experimental validation is needed to confirm the biological role of these homologs in NTMs and their contribution to virulence and survival in the host.
Author Contributions
Conceptualization, S.J.; writing, reviewing, and editing, S.J. and S.K.; methodology, software, and data curation, S.J. and S.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available.
Acknowledgments
We want to acknowledge the UMass Chan Library and City College of New York Library for providing access to the necessary resources.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Burroughs, A.M.; Allen, K.N.; Dunaway-Mariano, D.; Aravind, L. Evolutionary Genomics of the HAD Superfamily: Understanding the Structural Adaptations and Catalytic Diversity in a Superfamily of Phosphoesterases and Allied Enzymes. J. Mol. Biol. 2006, 361, 1003–1034. [Google Scholar] [CrossRef] [PubMed]
- Bach, H.; Wong, D.; Av-Gay, Y. Mycobacterium tuberculosis PtkA is a novel protein tyrosine kinase whose substrate is PtpA. Biochem J. 2009, 420, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Diseases, I.; Columbia, B. Protein tyrosine phosphorylation in Mycobacterium tuberculosis. FEMS Microbiol. Lett. 1994, 124, 203–207. [Google Scholar]
- Chao, J.D.; Wong, D.; Av-Gay, Y. Microbial protein-tyrosine kinases. J. Biol. Chem. 2014, 289, 9463–9472. [Google Scholar] [CrossRef] [PubMed]
- Alderwick, L.J.; Harrison, J.; Lloyd, G.S.; Birch, H.L. The Mycobacterial Cell Wall Peptidoglycan and Arabinogalactan. Cold Spring Harb. Perspect. Med. 2015, 5, a021113. [Google Scholar] [CrossRef] [PubMed]
- Maitra, A.; Munshi, T.; Healy, J.; Martin, L.T.; Vollmer, W.; Keep, N.H.; Bhakta, S. Cell wall peptidoglycan in Mycobacterium tuberculosis: An Achilles heel for the TB-causing pathogen. FEMS Microbiol. Rev. 2019, 43, 548–575. [Google Scholar] [CrossRef] [PubMed]
- Vilchèze, C. Mycobacterial Cell Wall: A Source of Successful Targets for Old and New Drugs. Appl. Sci. 2020, 10, 2278. [Google Scholar] [CrossRef]
- Morona, J.K.; Morona, R.; Paton, J.C. Attachment of capsular polysaccharide to the cell wall of Streptococcus pneumoniae type 2 is required for invasive disease. Proc. Natl. Acad. Sci. USA 2006, 103, 8505–8510. [Google Scholar] [CrossRef]
- Morona, J.K.; Paton, J.C.; Miller, D.C.; Morona, R. Tyrosine phosphorylation of CpsD negatively regulates capsular polysaccharide biosynthesis in Streptococcus pneumoniae. Mol. Microbiol. 2000, 35, 1431–1442. [Google Scholar] [CrossRef]
- Wu, J.; Ohta, N.; Zhao, J.-L.; Newton, A. A novel bacterial tyrosine kinase essential for cell division and differentiation. Proc. Natl. Acad. Sci. USA 1999, 96, 13068–13073. [Google Scholar] [CrossRef]
- Song, H.; Sandie, R.; Wang, Y.; Andrade-Navarro, M.A.; Niederweis, M. Identification of outer membrane proteins of Mycobacterium tuberculosis. Tuberculosis Edinb. 2008, 88, 526–544. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, S.; Kumar, S.; De la Cruz, E.V. Exploring the role of the protein tyrosine kinase a (PtkA) in mycobacterial intracellular survival. Tuberculosis 2023, 142, 102398. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, S.; Srivasatava, K.K. A brief report on the mycobacterial proteins regulated by the loss of PtkA in Mtb-H37Ra. BioRxiv 2023, 09.10.556525. [Google Scholar] [CrossRef]
- Nguyen, L.; Scherr, N.; Gatfield, J.; Walburger, A.; Pieters, J.; Thompson, C.J. Antigen 84, an Effector of Pleiomorphism inMycobacterium smegmatis. J. Bacteriol. 2007, 189, 7896–7910. [Google Scholar] [CrossRef]
- Grangeasse, C.; Nessler, S.; Mijakovic, I. Bacterial tyrosine kinases: Evolution, biological function and structural insights. Philos. Trans. R. Soc. B Biol. Sci. 2012, 367, 2640–2655. [Google Scholar] [CrossRef]
- Hajredini, F.; Alphonse, S.; Ghose, R. BY-kinases: Protein tyrosine kinases like no other. J. Biol. Chem. 2023, 299, 102737. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Whitmore, S.E.; Lamont, R.J. Tyrosine phosphorylation and bacterial virulence. Int. J. Oral. Sci. 2012, 4, 1–6. [Google Scholar] [CrossRef][Green Version]
- Getz, L.J.; Runte, C.S.; Rainey, J.K.; Thomas, N.A. Tyrosine Phosphorylation as a Widespread Regulatory Mechanism in Prokaryotes. J. Bacteriol. 2019, 201, e00205-19. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Grangeasse, C.; Terreux, R.; Nessler, S. Bacterial tyrosine-kinases: Structure-function analysis and therapeutic potential. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2010, 1804, 628–634. [Google Scholar] [CrossRef]
- Jaiswal, S.; Srivastava, K.K. Protein tyrosine kinase A modulates intracellular survival of mycobacteria through Galectin 3. Biochem. Biophys. Res. Commun. 2018, 498, 884–890. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Protein Sci. A Publ. Protein So Ciety 2021, 30, 70–82. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).