
Role of Transient Receptor Potential Vanilloid Channels in Gastrointestinal Physiology and Pathology

 , , , , ,
, , , , ,
Abstract
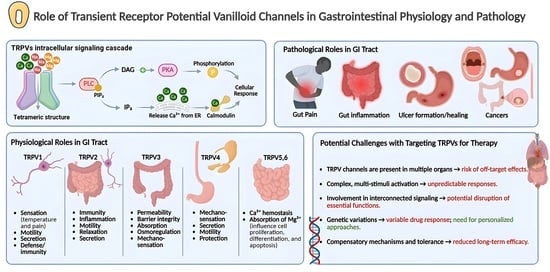
1. Introduction
2. Methods
3. The General Structure of TRPV Channels
4. TRPV Channels in GI Physiology
4.1. TRPV1
4.2. TRPV2
4.3. TRPV3
4.4. TRPV4
4.5. TRPV5 and TRPV6

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Channel | Key Activators | Localization and Expression | Physiological Functions in the GI Tract |
|---|---|---|---|
| TRPV1 | Activated by temperatures above 42 °C, hyperosmolarity, low pH, capsaicin, and endovanilloids such as anandamide [21,22]. Moreover, it is activated by the endogenous lipid mediator oleoylethanolamide (OLDA) [38]. | Primarily localized in sensory neurons of the peripheral nervous system, dorsal root ganglia, and trigeminal ganglia [9], as well as in enteric neurons within the gastrointestinal tract (GIT) [24]. | TRPV1 senses temperature and pain, mediating responses to noxious heat and chemical irritants [9]. It contributes to visceral pain perception [25] and regulates GI motility and secretion [25]. It modulates vagally mediated contractions of the striated part of the esophagus, regulates gastric acid secretion and gastric emptying [25], and is implicated in gastric mucosal defense by promoting mucus and bicarbonate release [27,28]. Additionally, it modulates immune responses in the stomach and small intestines [27,28]. |
| TRPV2 | Activated by high temperatures (above 50–52 °C), mechanical stimuli, and endogenous lipids such as 2-arachidonoylglycerol (2-AG) and lysophosphatidylcholine (LPC) [29,30,31]. | Exhibits widespread expression across various tissues [33], including enteric immune cells such as macrophages, dendritic cells, and T cells [34], and has been identified in enteric neurons in rodents [35,36]. | TRPV2 contributes to diverse cellular processes, including immune responses and inflammation [33], and regulates inflammatory responses while enhancing mucosal immunity within the GIT [34]. Its stimulation in enteric neurons influences downstream neurotransmitter release [35,36] and mediates nitric oxide (NO) release, a key inhibitory neurotransmitter that regulates intestinal motility by inducing smooth muscle relaxation [35,36]. |
| TRPV3 | Activated by temperatures exceeding 30–33 °C, natural compounds like camphor and 2-aminoethoxydiphenyl borate (2-APB), and endogenous lipids such as 12-hydroxyeicosatetraenoic acid (12-HETE) [37]. Additionally, Cannabidiol (CBD) potently agonizes TRPV3 [32]. | Primarily expressed in epidermal keratinocytes [39,40,41], with suggested expression in the brain [42]. In the GIT, it is expressed particularly in the cecum and colon [43], and is localized predominantly to the apical membranes of epithelial cells [44]. | TRPV3 plays a vital role in sensing warmth and maintaining skin barrier function [39,40,41]. In the gut, it plays a crucial role in ammonium (NH4+) transport through its divalent-sensitive cation conductance [43], contributing to nitrogen metabolism and gut homeostasis [43]. It influences epithelial permeability and maintains barrier integrity [44], modulates transepithelial ionic balance [44,45], and may have a potential role in host-microbe interactions and nutrient absorption [44,45]. |
| TRPV4 | Activated by mechanical stress, osmotic changes, moderate temperatures (25–35 °C), arachidonic acid metabolites, and the selective pharmacological agonist GSK1016790A [50,51,52,53,57]. | Widely expressed in multiple tissues and cell types [5,46,47,48,49], including endothelial cells, sensory neurons, urothelial cells, and chondrocytes [5,46,47,48,49]. It is present in the GIT milieu [54], and expressed in salivary glands, esophagus, and stomach [54,56,57] | TRPV4 is involved in regulating GI motility [54] and fluid secretion, which is essential for salivary flow [54]. It actively participates in GI mechanosensation, influencing gastric emptying and intestinal transit [55]. It regulates calcium influx and ATP release in the esophagus and stomach [56,57], is crucial for maintaining epithelial integrity and sensory transduction pathways [57], contributes to esophageal mechanosensation and regulation of peristalsis [55,58], and plays a role in regulating gastric motility and pancreatic fluid secretion [57,59]. |
| TRPV5 & TRPV6 | While TRPV5 is positively modulated by Klotho and sialidase [64], both channels are activated by extracellular calcium as well as vitamin D, 1,25-dihydroxyvitamin D3 (1,25[OH]2D3) [60,62,63] and the membrane phospholipid phosphatidylinositol 4,5-bisphosphate (PI(4,5)P2), which is essential for TRPV5 opening and enhances TRPV6 function by relieving auto-inhibition [65,66]. | Predominantly expressed in GI epithelial cells [60], specifically on the apical membrane of epithelial cells in the small intestine, particularly in the duodenum and jejunum [60]. | These channels are crucial for maintaining calcium homeostasis [60], being highly selective for calcium ions and enabling calcium influx [60]. They are instrumental in maintaining systemic calcium balance [67,68]. TRPV6 also participates in the absorption of magnesium within the intestine [67,68] and influences cell proliferation, differentiation, and apoptosis via cellular signaling pathways [67,68]. |
5. Potential Role of TRPV Channels in the Pathophysiology of Different GIT Disorders
5.1. Mediation of Gut Pain
5.1.1. TRPV1 Channel and Its Contribution to Gut Pain
5.1.2. TRPV4 Channel and Its Contribution to Gut Pain
5.2. Facilitation of Gut Motility and Regulation of Secretions
5.2.1. Role of TRPV1 Channel in the Motor Activity and Secretory Processes of the Gut
5.2.2. Role of TRPV2 Channel in Motor Activity of the Gut
5.2.3. Role of TRPV4 Channel in Motor Activity and Secretory Processes of the Gut
5.3. Role of TRPV Channels in Gut Inflammation
5.3.1. Role of TRPV1 Channel in Gut Inflammation
5.3.2. Role of TRPV2 Channel in Gut Inflammation
5.3.3. Role of TRPV4 Channel in Gut Inflammation
5.3.4. Role of TRPV6 Channel in Gut Inflammation
5.4. Role of TRPV Channels in Ulcer Formation and Healing
5.4.1. TRPV Channels and Ulcer Formation
5.4.2. Role of TRPV Channels in Ulcer Healing
5.4.3. Implications for Therapeutic Targeting of TRPV Channels in Ulcerative Diseases
6. Role of TRPV Channels in GIT Tumors
6.1. Oral Cavity Cancer
6.2. Esophageal Cancer
6.3. Gastric Cancer
6.4. Intestinal Adenoma
6.5. Colorectal Cancer
6.6. Neuroendocrine Tumors
7. Potential Challenges with Targeting TRPVs for Therapy
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| 2-AG | 2-Arachidonoylglycerol |
| 2-APB | 2-Aminoethoxydiphenyl Borate |
| 12-HETE | 12-Hydroxyeicosatetraenoic Acid |
| Akt | Protein Kinase B |
| AMPK | AMP-activated protein kinase |
| ARD | Ankyrin Repeat Domain |
| ATP | Adenosine Triphosphate |
| Ca2+ | Calcium Ion |
| CaMKII | Calcium/Calmodulin-Dependent Protein Kinase II |
| CaSR | Calcium-Sensing Receptor |
| CBD | Cannabidiol |
| CGRP | Calcitonin Gene-Related Peptide |
| CRC | Colorectal Cancer |
| DAG | Diacylglycerol |
| DRG | Dorsal Root Ganglia |
| DSS | Dextran Sulfate Sodium |
| EGFR | Epidermal Growth Factor Receptor |
| ERK1/2 | Extracellular Signal-Regulated Kinase 1/2 |
| ESCC | Esophageal Squamous Cell Carcinoma |
| GALT | Gut-Associated Lymphoid Tissue |
| GC | Gastric Cancer |
| GERD | Gastroesophageal Reflux Disease |
| GIT | Gastrointestinal Tract |
| GPCR | G Protein-Coupled Receptors |
| IBD | Inflammatory Bowel Disease |
| IBS | Irritable Bowel Syndrome |
| IFN-γ | Interferon Gamma |
| IL-11 | Interleukin-11 |
| IL-17 | Interleukin-17 |
| IL-2 | Interleukin-2 |
| IL-6 | Interleukin-6 |
| IP3 | Inositol 1,4,5-Trisphosphate |
| LPC | Lysophosphatidylcholine |
| MMs | Muscularis Macrophages |
| NFAT | Nuclear Factor of Activated T-cells |
| NF-κB | Nuclear Factor Kappa-Light-Chain-Enhancer of Activated B Cells |
| NH4+ | Ammonium ion |
| NO | Nitric Oxide |
| OLDA | Oleoylethanolamide |
| PACAP | Pituitary Adenylate Cyclase-Activating Peptide |
| PanNETs | Pancreatic Neuroendocrine Tumors |
| PAR-2 | Proteinase-Activated Receptor 2 |
| PH | Pore Helix |
| PI3K | Phosphoinositide 3-Kinase |
| PIP2 | Phosphatidylinositol 4,5-Bisphosphate |
| PKC | Protein Kinase C |
| PLC | Phospholipase C |
| PTEN | Phosphatase and Tensin Homolog |
| PGE2 | Prostaglandin E2 |
| ROS | Reactive Oxygen Species |
| SCC | Squamous Cell Carcinoma |
| SMCs | Smooth Muscle Cells |
| STAT3 | Signal Transducer and Activator of Transcription 3 |
| TRP | Transient Receptor Potential |
| TRPA | Transient Receptor Potential Ankyrin |
| TRPC | Transient Receptor Potential Canonical |
| TRPM | Transient Receptor Potential Melastatin |
| TRPML | Transient Receptor Potential Mucolipin |
| TRPN | Transient Receptor Potential No-Mechano-Potential |
| TRPP | Transient Receptor Potential Polycystin |
| TRPV | Transient Receptor Potential Vanilloid |
| VIP | Vasoactive Intestinal Peptide |
| VNUT | Vesicular Nucleotide Transporter |
| VSLD | Voltage-Sensing-Like Domain |
References
- Gieryńska, M.; Szulc-Dąbrowska, L.; Struzik, J.; Mielcarska, M.B.; Gregorczyk-Zboroch, K.P. Integrity of the Intestinal Barrier: The Involvement of Epithelial Cells and Microbiota-A Mutual Relationship. Animals 2022, 12, 145. [Google Scholar] [CrossRef]
- Farré, R.; Fiorani, M.; Abdu Rahiman, S.; Matteoli, G. Intestinal Permeability, Inflammation and the Role of Nutrients. Nutrients 2020, 12, 1185. [Google Scholar] [CrossRef]
- Koivisto, A.P.; Belvisi, M.G.; Gaudet, R.; Szallasi, A. Advances in TRP channel drug discovery: From target validation to clinical studies. Nat. Rev. Drug Discov. 2022, 21, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Samanta, A.; Hughes, T.E.T.; Moiseenkova-Bell, V.Y. Transient Receptor Potential (TRP) Channels. Subcell. Biochem. 2018, 87, 141–165. [Google Scholar] [CrossRef] [PubMed]
- Boudaka, A.; Al-Suleimani, M.; Al-Lawati, I.; Baomar, H.; Al-Siyabi, S.; Zadjali, F. Downregulation of endothelial transient receptor potential vanilloid type 4 channel underlines impaired endothelial nitric oxide-mediated relaxation in the mesenteric arteries of hypertensive rats. Physiol. Res. 2019, 68, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Boudaka, A.; Al-Yazeedi, M.; Al-Lawati, I. Role of Transient Receptor Potential Vanilloid 4 Channel in Skin Physiology and Pathology. Sultan Qaboos Univ. Med. J. 2020, 20, e138–e146. [Google Scholar] [CrossRef]
- Pedersen, S.F.; Owsianik, G.; Nilius, B. TRP channels: An overview. Cell Calcium 2005, 38, 233–252. [Google Scholar] [CrossRef]
- Zhang, M.; Ma, Y.; Ye, X.; Zhang, N.; Pan, L.; Wang, B. TRP (transient receptor potential) ion channel family: Structures, biological functions and therapeutic interventions for diseases. Signal Transduct. Target. Ther. 2023, 8, 261. [Google Scholar] [CrossRef]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef]
- Seebohm, G.; Schreiber, J.A. Beyond Hot and Spicy: TRPV Channels and their Pharmacological Modulation. Cell Physiol. Biochem. 2021, 55, 108–130. [Google Scholar] [CrossRef]
- Pumroy, R.A.; Fluck, E.C., 3rd; Ahmed, T.; Moiseenkova-Bell, V.Y. Structural insights into the gating mechanisms of TRPV channels. Cell Calcium 2020, 87, 102168. [Google Scholar] [CrossRef]
- Caviglia, G.P.; Garrone, A.; Bertolino, C.; Vanni, R.; Bretto, E.; Poshnjari, A.; Tribocco, E.; Frara, S.; Armandi, A.; Astegiano, M.; et al. Epidemiology of Inflammatory Bowel Diseases: A Population Study in a Healthcare District of North-West Italy. J. Clin. Med. 2023, 12, 641. [Google Scholar] [CrossRef]
- Gusenbauer, M. Beyond Google Scholar, Scopus, and Web of Science: An evaluation of the backward and forward citation coverage of 59 databases’ citation indices. Res. Synth. Methods 2024, 15, 802–817. [Google Scholar] [CrossRef]
- Hellmich, U.A.; Gaudet, R. Structural biology of TRP channels. Handb. Exp. Pharmacol. 2014, 223, 963–990. [Google Scholar] [CrossRef] [PubMed]
- Cao, E.; Liao, M.; Cheng, Y.; Julius, D. TRPV1 structures in distinct conformations reveal activation mechanisms. Nature 2013, 504, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Dosey, T.L.; Wang, Z.; Fan, G.; Zhang, Z.; Serysheva, I.I.; Chiu, W.; Wensel, T.G. Structures of TRPV2 in distinct conformations provide insight into role of the pore turret. Nat. Struct. Mol. Biol. 2019, 26, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Phelps, C.B.; Wang, R.R.; Choo, S.S.; Gaudet, R. Differential regulation of TRPV1, TRPV3, and TRPV4 sensitivity through a conserved binding site on the ankyrin repeat domain. J. Biol. Chem. 2010, 285, 731–740. [Google Scholar] [CrossRef]
- Rosenbaum, T.; Islas, L.D. Molecular Physiology of TRPV Channels: Controversies and Future Challenges. Annu. Rev. Physiol. 2023, 85, 293–316. [Google Scholar] [CrossRef]
- Veldhuis, N.A.; Poole, D.P.; Grace, M.; McIntyre, P.; Bunnett, N.W. The G protein-coupled receptor-transient receptor potential channel axis: Molecular insights for targeting disorders of sensation and inflammation. Pharmacol. Rev. 2015, 67, 36–73. [Google Scholar] [CrossRef]
- Adapala, R.K.; Katari, V.; Teegala, L.R.; Thodeti, S.; Paruchuri, S.; Thodeti, C.K. TRPV4 mechanotransduction in fibrosis. Cells 2021, 10, 3053. [Google Scholar] [CrossRef]
- Holzer, P. The pharmacological challenge to tame the transient receptor potential vanilloid-1 (TRPV1) nocisensor. Br. J. Pharmacol. 2008, 155, 1145–1162. [Google Scholar] [CrossRef]
- Ciura, S.; Liedtke, W.; Bourque, C.W. Hypertonicity sensing in organum vasculosum lamina terminalis neurons: A mechanical process involving TRPV1 but not TRPV4. J. Neurosci. 2011, 31, 14669–14676. [Google Scholar] [CrossRef] [PubMed]
- Laleh, P.; Yaser, K.; Alireza, O. Oleoylethanolamide: A novel pharmaceutical agent in the management of obesity-an updated review. J. Cell Physiol. 2019, 234, 7893–7902. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.A.; Leffler, A.; Niedermirtl, F.; Babes, A.; Zimmermann, K.; Filipović, M.R.; Izydorczyk, I.; Eberhardt, M.; Kichko, T.I.; Mueller-Tribbensee, S.M.; et al. TRPA1 and substance P mediate colitis in mice. Gastroenterology 2011, 141, 1346–1358. [Google Scholar] [CrossRef] [PubMed]
- Boudaka, A.; Wörl, J.; Shiina, T.; Neuhuber, W.L.; Kobayashi, H.; Shimizu, Y.; Takewaki, T. Involvement of TRPV1-dependent and -independent components in the regulation of vagally induced contractions in the mouse esophagus. Eur. J. Pharmacol. 2007, 556, 157–165. [Google Scholar] [CrossRef]
- Holzer, P. Taste receptors in the gastrointestinal tract. V. Acid sensing in the gastrointestinal tract. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G699–G705. [Google Scholar] [CrossRef]
- Holzer, P. TRPV1: A new target for treatment of visceral pain in IBS? Gut 2008, 57, 882–884. [Google Scholar] [CrossRef]
- Du, Q.; Liao, Q.; Chen, C.; Yang, X.; Xie, R.; Xu, J. The Role of Transient Receptor Potential Vanilloid 1 in Common Diseases of the Digestive Tract and the Cardiovascular and Respiratory System. Front. Physiol. 2019, 10, 1064. [Google Scholar] [CrossRef]
- Park, U.; Vastani, N.; Guan, Y.; Raja, S.N.; Koltzenburg, M.; Caterina, M.J. TRP vanilloid 2 knock-out mice are susceptible to perinatal lethality but display normal thermal and mechanical nociception. J. Neurosci. 2011, 31, 11425–11436. [Google Scholar] [CrossRef]
- Shibasaki, K.; Ishizaki, Y.; Mandadi, S. Astrocytes express functional TRPV2 ion channels. Biochem. Biophys. Res. Commun. 2013, 441, 327–332. [Google Scholar] [CrossRef]
- Ciardo, M.G.; Ferrer-Montiel, A. Lipids as central modulators of sensory TRP channels. Biochim. Biophys. Acta Biomembr. 2017, 1859 Pt B, 1615–1628. [Google Scholar] [CrossRef]
- Pumroy, R.A.; Samanta, A.; Liu, Y.; Hughes, T.E.; Zhao, S.; Yudin, Y.; Rohacs, T.; Han, S.; Moiseenkova-Bell, V.Y. Molecular mechanism of TRPV2 channel modulation by cannabidiol. Elife 2019, 8, e48792. [Google Scholar] [CrossRef]
- Parenti, A.; De Logu, F.; Geppetti, P.; Benemei, S. What is the evidence for the role of TRP channels in inflammatory and immune cells? Br. J. Pharmacol. 2016, 173, 953–969. [Google Scholar] [CrossRef] [PubMed]
- Santoni, G.; Farfariello, V.; Liberati, S.; Morelli, M.B.; Nabissi, M.; Santoni, M.; Amantini, C. The role of transient receptor potential vanilloid type-2 ion channels in innate and adaptive immune responses. Front. Immunol. 2013, 4, 34. [Google Scholar] [CrossRef] [PubMed]
- Kashiba, H.; Uchida, Y.; Takeda, D.; Nishigori, A.; Ueda, Y.; Kuribayashi, K.; Ohshima, M. TRPV2-immunoreactive intrinsic neurons in the rat intestine. Neurosci. Lett. 2004, 366, 193–196. [Google Scholar] [CrossRef] [PubMed]
- Mihara, H.; Boudaka, A.; Shibasaki, K.; Yamanaka, A.; Sugiyama, T.; Tominaga, M. Involvement of TRPV2 activation in intestinal movement through nitric oxide production in mice. J. Neurosci. 2010, 30, 16536–16544. [Google Scholar] [CrossRef]
- Doerner, J.F.; Hatt, H.; Ramsey, I.S. Voltage- and temperature-dependent activation of TRPV3 channels is potentiated by receptor-mediated PI(4,5)P2 hydrolysis. J. Gen. Physiol. 2011, 137, 271–288. [Google Scholar] [CrossRef]
- Vogt-Eisele, A.K.; Weber, K.; Sherkheli, M.A.; Vielhaber, G.; Panten, J.; Gisselmann, G.; Hatt, H. Monoterpenoid agonists of TRPV3. Br. J. Pharmacol. 2007, 151, 530–540. [Google Scholar] [CrossRef]
- Nilius, B.; Bíró, T.; Owsianik, G. TRPV3: Time to decipher a poorly understood family member! J. Physiol. 2014, 592, 295–304. [Google Scholar] [CrossRef]
- Xu, H.; Blair, N.T.; Clapham, D.E. Camphor activates and strongly desensitizes the transient receptor potential vanilloid subtype 1 channel in a vanilloid-independent mechanism. J. Neurosci. 2005, 25, 8924–8937. [Google Scholar] [CrossRef]
- Colton, C.K.; Zhu, M.X. 2-Aminoethoxydiphenyl borate as a common activator of TRPV1, TRPV2, and TRPV3 channels. Handb. Exp. Pharmacol. 2007, 179, 173–187. [Google Scholar] [CrossRef]
- Singh, R.; Bansal, Y.; Sodhi, R.K.; Khare, P.; Bishnoi, M.; Kondepudi, K.K.; Medhi, B.; Kuhad, A. Role of TRPV1/TRPV3 channels in olanzapine-induced metabolic alteration: Possible involvement in hypothalamic energy-sensing, appetite regulation, inflammation and mesolimbic pathway. Toxicol. Appl. Pharmacol. 2020, 402, 115124. [Google Scholar] [CrossRef] [PubMed]
- Schrapers, K.T.; Sponder, G.; Liebe, F.; Liebe, H.; Stumpff, F. The bovine TRPV3 as a pathway for the uptake of Na(+), Ca(2+), and NH(4)(+). PLoS ONE 2018, 13, e0193519. [Google Scholar] [CrossRef] [PubMed]
- Liebe, F.; Liebe, H.; Kaessmeyer, S.; Sponder, G.; Stumpff, F. The TRPV3 channel of the bovine rumen: Localization and functional characterization of a protein relevant for ruminal ammonia transport. Pflug. Arch. 2020, 472, 693–710. [Google Scholar] [CrossRef] [PubMed]
- Manneck, D.; Braun, H.S.; Schrapers, K.T.; Stumpff, F. TRPV3 and TRPV4 as candidate proteins for intestinal ammonium absorption. Acta Physiol. 2021, 233, e13694. [Google Scholar] [CrossRef]
- Mochizuki, T.; Sokabe, T.; Araki, I.; Fujishita, K.; Shibasaki, K.; Uchida, K.; Naruse, K.; Koizumi, S.; Takeda, M.; Tominaga, M. The TRPV4 cation channel mediates stretch-evoked Ca2+ influx and ATP release in primary urothelial cell cultures. J. Biol. Chem. 2009, 284, 21257–21264. [Google Scholar] [CrossRef]
- O’Conor, C.J.; Leddy, H.A.; Benefield, H.C.; Liedtke, W.B.; Guilak, F. TRPV4-mediated mechanotransduction regulates the metabolic response of chondrocytes to dynamic loading. Proc. Natl. Acad. Sci. USA 2014, 111, 1316–1321. [Google Scholar] [CrossRef]
- Darby, W.G.; Grace, M.S.; Baratchi, S.; McIntyre, P. Modulation of TRPV4 by diverse mechanisms. Int. J. Biochem. Cell Biol. 2016, 78, 217–228. [Google Scholar] [CrossRef]
- Maqboul, A.; Elsadek, B. Expression profiles of TRPV1, TRPV4, TLR4 and ERK1/2 in the dorsal root ganglionic neurons of a cancer-induced neuropathy rat model. PeerJ 2018, 6, e4622. [Google Scholar] [CrossRef]
- Güler, A.D.; Lee, H.; Iida, T.; Shimizu, I.; Tominaga, M.; Caterina, M. Heat-evoked activation of the ion channel, TRPV4. J. Neurosci. 2002, 22, 6408–6414. [Google Scholar] [CrossRef]
- Suzuki, M.; Mizuno, A.; Kodaira, K.; Imai, M. Impaired pressure sensation in mice lacking TRPV4. J. Biol. Chem. 2003, 278, 22664–22668. [Google Scholar] [CrossRef] [PubMed]
- Berna-Erro, A.; Izquierdo-Serra, M.; Sepúlveda, R.V.; Rubio-Moscardo, F.; Doñate-Macián, P.; Serra, S.A.; Carrillo-Garcia, J.; Perálvarez-Marín, A.; González-Nilo, F.; Fernández-Fernández, J.M.; et al. Structural determinants of 5′,6′-epoxyeicosatrienoic acid binding to and activation of TRPV4 channel. Sci. Rep. 2017, 7, 10522. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Anishkin, A.; Zinkevich, N.S.; Nishijima, Y.; Korishettar, A.; Wang, Z.; Fang, J.; Wilcox, D.A.; Zhang, D.X. Transient receptor potential vanilloid 4 (TRPV4) activation by arachidonic acid requires protein kinase A-mediated phosphorylation. J. Biol. Chem. 2018, 293, 5307–5322. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Ma, D.; Zhu, X.; Wu, Z.; An, Q.; Zhao, J.; Gao, X.; Zhang, L. Roles of TRP and PIEZO receptors in autoimmune diseases. Expert. Rev. Mol. Med. 2024, 26, e10. [Google Scholar] [CrossRef]
- Boudaka, A.; Tominaga, M. Physiological and Pathological Significance of Esophageal TRP Channels: Special Focus on TRPV4 in Esophageal Epithelial Cells. Int. J. Mol. Sci. 2022, 23, 4550. [Google Scholar] [CrossRef]
- Mihara, H.; Boudaka, A.; Tominaga, M.; Sugiyama, T. Transient Receptor Potential Vanilloid 4 Regulation of Adenosine Triphosphate Release by the Adenosine Triphosphate Transporter Vesicular Nucleotide Transporter, a Novel Therapeutic Target for Gastrointestinal Baroreception and Chronic Inflammation. Digestion 2020, 101, 6–11. [Google Scholar] [CrossRef]
- Mihara, H.; Suzuki, N.; Boudaka, A.A.; Muhammad, J.S.; Tominaga, M.; Tabuchi, Y.; Sugiyama, T. Transient receptor potential vanilloid 4-dependent calcium influx and ATP release in mouse and rat gastric epithelia. World J. Gastroenterol. 2016, 22, 5512–5519. [Google Scholar] [CrossRef]
- Mihara, H.; Boudaka, A.; Sugiyama, T.; Moriyama, Y.; Tominaga, M. Transient receptor potential vanilloid 4 (TRPV4)-dependent calcium influx and ATP release in mouse oesophageal keratinocytes. J. Physiol. 2011, 589 Pt 14, 3471–3482. [Google Scholar] [CrossRef]
- Boudaka, A.; Saito, C.T.; Tominaga, M. Deletion of TRPV4 enhances in vitro wound healing of murine esophageal keratinocytes. Sci. Rep. 2020, 10, 11349. [Google Scholar] [CrossRef]
- Hoenderop, J.G.; Nilius, B.; Bindels, R.J. Calcium absorption across epithelia. Physiol. Rev. 2005, 85, 373–422. [Google Scholar] [CrossRef]
- Peng, J.B.; Suzuki, Y.; Gyimesi, G.; Hediger, M.A. TRPV5 and TRPV6 Calcium-Selective Channels. In Calcium Entry Channels in Non-Excitable Cells; Kozak, J.A., Putney, J.W., Jr., Eds.; CRC Press/Taylor & Francis, LLC.: Boca Raton, FL, USA, 2018; pp. 241–274. [Google Scholar]
- Na, T.; Peng, J.B. TRPV5: A Ca(2+) channel for the fine-tuning of Ca(2+) reabsorption. Handb. Exp. Pharmacol. 2014, 222, 321–357. [Google Scholar] [CrossRef] [PubMed]
- Fecher-Trost, C.; Weissgerber, P.; Wissenbach, U. TRPV6 channels. Handb. Exp. Pharmacol. 2014, 222, 359–384. [Google Scholar] [CrossRef]
- Leunissen, E.H.; Nair, A.V.; Büll, C.; Lefeber, D.J.; van Delft, F.L.; Bindels, R.J.; Hoenderop, J.G. The epithelial calcium channel TRPV5 is regulated differentially by klotho and sialidase. J. Biol. Chem. 2013, 288, 29238–29246. [Google Scholar] [CrossRef]
- Hughes, T.E.; Del Rosario, J.S.; Kapoor, A.; Yazici, A.T.; Yudin, Y.; Fluck, E.C., 3rd; Filizola, M.; Rohacs, T.; Moiseenkova-Bell, V.Y. Structure-based characterization of novel TRPV5 inhibitors. Elife 2019, 8, e49572. [Google Scholar] [CrossRef]
- Cai, R.; Wang, L.; Liu, X.; Michalak, M.; Tang, J.; Peng, J.-B.; Chen, X.-Z. Auto-inhibitory intramolecular S5/S6 interaction in the TRPV6 channel regulates breast cancer cell migration and invasion. Commun. Biol. 2021, 4, 990. [Google Scholar] [CrossRef]
- Stewart, J.M. TRPV6 as A Target for Cancer Therapy. J. Cancer 2020, 11, 374–387. [Google Scholar] [CrossRef]
- Khattar, V.; Wang, L.; Peng, J.B. Calcium selective channel TRPV6: Structure, function, and implications in health and disease. Gene 2022, 817, 146192. [Google Scholar] [CrossRef]
- Lai, N.Y.; Mills, K.; Chiu, I.M. Sensory neuron regulation of gastrointestinal inflammation and bacterial host defence. J. Intern. Med. 2017, 282, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Lagomarsino, V.N.; Kostic, A.D.; Chiu, I.M. Mechanisms of microbial-neuronal interactions in pain and nociception. Neurobiol. Pain. 2021, 9, 100056. [Google Scholar] [CrossRef]
- Alaimo, A.; Rubert, J. The Pivotal Role of TRP Channels in Homeostasis and Diseases throughout the Gastrointestinal Tract. Int. J. Mol. Sci. 2019, 20, 5277. [Google Scholar] [CrossRef] [PubMed]
- Holzer, P. Transient receptor potential (TRP) channels as drug targets for diseases of the digestive system. Pharmacol. Ther. 2011, 131, 142–170. [Google Scholar] [CrossRef] [PubMed]
- Holzer, P. TRP channels in the digestive system. Curr. Pharm. Biotechnol. 2011, 12, 24–34. [Google Scholar] [CrossRef] [PubMed]
- Hicks, G.A. TRP channels as therapeutic targets: Hot property, or time to cool down? Neurogastroenterol. Motil. 2006, 18, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Akbar, A.; Yiangou, Y.; Facer, P.; Walters, J.R.; Anand, P.; Ghosh, S. Increased capsaicin receptor TRPV1-expressing sensory fibres in irritable bowel syndrome and their correlation with abdominal pain. Gut 2008, 57, 923–929. [Google Scholar] [CrossRef]
- Akbar, A.; Yiangou, Y.; Facer, P.; Brydon, W.G.; Walters, J.R.; Anand, P.; Ghosh, S. Expression of the TRPV1 receptor differs in quiescent inflammatory bowel disease with or without abdominal pain. Gut 2010, 59, 767–774. [Google Scholar] [CrossRef]
- Chan, C.L.; Facer, P.; Davis, J.B.; Smith, G.D.; Egerton, J.; Bountra, C.; Williams, N.S.; Anand, P. Sensory fibres expressing capsaicin receptor TRPV1 in patients with rectal hypersensitivity and faecal urgency. Lancet 2003, 361, 385–391. [Google Scholar] [CrossRef]
- Bhat, Y.M.; Bielefeldt, K. Capsaicin receptor (TRPV1) and non-erosive reflux disease. Eur. J. Gastroenterol. Hepatol. 2006, 18, 263–270. [Google Scholar] [CrossRef]
- Hammer, J.; Führer, M.; Pipal, L.; Matiasek, J. Hypersensitivity for capsaicin in patients with functional dyspepsia. Neurogastroenterol. Motil. 2008, 20, 125–133. [Google Scholar] [CrossRef]
- Gonlachanvit, S.; Mahayosnond, A.; Kullavanijaya, P. Effects of chili on postprandial gastrointestinal symptoms in diarrhoea predominant irritable bowel syndrome: Evidence for capsaicin-sensitive visceral nociception hypersensitivity. Neurogastroenterol. Motil. 2009, 21, 23–32. [Google Scholar] [CrossRef]
- Sipe, W.E.; Brierley, S.M.; Martin, C.M.; Phillis, B.D.; Cruz, F.B.; Grady, E.F.; Liedtke, W.; Cohen, D.M.; Vanner, S.; Blackshaw, L.A.; et al. Transient receptor potential vanilloid 4 mediates protease activated receptor 2-induced sensitization of colonic afferent nerves and visceral hyperalgesia. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G1288–G1298. [Google Scholar] [CrossRef]
- Brierley, S.M.; Page, A.J.; Hughes, P.A.; Adam, B.; Liebregts, T.; Cooper, N.J.; Holtmann, G.; Liedtke, W.; Blackshaw, L.A. Selective role for TRPV4 ion channels in visceral sensory pathways. Gastroenterology 2008, 134, 2059–2069. [Google Scholar] [CrossRef] [PubMed]
- Cenac, N.; Altier, C.; Chapman, K.; Liedtke, W.; Zamponi, G.; Vergnolle, N. Transient receptor potential vanilloid-4 has a major role in visceral hypersensitivity symptoms. Gastroenterology 2008, 135, 937–946, 946.e1–946e2. [Google Scholar] [CrossRef] [PubMed]
- Alessandri-Haber, N.; Dina, O.A.; Joseph, E.K.; Reichling, D.; Levine, J.D. A transient receptor potential vanilloid 4-dependent mechanism of hyperalgesia is engaged by concerted action of inflammatory mediators. J. Neurosci. 2006, 26, 3864–3874. [Google Scholar] [CrossRef] [PubMed]
- Grant, A.D.; Cottrell, G.S.; Amadesi, S.; Trevisani, M.; Nicoletti, P.; Materazzi, S.; Altier, C.; Cenac, N.; Zamponi, G.W.; Bautista-Cruz, F.; et al. Protease-activated receptor 2 sensitizes the transient receptor potential vanilloid 4 ion channel to cause mechanical hyperalgesia in mice. J. Physiol. 2007, 578 Pt 3, 715–733. [Google Scholar] [CrossRef]
- Zhao, P.; Lieu, T.; Barlow, N.; Metcalf, M.; Veldhuis, N.A.; Jensen, D.D.; Kocan, M.; Sostegni, S.; Haerteis, S.; Baraznenok, V.; et al. Cathepsin S causes inflammatory pain via biased agonism of PAR2 and TRPV4. J. Biol. Chem. 2014, 289, 27215–27234. [Google Scholar] [CrossRef]
- Zhao, P.; Lieu, T.; Barlow, N.; Sostegni, S.; Haerteis, S.; Korbmacher, C.; Liedtke, W.; Jimenez-Vargas, N.N.; Vanner, S.J.; Bunnett, N.W. Neutrophil Elastase Activates Protease-activated Receptor-2 (PAR2) and Transient Receptor Potential Vanilloid 4 (TRPV4) to Cause Inflammation and Pain. J. Biol. Chem. 2015, 290, 13875–13887. [Google Scholar] [CrossRef]
- Cenac, N.; Altier, C.; Motta, J.P.; d’Aldebert, E.; Galeano, S.; Zamponi, G.W.; Vergnolle, N. Potentiation of TRPV4 signalling by histamine and serotonin: An important mechanism for visceral hypersensitivity. Gut 2010, 59, 481–488. [Google Scholar] [CrossRef]
- Weber, E.; Neunlist, M.; Schemann, M.; Frieling, T. Neural components of distension-evoked secretory responses in the guinea-pig distal colon. J. Physiol. 2001, 536 Pt 3, 741–751. [Google Scholar] [CrossRef]
- Boudaka, A.; Wörl, J.; Shiina, T.; Saito, S.; Atoji, Y.; Kobayashi, H.; Shimizu, Y.; Takewaki, T. Key role of mucosal primary afferents in mediating the inhibitory influence of capsaicin on vagally mediated contractions in the mouse esophagus. J. Vet. Med. Sci. 2007, 69, 365–372. [Google Scholar] [CrossRef]
- Boudaka, A.; Wörl, J.; Shiina, T.; Shimizu, Y.; Takewaki, T.; Neuhuber, W.L. Galanin modulates vagally induced contractions in the mouse oesophagus. Neurogastroenterol. Motil. 2009, 21, 180–188. [Google Scholar] [CrossRef]
- Benko, R.; Lazar, Z.; Undi, S.; Illenyi, L.; Antal, A.; Horvath, O.P.; Rumbus, Z.; Wolf, M.; Maggi, C.A.; Bartho, L. Inhibition of nitric oxide synthesis blocks the inhibitory response to capsaicin in intestinal circular muscle preparations from different species. Life Sci. 2005, 76, 2773–2782. [Google Scholar] [CrossRef]
- Holzer, P.; Lippe, I.T.; Holzer-Petsche, U. Inhibition of gastrointestinal transit due to surgical trauma or peritoneal irritation is reduced in capsaicin-treated rats. Gastroenterology 1986, 91, 360–363. [Google Scholar] [CrossRef]
- De Schepper, H.U.; De Man, J.G.; Ruyssers, N.E.; Deiteren, A.; Van Nassauw, L.; Timmermans, J.P.; Martinet, W.; Herman, A.G.; Pelckmans, P.A.; De Winter, B.Y. TRPV1 receptor signaling mediates afferent nerve sensitization during colitis-induced motility disorders in rats. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 294, G245–G253. [Google Scholar] [CrossRef]
- Ding, Q.W.; Zhang, Y.; Wang, Y.; Wang, Y.N.; Zhang, L.; Ding, C.; Wu, L.L.; Yu, G.Y. Functional vanilloid receptor-1 in human submandibular glands. J. Dent. Res. 2010, 89, 711–716. [Google Scholar] [CrossRef]
- Cheng, L.; de la Monte, S.; Ma, J.; Hong, J.; Tong, M.; Cao, W.; Behar, J.; Biancani, P.; Harnett, K.M. HCl-activated neural and epithelial vanilloid receptors (TRPV1) in cat esophageal mucosa. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G135–G143. [Google Scholar] [CrossRef] [PubMed]
- Ericson, A.; Nur, E.M.; Petersson, F.; Kechagias, S. The effects of capsaicin on gastrin secretion in isolated human antral glands: Before and after ingestion of red chilli. Dig. Dis. Sci. 2009, 54, 491–498. [Google Scholar] [CrossRef] [PubMed]
- Schicho, R.; Krueger, D.; Zeller, F.; Von Weyhern, C.W.; Frieling, T.; Kimura, H.; Ishii, I.; De Giorgio, R.; Campi, B.; Schemann, M. Hydrogen sulfide is a novel prosecretory neuromodulator in the Guinea-pig and human colon. Gastroenterology 2006, 131, 1542–1552. [Google Scholar] [CrossRef] [PubMed]
- Gradilone, S.A.; Masyuk, A.I.; Splinter, P.L.; Banales, J.M.; Huang, B.Q.; Tietz, P.S.; Masyuk, T.V.; Larusso, N.F. Cholangiocyte cilia express TRPV4 and detect changes in luminal tonicity inducing bicarbonate secretion. Proc. Natl. Acad. Sci. USA 2007, 104, 19138–19143. [Google Scholar] [CrossRef]
- Gradilone, S.A.; Masyuk, T.V.; Huang, B.Q.; Banales, J.M.; Lehmann, G.L.; Radtke, B.N.; Stroope, A.; Masyuk, A.I.; Splinter, P.L.; LaRusso, N.F. Activation of Trpv4 reduces the hyperproliferative phenotype of cystic cholangiocytes from an animal model of ARPKD. Gastroenterology 2010, 139, 304–314.e302. [Google Scholar] [CrossRef]
- Luo, J.; Qian, A.; Oetjen, L.K.; Yu, W.; Yang, P.; Feng, J.; Xie, Z.; Liu, S.; Yin, S.; Dryn, D.; et al. TRPV4 Channel Signaling in Macrophages Promotes Gastrointestinal Motility via Direct Effects on Smooth Muscle Cells. Immunity 2018, 49, 107–119.e104. [Google Scholar] [CrossRef]
- Gazzieri, D.; Trevisani, M.; Springer, J.; Harrison, S.; Cottrell, G.S.; Andre, E.; Nicoletti, P.; Massi, D.; Zecchi, S.; Nosi, D.; et al. Substance P released by TRPV1-expressing neurons produces reactive oxygen species that mediate ethanol-induced gastric injury. Free Radic. Biol. Med. 2007, 43, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Chen, J.; Shen, L.; Wang, B. TRP channels in inflammatory bowel disease: Potential therapeutic targets. Biochem. Pharmacol. 2022, 203, 115195. [Google Scholar] [CrossRef]
- Shibasaki, K. Physiological significance of TRPV2 as a mechanosensor, thermosensor and lipid sensor. J. Physiol. Sci. 2016, 66, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Levine, J.D.; Alessandri-Haber, N. TRP channels: Targets for the relief of pain. Biochim. Biophys. Acta 2007, 1772, 989–1003. [Google Scholar] [CrossRef] [PubMed]
- Issa, C.M.; Hambly, B.D.; Wang, Y.; Maleki, S.; Wang, W.; Fei, J.; Bao, S. TRPV2 in the development of experimental colitis. Scand. J. Immunol. 2014, 80, 307–312. [Google Scholar] [CrossRef]
- D’Aldebert, E.; Cenac, N.; Rousset, P.; Martin, L.; Rolland, C.; Chapman, K.; Selves, J.; Alric, L.; Vinel, J.P.; Vergnolle, N. Transient receptor potential vanilloid 4 activated inflammatory signals by intestinal epithelial cells and colitis in mice. Gastroenterology 2011, 140, 275–285. [Google Scholar] [CrossRef]
- Vincent, F.; Acevedo, A.; Nguyen, M.T.; Dourado, M.; DeFalco, J.; Gustafson, A.; Spiro, P.; Emerling, D.E.; Kelly, M.G.; Duncton, M.A. Identification and characterization of novel TRPV4 modulators. Biochem. Biophys. Res. Commun. 2009, 389, 490–494. [Google Scholar] [CrossRef]
- Meena, A.S.; Shukla, P.K.; Bell, B.; Giorgianni, F.; Caires, R.; Fernández-Peña, C.; Beranova, S.; Aihara, E.; Montrose, M.H.; Chaib, M.; et al. TRPV6 channel mediates alcohol-induced gut barrier dysfunction and systemic response. Cell Rep. 2022, 39, 110937. [Google Scholar] [CrossRef]
- Munjuluri, S.; Wilkerson, D.A.; Sooch, G.; Chen, X.; White, F.A.; Obukhov, A.G. Capsaicin and TRPV1 Channels in the Cardiovascular System: The Role of Inflammation. Cells 2021, 11, 18. [Google Scholar] [CrossRef]
- Chen, J.; Sun, W.; Zhu, Y.; Zhao, F.; Deng, S.; Tian, M.; Wang, Y.; Gong, Y. TRPV1: The key bridge in neuroimmune interactions. J. Intensive Med. 2024, 4, 442–452. [Google Scholar] [CrossRef]
- Devesa, I.; Planells-Cases, R.; Fernández-Ballester, G.; González-Ros, J.M.; Ferrer-Montiel, A.; Fernández-Carvajal, A. Role of the transient receptor potential vanilloid 1 in inflammation and sepsis. J. Inflamm. Res. 2011, 4, 67–81. [Google Scholar] [CrossRef]
- Dunne, O.M.; Martin, S.L.; Sergeant, G.P.; McAuley, D.F.; O’Kane, C.M.; Button, B.; McGarvey, L.P.; Lundy, F.T. TRPV2 modulates mechanically Induced ATP Release from Human bronchial epithelial cells. Respir. Res. 2024, 25, 188. [Google Scholar] [CrossRef] [PubMed]
- Dvornikova, K.A.; Platonova, O.N.; Bystrova, E.Y. The Role of TRP Channels in Sepsis and Colitis. Int. J. Mol. Sci. 2024, 25, 4784. [Google Scholar] [CrossRef] [PubMed]
- Pacífico, D.M.; Costa, D.; Lima Barbosa, M.L.; Rebouças, C.S.M.; Simonato, S.G.; Warren, C.A.; Morais, M.; Leitao, R.F.C.; Brito, G.A.C. TRPV4 modulates inflammatory responses and apoptosis in enteric glial cells triggered by Clostridioides difficile toxins A and B. J. Inflamm. 2025, 22, 3. [Google Scholar] [CrossRef] [PubMed]
- Thodeti, C.K.; Matthews, B.; Ravi, A.; Mammoto, A.; Ghosh, K.; Bracha, A.L.; Ingber, D.E. TRPV4 channels mediate cyclic strain-induced endothelial cell reorientation through integrin-to-integrin signaling. Circ. Res. 2009, 104, 1123–1130. [Google Scholar] [CrossRef]
- Liu, J.; Guo, Y.; Zhang, R.; Xu, Y.; Luo, C.; Wang, R.; Xu, S.; Wei, L. Inhibition of TRPV4 remodels single cell polarity and suppresses the metastasis of hepatocellular carcinoma. Cell Death Dis. 2023, 14, 379. [Google Scholar] [CrossRef]
- Jiang, D.; Guo, R.; Dai, R.; Knoedler, S.; Tao, J.; Machens, H.G.; Rinkevich, Y. The Multifaceted Functions of TRPV4 and Calcium Oscillations in Tissue Repair. Int. J. Mol. Sci. 2024, 25, 1179. [Google Scholar] [CrossRef]
- Miyamoto, T.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. TRPV3 regulates nitric oxide synthase-independent nitric oxide synthesis in the skin. Nat. Commun. 2011, 2, 369. [Google Scholar] [CrossRef]
- Aijima, R.; Wang, B.; Takao, T.; Mihara, H.; Kashio, M.; Ohsaki, Y.; Zhang, J.Q.; Mizuno, A.; Suzuki, M.; Yamashita, Y.; et al. The thermosensitive TRPV3 channel contributes to rapid wound healing in oral epithelia. FASEB J. 2015, 29, 182–192. [Google Scholar] [CrossRef]
- Ishii, T.; Uchida, K.; Hata, S.; Hatta, M.; Kita, T.; Miyake, Y.; Okamura, K.; Tamaoki, S.; Ishikawa, H.; Yamazaki, J. TRPV2 channel inhibitors attenuate fibroblast differentiation and contraction mediated by keratinocyte-derived TGF-β1 in an in vitro wound healing model of rats. J. Dermatol. Sci. 2018, 90, 332–342. [Google Scholar] [CrossRef]
- Varela-López, E.; Del Valle-Mondragón, L.; Castrejón-Téllez, V.; Pérez-Torres, I.; Arenas, A.P.; Rojas, F.M.; Guarner-Lans, V.; Vargas-González, A.; Pastelín-Hernández, G.; Torres-Narváez, J.C. Role of the Transient Receptor Potential Vanilloid Type 1 (TRPV1) in the Regulation of Nitric Oxide Release in Wistar Rat Aorta. Oxid. Med. Cell Longev. 2021, 2021, 8531975. [Google Scholar] [CrossRef]
- Zhu, Z.; Jiang, Y.; Li, Z.; Du, Y.; Chen, Q.; Guo, Q.; Ban, Y.; Gong, P. Sensory neuron transient receptor potential vanilloid-1 channel regulates angiogenesis through CGRP in vivo. Front. Bioeng. Biotechnol. 2024, 12, 1338504. [Google Scholar] [CrossRef]
- Peles, S.; Medda, B.K.; Zhang, Z.; Banerjee, B.; Lehmann, A.; Shaker, R.; Sengupta, J.N. Differential effects of transient receptor vanilloid one (TRPV1) antagonists in acid-induced excitation of esophageal vagal afferent fibers of rats. Neuroscience 2009, 161, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Stokłosa, P.; Borgström, A.; Kappel, S.; Peinelt, C. TRP Channels in Digestive Tract Cancers. Int. J. Mol. Sci. 2020, 21, 1877. [Google Scholar] [CrossRef] [PubMed]
- Marini, M.; Titiz, M.; Souza Monteiro de Araújo, D.; Geppetti, P.; Nassini, R.; De Logu, F. TRP Channels in Cancer: Signaling Mechanisms and Translational Approaches. Biomolecules 2023, 13, 1557. [Google Scholar] [CrossRef] [PubMed]
- Kärki, T.; Tojkander, S. TRPV Protein Family-From Mechanosensing to Cancer Invasion. Biomolecules 2021, 11, 1019. [Google Scholar] [CrossRef]
- Marincsák, R.; Tóth, B.I.; Czifra, G.; Márton, I.; Rédl, P.; Tar, I.; Tóth, L.; Kovács, L.; Bíró, T. Increased expression of TRPV1 in squamous cell carcinoma of the human tongue. Oral. Dis. 2009, 15, 328–335. [Google Scholar] [CrossRef]
- Gonzales, C.B.; Kirma, N.B.; De La Chapa, J.J.; Chen, R.; Henry, M.A.; Luo, S.; Hargreaves, K.M. Vanilloids induce oral cancer apoptosis independent of TRPV1. Oral. Oncol. 2014, 50, 437–447. [Google Scholar] [CrossRef]
- NavaneethaKrishnan, S.; Rosales, J.L.; Lee, K.Y. ROS-Mediated Cancer Cell Killing through Dietary Phytochemicals. Oxid. Med. Cell Longev. 2019, 2019, 9051542. [Google Scholar] [CrossRef]
- Sakakibara, A.; Sakakibara, S.; Kusumoto, J.; Takeda, D.; Hasegawa, T.; Akashi, M.; Minamikawa, T.; Hashikawa, K.; Terashi, H.; Komori, T. Upregulated Expression of Transient Receptor Potential Cation Channel Subfamily V Receptors in Mucosae of Patients with Oral Squamous Cell Carcinoma and Patients with a History of Alcohol Consumption or Smoking. PLoS ONE 2017, 12, e0169723. [Google Scholar] [CrossRef]
- Waning, J.; Vriens, J.; Owsianik, G.; Stüwe, L.; Mally, S.; Fabian, A.; Frippiat, C.; Nilius, B.; Schwab, A. A novel function of capsaicin-sensitive TRPV1 channels: Involvement in cell migration. Cell Calcium 2007, 42, 17–25. [Google Scholar] [CrossRef]
- Fujii, S.; Tajiri, Y.; Hasegawa, K.; Matsumoto, S.; Yoshimoto, R.U.; Wada, H.; Kishida, S.; Kido, M.A.; Yoshikawa, H.; Ozeki, S.; et al. The TRPV4-AKT axis promotes oral squamous cell carcinoma cell proliferation via CaMKII activation. Lab. Investig. 2020, 100, 311–323. [Google Scholar] [CrossRef]
- Huang, R.; Wang, F.; Yang, Y.; Ma, W.; Lin, Z.; Cheng, N.; Long, Y.; Deng, S.; Li, Z. Recurrent activations of transient receptor potential vanilloid-1 and vanilloid-4 promote cellular proliferation and migration in esophageal squamous cell carcinoma cells. FEBS Open Bio 2019, 9, 206–225. [Google Scholar] [CrossRef]
- Valenzano, K.J.; Grant, E.R.; Wu, G.; Hachicha, M.; Schmid, L.; Tafesse, L.; Sun, Q.; Rotshteyn, Y.; Francis, J.; Limberis, J.; et al. N-(4-tertiarybutylphenyl)-4-(3-chloropyridin-2-yl)tetrahydropyrazine -1(2H)-carbox-amide (BCTC), a novel, orally effective vanilloid receptor 1 antagonist with analgesic properties: I. in vitro characterization and pharmacokinetic properties. J. Pharmacol. Exp. Ther. 2003, 306, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Gunthorpe, M.J.; Hannan, S.L.; Smart, D.; Jerman, J.C.; Arpino, S.; Smith, G.D.; Brough, S.; Wright, J.; Egerton, J.; Lappin, S.C.; et al. Characterization of SB-705498, a potent and selective vanilloid receptor-1 (VR1/TRPV1) antagonist that inhibits the capsaicin-, acid-, and heat-mediated activation of the receptor. J. Pharmacol. Exp. Ther. 2007, 321, 1183–1192. [Google Scholar] [CrossRef] [PubMed]
- Rami, H.K.; Thompson, M.; Stemp, G.; Fell, S.; Jerman, J.C.; Stevens, A.J.; Smart, D.; Sargent, B.; Sanderson, D.; Randall, A.D.; et al. Discovery of SB-705498: A potent, selective and orally bioavailable TRPV1 antagonist suitable for clinical development. Bioorg Med. Chem. Lett. 2006, 16, 3287–3291. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Zhang, S.S.; Yan, Y.; Zhao, S. Overexpression of transient receptor potential vanilloid 2 is associated with poor prognosis in patients with esophageal squamous cell carcinoma. Med. Oncol. 2014, 31, 17. [Google Scholar] [CrossRef]
- Kudou, M.; Shiozaki, A.; Yamazato, Y.; Katsurahara, K.; Kosuga, T.; Shoda, K.; Arita, T.; Konishi, H.; Komatsu, S.; Kubota, T.; et al. The expression and role of TRPV2 in esophageal squamous cell carcinoma. Sci. Rep. 2019, 9, 16055. [Google Scholar] [CrossRef]
- Shiozaki, A.; Kudou, M.; Ichikawa, D.; Fujiwara, H.; Shimizu, H.; Ishimoto, T.; Arita, T.; Kosuga, T.; Konishi, H.; Komatsu, S.; et al. Esophageal cancer stem cells are suppressed by tranilast, a TRPV2 channel inhibitor. J. Gastroenterol. 2018, 53, 197–207. [Google Scholar] [CrossRef]
- Huang, R.; Li, S.; Tian, C.; Zhou, P.; Zhao, H.; Xie, W.; Xiao, J.; Wang, L.; Habimana, J.D.; Lin, Z.; et al. Thermal stress involved in TRPV2 promotes tumorigenesis through the pathways of HSP70/27 and PI3K/Akt/mTOR in esophageal squamous cell carcinoma. Br. J. Cancer 2022, 127, 1424–1439. [Google Scholar] [CrossRef]
- Gao, N.; Yang, F.; Chen, S.; Wan, H.; Zhao, X.; Dong, H. The role of TRPV1 ion channels in the suppression of gastric cancer development. J. Exp. Clin. Cancer Res. 2020, 39, 206. [Google Scholar] [CrossRef] [PubMed]
- Zoppoli, P.; Calice, G.; Laurino, S.; Ruggieri, V.; La Rocca, F.; La Torre, G.; Ciuffi, M.; Amendola, E.; De Vita, F.; Petrillo, A.; et al. TRPV2 Calcium Channel Gene Expression and Outcomes in Gastric Cancer Patients: A Clinically Relevant Association. J. Clin. Med. 2019, 8, 662. [Google Scholar] [CrossRef] [PubMed]
- Xie, R.; Xu, J.; Xiao, Y.; Wu, J.; Wan, H.; Tang, B.; Liu, J.; Fan, Y.; Wang, S.; Wu, Y.; et al. Calcium Promotes Human Gastric Cancer via a Novel Coupling of Calcium-Sensing Receptor and TRPV4 Channel. Cancer Res. 2017, 77, 6499–6512. [Google Scholar] [CrossRef] [PubMed]
- de Jong, P.R.; Takahashi, N.; Harris, A.R.; Lee, J.; Bertin, S.; Jeffries, J.; Jung, M.; Duong, J.; Triano, A.I.; Lee, J.; et al. Ion channel TRPV1-dependent activation of PTP1B suppresses EGFR-associated intestinal tumorigenesis. J. Clin. Investig. 2014, 124, 3793–3806. [Google Scholar] [CrossRef]
- de Jong, P.R.; Bertin, S.; Raz, E. TRPV1: Turning up the heat on intestinal tumorigenesis. Mol. Cell Oncol. 2015, 2, e975619. [Google Scholar] [CrossRef][Green Version]
- Du, Y.; Karatekin, F.; Wang, W.K.; Hong, W.; Boopathy, G.T.K. Cracking the EGFR code: Cancer biology, resistance mechanisms, and future therapeutic frontiers. Pharmacol. Rev. 2025, 77, 100076. [Google Scholar] [CrossRef]
- Dumitru, C.S.; Raica, M. Vascular Endothelial Growth Factor Family and Head and Neck Squamous Cell Carcinoma. Anticancer. Res. 2023, 43, 4315–4326. [Google Scholar] [CrossRef]
- Yang, Y.; Wen, W.; Chen, F.L.; Zhang, Y.J.; Liu, X.C.; Yang, X.Y.; Hu, S.S.; Jiang, Y.; Yuan, J. Expression and significance of pigment epithelium-derived factor and vascular endothelial growth factor in colorectal adenoma and cancer. World J. Gastrointest. Oncol. 2024, 16, 670–686. [Google Scholar] [CrossRef]
- Rizopoulos, T.; Assimakopoulou, M. Transient receptor potential (TRP) channels in human colorectal cancer: Evidence and perspectives. Histol. Histopathol. 2021, 36, 515–526. [Google Scholar] [CrossRef]
- Rindi, G.; Klöppel, G. Endocrine tumors of the gut and pancreas tumor biology and classification. Neuroendocrinology 2004, 80 (Suppl. S1), 12–15. [Google Scholar] [CrossRef]
- Islam, M.S. Molecular Regulations and Functions of the Transient Receptor Potential Channels of the Islets of Langerhans and Insulinoma Cells. Cells 2020, 9, 685. [Google Scholar] [CrossRef]
- Mergler, S.; Skrzypski, M.; Sassek, M.; Pietrzak, P.; Pucci, C.; Wiedenmann, B.; Strowski, M.Z. Thermo-sensitive transient receptor potential vanilloid channel-1 regulates intracellular calcium and triggers chromogranin A secretion in pancreatic neuroendocrine BON-1 tumor cells. Cell Signal 2012, 24, 233–246. [Google Scholar] [CrossRef]
- Skrzypski, M.; Sassek, M.; Abdelmessih, S.; Mergler, S.; Grötzinger, C.; Metzke, D.; Wojciechowicz, T.; Nowak, K.W.; Strowski, M.Z. Capsaicin induces cytotoxicity in pancreatic neuroendocrine tumor cells via mitochondrial action. Cell Signal 2014, 26, 41–48. [Google Scholar] [CrossRef]
- Menon, G.; Pandit, S.; Annamaraju, P.; Bhusal, K. Carcinoid Syndrome. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2025. [Google Scholar]
- Doihara, H.; Nozawa, K.; Kojima, R.; Kawabata-Shoda, E.; Yokoyama, T.; Ito, H. QGP-1 cells release 5-HT via TRPA1 activation; a model of human enterochromaffin cells. Mol. Cell Biochem. 2009, 331, 239–245. [Google Scholar] [CrossRef]
- Nozawa, K.; Kawabata-Shoda, E.; Doihara, H.; Kojima, R.; Okada, H.; Mochizuki, S.; Sano, Y.; Inamura, K.; Matsushime, H.; Koizumi, T.; et al. TRPA1 regulates gastrointestinal motility through serotonin release from enterochromaffin cells. Proc. Natl. Acad. Sci. USA 2009, 106, 3408–3413. [Google Scholar] [CrossRef] [PubMed]
- Chaigne, S.; Barbeau, S.; Ducret, T.; Guinamard, R.; Benoist, D. Pathophysiological Roles of the TRPV4 Channel in the Heart. Cells 2023, 12, 1654. [Google Scholar] [CrossRef] [PubMed]
- Filosa, J.A.; Yao, X.; Rath, G. TRPV4 and the regulation of vascular tone. J. Cardiovasc. Pharmacol. 2013, 61, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J. Molecular mechanism of TRP channels. Compr. Physiol. 2013, 3, 221–242. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Li, J.; Zhang, H.R.; Bai, S.W.; Yang, H.Y.; Shen, B.; Du, J.; Xia, X.M. The effect of transient receptor potential vanilloid 4 on the intestinal epithelial barrier and human colonic cells was affected by tyrosine-phosphorylated claudin-7. Biomed. Pharmacother. 2020, 122, 109697. [Google Scholar] [CrossRef]
- Khairatkar-Joshi, N.; Szallasi, A. TRPV1 antagonists: The challenges for therapeutic targeting. Trends Mol. Med. 2009, 15, 14–22. [Google Scholar] [CrossRef]
- Gavva, N.R.; Treanor, J.J.; Garami, A.; Fang, L.; Surapaneni, S.; Akrami, A.; Alvarez, F.; Bak, A.; Darling, M.; Gore, A.; et al. Pharmacological blockade of the vanilloid receptor TRPV1 elicits marked hyperthermia in humans. Pain 2008, 136, 202–210. [Google Scholar] [CrossRef]
- Chizh, B.A.; O’Donnell, M.B.; Napolitano, A.; Wang, J.; Brooke, A.C.; Aylott, M.C.; Bullman, J.N.; Gray, E.J.; Lai, R.Y.; Williams, P.M.; et al. The effects of the TRPV1 antagonist SB-705498 on TRPV1 receptor-mediated activity and inflammatory hyperalgesia in humans. Pain 2007, 132, 132–141. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tluli, O.; Arabi, A.; Rajha, H.E.; Abugharbieh, L.; Al Zeir, F.; Hamdan, M.; Abuqaba, A.; Boudaka, A. Role of Transient Receptor Potential Vanilloid Channels in Gastrointestinal Physiology and Pathology. Receptors 2025, 4, 24. https://doi.org/10.3390/receptors4040024
Tluli O, Arabi A, Rajha HE, Abugharbieh L, Al Zeir F, Hamdan M, Abuqaba A, Boudaka A. Role of Transient Receptor Potential Vanilloid Channels in Gastrointestinal Physiology and Pathology. Receptors. 2025; 4(4):24. https://doi.org/10.3390/receptors4040024
Chicago/Turabian StyleTluli, Omar, Ahmed Arabi, Humam Emad Rajha, Lana Abugharbieh, Faissal Al Zeir, Maryam Hamdan, Ayeda Abuqaba, and Ammar Boudaka. 2025. "Role of Transient Receptor Potential Vanilloid Channels in Gastrointestinal Physiology and Pathology" Receptors 4, no. 4: 24. https://doi.org/10.3390/receptors4040024
APA StyleTluli, O., Arabi, A., Rajha, H. E., Abugharbieh, L., Al Zeir, F., Hamdan, M., Abuqaba, A., & Boudaka, A. (2025). Role of Transient Receptor Potential Vanilloid Channels in Gastrointestinal Physiology and Pathology. Receptors, 4(4), 24. https://doi.org/10.3390/receptors4040024