
Maize Breeding for Low Nitrogen Inputs in Agriculture: Mechanisms Underlying the Tolerance to the Abiotic Stress




Abstract
1. Introduction
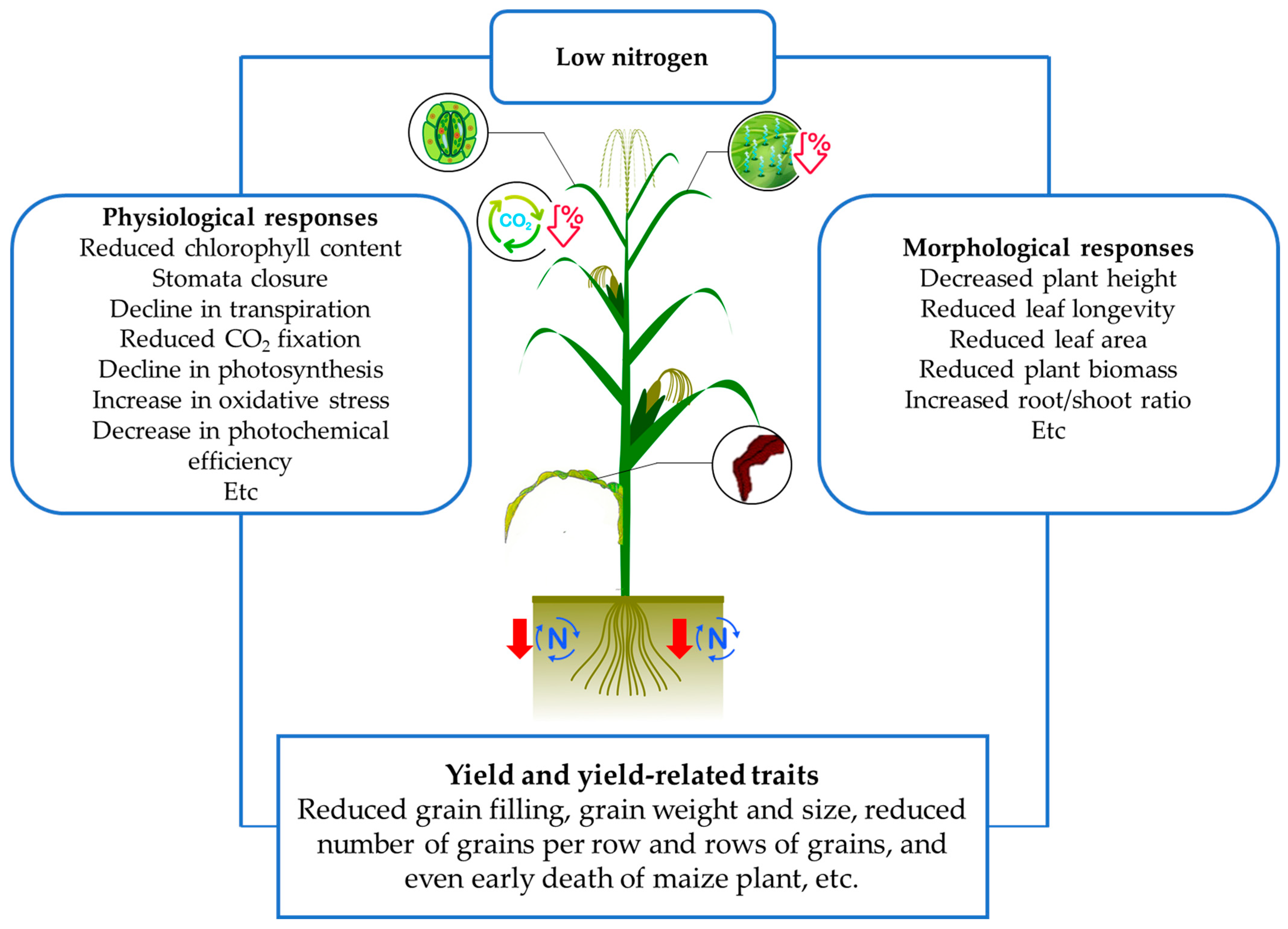
2. The Physiological and Morphological Shoot Responses of Maize under Low Nitrogen Conditions
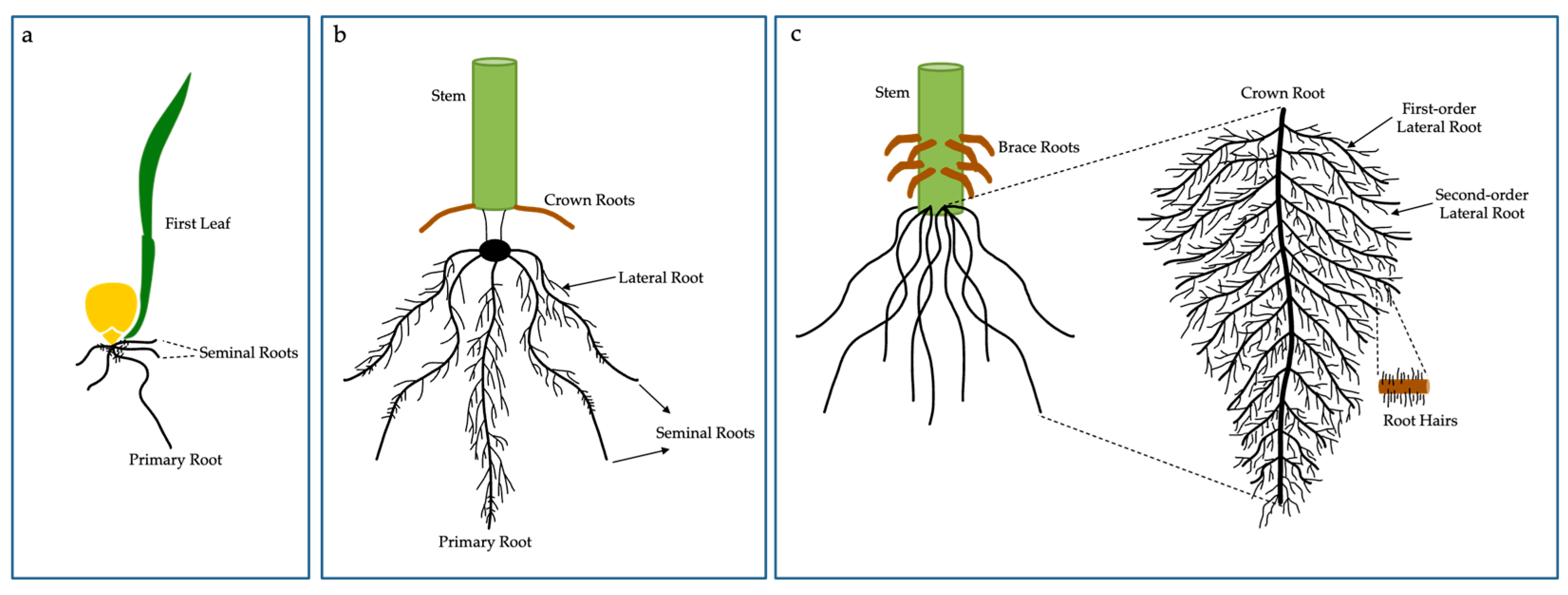
3. The Root Adaptation of Maize under Low N Conditions
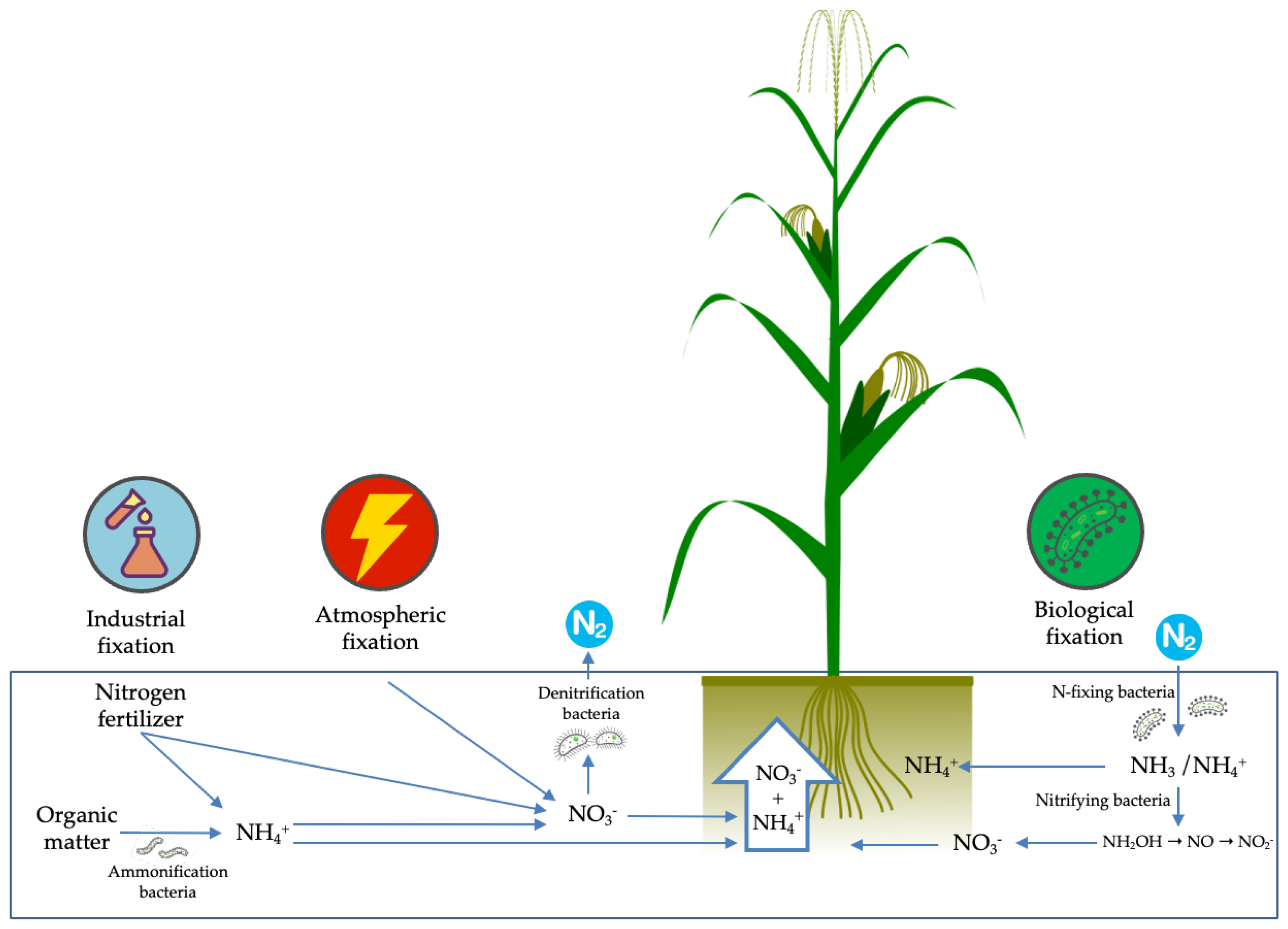
4. The Molecular Mechanisms Involved in Nitrogen-Use Efficiency in Maize
4.1. Nitrogen Uptake and Transport
4.2. Nitrogen Reduction and Assimilation
4.3. Translocation and Remobilization of Nitrogen
5. Maize Improvement for Low N Conditions
The Conventional Breeding Approach and the Genetic Basis of Nitrogen-Use Efficiency under Conditions of Limited N in Soil
6. Conclusions and the Way Forward
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, X.; Davidson, E.A.; Mauzerall, D.L.; Searchinger, T.D.; Dumas, P.; Shen, Y. Managing Nitrogen for Sustainable Development. Nature 2015, 528, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Almeida, V.C.; Viana, J.M.S.; Risso, L.A.; Ribeiro, C.; DeLima, R.O. Generation Mean Analysis for Nitrogen and Phosphorus Uptake, Utilization, and Translocation Indexes at Vegetative Stage in Tropical Popcorn. Euphytica 2018, 214, 103. [Google Scholar] [CrossRef]
- Tilman, D.; Reich, P.B.; Knops, J.; Wedin, D.; Mielke, T.; Lehman, C. Diversity and Productivity in a Long-Term Grassland Experiment. Science 2001, 294, 843–845. [Google Scholar] [CrossRef] [PubMed]
- Mascia, M.; Sega, D.; Zamboni, A.; Varanini, Z. Nitrogen Starvation Differentially Influences Transcriptional and Uptake Rate Profiles in Roots of Two Maize Inbred Lines with Different NUE. Int. J. Mol. Sci. 2019, 20, 4856. [Google Scholar] [CrossRef]
- Khan, S.; Amaral Júnior, A.T.d.; Ferreira, F.R.A.; Kamphorst, S.H.; Gonçalves, G.M.B.; Simone Mendonça Freitas, M.; Silveira, V.; Apolinário de Souza Filho, G.; Francisco Teixeira do Amaral, J.; Enrique Bresssan Smith, R.; et al. Limited Nitrogen and Plant Growth Stages Discriminate Well Nitrogen Use, Uptake and Utilization Efficiency in Popcorn. Plants 2020, 9, 893. [Google Scholar] [CrossRef]
- Ren, B.; Guo, Y.; Liu, P.; Zhao, B.; Zhang, J. Effects of Urea-Ammonium Nitrate Solution on Yield, N2O Emission, and Nitrogen Efficiency of Summer Maize Under Integration of Water and Fertilizer. Front. Plant Sci. 2021, 12, 700331. [Google Scholar] [CrossRef]
- Wang, Z.; Ma, B.-L.; Yu, X.; Gao, J.; Sun, J.; Su, Z.; Yu, S. Physiological Basis of Heterosis for Nitrogen Use Efficiency of Maize. Sci. Rep. 2019, 9, 18708. [Google Scholar] [CrossRef] [PubMed]
- Andrews, M.; Lea, P.J. Our Nitrogen ‘Footprint’: The Need for Increased Crop Nitrogen Use Efficiency. Ann. Appl. Biol. 2013, 163, 165–169. [Google Scholar] [CrossRef]
- USDA Agricultural Baseline Database. Available online: https://www.ers.usda.gov/data-products/agricultural-baseline-database (accessed on 4 January 2023).
- Heffer, P.; Gruère, A.; Roberts, T. Assessment of Fertilizer Use by Crop at the Global Level; International Fertilizer Industry Association: Paris, France, 2017. [Google Scholar]
- Sheoran, S.; Kumar, S.; Kumar, P.; Meena, R.S.; Rakshit, S. Nitrogen Fixation in Maize: Breeding Opportunities. Theor. Appl. Genet. 2021, 134, 1263–1280. [Google Scholar] [CrossRef]
- Han, M.; Wong, J.; Su, T.; Beatty, P.H.; Good, A.G. Identification of Nitrogen Use Efficiency Genes in Barley: Searching for QTLs Controlling Complex Physiological Traits. Front. Plant Sci. 2016, 7, 1587. [Google Scholar] [CrossRef]
- Pampana, S.; Mariotti, M. Durum Wheat Yield and N Uptake as Affected by N Source, Timing, and Rate in Two Mediterranean Environments. Agronomy 2021, 11, 1299. [Google Scholar] [CrossRef]
- Hawkesford, M.J. Genetic Variation in Traits for Nitrogen Use Efficiency in Wheat. J. Exp. Bot. 2017, 68, 2627–2632. [Google Scholar] [CrossRef] [PubMed]
- Hawkesford, M.J.; Griffiths, S. Exploiting Genetic Variation in Nitrogen Use Efficiency for Cereal Crop Improvement. Curr. Opin. Plant Biol. 2019, 49, 35–42. [Google Scholar] [CrossRef]
- Congreves, K.A.; Otchere, O.; Ferland, D.; Farzadfar, S.; Williams, S.; Arcand, M.M. Nitrogen Use Efficiency Definitions of Today and Tomorrow. Front. Plant Sci. 2021, 12, 637108. [Google Scholar] [CrossRef]
- Quan, X.; Zeng, J.; Chen, G.; Zhang, G. Transcriptomic Analysis Reveals Adaptive Strategies to Chronic Low Nitrogen in Tibetan Wild Barley. BMC Plant. Biol. 2019, 19, 68. [Google Scholar] [CrossRef]
- He, X.; Ma, H.; Zhao, X.; Nie, S.; Li, Y.; Zhang, Z.; Shen, Y.; Chen, Q.; Lu, Y.; Lan, H.; et al. Comparative RNA-Seq Analysis Reveals That Regulatory Network of Maize Root Development Controls the Expression of Genes in Response to N Stress. PLoS ONE 2016, 11, e0151697. [Google Scholar] [CrossRef]
- Kumari, S.; Sharma, N.; Raghuram, N. Meta-Analysis of Yield-Related and N-Responsive Genes Reveals Chromosomal Hotspots, Key Processes and Candidate Genes for Nitrogen-Use Efficiency in Rice. Front. Plant Sci. 2021, 12, 627955. [Google Scholar] [CrossRef] [PubMed]
- Gan, H.; Jiao, Y.; Jia, J.; Wang, X.; Li, H.; Shi, W.; Peng, C.; Polle, A.; Luo, Z.-B. Phosphorus and Nitrogen Physiology of Two Contrasting Poplar Genotypes When Exposed to Phosphorus and/or Nitrogen Starvation. Tree Physiol. 2016, 36, 22–38. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 6th ed.; Sinauer Associates: Sunderland, MA, USA, 2016. [Google Scholar]
- Mu, X.; Chen, Y. The Physiological Response of Photosynthesis to Nitrogen Deficiency. Plant Physiol. Biochem. 2021, 158, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Urban, A.; Rogowski, P.; Wasilewska-Dębowska, W.; Romanowska, E. Understanding Maize Response to Nitrogen Limitation in Different Light Conditions for the Improvement of Photosynthesis. Plants 2021, 10, 1932. [Google Scholar] [CrossRef] [PubMed]
- Buchert, F.; Scholz, M.; Hippler, M. Electron Transfer via Cytochrome b 6 f Complex Displays Sensitivity to Antimycin A upon STT7 Kinase Activation. Biochem. J. 2022, 479, 111–127. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, L.; Liu, J. Proteomic Analysis of Hydrogen Production in Chlorella pyrenoidosa under Nitrogen Deprivation. Algal Res. 2021, 53, 102143. [Google Scholar] [CrossRef]
- Thi Nong, H.; Tateishi, R.; Suriyasak, C.; Kobayashi, T.; Oyama, Y.; Chen, W.J.; Matsumoto, R.; Hamaoka, N.; Iwaya-Inoue, M.; Ishibashi, Y. Effect of Seedling Nitrogen Condition on Subsequent Vegetative Growth Stages and Its Relationship to the Expression of Nitrogen Transporter Genes in Rice. Plants 2020, 9, 861. [Google Scholar] [CrossRef]
- Sanagi, M.; Aoyama, S.; Kubo, A.; Lu, Y.; Sato, Y.; Ito, S.; Abe, M.; Mitsuda, N.; Ohme-Takagi, M.; Kiba, T.; et al. Low Nitrogen Conditions Accelerate Flowering by Modulating the Phosphorylation State of FLOWERING BHLH 4 in Arabidopsis. Proc. Natl. Acad. Sci. USA 2021, 118, e2022942118. [Google Scholar] [CrossRef]
- Liu, H.; Able, A.J.; Able, J.A. Nitrogen Starvation-Responsive MicroRNAs Are Affected by Transgenerational Stress in Durum Wheat Seedlings. Plants 2021, 10, 826. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.d.; Amaral Júnior, A.T.d.; Fritsche-Neto, R.; Kamphorst, S.H.; Ferreira, F.R.A.; Amaral, J.F.T.d.; Vivas, J.M.S.; Santos, P.H.A.D.; Lima, V.J.d.; Khan, S.; et al. Relative Importance of Gene Effects for Nitrogen-Use Efficiency in Popcorn. PLoS ONE 2019, 14, e0222726. [Google Scholar] [CrossRef]
- Hammad, H.M.; Chawla, M.S.; Jawad, R.; Alhuqail, A.; Bakhat, H.F.; Farhad, W.; Khan, F.; Mubeen, M.; Shah, A.N.; Liu, K.; et al. Evaluating the Impact of Nitrogen Application on Growth and Productivity of Maize Under Control Conditions. Front. Plant Sci. 2022, 13, 885479. [Google Scholar] [CrossRef]
- Abubakar, A.W.; Manga, A.A.; Kamara, A.Y.; Tofa, A.I. Physiological Evaluations of Maize Hybrids under Low Nitrogen. Adv. Agric. 2019, 2019, 2624707. [Google Scholar] [CrossRef]
- D’Andrea, K.E.; Parco, M.; Maddonni, G.Á. Maize Prolificacy under Contrasting Plant Densities and N Supplies: II. Growth per Plant, Biomass Partitioning to Apical and Sub-Apical Ears during the Critical Period and Kernel Setting. Field Crops Res. 2022, 284, 108557. [Google Scholar] [CrossRef]
- Ludemann, C.I.; Hijbeek, R.; van Loon, M.P.; Murrell, T.S.; Dobermann, A.; van Ittersum, M.K. Estimating Maize Harvest Index and Nitrogen Concentrations in Grain and Residue Using Globally Available Data. Field Crops Res. 2022, 284, 108578. [Google Scholar] [CrossRef]
- Liu, S.; Qin, F. Genetic Dissection of Maize Drought Tolerance for Trait Improvement. Mol. Breed. 2021, 41, 8. [Google Scholar] [CrossRef]
- Makino, A.; Shimada, T.; Takumi, S.; Kaneko, K.; Matsuoka, M.; Shimamoto, K.; Nakano, H.; Miyao-Tokutomi, M.; Mae, T.; Yamamoto, N. Does Decrease in Ribulose-1,5-Bisphosphate Carboxylase by Antisense RbcS Lead to a Higher N-Use Efficiency of Photosynthesis under Conditions of Saturating CO2 and Light in Rice Plants? Plant Physiol. 1997, 114, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Chen, Q.; Chen, F.; Yuan, L.; Mi, G. Within-Leaf Nitrogen Allocation in Adaptation to Low Nitrogen Supply in Maize during Grain-Filling Stage. Front. Plant Sci. 2016, 7, 699. [Google Scholar] [CrossRef] [PubMed]
- Khamis, S.; Lamaze, T.; Lemoine, Y.; Foyer, C. Adaptation of the Photosynthetic Apparatus in Maize Leaves as a Result of Nitrogen Limitation. Plant Physiol. 1990, 94, 1436–1443. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Zhang, J.; Zhang, Q.; Li, L.; Kuang, T. Modification of Photosystem II Photochemistry in Nitrogen Deficient Maize and Wheat Plants. J. Plant Physiol. 2001, 158, 1423–1430. [Google Scholar] [CrossRef]
- Fu, W.; Wang, Y.; Ye, Y.; Zhen, S.; Zhou, B.; Wang, Y.; Hu, Y.; Zhao, Y.; Huang, Y. Grain Yields and Nitrogen Use Efficiencies in Different Types of Stay-Green Maize in Response to Nitrogen Fertilizer. Plants 2020, 9, 474. [Google Scholar] [CrossRef]
- Lee, E.A.; Tollenaar, M. Physiological Basis of Successful Breeding Strategies for Maize Grain Yield. Crop. Sci. 2007, 47, S-202–S-215. [Google Scholar] [CrossRef]
- Hirel, B.; le Gouis, J.; Ney, B.; Gallais, A. The Challenge of Improving Nitrogen Use Efficiency in Crop Plants: Towards a More Central Role for Genetic Variability and Quantitative Genetics within Integrated Approaches. J. Exp. Bot. 2007, 58, 2369–2387. [Google Scholar] [CrossRef]
- Chen, Y.; Xiao, C.; Chen, X.; Li, Q.; Zhang, J.; Chen, F.; Yuan, L.; Mi, G. Characterization of the Plant Traits Contributed to High Grain Yield and High Grain Nitrogen Concentration in Maize. Field Crops Res. 2014, 159, 1–9. [Google Scholar] [CrossRef]
- Mu, X.; Chen, Q.; Chen, F.; Yuan, L.; Mi, G. Dynamic Remobilization of Leaf Nitrogen Components in Relation to Photosynthetic Rate during Grain Filling in Maize. Plant Physiol. Biochem. 2018, 129, 27–34. [Google Scholar] [CrossRef]
- Nasielski, J.; Earl, H.; Deen, B. Luxury Vegetative Nitrogen Uptake in Maize Buffers Grain Yield Under Post-Silking Water and Nitrogen Stress: A Mechanistic Understanding. Front. Plant Sci. 2019, 10, 318. [Google Scholar] [CrossRef]
- Plénet, D.; Lemaire, G. Relationships between Dynamics of Nitrogen Uptake and Dry Matter Accumulation in Maize Crops. Determination of Critical N Concentration. Plant Soil 1999, 216, 65–82. [Google Scholar] [CrossRef]
- Ciampitti, I.A.; Vyn, T.J. Physiological Perspectives of Changes over Time in Maize Yield Dependency on Nitrogen Uptake and Associated Nitrogen Efficiencies: A Review. Field Crops Res. 2012, 133, 48–67. [Google Scholar] [CrossRef]
- Paponov, I.A.; Engels, C. Effect of Nitrogen Supply on Carbon and Nitrogen Partitioning after Flowering in Maize. J. Plant Nutr. Soil Sci. 2005, 168, 447–453. [Google Scholar] [CrossRef]
- De Oliveira Silva, A.; Camberato, J.J.; Coram, T.; Filley, T.; Vyn, T.J. Applicability of a “Multi-Stage Pulse Labeling” 15N Approach to Phenotype N Dynamics in Maize Plant Components during the Growing Season. Front. Plant Sci. 2017, 8, 1360. [Google Scholar] [CrossRef]
- Gao, K.; Chen, F.; Yuan, L.; Zhang, F.; Mi, G. A Comprehensive Analysis of Root Morphological Changes and Nitrogen Allocation in Maize in Response to Low Nitrogen Stress. Plant Cell Environ. 2015, 38, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Schneider, H.M.; Yang, J.T.; Brown, K.M.; Lynch, J.P. Nodal Root Diameter and Node Number in Maize (Zea mays L.) Interact to Influence Plant Growth under Nitrogen Stress. Plant Direct. 2021, 5, e00310. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.P. Root Phenes That Reduce the Metabolic Costs of Soil Exploration: Opportunities for 21st Century Agriculture. Plant Cell Environ. 2015, 38, 1775–1784. [Google Scholar] [CrossRef] [PubMed]
- Hochholdinger, F.; Woll, K.; Sauer, M.; Dembinsky, D. Genetic Dissection of Root Formation in Maize (Zea mays) Reveals Root-Type Specific Developmental Programmes. Ann. Bot. 2004, 93, 359–368. [Google Scholar] [CrossRef]
- Lynch, J.P. Steep, Cheap and Deep: An Ideotype to Optimize Water and N Acquisition by Maize Root Systems. Ann. Bot. 2013, 112, 347–357. [Google Scholar] [CrossRef]
- Wang, Y.; Mi, G.; Chen, F.; Zhang, J.; Zhang, F. Response of Root Morphology to Nitrate Supply and Its Contribution to Nitrogen Accumulation in Maize. J. Plant. Nutr. 2005, 27, 2189–2202. [Google Scholar] [CrossRef]
- Trachsel, S.; Kaeppler, S.M.; Brown, K.M.; Lynch, J.P. Maize Root Growth Angles Become Steeper under Low N Conditions. Field Crops Res. 2013, 140, 18–31. [Google Scholar] [CrossRef]
- Burridge, J.D.; Findeis, J.L.; Jochua, C.N.; Miguel, M.A.; Mubichi-Kut, F.M.; Quinhentos, M.L.; Xerinda, S.A.; Lynch, J.P. A Case Study on the Efficacy of Root Phenotypic Selection for Edaphic Stress Tolerance in Low-Input Agriculture: Common Bean Breeding in Mozambique. Field Crops Res. 2019, 244, 107612. [Google Scholar] [CrossRef]
- Santos, D.L.; Coelho, E.F.; Cunha, F.F.d.; Donato, S.L.R.; Bernado, W.d.P.; Rodrigues, W.P.; Campostrini, E. Partial Root-Zone Drying in Field-Grown Papaya: Gas Exchange, Yield, and Water Use Efficiency. Agric. Water Manag. 2021, 243, 106421. [Google Scholar] [CrossRef]
- Santos, T.d.O.; Amaral Junior, A.T.d.; Bispo, R.B.; Lima, V.J.d.; Kamphorst, S.H.; Leite, J.T.; Santos Júnior, D.R.d.; Santos, P.H.A.D.; Oliveira, U.A.d.; Schmitt, K.F.M.; et al. Phenotyping Latin American Open-Pollinated Varieties of Popcorn for Environments with Low Water Availability. Plants 2021, 10, 1211. [Google Scholar] [CrossRef]
- Favela, A.; Bohn, M.O.; Kent, A.D. Maize Germplasm Chronosequence Shows Crop Breeding History Impacts Recruitment of the Rhizosphere Microbiome. ISME J. 2021, 15, 2454–2464. [Google Scholar] [CrossRef]
- Van Deynze, A.; Zamora, P.; Delaux, P.-M.; Heitmann, C.; Jayaraman, D.; Rajasekar, S.; Graham, D.; Maeda, J.; Gibson, D.; Schwartz, K.D.; et al. Nitrogen Fixation in a Landrace of Maize Is Supported by a Mucilage-Associated Diazotrophic Microbiota. PLoS Biol. 2018, 16, e2006352. [Google Scholar] [CrossRef]
- Jones, P.; Garcia, B.J.; Furches, A.; Tuskan, G.A.; Jacobson, D. Plant Host-Associated Mechanisms for Microbial Selection. Front. Plant Sci. 2019, 10, 862 . [Google Scholar] [CrossRef]
- Bais, H.P.; Park, S.-W.; Weir, T.L.; Callaway, R.M.; Vivanco, J.M. How Plants Communicate Using the Underground Information Superhighway. Trends Plant Sci. 2004, 9, 26–32. [Google Scholar] [CrossRef]
- Roesch, L.F.W.; Camargo, F.A.O.; Bento, F.M.; Triplett, E.W. Biodiversity of Diazotrophic Bacteria within the Soil, Root and Stem of Field-Grown Maize. Plant Soil 2008, 302, 91–104. [Google Scholar] [CrossRef]
- Santoyo, G. How Plants Recruit Their Microbiome? New Insights into Beneficial Interactions. J. Adv. Res. 2022, 40, 45–58. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; van der Putten, W.H. Going Back to the Roots: The Microbial Ecology of the Rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Bulgarelli, D.; Schlaeppi, K.; Spaepen, S.; van Themaat, E.V.L.; Schulze-Lefert, P. Structure and Functions of the Bacterial Microbiota of Plants. Annu. Rev. Plant Biol. 2013, 64, 807–838. [Google Scholar] [CrossRef] [PubMed]
- De Mello Prado, R. Nitrogen. In Mineral Nutrition of Tropical Plants; Springer International Publishing: Cham, Switzerland, 2021; pp. 69–98. [Google Scholar]
- Bi, Y.-M.; Meyer, A.; Downs, G.S.; Shi, X.; El-kereamy, A.; Lukens, L.; Rothstein, S.J. High Throughput RNA Sequencing of a Hybrid Maize and Its Parents Shows Different Mechanisms Responsive to Nitrogen Limitation. BMC Genom. 2014, 15, 77. [Google Scholar] [CrossRef]
- Cao, H.; Qi, S.; Sun, M.; Li, Z.; Yang, Y.; Crawford, N.M.; Wang, Y. Overexpression of the Maize ZmNLP6 and ZmNLP8 Can Complement the Arabidopsis Nitrate Regulatory Mutant Nlp7 by Restoring Nitrate Signaling and Assimilation. Front. Plant Sci. 2017, 8, 1703. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Turano, F.J. The Putative Glutamate Receptor 1.1 (AtGLR1.1) Functions as a Regulator of Carbon and Nitrogen Metabolism in Arabidopsis Thaliana. Proc. Natl. Acad. Sci. USA 2003, 100, 6872–6877. [Google Scholar] [CrossRef]
- Raddatz, N.; Morales de los Ríos, L.; Lindahl, M.; Quintero, F.J.; Pardo, J.M. Coordinated Transport of Nitrate, Potassium, and Sodium. Front. Plant Sci. 2020, 11, 247. [Google Scholar] [CrossRef] [PubMed]
- Zuluaga, D.L.; Sonnante, G. The Use of Nitrogen and Its Regulation in Cereals: Structural Genes, Transcription Factors, and the Role of MiRNAs. Plants 2019, 8, 294. [Google Scholar] [CrossRef] [PubMed]
- Simons, M.; Saha, R.; Guillard, L.; Clement, G.; Armengaud, P.; Canas, R.; Maranas, C.D.; Lea, P.J.; Hirel, B. Nitrogen-Use Efficiency in Maize (Zea mays L.): From “omics” Studies to Metabolic Modelling. J. Exp. Bot. 2014, 65, 5657–5671. [Google Scholar] [CrossRef]
- Plett, D.C.; Holtham, L.R.; Okamoto, M.; Garnett, T.P. Nitrate Uptake and Its Regulation in Relation to Improving Nitrogen Use Efficiency in Cereals. Semin. Cell Dev. Biol. 2018, 74, 97–104. [Google Scholar] [CrossRef]
- Engineer, C.B.; Kranz, R.G. Reciprocal Leaf and Root Expression of AtAmt1.1 and Root Architectural Changes in Response to Nitrogen Starvation. Plant Physiol. 2007, 143, 236–250. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; Dong, L.; Lü, W.; Lü, J.; Meng, Q.; Liu, P. Transcriptome Analysis of Maize Seedling Roots in Response to Nitrogen-, Phosphorus-, and Potassium Deficiency. Plant Soil 2020, 447, 637–658. [Google Scholar] [CrossRef]
- Vidal, E.A.; Moyano, T.C.; Krouk, G.; Katari, M.S.; Tanurdzic, M.; McCombie, W.R.; Coruzzi, G.M.; Gutiérrez, R.A. Integrated RNA-Seq and SRNA-Seq Analysis Identifies Novel Nitrate-Responsive Genes in Arabidopsis Thaliana Roots. BMC Genom. 2013, 14, 701. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Naz, M.; Fan, X.; Xuan, W.; Miller, A.J.; Xu, G. Plant Nitrate Transporters: From Gene Function to Application. J. Exp. Bot. 2017, 68, 2463–2475. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, Z.; Yang, C.; Yang, Z.; Li, H.; Wu, Y. Physiological Responses and Small RNAs Changes in Maize under Nitrogen Deficiency and Resupply. Genes Genom. 2019, 41, 1183–1194. [Google Scholar] [CrossRef]
- Wang, Y.; Yao, Q.; Zhang, Y.; Zhang, Y.; Xing, J.; Yang, B.; Mi, G.; Li, Z.; Zhang, M. The Role of Gibberellins in Regulation of Nitrogen Uptake and Physiological Traits in Maize Responding to Nitrogen Availability. Int. J. Mol. Sci. 2020, 21, 1824. [Google Scholar] [CrossRef]
- Dechorgnat, J.; Francis, K.L.; Dhugga, K.S.; Rafalski, J.A.; Tyerman, S.D.; Kaiser, B.N. Tissue and Nitrogen-Linked Expression Profiles of Ammonium and Nitrate Transporters in Maize. BMC Plant Biol. 2019, 19, 206. [Google Scholar] [CrossRef] [PubMed]
- Ohkubo, Y.; Kuwata, K.; Matsubayashi, Y. A Type 2C Protein Phosphatase Activates High-Affinity Nitrate Uptake by Dephosphorylating NRT2.1. Nat. Plants 2021, 7, 310–316. [Google Scholar] [CrossRef]
- Trevisan, S.; Borsa, P.; Botton, A.; Varotto, S.; Malagoli, M.; Ruperti, B.; Quaggiotti, S. Expression of Two Maize Putative Nitrate Transporters in Response to Nitrate and Sugar Availability. Plant Biol. 2008, 10, 462–475. [Google Scholar] [CrossRef]
- Sinha, S.; Sevanthi, V.A.; Chaudhary, S.; Tyagi, P.; Venkadesan, S.; Rani, M.; Mandal, P. Transcriptome Analysis of Two Rice Varieties Contrasting for Nitrogen Use Efficiency under Chronic N Starvation Reveals Differences in Chloroplast and Starch Metabolism-Related Genes. Genes 2018, 9, 206. [Google Scholar] [CrossRef]
- Goel, P.; Sharma, N.K.; Bhuria, M.; Sharma, V.; Chauhan, R.; Pathania, S.; Swarnkar, M.K.; Chawla, V.; Acharya, V.; Shankar, R.; et al. Transcriptome and Co-Expression Network Analyses Identify Key Genes Regulating Nitrogen Use Efficiency in Brassica juncea L. Sci. Rep. 2018, 8, 7451. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, N.; Liu, S.; Dong, A.; Zenda, T.; Liu, X.; Li, J.; Duan, H. Comparative Proteomic Analysis of Two Contrasting Maize Hybrids’ Responses to Low Nitrogen Stress at the Twelve Leaf Stage and Function Verification of ZmTGA Gene. Genes 2022, 13, 670. [Google Scholar] [CrossRef] [PubMed]
- Jakoby, M.; Weisshaar, B.; Dröge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F. BZIP Transcription Factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, J.M.; Riveras, E.; Vidal, E.A.; Gras, D.E.; Contreras-López, O.; Tamayo, K.P.; Aceituno, F.; Gómez, I.; Ruffel, S.; Lejay, L.; et al. Systems Approach Identifies TGA1 and TGA4 Transcription Factors as Important Regulatory Components of the Nitrate Response of Arabidopsis Thaliana Roots. Plant J. 2014, 80, 12618. [Google Scholar] [CrossRef]
- Zhong, L.; Chen, D.; Min, D.; Li, W.; Xu, Z.; Zhou, Y.; Li, L.; Chen, M.; Ma, Y. AtTGA4, a BZIP Transcription Factor, Confers Drought Resistance by Enhancing Nitrate Transport and Assimilation in Arabidopsis Thaliana. Biochem. Biophys. Res. Commun. 2015, 457, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Marschner, H. Marschner’s Mineral Nutrition of Higher Plants; Elsevier: Amsterdam, The Netherlands, 2012; ISBN 9780123849052. [Google Scholar]
- Taylor, A.R.; Bloom, A.J. Ammonium, Nitrate, and Proton Fluxes along the Maize Root. Plant Cell Environ. 1998, 21, 1255–1263. [Google Scholar] [CrossRef]
- Bloom, A.J.; Meyerhoff, P.A.; Taylor, A.R.; Rost, T.L. Root Development and Absorption of Ammonium and Nitrate from the Rhizosphere. J. Plant Growth Regul. 2002, 21, 416–431. [Google Scholar] [CrossRef]
- Gu, R.; Duan, F.; An, X.; Zhang, F.; von Wirén, N.; Yuan, L. Characterization of AMT-Mediated High-Affinity Ammonium Uptake in Roots of Maize (Zea mays L.). Plant Cell Physiol. 2013, 54, 1515–1524. [Google Scholar] [CrossRef]
- Zhao, S.; Li, C.-I.; Guo, Y.; Sheng, Q.; Shyr, Y. RnaSeqSampleSize: Real Data Based Sample Size Estimation for RNA Sequencing. BMC Bioinform. 2018, 19, 191. [Google Scholar] [CrossRef]
- Meyer, C.; Stitt, M. Nitrate Reduction and Signalling. In Plant Nitrogen; Springer: Berlin/Heidelberg, Germany, 2001; pp. 37–59. [Google Scholar]
- Wang, Y.-Y.; Cheng, Y.-H.; Chen, K.-E.; Tsay, Y.-F. Nitrate Transport, Signaling, and Use Efficiency. Annu. Rev. Plant Biol. 2018, 69, 85–122. [Google Scholar] [CrossRef]
- Masclaux-Daubresse, C.; Daniel-Vedele, F.; Dechorgnat, J.; Chardon, F.; Gaufichon, L.; Suzuki, A. Nitrogen Uptake, Assimilation and Remobilization in Plants: Challenges for Sustainable and Productive Agriculture. Ann. Bot. 2010, 105, 1141–1157. [Google Scholar] [CrossRef]
- Asibi, A.E.; Chai, Q.; Coulter, J.A. Mechanisms of Nitrogen Use in Maize. Agronomy 2019, 9, 775. [Google Scholar] [CrossRef]
- Sakakibara, H.; Kawabata, S.; Hase, T.; Sugiyama, T. Differential Effects of Nitrate and Light on the Expression of Glutamine Synthetases and Ferredoxin-Dependent Glutamate Synthase in Maize. Plant Cell Physiol. 1992, 33, 1193–1198. [Google Scholar] [CrossRef]
- Martin, A.; Lee, J.; Kichey, T.; Gerentes, D.; Zivy, M.; Tatout, C.; Dubois, F.; Balliau, T.; Valot, B.; Davanture, M.; et al. Two Cytosolic Glutamine Synthetase Isoforms of Maize Are Specifically Involved in the Control of Grain Production. Plant Cell 2006, 18, 3252–3274. [Google Scholar] [CrossRef]
- Liseron-Monfils, C.; Bi, Y.-M.; Downs, G.S.; Wu, W.; Signorelli, T.; Lu, G.; Chen, X.; Bondo, E.; Zhu, T.; Lukens, L.N.; et al. Nitrogen Transporter and Assimilation Genes Exhibit Developmental Stage-Selective Expression in Maize (Zea mays L.) Associated with Distinct Cis -Acting Promoter Motifs. Plant Signal. Behav. 2013, 8, e26056. [Google Scholar] [CrossRef]
- Suzuki, A.; Knaff, D.B. Glutamate Synthase: Structural, Mechanistic and Regulatory Properties, and Role in the Amino Acid Metabolism. Photosynth. Res. 2005, 83, 191–217. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, T.; Gaufichon, L.; Boutet-Mercey, S.; Christ, A.; Masclaux-Daubresse, C. Enzymatic and Metabolic Diagnostic of Nitrogen Deficiency in Arabidopsis thaliana Wassileskija Accession. Plant Cell Physiol. 2008, 49, 1056–1065. [Google Scholar] [CrossRef]
- Tabuchi, M.; Abiko, T.; Yamaya, T. Assimilation of Ammonium Ions and Reutilization of Nitrogen in Rice (Oryza sativa L.). J. Exp. Bot. 2007, 58, 2319–2327. [Google Scholar] [CrossRef]
- Uauy, C.; Distelfeld, A.; Fahima, T.; Blechl, A.; Dubcovsky, J. A NAC Gene Regulating Senescence Improves Grain Protein, Zinc, and Iron Content in Wheat. Science 2006, 314, 1298–1301. [Google Scholar] [CrossRef]
- Bänziger, M.; Betrán, F.J.; Lafitte, H.R. Efficiency of High-Nitrogen Selection Environments for Improving Maize for Low-Nitrogen Target Environments. Crop Sci. 1997, 37, 1103–1109. [Google Scholar] [CrossRef]
- Bänziger, M.; Lafitte, H.R. Efficiency of Secondary Traits for Improving Maize for Low-Nitrogen Target Environments. Crop Sci. 1997, 37, 1110–1117. [Google Scholar] [CrossRef]
- Ertiro, B.T.; Olsen, M.; Das, B.; Gowda, M.; Labuschagne, M. Efficiency of Indirect Selection for Grain Yield in Maize (Zea mays L.) under Low Nitrogen Conditions through Secondary Traits under Low Nitrogen and Grain Yield under Optimum Conditions. Euphytica 2020, 216, 134. [Google Scholar] [CrossRef]
- Caixeta, D.S.; Fritsche-Neto, R.; Granato, Í.S.C.; Oliveira, L.R.d.; Galvão, J.C.C. Early Indirect Selection for Nitrogen Use Efficiency in Maize. Rev. Ciência Agronômica 2015, 46, 369–378. [Google Scholar] [CrossRef]
- Ertiro, B.T.; Labuschagne, M.; Olsen, M.; Das, B.; Prasanna, B.M.; Gowda, M. Genetic Dissection of Nitrogen Use Efficiency in Tropical Maize Through Genome-Wide Association and Genomic Prediction. Front. Plant Sci. 2020, 11, 474. [Google Scholar] [CrossRef]
- Leite, J.T.; Amaral Junior, A.T.d.; Kamphorst, S.H.; Lima, V.J.d.; Santos Junior, D.R.d.; Schmitt, K.F.M.; Souza, Y.P.d.; Santos, T.d.O.; Bispo, R.B.; Mafra, G.S.; et al. Water Use Efficiency in Popcorn (Zea mays L. Var. Everta): Which Physiological Traits Would Be Useful for Breeding? Plants 2021, 10, 1450. [Google Scholar] [CrossRef] [PubMed]
- Scharf, P.C.; Brouder, S.M.; Hoeft, R.G. Chlorophyll Meter Readings Can Predict Nitrogen Need and Yield Response of Corn in the North-Central USA. Agron. J. 2006, 98, 655–665. [Google Scholar] [CrossRef]
- Yang, Y.; Timlin, D.J.; Fleisher, D.H.; Lokhande, S.B.; Chun, J.A.; Kim, S.-H.; Staver, K.; Reddy, V.R. Nitrogen Concentration and Dry-Matter Accumulation in Maize Crop: Assessing Maize Nitrogen Status with an Allometric Function and a Chlorophyll Meter. Commun. Soil Sci. Plant Anal. 2012, 43, 1563–1575. [Google Scholar] [CrossRef]
- Zhang, J.; Blackmer, A.M.; Ellsworth, J.W.; Koehler, K.J. Sensitivity of Chlorophyll Meters for Diagnosing Nitrogen Deficiencies of Corn in Production Agriculture. Agron. J. 2008, 100, 543–550. [Google Scholar] [CrossRef]
- Hawkins, J.A.; Sawyer, J.E.; Barker, D.W.; Lundvall, J.P. Using Relative Chlorophyll Meter Values to Determine Nitrogen Application Rates for Corn. Agron. J. 2007, 99, 1034–1040. [Google Scholar] [CrossRef]
- Rashid, M.T.; Voroney, P.; Parkin, G. Predicting Nitrogen Fertilizer Requirements for Corn by Chlorophyll Meter under Different N Availability Conditions. Can. J. Soil Sci. 2005, 85, 149–159. [Google Scholar] [CrossRef]
- Blackmer, T.M.; Schepers, J.S. Techniques for Monitoring Crop Nitrogen Status in Corn. Commun. Soil Sci. Plant Anal. 1994, 25, 1791–1800. [Google Scholar] [CrossRef]
- Zhang, H.; Zhao, Q.; Wang, Z.; Wang, L.; Li, X.; Fan, Z.; Zhang, Y.; Li, J.; Gao, X.; Shi, J.; et al. Effects of Nitrogen Fertilizer on Photosynthetic Characteristics, Biomass, and Yield of Wheat under Different Shading Conditions. Agronomy 2021, 11, 1989. [Google Scholar] [CrossRef]
- Jin, X.; Yang, G.; Tan, C.; Zhao, C. Effects of Nitrogen Stress on the Photosynthetic CO2 Assimilation, Chlorophyll Fluorescence and Sugar-Nitrogen Ratio in Corn. Sci. Rep. 2015, 5, 9311. [Google Scholar] [CrossRef]
- Santos, T.d.O.; de Oliveira, F.T.; Amaral Junior, A.T.d.; de Almeida Filho, J.E.; Bispo, R.B.; de Freitas, M.S.M.; Amaral, J.F.T.d.; Kamphorst, S.H.; de Lima, V.J.; Viana, F.N.; et al. Additive and Non-Additive Effects on the Control of Key Agronomic Traits in Popcorn Lines under Contrasting Phosphorus Conditions. Plants 2022, 11, 2216. [Google Scholar] [CrossRef]
- Mastrodomenico, A.T.; Bohn, M.O.; Lipka, A.E.; Below, F.E. Genomic Selection Using Maize Ex-Plant Variety Protection Germplasm for the Prediction of Nitrogen-Use Traits. Crop Sci. 2019, 59, 212–220. [Google Scholar] [CrossRef]
- Guedes, F.L.; Diniz, R.P.; Balestre, M.; Ribeiro, C.B.; Camargos, R.B.; Souza, J.C. Inheritance of Nitrogen Use Efficiency in Inbred Progenies of Tropical Maize Based on Multivariate Diallel Analysis. Sci. World J. 2014, 2014, 894710. [Google Scholar] [CrossRef] [PubMed]
- DoVale, J.C.; Fritsche-Neto, R.; Bermudez, F.; Miranda, G.V. Efeitos Gênicos de Caracteres Associados à Eficiência No Uso de Nitrogênio Em Milho. Pesqui. Agropecuária Bras. 2012, 47, 385–392. [Google Scholar] [CrossRef]
- Makumbi, D.; Betrán, J.F.; Bänziger, M.; Ribaut, J.-M. Combining Ability, Heterosis and Genetic Diversity in Tropical Maize (Zea mays L.) under Stress and Non-Stress Conditions. Euphytica 2011, 180, 143–162. [Google Scholar] [CrossRef]
- Souza, L.V.d.; Miranda, G.V.; Galvão, J.C.C.; Eckert, F.R.; Mantovani, É.E.; Lima, R.O.; Guimarães, L.J.M. Genetic Control of Grain Yield and Nitrogen Use Efficiency in Tropical Maize. Pesqui. Agropecuária Bras. 2008, 43, 1517–1523. [Google Scholar] [CrossRef]
- Riache, M.; Revilla, P.; Malvar, R.A.; Djemel, A.; Chemlal, A.; Mefti, M. Assessment of Nitrogen Use Efficiency in Algerian Saharan Maize Populations for Tolerance under Drought and No-Nitrogen Stresses. Agronomy 2022, 12, 1123. [Google Scholar] [CrossRef]
- Amegbor, I.K.; Abe, A.; Adjebeng-Danquah, J.; Adu, G.B. Genetic Analysis and Yield Assessment of Maize Hybrids under Low and Optimal Nitrogen Environments. Heliyon 2022, 8, e09052. [Google Scholar] [CrossRef] [PubMed]
- Almeida, V.C.; Viana, J.M.S.; DeOliveira, H.M.; Risso, L.A.; Ribeiro, A.F.S.; DeLima, R.O. Genetic Diversity and Path Analysis for Nitrogen Use Efficiency of Tropical Popcorn (Zea mays Ssp. Everta) Inbred Lines in Adult Stage. Plant Breed. 2018, 137, 839–847. [Google Scholar] [CrossRef]
- Riache, M.; Revilla, P.; Maafi, O.; Malvar, R.A.; Djemel, A. Combining Ability and Heterosis of Algerian Saharan Maize Populations (Zea mays L.) for Tolerance to No-Nitrogen Fertilization and Drought. Agronomy 2021, 11, 492. [Google Scholar] [CrossRef]
- Peterlini, E.; Pinto, R.J.B.; Scapim, C.A.; Rizzardi, D.A.; Bertagna, F.A.B.; Amaral Júnior, A.T. do Diallel Analysis of Popcorn Populations for Yield, Popping Expansion and Resistance to Fall Armyworm. Rev. Ceres 2020, 67, 288–295. [Google Scholar] [CrossRef]
- Schegoscheski Gerhardt, I.F.; Teixeira do Amaral Junior, A.; Ferreira Pena, G.; Moreira Guimarães, L.J.; de Lima, V.J.; Vivas, M.; Araújo Diniz Santos, P.H.; Alves Ferreira, F.R.; Mendonça Freitas, M.S.; Kamphorst, S.H. Genetic Effects on the Efficiency and Responsiveness to Phosphorus Use in Popcorn as Estimated by Diallel Analysis. PLoS ONE 2019, 14, e0216980. [Google Scholar] [CrossRef]
- Schwantes, I.A.; Amaral Júnior, A.T.d.; Vivas, M.; Almeida Filho, J.E.d.; Kamphorst, S.H.; Guimarães, A.G.; Khan, S. Inheritance of Resistance to Fusarium Ear Rot in Popcorn. Crop Breed. Appl. Biotechnol. 2018, 18, 81–88. [Google Scholar] [CrossRef]
- Silva, V.; Amaral JÚnior, A.; Scapim, C.; Freitas Júnior, S.; Gonçalves, L. Inheritance for Economically Important Traits in Popcorn from Distinct Heterotic Groups by Hayman’s Diallel. Cereal Res. Commun. 2010, 38, 272–284. [Google Scholar] [CrossRef]
- Lima, V.J.d.; Amaral Júnior, A.T.d.; Kamphorst, S.H.; Bispo, R.B.; Leite, J.T.; Santos, T.d.O.; Schmitt, K.F.M.; Chaves, M.M.; Oliveira, U.A.d.; Santos, P.H.A.D.; et al. Combined Dominance and Additive Gene Effects in Trait Inheritance of Drought-Stressed and Full Irrigated Popcorn. Agronomy 2019, 9, 782. [Google Scholar] [CrossRef]
- Dofing, S.M.; D’Croz-Mason, N.; Thomas-Compton, M.A. Inheritance of Expansion Volume and Yield in Two Popcorn × Dent Corn Crosses. Crop Sci. 1991, 31, 715–718. [Google Scholar] [CrossRef]
- Coan, M.M.D.; Pinto, R.J.B.; Kuki, M.C.; Amaral Júnior, A.T.; Figueiredo, A.S.T.; Scapim, C.A.; Warburton, M. Inheritance Study for Popping Expansion in Popcorn vs Flint Corn Genotypes. Agron. J. 2019, 111, 2174–2183. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Traits | Type of Inheritance | Population | Number of Environments | Method | Authors |
|---|---|---|---|---|---|---|
| Zea mays everta | Grain yield and two NUE indices (Harmonic Mean of the Relative Performance and Agronomic Efficiency under Low Nitrogen Availability) | Additive and non-additive effects | 90 temperate/tropical hybrids | 2 environments and 2 N conditions | Classical (SCA and GCA) | Santos et al. [29] |
| Daily growth, shoot, root and total dry weight, root shoot ratio, NUpE, NUtE and NtrE efficiencies, lateral and axial root length, root surface area and root volume | Non-additive effects | 2 F1 hybrids, F2 and backcross | 1 environment and 2 N conditions | Generation Mean Analysis | Almeida et al. [2] | |
| Z. mays | Grain yield, anthesis date and silking interval, plant height, ear height and position, ears per plant and senescence | Polygenic | 411 testcrosses tropical hybrids | 9 high N and 13 low N sites | Genomics (GWAS and GP) | Ertiro et al. [110] |
| Grain yield, harvest index, nitrogen harvest index, grain protein concentration, NUE, NUpE, NUtE and genetic utilization | Polygenic | 89 ex-PVP germplasm and 2 public lines (B73 and Mo17) | 11 environments from (location - year, 2011 to 2016) | Classical (GCA, SCA) and genomics (GP) | Mastrodomenico et al. [121] | |
| Grain yield and yield related traits (ear length, ear diameter, cob diameter, and grain size) | Non-additive effects | 55 tropical hybrids | 2 environments and 2 N conditions | Classical (SCA and GCA) | Guedes et al. [122] | |
| Shoot dry weight, lateral and axial root length, NUE and its components (NUpE and NUtE) | Additive effects | 41 hybrids | 1 environment and 2 N conditions | Classical (SCA and GCA) | DoVale et al. [123] | |
| Grain yield | Non-additive effects | 105 tropical/ subtropical hybrids | Two trials under high and low N conditions each | Classical (SCA and GCA) | Makumbi et al. [124] | |
| Grain yield, NUE and its components (NUpE and NUtE) | Additive and non-additive effects | 28 hybrids | 1 environment and 2 N conditions | Classical (SCA and GCA) | Souza et al. [125] | |
| Partial factor productivity, agronomic nitrogen use efficiency, grain nutrient utilization efficiency and protein content | Additive and non-additive effects | 15 hybrids | 2 N conditions and 2 years | Classical (SCA and GCA) | Riache et al. [126] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, T.d.O.; Amaral Junior, A.T.d.; Moulin, M.M. Maize Breeding for Low Nitrogen Inputs in Agriculture: Mechanisms Underlying the Tolerance to the Abiotic Stress. Stresses 2023, 3, 136-152. https://doi.org/10.3390/stresses3010011
Santos TdO, Amaral Junior ATd, Moulin MM. Maize Breeding for Low Nitrogen Inputs in Agriculture: Mechanisms Underlying the Tolerance to the Abiotic Stress. Stresses. 2023; 3(1):136-152. https://doi.org/10.3390/stresses3010011
Chicago/Turabian StyleSantos, Talles de Oliveira, Antônio Teixeira do Amaral Junior, and Monique Moreira Moulin. 2023. "Maize Breeding for Low Nitrogen Inputs in Agriculture: Mechanisms Underlying the Tolerance to the Abiotic Stress" Stresses 3, no. 1: 136-152. https://doi.org/10.3390/stresses3010011
APA StyleSantos, T. d. O., Amaral Junior, A. T. d., & Moulin, M. M. (2023). Maize Breeding for Low Nitrogen Inputs in Agriculture: Mechanisms Underlying the Tolerance to the Abiotic Stress. Stresses, 3(1), 136-152. https://doi.org/10.3390/stresses3010011