Abstract
Swimming pool water treatment by chemicals is an essential step to avoid microbial proliferation and infections namely caused by free living amoeba such as, for example, primary amebic meningoencephalitis and Acanthamoeba keratitis. In the present study, a commercial reactive, CLORICAN, based on chlorine dioxide, was evaluated against Acanthamoeba spp. and Naegleria fowleri. We observed that CLORICAN could eliminate in a short period of incubation time both amoebae. Furthermore, Naegleria fowleri’s trophozoites were more sensitive than those of Acanthamoeba spp. By means of inverted microscopy, the chlorine dioxide was found to greatly affect morphology shape by increasing the cell size shrinkage.
1. Introduction
Free-living amoebae (FLA) are ubiquitous protozoa found in natural habitats, namely soil and water, and artificial habitats such as swimming pools. FLA act as pathogens, as parasites, as carries, or as vehicles of pathogenic microorganisms such as as bacteria or viruses [1]. At present, multiple species have been reported as being pathogens and opportunistic, namely Acanthamoeba spp., Naegleria fowleri, Balamuthia mandrillaris, Vahlkampfia, Vermamoeba and Sappinia. Based on the literature, Acanthamoeba spp., Naegleria fowleri and Balamuthia mandrillaris are the most common FLA, with severe pathogenesis. WHO have defined Acanthamoeba spp. and Naegleria fowleri as quality indicator microorganisms in recreational water [2].
Some Acanthamoeba species or genotypes are the causative agents of severe medical complications including pneumonitis, fatal granulomatous encephalitis, skin infections and Acanthamoeba keratitis (AK) [3,4,5]. Based on the small ribosomal subunit gene (18S rDNA), 22 different genotypes were identified as T1 to T22 [6]. Even though genotype T4 is the most abundant species in both clinical and environmental samples [7], other genotypes such as T2, T3, T5, T6, T11, T13, and T15 were isolated from AK and amoebic encephalitis cases [8]. Acanthamoeba are widely presented in freshwater and marine habitats such as rivers, tap water, swimming pools, and sea water, among others [9]. Overall, 25% of Acanthamoeba sp. isolated from the different habitats host and protect microorganisms from adverse environmental conditions and chemical factors [10,11]. Free living amoebae can be a reservoir of pathogenic bacteria such as Vibrio cholerae, Mycobacterium tuberculosis, Listeria monocytogenes, Helicobacter pylori, Legionella pneumophila, and Klebsiella pneumoniae, among others. Virus such herpes virus, enteroviruses, pandoraviruses and mimivirus have also been isolated from FLA [12,13,14].
Nevertheless, the amoebae from the Naegleria genus were isolated particularly from fresh warm water such as hot springs, swimming pools, and lagoons, among others. Even though more than 40 species of Naegleria have been identified, only Naegleria fowleri, “the brain-eating amoeba”, has been reported to induce a rare and fatal disease, primary amoebic meningoencephalitis (PAM) [15,16].
In order to ensure the quality of recreational water such as swimming pools and so on, several water treatments are conducted, namely filtration, coagulation and disinfection [2]. Disinfection of recreational water is fundamental to eliminate any microbial hazards [17]. During this treatment, chemical agents (e.g., chlorination) and physical methods (e.g., UV-C radiation) are used in order to deactivate the present germs [2]. The CDC recommend a free chlorine concentration of at least 1 ppm in pools and at least 3 ppm in hot tubs/spas and a pH of 7.2–7.8 [18]. Chlorine dioxide has been reported as a strong disinfectant with numerous applications in wastewater treatment, environment and food disinfection, and medicine [19]. For instance, Dupuy et al. (2014) reported the efficacy of chlorine dioxide in inhibiting three different FLA strains [20]. In the present study, the efficacy of a new formulation based on chlorine dioxide was tested against four FLA strains—three Acanthamoeba strains and Naegleria fowleri. The protozoa were analyzed during the time using inverted microscopy.
2. Results
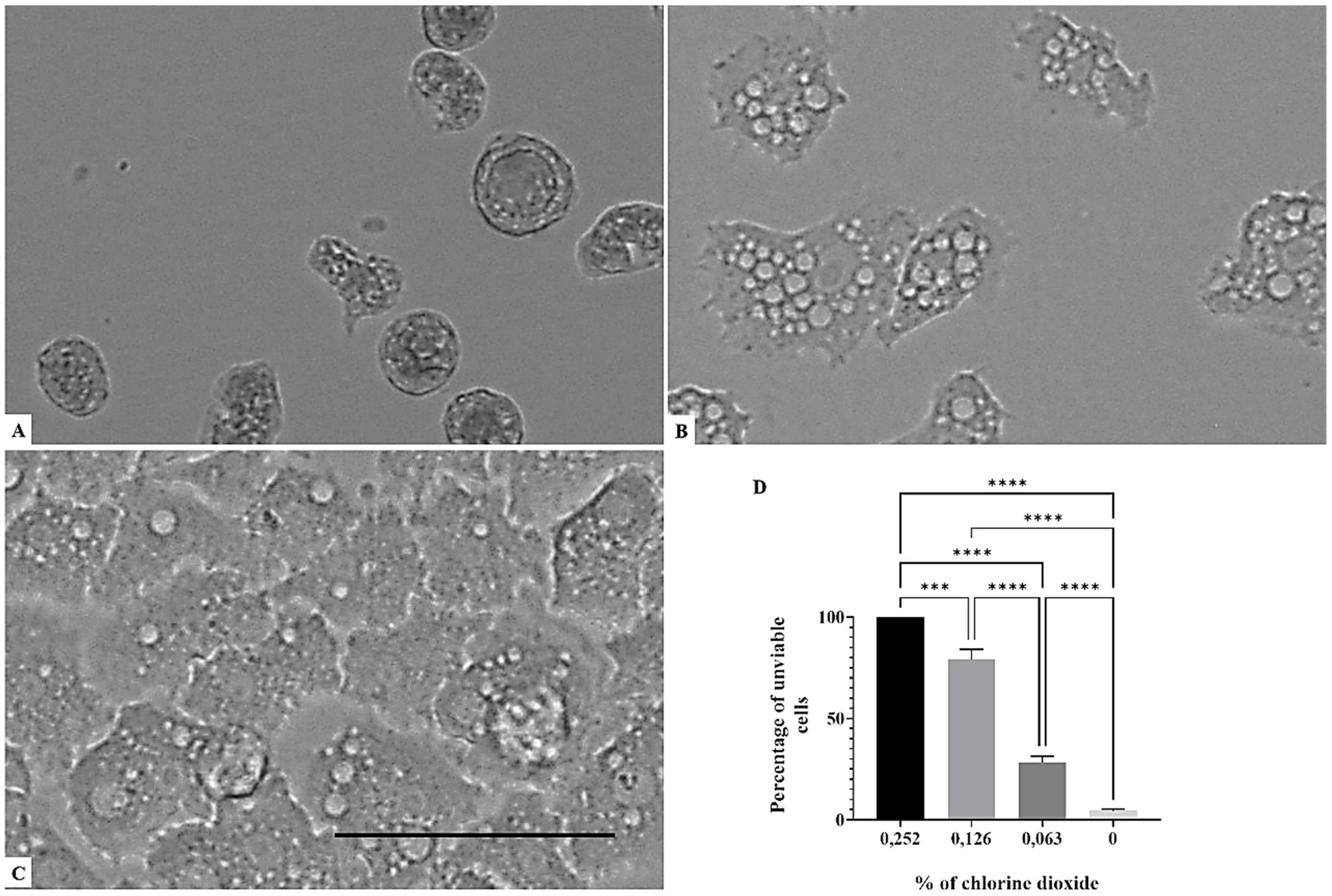
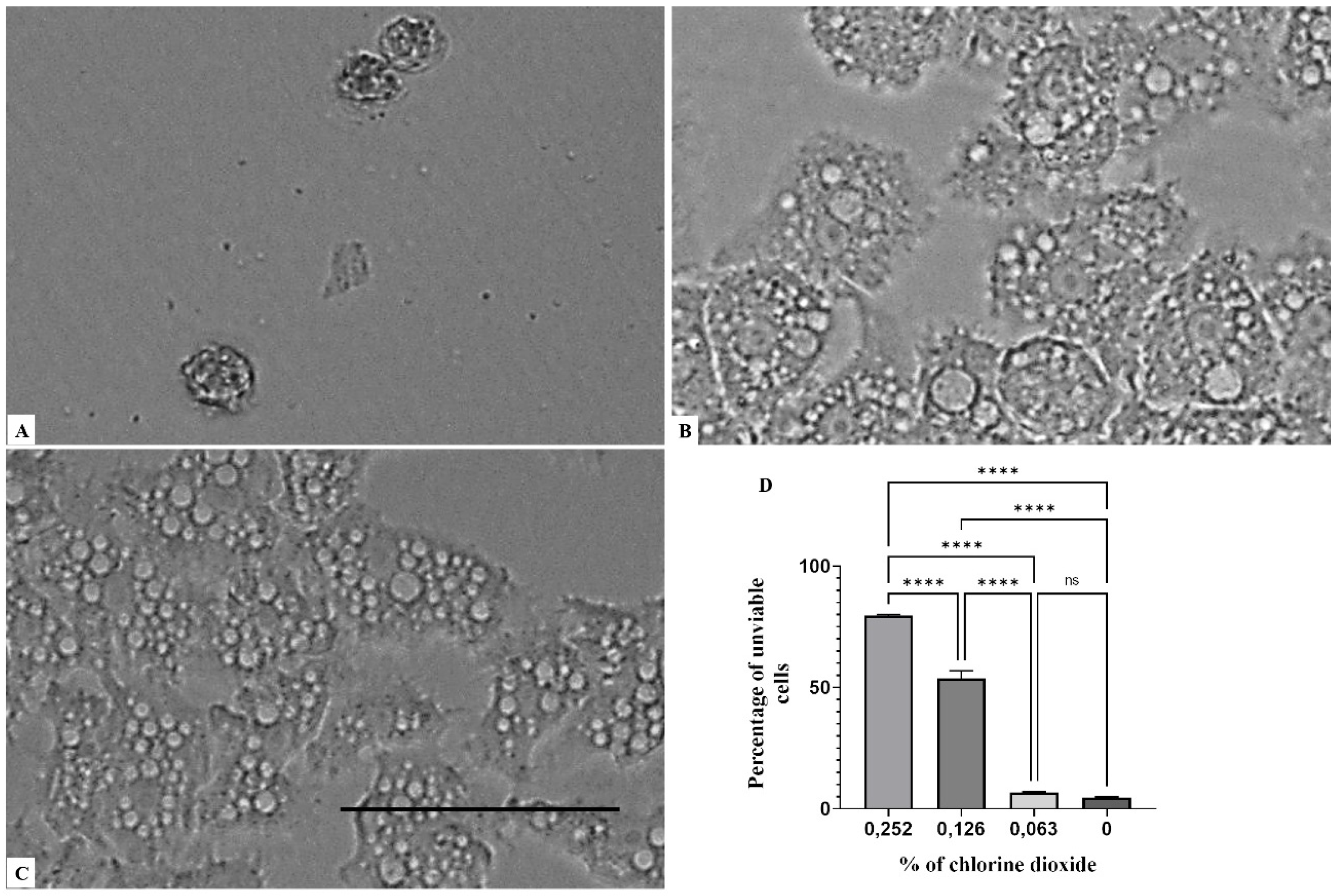
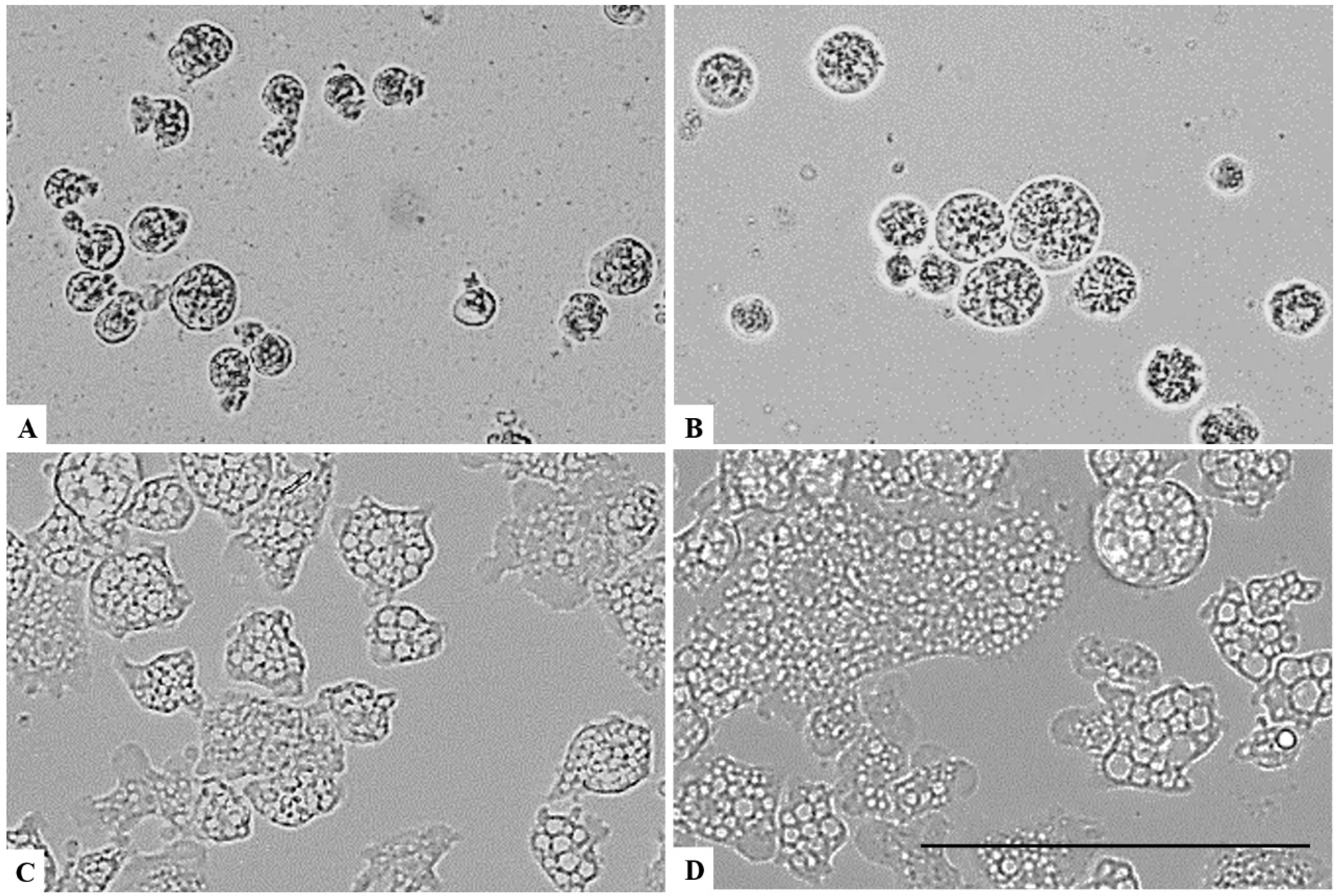
Figure 1, Figure 2 and Figure 3 illustrates that a concentration above 0.063% of chlorine dioxide turned out to be lethal for Acanthamoeba castellanii Neff. In fact, at this concentration, the trophozoites seemed to lose any motility and appeared as fixed cells. After 24 h of incubation with chlorine dioxide at a concentration of 0.063% (Figure 2C), the trophozoites do not show their pseudopodia and become rounded cells. At the same concentration and after 96 h of incubation, we noticed that the rounded trophozoites were not able to encyst in order to remain as pseudo-cysts (Figure 3B). The same effect was observed for Acanthamoeba polyphaga and Acanthamoeba griffini, although both strains were more resistant to CLORICAN. As shown by Figure 4 for Acanthamoeba griffini and Figure 5 for Acanthamoeba polyphaga, a chlorine dioxide concentration of 0.126% was effective to prevent the encystation process. A concentration of 0.063% was able to eliminate 83% and 30%, respectively, of Acanthamoeba castellanii Neff and Acanthamoeba griffini. This dose was ineffective in Acanthamoeba polyphaga.
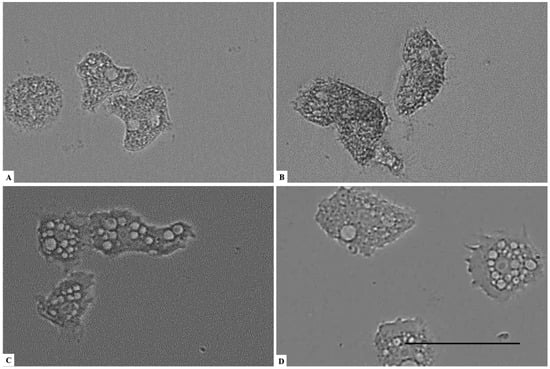
Figure 1.
Effect of CLORICAN on Acanthamoeba castellanii Neff trophozoite after 3 h of incubation at 0.252% (A), 0.126% (B), 0.063% (C) of chlorine dioxide and negative control (D). At these concentrations, trophozoites seems to lose any motility and appeared as fixed cells (A,B). All images (×40) are representative of the population of treated amoebae and are based on the live cell imaging microscope EVOS FL cell imaging system. Scale bar = 50 µm.
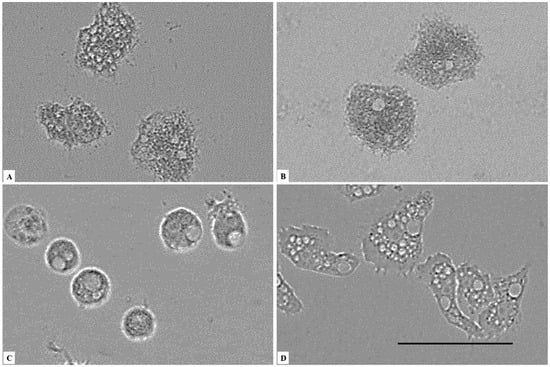
Figure 2.
Effect of CLORICAN on Acanthamoeba castellanii Neff trophozoite after 24 h of incubation at concentrations of 0.252% (A), 0.126% (B), and 0.063% (C) chlorine dioxide, and the negative control (D). The characteristic pseudopodia of Acanthamoeba castellanii Neff cannot be observed in treated cells. (A–C) All images (×40) are representative of the population of treated amoebae and are based on the live cell imaging microscope EVOS FL cell imaging system. Scale bar = 50 µm.
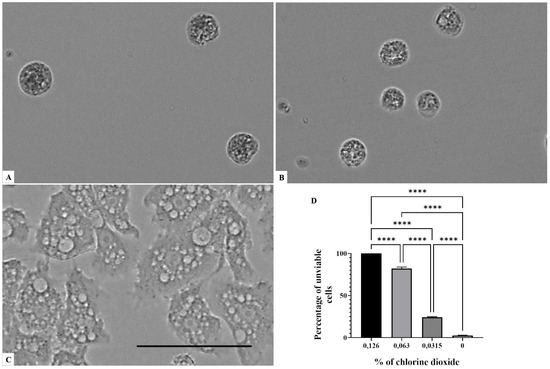
Figure 3.
Growth inhibition of Acanthamoeba castellanii Neff after 96 h of incubation. Acanthamoeba spp. cells were incubated with a concentration of 0.126% chlorine dioxide (A). Acanthamoeba cells were incubated with a concentration of 0.063% chlorine dioxide (B). Negative control (C). Histograms of cells’ unviability percentages for each tested concentration of chlorine dioxide (D). All images (×40) are representative of the population of treated amoebae and are based on the live cell imaging microscope EVOS FL cell imaging system. ** p < 0.01; *** p < 0.001; **** p < 0.0001 significance differences among different doses of chlorine dioxide. Scale bar = 50 µm.
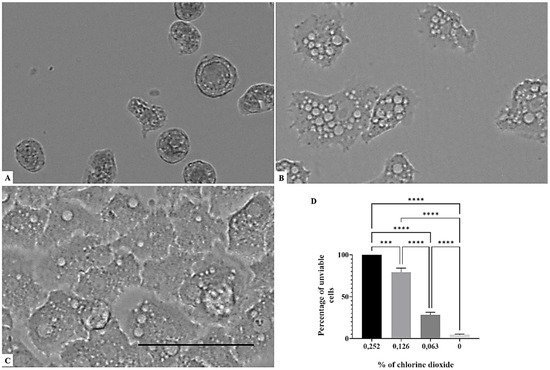
Figure 4.
Growth inhibition of Acanthamoeba griffini after 96 h of incubation. Acanthamoeba spp. cells were incubated with a concentration of 0.126% chlorine dioxide (A). Acanthamoeba cells were incubated with a concentration of 0.063% chlorine dioxide (B). Negative control (C). Histograms of cells’ unviability percentages for each tested concentration of chlorine dioxide (D). All images (×40) are representative of the population of treated amoebae and are based on the live cell imaging microscope EVOS FL cell imaging system. *** p < 0.001; **** p < 0.0001 significance differences among different doses of chlorine dioxide. Scale bar = 50µm.
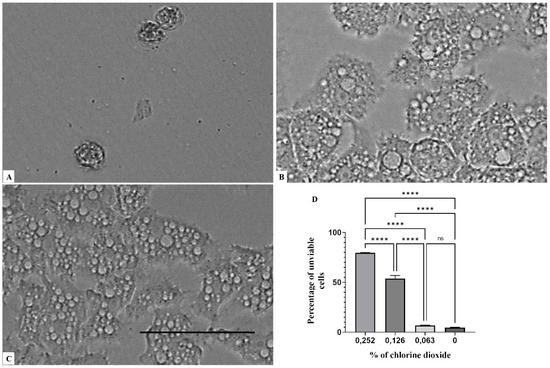
Figure 5.
Growth inhibition of Acanthamoeba polyphaga; after 96 h of incubation. Acanthamoeba spp. cells were incubated with a concentration of 0.137% chlorine dioxide (A). Acanthamoeba cells were incubated with a concentration of 0.063% chlorine dioxide (B). Negative control (C). Histograms of cells’ unviability percentages for each tested concentration of chlorine dioxide (D). All images (×40) are representative of the population of treated amoebae and are based on the live cell imaging microscope EVOS FL cell imaging system. Data are presented as means ± SD (N = 3) **** p < 0.0001 significance differences among different doses of chlorine dioxide. Scale bar = 50 µm.
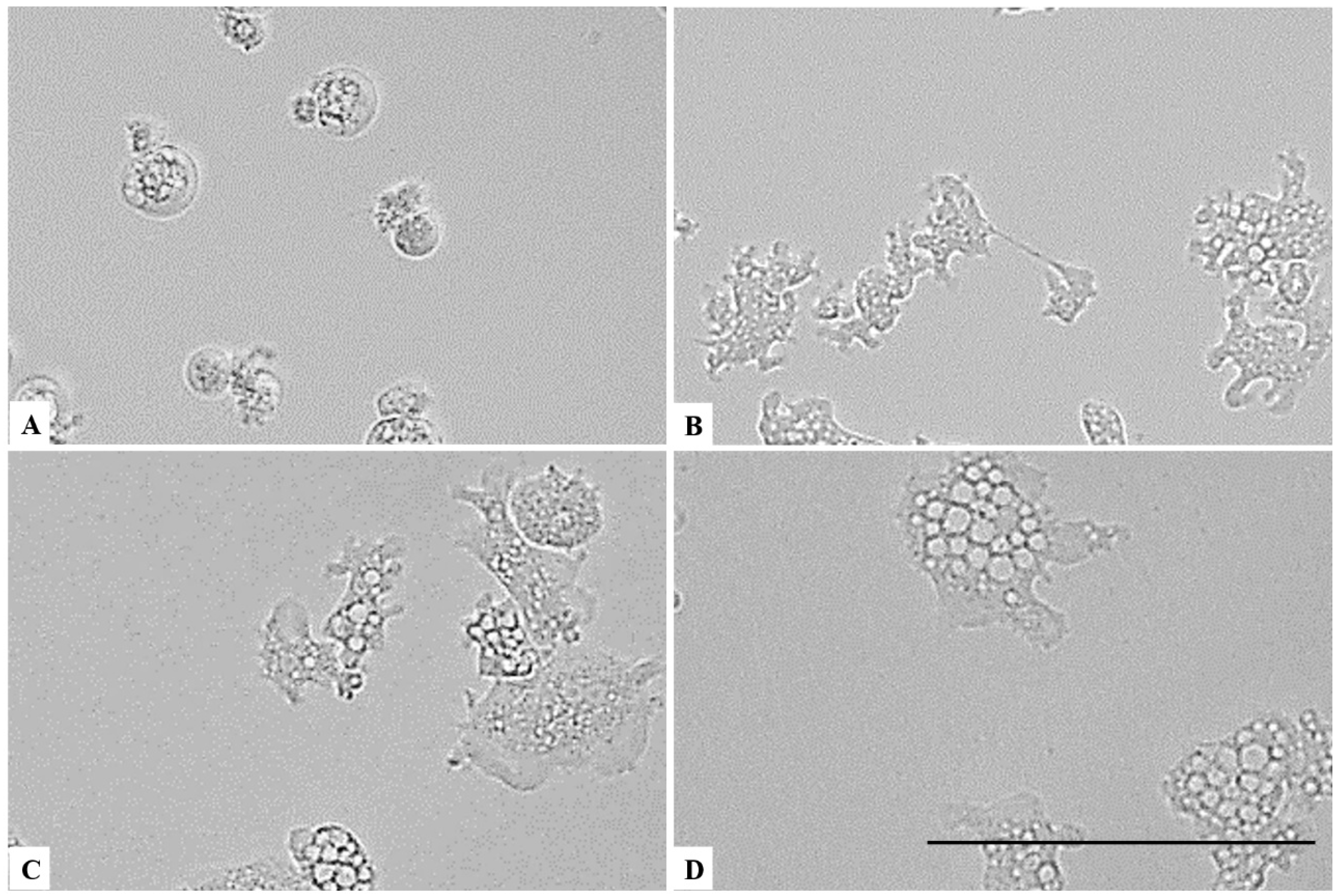
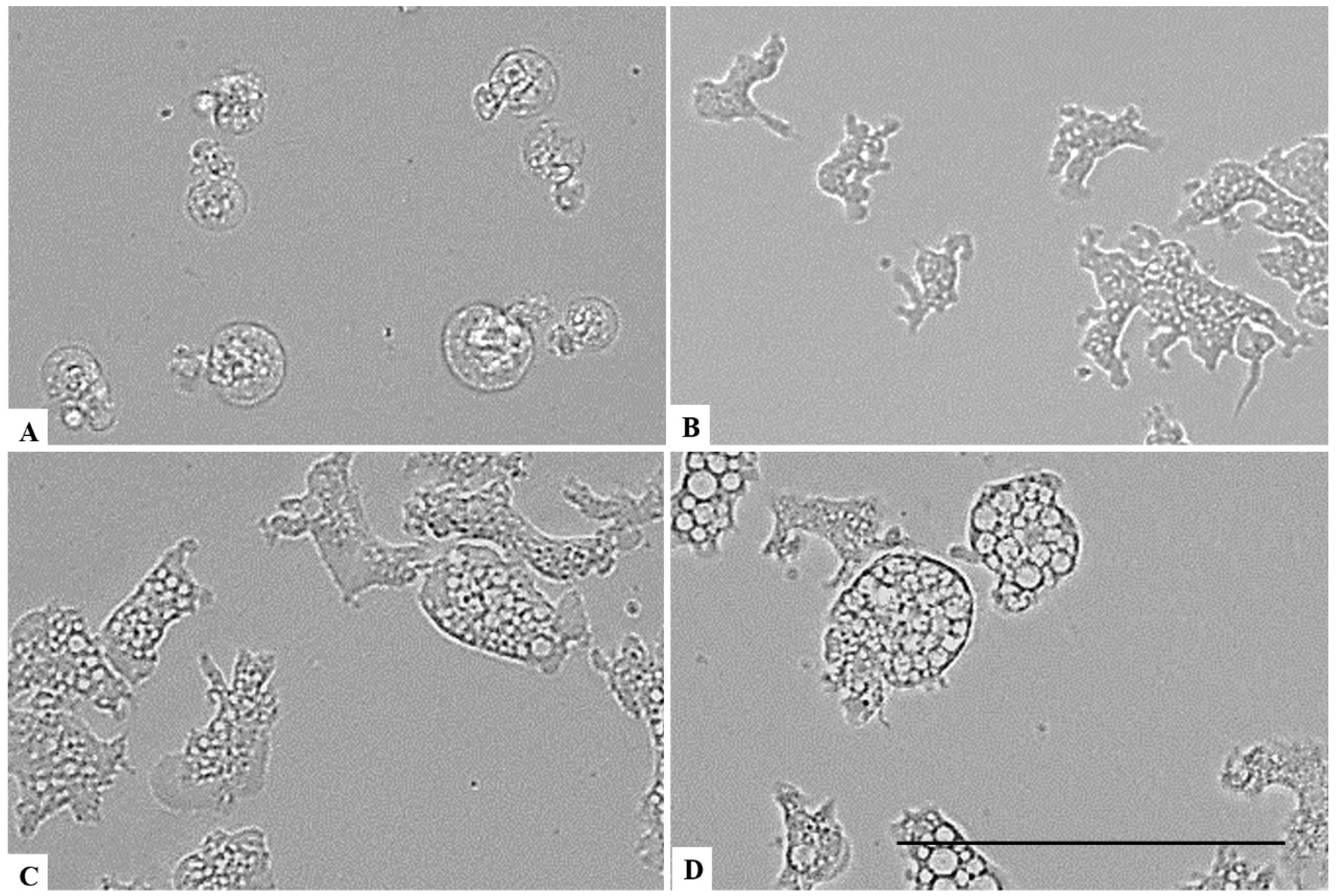
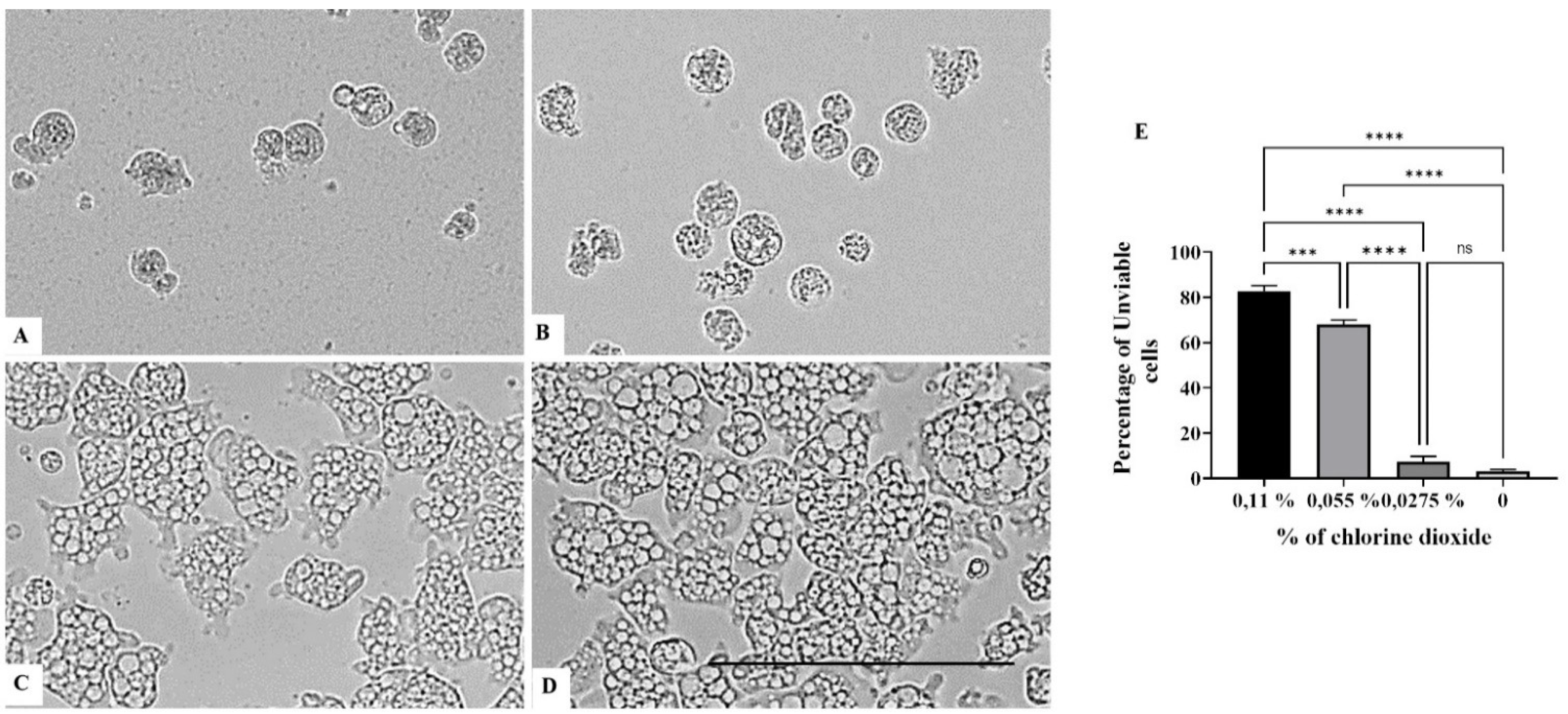
On the other hand, a chlorine dioxide concentration of up to 0.055 % on the Naegleria fowleri trophozoites, and a strong change in morphology was detected after 15 min of incubation; in fact, most of the amoebae lost their amoeboid shape, leading them to be rounded (Figure 6). This effect persists for at least 24 h (see Figure 7, Figure 8 and Figure 9). At a lower concentration of chlorine dioxide, precisely 0.0275%, the trophozoite showed slight morphological changes as well as in the number of intracellular vesicles after 1 h of incubation (Figure 7). After 24 h of incubation with a 0.055% concentration of chlorine dioxide, we noticed the presence of pseudo-cyst cells; nevertheless, these cells are not able to complete the encystation process, as shown in Figure 9B.
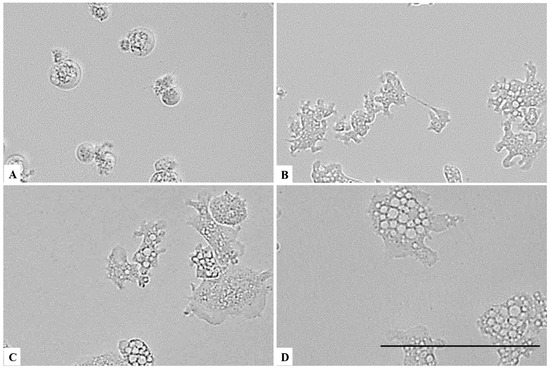
Figure 6.
Effect of CLORICAN on Naegleria fowleri trophozoites after 15 min of incubation at chlorine dioxide concentrations of 0.11% (A), 0.055% (B), and 0.0275% (C) and the negative control (D). The Naegleria fowleri trophozoites lost their amoeboid morphology. All images (×40) are representative of the population of treated amoebae and are based on the live cell imaging microscope EVOS FL cell imaging system. Scale bar = 50 µm.
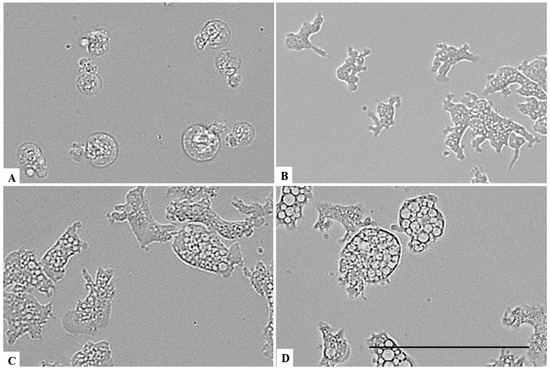
Figure 7.
Effect of CLORICAN on Naegleria fowleri trophozoites after 1 h of incubation at chlorine dioxide concentrations of 0.11% (A), 0.055% (B), and 0.0275% (C) and the negative control (D). All images (×40) are representative of the population of treated amoebae and are based on the live cell imaging microscope EVOS FL cell imaging system. Scale bar = 50 µm.
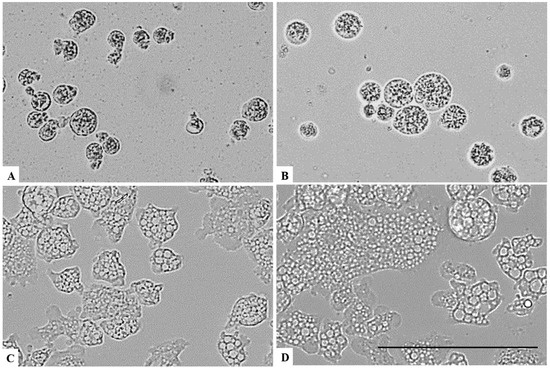
Figure 8.
Effect of CLORICAN on Naegleria fowleri trophozoite after 24 h of incubation at chlorine dioxide concentrations of 0.11% (A), 0.055% (B), and 0.0275% (C) and the negative control (D). All images (×40) are representative of the population of treated amoebae and are based on the live cell imaging microscope EVOS FL cell imaging system. Scale bar = 50µm.
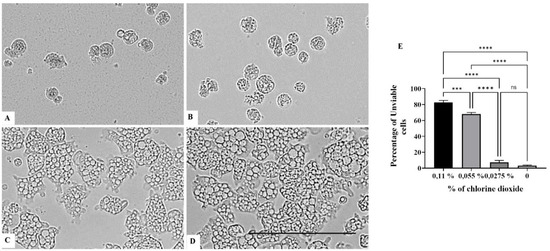
Figure 9.
Effect of CLORICAN on Naegleria fowleri trophozoite after 48 h of incubation at chlorine dioxide concentrations of 0.11% (A), 0.055% (B), and 0.0275% (C) and the negative control (D). All images (×40) are representative of the population of treated amoebae and are based on the live cell imaging microscope EVOS FL cell imaging system. (E) Data are presented as means ± SD (N = 3). *** p < 0.001; **** p < 0.0001 significance differences among different doses of chlorine dioxide. Scale bar = 50 µm.
3. Discussion
Several free-living amoebae including Naegleria fowleri and some species from the Acanthamoeba genus represent a potential threat to human health due to their intrinsic pathogenicity and their role as protective and disseminating hosts for other pathogenic microorganisms [21]. Due to their higher resistance to diverse water treatments (physical and chemical), FLA may act as a Trojan horse transporting pathogenic bacteria such as Legionella spp., Mycobacterium avium and Listeria monocytogenes, among others [22]. Despite the fact that FLA have recently gained considerable attention due to their potential to develop illnesses, there is still a lack of information in methods to eliminate them [21]. Even though the disinfection treatment recommendations for recreational water are detailed by the WHO, CDC, and Commission for the Environment, Climate Change and Energy, among others, none of them include a specific protocol for free living amoeba eradication. The chemical disinfectants that are mostly used to eliminate pathogens are chlorine, chlorine dioxide and bromochlorodimethylhydantoin (BCDMH). In the WHO guideline, it is announced that the residual free chlorine level should not exceed 3 mg/L in public and semi-public pools and should not exceed 5 mg/l in public and semi-public hot tubs [2].
Chlorine dioxide, a potent oxidant and antimicrobial, is a disinfectant widely used in paper bleaching, surface disinfection, biofilm removal in water distribution systems, and, increasingly, in water treatment to produce potable water [18]. Several authors have reported the higher efficacy of chlorine dioxide than chlorine to eliminate free living amoeba cells [19]. Mogoa et al. (2010) reported that a concentration of free chlorine lower than 3 mg/L was not able to inhibit Acanthamoeba castellanii Neff, whereas in this study, a concentration of 3 mg/L was able to inhibit the growth of 60% of the cell culture [23]. On the other hand, the CDC recommends a concentration of 1–3 mg/L [18] of free chlorine to avoid the growth of Naegleria fowleri in swimming pools. Moreover, according to the Spanish recreational swimming pool regulation (RD 742/2013 27th of September), the recommended parametric value of free chlorine remains between 0.5 and 2 mg/L [24], while the CLORICAN product’s indications establish a concentration of 0.4 mg/L for biocidal activity.
The amoebicidal capacity of CLORICAN on the trophozoite stage of Acanthamoeba spp. and Naegleria fowleri was studied over 48 h using an inverted light-microscope. In Acanthamoeba castellanii (ATCC 30234), Mogoa et al. (2010) reported that chlorine treatment led to the loss of pseudopodia and shrinking in the cell size and membrane permeabilization [23]. The same authors proved in a second paper that chlorine dioxide was as effective as chlorine in eliminating Acanthamoeba castellanii Neff by reducing the cell size more more and inducing membrane permeabilization, as shown by propidium iodide staining [21].
In our study, CLORICAN was shown to have a stronger effect on Naegleria fowleri than Acanthamoeba spp. Several authors have mentioned the higher sensitivity of Naegleria fowleri to chlorine and chlorine dioxide compared to non-pathogenic Naegleria, namely N. gruberi and Acanthamoeba spp. [25]. The chlorine dioxide did affect Naegleria fowleri cells by altering the amoeboid shape and inducing cell shrinking.
4. Materials and Methods
4.1. Chemical
CLORICAN®, a commercial liquid form of chlorine dioxide, is used to disinfect swimming pools. The reactive is produced by Vadeguas company, Canary Islands, Spain. The tested solution has a pH of 4.5–6 and a chlorine dioxide concentration of 0.75% (w/w).
4.2. Cell Culture
The strains used in this study were: Acanthamoeba castellanii Neff (ATCC 30010), Acanthamoeba polyphaga, genotype T4 (ATCC 30461) and Acanthamoeba griffini, genotype T3 obtained in previous studies [17]. Those strains were axenically grown in PYG medium (0.75% (w/v) proteose peptone, 0.75% (w/v) yeast extract and 1.5% (w/v) glucose) containing 40 µg/mL of gentamicin (Biochrom AG, Cultek, Granollers, Barcelona, Spain).
The Naegleria strain used was Naegleria fowleri (ATCC® 30808™). This strain was kept in the biological security facilities level 3 of our institution following Spanish biosafety guidelines for this pathogen. It was axenically cultured at 37 °C in 2% (w/v) bactocasitone medium (Thermo Fisher Scientific, Madrid, Spain) supplemented with 10% (v/v) fetal bovine serum (FBS), containing 0.5 mg/mL of streptomycin sulfate (Sigma-Aldrich, Madrid, Spain) and 0.3 µg/mL of penicillin G sodium salt (Sigma-Aldrich, Madrid, Spain).
4.3. In Vitro Activity Against Trophozoite Stage
The amoebicidal activity experiments were carried out using an inverted light-microscope Leica DM IL (Wetzlar, Germany). Initially, the trophozoites were counted using a Countess II FL automatic cell counter (Thermo Fisher Scientific, Madrid, Spain) to prepare a working concentration of 5.104 trophozoites/well and 105 cells/well, respectively, for Acanthamoeba spp. and Naegleria fowleri strains), and 50 µL per well was seeded in triplicate in a 96-well plate (Thermo Fisher Scientific, Madrid, Spain).
After that, a serial dilution of CLORICAN in the same culture medium was added to the plate (50 µL per well). As a negative control, the trophozoites were incubated with the medium alone. Plates were incubated with slight agitation for 96 h at 26 °C for Acanthamoeba spp. and 48 h at 37 °C for Naegleria. Subsequently, the plates were observed during the incubation time with an inverted light-microscope (15 min, 1 h, 24 h, 48 h and 96 h). The numbers of viable and nonviable trophozoite were counted using a Countess II FL automatic cell counter (Thermo Fisher Scientific, Madrid, Spain) at 96 h for Acanthamoeba spp. and 48 h for Naegleria fowleri using an inverted microscope and confirmed with trypan blue (0.4%) staining using the Countess II FL (Thermo Fisher Scientific, Madrid, Spain)
4.4. Statistical Analysis
All data are expressed as the mean ± standard deviation of at least three independent experiments. To highlight the chlorine dioxide’s effect on Acanthamoeba spp., a statistical comparison was conducted using the t-test and one-way analysis of variance (ANOVA). All the analysis and the graphics were done by GraphPad Prism version 8.0. Statistical significance was set at p < 0.05.
5. Conclusions
The CLORICAN reactive was highly effective at eliminating both amoeba strains tested in vitro. Moreover, Naegleria fowleri’s trophozoites were more sensitive than those of Acanthamoeba spp. The chlorine dioxide greatly affected the morphology shape and increased the cell size shrinkage. Further experiments are needed to better understand the mode of action of CLORICAN and to evaluate its in situ efficacy in a scaled-up process, e.g., in a swimming pool.
Author Contributions
Conceptualization, I.S., M.R.-B., J.E.P., J.L.-M.; methodology, I.S., M.R.-B., J.E.P., J.L.-M.; software, I.S., A.R.-L., I.A.-J., R.L.R.-E.; validation, I.S., M.R.-B., J.E.P., and J.L.-M.; formal analysis, I.S., A.R.-L., I.A.-J., R.L.R.-E.; investigation, I.S., A.R.-L., I.A.-J., R.L.R.-E., M.R.-B.; resources, I.S., M.R.-B.; data curation, A.R.-L., I.A.-J., R.L.R.-E.; writing—original draft preparation, I.S., A.R.-L., I.A.-J., R.L.R.-E.; writing—review and editing, All the authors.; visualization, I.S., M.R.-B., J.E.P., J.L.-M.; supervision, J.E.P., J.L.-M.; project administration, J.E.P. and J.L.-M.; funding acquisition, J.E.P. and J.L.-M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the grants PI18/01380, Fondo Europeo de Desarollo Regional (FEDER) and RICET (project no. RD16/0027/0001 of the program of Redes Temáticas de Investigación Cooperativa, FIS), Spanish Ministry of Science, Innovation and Universities, Madrid, Spain. IS and MRB was also funded by RICET. ARL (TESIS2020010054), IAJ (TESIS2020010063) and RLRE (TESIS2020010117) were funded by the Agencia Canaria De Investigación Innovación Y Sociedad De La Información Gobierno De Canarias. (ACIISI), y Fondo Social Europeo (FSE).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to thank Octavio Santana from VADEAGUAS for providing the CLORICAN and for their support.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Javanmard, E.; Niyyati, M.; Lorenzo-Morales, J.; Lasjerdi, Z.; Behniafar, H.; Mirjalali, H. Molecular identification of waterborne free living amoebae (Acanthamoeba, Naegleria and Vermamoeba) isolated from municipal drinking water and environmental sources, Semnan province, north half of Iran. Exp. Parasitol. 2017, 183, 240–244. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. World Health Organization Guidelines for Safe Recreational Water Environments: Volume 2: Swimming Pools and Similar Environments; World Health Organization: Geneva, Switzerland, 2006; p. 118, xiii. [Google Scholar]
- Marciano-Cabral, F.; Puffenbarger, A.R.; Cabral, G.A. The Increasing Importance of Acanthamoeba Infections1. J. Eukaryot. Microbiol. 2000, 47, 29–36. [Google Scholar] [CrossRef]
- Scheid, P.; Zöller, L.; Pressmar, S.; Richard, G.; Michel, R. An extraordinary endocytobiont in Acanthamoeba sp. isolated from a patient with keratitis. Parasitol. Res. 2008, 102, 945–950. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo-Morales, J.; Khan, N.; Walochnik, J. An update onAcanthamoebakeratitis: Diagnosis, pathogenesis and treatment. Parasite 2015, 22, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corsaro, D. Update on Acanthamoeba phylogeny. Parasitol. Res. 2020, 119, 3327–3338. [Google Scholar] [CrossRef]
- Maciver, S.K.; Asif, M.; Simmen, M.W.; Lorenzo-Morales, J. A systematic analysis of Acanthamoeba genotype frequency correlated with source and pathogenicity: T4 is confirmed as a pathogen-rich genotype. Eur. J. Protistol. 2013, 49, 217–221. [Google Scholar] [CrossRef]
- Grün, A.-L.; Stemplewitz, B.; Scheid, P. First report of an Acanthamoeba genotype T13 isolate as etiological agent of a keratitis in humans. Parasitol. Res. 2014, 113, 2395–2400. [Google Scholar] [CrossRef]
- Al-Herrawy, A.; Bahgat, M.; Mohammed, A.-E.; Ashour, A.; Hikal, W. Acanthamoeba species in Swimming Pools of Cairo, Egypt. Iran. J. Parasitol. 2014, 9, 194–201. [Google Scholar]
- Kot, K.; Łanocha-Arendarczyk, N.A.; Kosik-Bogacka, D.I. Amoebas from the genus Acanthamoeba and their pathogenic properties. Ann. Parasitol. 2018, 64, 299–308. [Google Scholar] [CrossRef]
- Balczun, C.; Scheid, P.L. Free-Living Amoebae as Hosts for and Vectors of Intracellular Microorganisms with Public Health Significance. Viruses 2017, 9, 65. [Google Scholar] [CrossRef]
- Scheid, P.; Balczun, C.; Schaub, G.A. Some secrets are revealed: Parasitic keratitis amoebae as vectors of the scarcely described pandoraviruses to humans. Parasitol. Res. 2014, 113, 3759–3764. [Google Scholar] [CrossRef]
- Rumelt, S.; Cohen, I.; Rehany, U. Spontaneous Corneal Graft Ulcerative Perforation Due to Mixed Acanthamoeba and Herpes Simplex Keratitis: A Clinicopathologic Study. Cornea 2000, 19, 240–242. [Google Scholar] [CrossRef]
- Guimaraes, A.J.; Gomes, K.X.; Cortines, J.; Peralta, J.M.; Peralta, R.H. Acanthamoeba spp. as a universal host for pathogenic microorganisms: One bridge from environment to host virulence. Microbiol. Res. 2016, 193, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Jonckheere, J.F.D. A Century of Research on the Amoeboflagellate Genus Naegleria. Acta Protozool. 2002, 41, 309. [Google Scholar]
- Dunn, A.L.; Reed, T.; Stewart, C.; Levy, R.A. Naegleria fowleriThat Induces Primary Amoebic Meningoencephalitis: Rapid Diagnosis and Rare Case of Survival in a 12-Year-Old Caucasian Girl. Lab. Med. 2016, 47, 149–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liviac, D.; Wagner, E.D.; Mitch, W.A.; Altonji, M.J.; Plewa, M.J. Genotoxicity of Water Concentrates from Recreational Pools after Various Disinfection Methods. Environ. Sci. Technol. 2010, 44, 3527–3532. [Google Scholar] [CrossRef]
- CDC Healthy Swimming/Recreational Water. Available online: https://www.cdc.gov/healthywater/swimming/index.html (accessed on 8 March 2021).
- Ma, J.-W.; Huang, B.-S.; Hsu, C.-W.; Peng, C.-W.; Cheng, M.-L.; Kao, J.-Y.; Way, T.-D.; Yin, H.-C.; Wang, S.-S. Efficacy and Safety Evaluation of a Chlorine Dioxide Solution. Int. J. Environ. Res. Public Health 2017, 14, 329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupuy, M.; Berne, F.; Herbelin, P.; Binet, M.; Berthelot, N.; Rodier, M.-H.; Soreau, S.; Héchard, Y. Sensitivity of free-living amoeba trophozoites and cysts to water disinfectants. Int. J. Hyg. Environ. Health 2014, 217, 335–339. [Google Scholar] [CrossRef]
- Coulon, C.; Collignon, A.; McDonnell, G.; Thomas, V. Resistance of Acanthamoeba Cysts to Disinfection Treatments Used in Health Care Settings. J. Clin. Microbiol. 2010, 48, 2689–2697. [Google Scholar] [CrossRef] [Green Version]
- Greub, G.; Raoult, D. Microorganisms Resistant to Free-Living Amoebae. Clin. Microbiol. Rev. 2004, 17, 413–433. [Google Scholar] [CrossRef] [Green Version]
- Mogoa, E.; Bodet, C.; Morel, F.; Rodier, M.-H.; Legube, B.; Héchard, Y. Cellular Response of the Amoeba Acanthamoeba castellanii to Chlorine, Chlorine Dioxide, and Monochloramine Treatments. Appl. Environ. Microbiol. 2011, 77, 4974–4980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agencia Estatal Boletín Oficial del Estado, Real Decreto 742/2013, de 27 de Septiembre, por el que se Establecen los Criterios técnico-Sanitarios de las Piscinas. Available online: https://www.boe.es/buscar/doc.php?id=BOE-A-2013-10580 (accessed on 8 March 2021).
- Ahmad, T.; Shukla, K.; Sharma, A.K. Effect of Chlorine (Disinfectant) on Viability of Pathogenic Free Living Amoebae. Indian J. Fundam. Appl. Life Sci. 2012, 2, 132. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).