
Molecular Characterization of Dehydrin in Azraq Saltbush among Related Atriplex Species




Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Cloning DHNs
2.3. Analysis of Atriplex DHNs
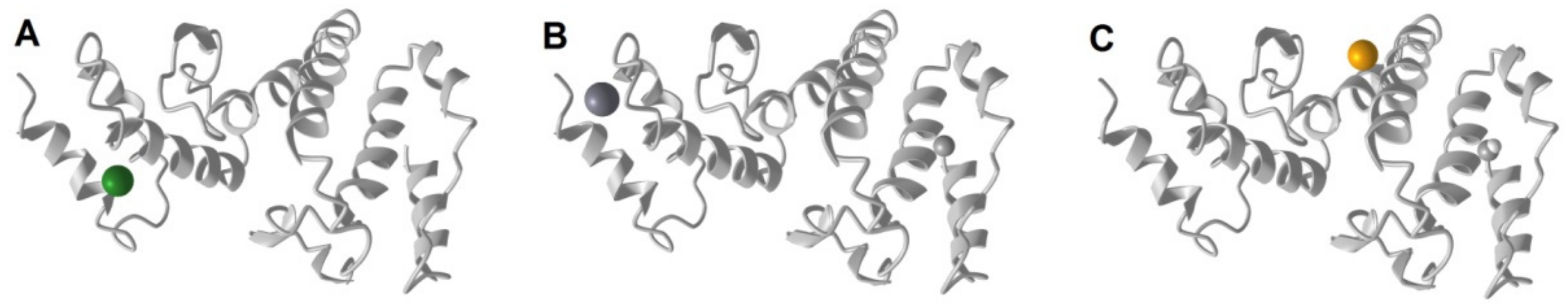
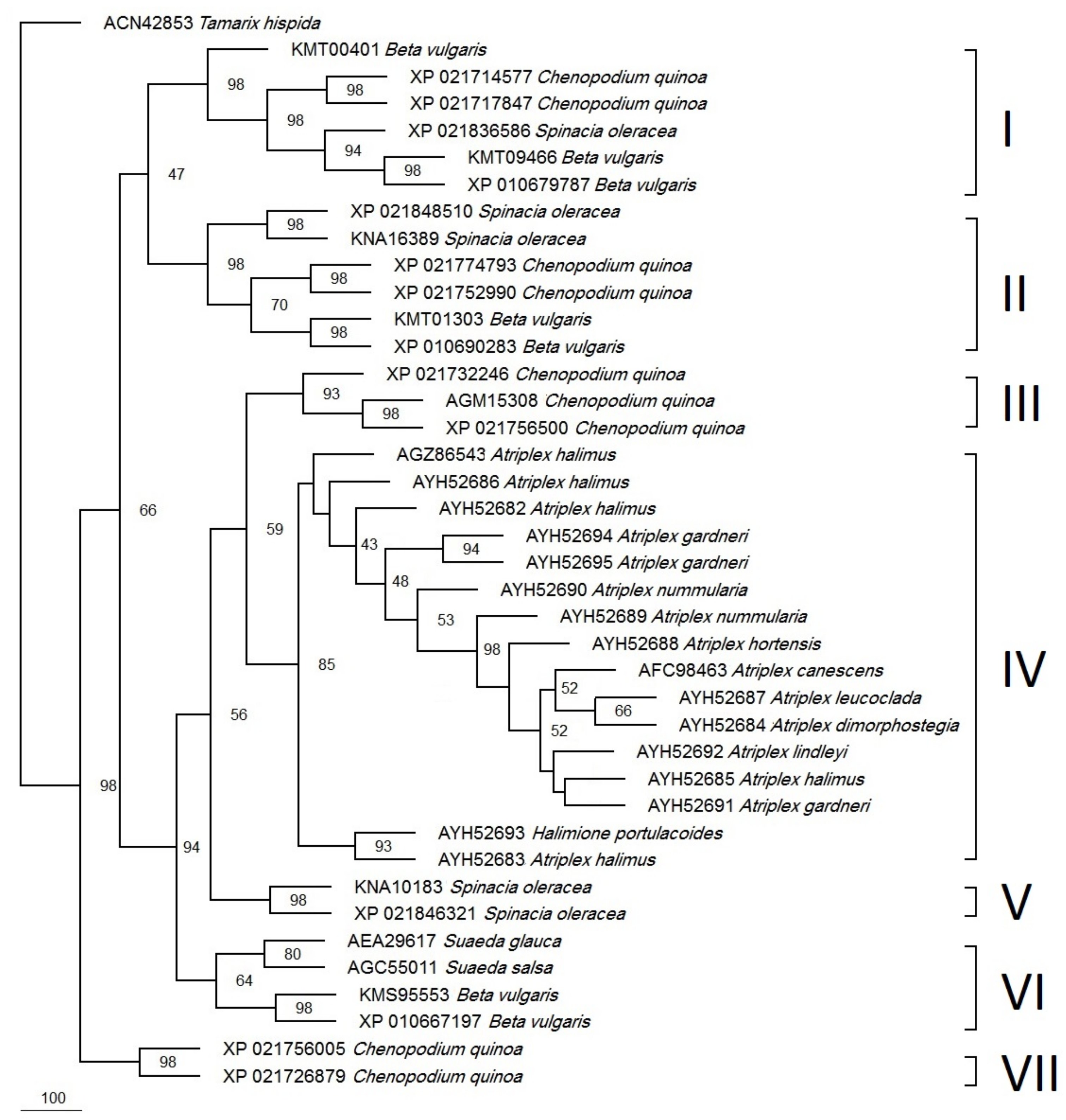
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alsadon, A.A.; Sadder, M.T.; Wahb-Allah, M.A. Responsive gene screening and exploration of genotypes responses to salinity tolerance in tomato. Aust. J. Crop Sci. 2013, 7, 1383–1395. [Google Scholar]
- Sadder, M.T.; Anwar, F.; Al-Doss, A.A. Gene expression and physiological analysis of Atriplex halimus (L.) under salt stress. Aust. J. Crop Sci. 2013, 7, 112–118. [Google Scholar]
- Brake, M.; Al-Qadumii, L.; Hamasha, H.; Migdadi, H.; Awad, A.; Haddad, N.; Sadder, M.T. Development of SSR Markers linked to stress responsive genes along tomato chromosome 3 (Solanum lycopersicum L.). BioTech 2022, 11, 34. [Google Scholar] [CrossRef]
- Ruas, C.F.; Ruas, P.M.; Stutz, H.C.; Fairbanks, D.J. Cytogenetic studies in the genus Atriplex (Chenopodiaceae). Caryologia 2001, 54, 129–145. [Google Scholar] [CrossRef]
- Frankton, C.; Bassett, I.J. The genus Atriplex (Chenopodiaceae) in Canada. I. Three introduced species: A. heterosperma, A. oblongifolia, and A. hortensis. Canad. J. Bot. 1968, 46, 1309–1313. [Google Scholar] [CrossRef]
- Nord, E.C.; Christensen, D.R.; Plummer, A.P. Atriplex species [or Taxa] that spread by root spouts, stem layers, and by seed. Ecology 1969, 50, 324–326. [Google Scholar] [CrossRef]
- Wilson, P.G. Chenopodiaceae. In Flora of Australia; Australian Government Publishing Service: Canberra, Australia, 1984; Volume 4, pp. 81–330. [Google Scholar]
- Le Houérou, H.N. The role of saltbushes (Atriplex spp.) in arid land rehabilitation in the Mediterranean Basin: A review. Agrofor. Syst. 1992, 18, 107–148. [Google Scholar] [CrossRef]
- Sadder, M.T.; Alshomali, I.; Ateyyeh, A.; Musallam, A. Physiological and molecular responses for long term salinity stress in common fig (Ficus carica L.). Physiol. Mol. Biol. Plants 2021, 27, 107–117. [Google Scholar] [CrossRef]
- Sadder, M.T.; Ateyyeh, A.F.; Alswalmah, H.; Zakri, A.M.; Alsadon, A.A.; Al-Doss, A.A. Characterization of putative salinity-responsive biomarkers in olive (Olea europaea L.). Plant Genet. Resour. 2021, 19, 133–143. [Google Scholar] [CrossRef]
- Dunn, G.M.; Neales, T.F. Are the effects of salinity on growth and leaf gas exchange related? Photosynthetica 1993, 29, 33–42. [Google Scholar]
- Xu, C.; Tang, X.; Shao, H.; Wang, H. Salinity tolerance mechanism of economic halophytes from physiological to molecular hierarchy for improving food quality. Curr. Genom. 2016, 17, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Sadder, M.T.; Al-Doss, A.A. Characterization of dehydrin AhDHN from Mediterranean saltbush (Atriplex halimus). Turk. J. Biol. 2014, 38, 469–477. [Google Scholar] [CrossRef]
- Hou, C.; Tian, W.; Kleist, T.; He, K.; Garcia, V.; Bai, F.; Hao, Y.; Luan, S.; Li, L. DUF221 proteins are a family of osmosensitive calcium-permeable cation channels conserved across eukaryotes. Cell Res. 2014, 24, 632–635. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Yang, H.; Xue, Y.; Kong, D.; Ye, R.; Li, C.; Zhang, J.; Theprungsirikul, L.; Shrift, T.; Krichilsky, B.; et al. OSCA1 mediates osmotic-stress-evoked Ca 2+ increases vital for osmosensing in Arabidopsis. Nature 2014, 514, 367–371. [Google Scholar] [CrossRef]
- Sadder, M.T.; Alsadon, A.A.; Wahb-Allah, M.A. Transcriptomic analysis of tomato lines reveals putative stress-specific biomarkers. Turk. J. Agric. For. 2014, 38, 700–715. [Google Scholar] [CrossRef]
- Hanin, M.; Brini, F.; Ebel, C.; Toda, Y.; Takeda, S.; Masmoudi, K. Plant dehydrins and stress tolerance: Versatile proteins for complex mechanisms. Plant Signal. Behav. 2011, 6, 1503–1509. [Google Scholar] [CrossRef]
- Koag, M.C.; Wilkens, S.; Fenton, R.D.; Resnik, J.; Vo, E.; Close, T.J. The K-segment of maize DHN1 mediates binding to anionic phospholipid vesicles and concomitant structural changes. Plant Physiol. 2009, 150, 1503–1514. [Google Scholar] [CrossRef]
- Rinne, P.L.; Kaikuranta, P.L.; van der Plas, L.H.; van der Schoot, C. Dehydrins in cold-acclimated apices of birch (Betula pubescens Ehrh.): Production, localization and potential role in rescuing enzyme function during dehydration. Planta 1999, 209, 377–388. [Google Scholar] [CrossRef]
- Hara, M.; Fujinaga, M.; Kuboi, T. Radical scavenging activity and oxidative modification of citrus dehydrin. Plant Physiol. Biochem. 2004, 42, 657–662. [Google Scholar] [CrossRef]
- NCBI, The National Center for Biotechnology Information. 2023. Available online: https://www.ncbi.nlm.nih.gov/ (accessed on 1 January 2023).
- Kelley, L.A.; Sternberg, M.J. Protein structure prediction on the Web: A case study using the Phyre server. Nat. Protoc. 2009, 4, 363. [Google Scholar] [CrossRef]
- Buchan, D.W.; Ward, S.M.; Lobley, A.E.; Nugent, T.C.; Bryson, K.; Jones, D.T. Protein annotation and modelling servers at University College London. Nucl. Acids Res. 2010, 38 (Suppl. S2), W563–W568. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Felsenstein, J. PHYLIP (Phylogeny Inference Package), version 3.5 c; Department of Genetics, University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Page, R.D.M. TREEVIEW: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar]
- Fernandez, M.; Águila, S.V.; Arora, R.; Chen, K. Isolation and characterization of three cold acclimation-responsive dehydrin genes from Eucalyptus globulus. Tree Genet. Genomes 2012, 8, 149–162. [Google Scholar] [CrossRef]
- Lin, C.H.; Peng, P.H.; Ko, C.Y.; Markhart, A.H.; Lin, T.Y. Characterization of a novel Y2K-type dehydrin VrDhn1 from Vigna radiata. Plant Cell Physiol. 2012, 53, 930–942. [Google Scholar] [CrossRef]
- He, Q.; Jones, D.C.; Li, W.; Xie, F.; Ma, J.; Sun, R.; Wang, Q.; Zhu, S.; Zhang, B. Genome-wide identification of R2R3-MYB genes and expression analyses during abiotic stress in Gossypium raimondii. Sci. Rep. 2016, 6, 22980. [Google Scholar] [CrossRef]
- Speth, C.; Szabo, E.X.; Martinho, C.; Collani, S.; zur Oven-Krockhaus, S.; Richter, S.; Droste-Borel, I.; Macek, B.; Stierhof, Y.D.; Schmid, M.; et al. Arabidopsis RNA processing factor SERRATE regulates the transcription of intronless genes. Elife 2018, 7, e37078. [Google Scholar] [CrossRef]
- Jeffares, D.C.; Penkett, C.J.; Bähler, J. Rapidly regulated genes are intron poor. Trends Genet. 2008, 24, 375–378. [Google Scholar] [CrossRef]
- Fang, H.; Huangfu, L.; Chen, R.; Li, P.; Xu, S.; Zhang, E.; Cao, W.; Liu, L.; Yao, Y.; Liang, G.; et al. Ancestor of land plants acquired the DNA-3-methyladenine glycosylase (MAG) gene from bacteria through horizontal gene transfer. Sci. Rep. 2017, 7, 9324. [Google Scholar] [CrossRef]
- Morozov, S.Y.; Solovyev, A.G. Emergence of intronless evolutionary forms of stress response genes: Possible relation to terrestrial adaptation of green plants. Front. Plant Sci. 2019, 10, 83. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yu, G.; Chen, X.; Fan, Z.; Liu, J.; Pan, H. Cloning and stress resistance functional characterization of an AcDHN gene from Atriplex canescens. Plant Physiol. Commun. 2012, 48, 676–682. [Google Scholar]
- Burrieza, H.P.; Koyro, H.W.; Tosar, L.M.; Kobayashi, K.; Maldonado, S. High salinity induces dehydrin accumulation in Chenopodium quinoa Willd. cv. Hualhuas embryos. Plant Soil 2012, 354, 69–79. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Dhar, S.; Banerjee, A.; Ray, S. Structural, functional, and evolutionary analysis of late embryogenesis abundant proteins (LEA) in Triticum aestivum: A detailed molecular level biochemistry using in silico approach. Comput. Biol. Chem. 2019, 82, 9–24. [Google Scholar] [CrossRef]
- Hernández-Sánchez, I.E.; Maruri-López, I.; Molphe-Balch, E.P.; Becerra-Flora, A.; Jaimes-Miranda, F.; Jiménez-Bremont, J.F. Evidence for in vivo interactions between dehydrins and the aquaporin AtPIP2B. Biochem. Biophys. Res. Commun. 2019, 510, 545–550. [Google Scholar] [CrossRef]
- Nguyen, P.N.; Tossounian, M.A.; Kovacs, D.S.; Thu, T.T.; Stijlemans, B.; Vertommen, D.; Pauwels, J.; Gevaert, K.; Angenon, G.; Messens, J.; et al. Dehydrin ERD14 activates glutathione transferase Phi9 in Arabidopsis thaliana under osmotic stress. Biochim. Biophys. Acta. Gen. Subj. 2020, 1864, 129506. [Google Scholar] [CrossRef]
- Sun, M.; Shen, Y.; Yin, K.; Guo, Y.; Cai, X.; Yang, J.; Zhu, Y.; Jia, B.; Sun, X. A late embryogenesis abundant protein GsPM30 interacts with a receptor like cytoplasmic kinase GsCBRLK and regulates environmental stress responses. Plant Sci. 2019, 283, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Feng, H.; Wen, J.; Dong, J.; Wang, T. MtCAS31 aids symbiotic nitrogen fixation by protecting the leghemoglobin MtLb120-1 under drought stress in Medicago truncatula. Front. Plant Sci. 2018, 9, 633. [Google Scholar] [CrossRef]
- Alsheikh, M.K.; Heyen, B.J.; Randall, S.K. Ion binding properties of the dehydrin ERD14 are dependent upon phosphorylation. J. Biol. Chem. 2003, 278, 40882–40889. [Google Scholar] [CrossRef]
- Sun, Z.; Li, S.; Chen, W.; Zhang, J.; Zhang, L.; Sun, W.; Wang, Z. Plant dehydrins: Expression, regulatory networks, and protective roles in plants challenged by abiotic stress. Int. J. Mol. Sci. 2021, 22, 12619. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, L.; Zhang, T.; Yang, X.; Li, D. Functional characterization of KS-type dehydrin ZmDHN13 and its related conserved domains under oxidative stress. Sci. Rep. 2017, 7, 7361. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Wang, X.; Zhang, L. Structural and functional dynamics of dehydrins: A plant protector protein under abiotic stress. Int. J. Mol. Sci. 2018, 19, 3420. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, P.; Chakrabarty, D. Dehydrin in the past four decades: From chaperones to transcription co-regulators in regulating abiotic stress response. Curr. Res. Biotechnol. 2021, 3, 249–259. [Google Scholar] [CrossRef]
- Boddington, K.F.; Graether, S.P. Binding of a Vitis riparia dehydrin to DNA. Plant Sci. 2019, 287, 110172. [Google Scholar] [CrossRef] [PubMed]
- Kadereit, G.; Mavrodiev, E.V.; Zacharias, E.H.; Sukhorukov, A.P. Molecular phylogeny of Atripliceae (Chenopodioideae, Chenopodiaceae): Implications for systematics, biogeography, flower and fruit evolution, and the origin of C4 photosynthesis. Am. J. Bot. 2010, 97, 1664–1687. [Google Scholar] [CrossRef]
- del-Pino, I.S.; Borsch, T.; Motley, T.J. trnL-F and rpl16 sequence data and dense taxon sampling reveal monophyly of unilocular anthered Gomphrenoideae (Amaranthaceae) and an improved picture of their internal relationships. Syst. Bot. 2009, 34, 57–67. [Google Scholar] [CrossRef]
- Kadereit, G.; Hohmann, S.; Kadereit, J.W. A synopsis of Chenopodiaceae subfam. Betoideae and notes on the taxonomy of Beta. Willdenowia 2006, 36, 9–19. [Google Scholar] [CrossRef]
- Müller, K.; Borsch, T. Phylogenetics of Amaranthaceae based on matK/trnK sequence data: Evidence from parsimony, likelihood, and Bayesian analyses. Ann. Mo. Bot. Gard. 2005, 92, 66–102. [Google Scholar]
- Hong, S.Y.; Cheon, K.S.; Yoo, K.O.; Lee, H.O.; Cho, K.S.; Suh, J.T.; Kim, S.J.; Nam, J.H.; Sohn, H.B.; Kim, Y.H. Complete chloroplast genome sequences and comparative analysis of Chenopodium quinoa and C. album. Front. Plant Sci. 2017, 8, 1696. [Google Scholar] [CrossRef]
- Jarvis, D.E.; Ho, Y.S.; Lightfoot, D.J.; Schmöckel, S.M.; Li, B.; Borm, T.J.; Ohyanagi, H.; Mineta, K.; Michell, C.T.; Saber, N.; et al. The genome of Chenopodium quinoa. Nature 2017, 542, 307–312. [Google Scholar] [CrossRef]
- Malik, A.A.; Veltri, M.; Boddington, K.F.; Singh, K.K.; Graether, S.P. Genome analysis of conserved dehydrin motifs in vascular plants. Front. Plant Sci. 2017, 108, 209. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yang, Y.; Xie, L.; Li, X.; Feng, C.; Chen, J.; Xu, C. Involvement of multiple types of dehydrins in the freezing response in loquat (Eriobotrya japonica). PLoS ONE 2019, 9, e87575. [Google Scholar] [CrossRef] [PubMed]
- Hinniger, C.; Caillet, V.; Michoux, F.; Ben Amor, M.; Tanksley, S.; Lin, C.; McCarthy, J. Isolation and characterization of cDNA encoding three dehydrins expressed during Coffea canephora (Robusta) grain development. Ann. Bot. 2006, 97, 755–765. [Google Scholar] [CrossRef]
- Zhang, X.H.; Moloney, M.; Chinnappa, C.C. Nucleotide sequence of a cDNA clone encoding a dehydrin-like protein from Stellaria longipes. Plant Physiol. 1993, 103, 1029. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.S.; Kim, H.Y.; Kim, Y.S.; Choi, H.G.; Kang, S.H.; Yoon, H.S. Expression of dehydrin gene from Arctic Cerastium arcticum increases abiotic stress tolerance and enhance the fermentation capacity of a genetically engineered Saccharomyces cerevisiae laboratory strain. Appl. Microbiol. Biotechnol. 2013, 97, 8997–9009. [Google Scholar] [CrossRef]
- Rosales, R.; Romero, I.; Escribano, M.I.; Merodio, C.; Sanchez-Ballesta, M.T. The crucial role of Φ-and K-segments in the in vitro functionality of Vitis vinifera dehydrin DHN1a. Phytochem 2014, 108, 17–25. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Species | Source | Lab Code | Gene | EST Accession | Protein Accession |
|---|---|---|---|---|---|---|
| 1 | Atriplex canescens | NCBI | - | AcaDHN1 | JN974246 | AFC98463 |
| 2 | Atriplex halimus | NCBI | - | AhaDHN1 | KF578414 | AGZ86543 |
| 3 | Atriplex halimus | Amman, Jordan | JO372 | AhaDHN1 | MH591427 | AYH52682 |
| 4 | Atriplex halimus | Azraq, Jordan | JO2991 | AhaDHN1 | MH591428 | AYH52683 |
| 5 | Atriplex dimorphostegia | Aqaba, Jordan | JO3111 | AdiDHN1 | MH591429 | AYH52684 |
| 6 | Atriplex halimus | Al Jouf, Saudi Arabia | KSA | AhaDHN2 | MH591430 | AYH52685 |
| 7 | Atriplex halimus | Bir Seb’a, Palestine | I4 | AhaDHN1 | MH591431 | AYH52686 |
| 8 | Atriplex leucoclada | Al-Naqab, Palestine | I5 | AleDHN1 | MH591432 | AYH52687 |
| 9 | Atriplex hortensis | Former Serbia and Montenegro | I15 | AhoDHN1 | MH591433 | AYH52688 |
| 10 | Atriplex numimularia | South Africa | I18U | AnuDHN1 | MH591434 | AYH52689 |
| I18L | AnuDHN2 | MH591435 | AYH52690 | |||
| 11 | Atriplex gardneri var. utahensis | USA | I19 | AgaDHN1 | MH591436 | AYH52691 |
| 12 | Atriplex lindleyi subsp. conduplicata | South Africa | I6 | AliDHN1 | MH591437 | AYH52692 |
| 13 | Halimione portulacoides | Egypt | E2 | HpoDHN1 | MH591438 | AYH52693 |
| 14 | Atriplex gardneri | USA | I11U | AgaDHN1 | MH591439 | AYH52694 |
| I11L | AgaDHN2 | MH591440 | AYH52695 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Musallam, A.; Abu-Romman, S.; Sadder, M.T. Molecular Characterization of Dehydrin in Azraq Saltbush among Related Atriplex Species. BioTech 2023, 12, 27. https://doi.org/10.3390/biotech12020027
Musallam A, Abu-Romman S, Sadder MT. Molecular Characterization of Dehydrin in Azraq Saltbush among Related Atriplex Species. BioTech. 2023; 12(2):27. https://doi.org/10.3390/biotech12020027
Chicago/Turabian StyleMusallam, Anas, Saeid Abu-Romman, and Monther T. Sadder. 2023. "Molecular Characterization of Dehydrin in Azraq Saltbush among Related Atriplex Species" BioTech 12, no. 2: 27. https://doi.org/10.3390/biotech12020027
APA StyleMusallam, A., Abu-Romman, S., & Sadder, M. T. (2023). Molecular Characterization of Dehydrin in Azraq Saltbush among Related Atriplex Species. BioTech, 12(2), 27. https://doi.org/10.3390/biotech12020027