

Role of the Androgen Receptor in Gender-Related Cancers

 ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Highlights
- In recent years, many studies have highlighted the involvement of sex steroid hormones, such as estrogens and androgens, and their receptors in cancer pathobiology. Their roles have been detailed in investigations of hormone-dependent cancers. However, new findings are also pointing towards their contribution to neoplasms that are not usually controlled by hormones but are identified as gender-related cancers with different incidence and mortality rates between men and women.
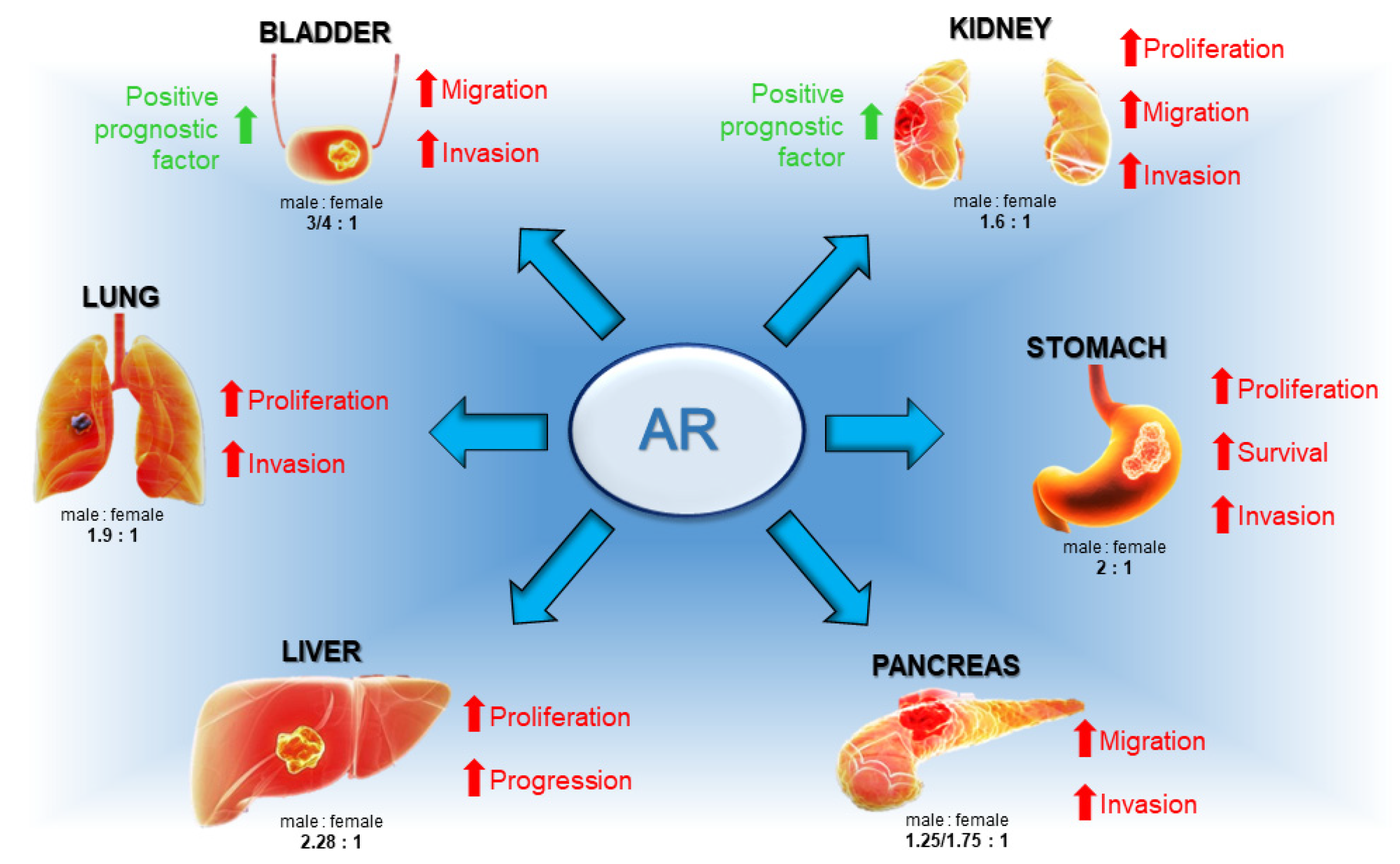
- The androgen receptor (AR) is detected in various gender-related tumors, with a higher incidence in men, and is linked to cancers such as gastric, liver, lung, bladder, kidney, and pancreatic. Although AR activity is not universally pro-tumorigenic, it frequently supports cancer progression, indicating its potential as a therapeutic target and prognostic marker in a wide array of human neoplasms.
- AR functions upon ligand activation, and understanding the impact of androgens and AR in gender-related cancers is crucial not only for developing new therapies but also for identifying at-risk groups and implementing preventive measures to reduce cancer incidence.
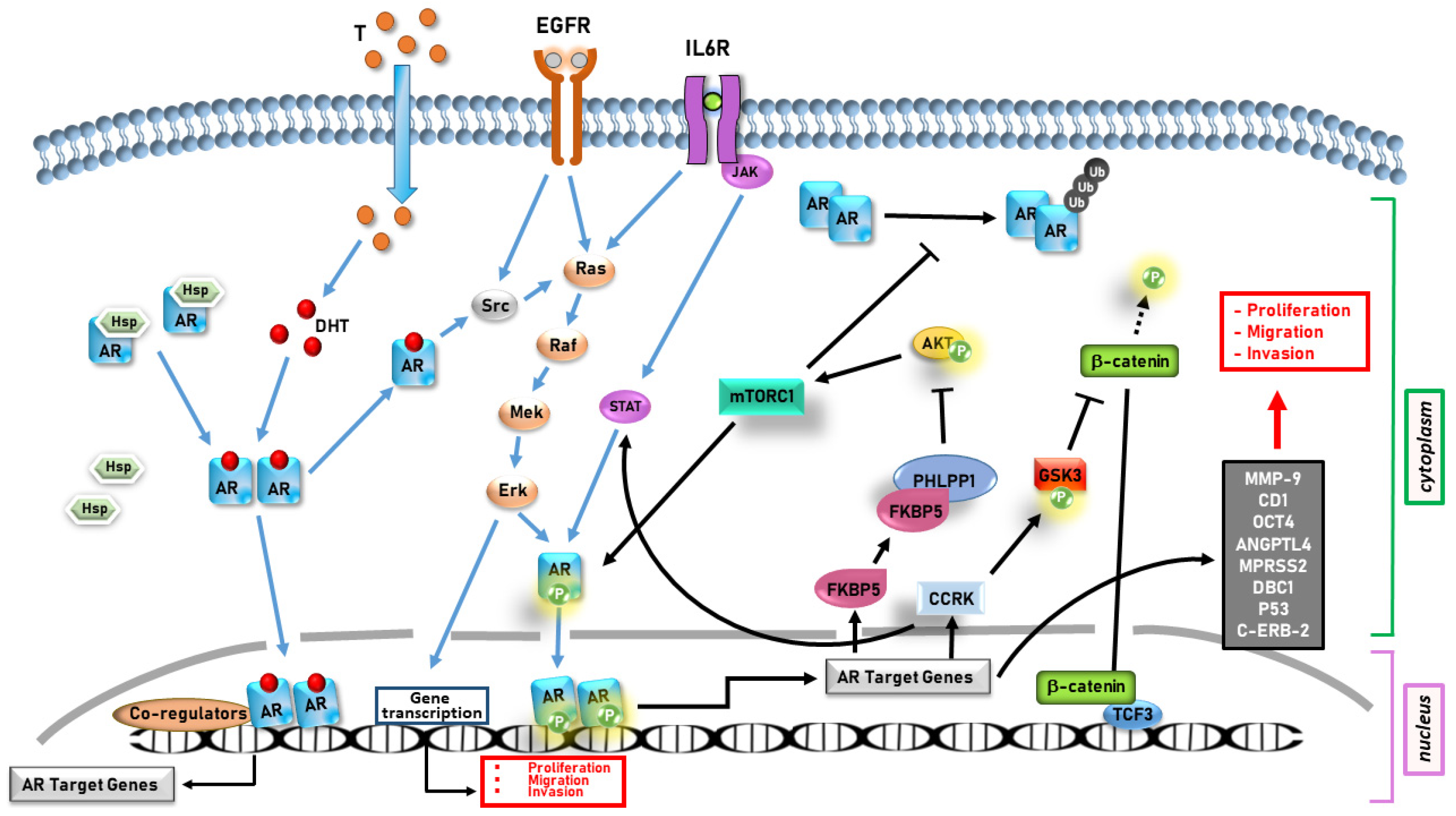
- AR appears to promote cancer progression in several gender-related cancers through both genomic and non-genomic actions, influencing cell proliferation, migration, invasion, and survival.
- Although some findings might still be considered unclear or controversial, the reviewed works highlight the importance of AR and circulating androgens, as well as the potential of AR to serve as both a therapeutic target and a prognostic marker.
- Selective androgen receptor modulators (SARMs) could complement standard treatments to enhance efficacy. Additionally, drug repurposing offers a promising approach, potentially extending therapies proven effective in hormone-dependent cancers to treat other gender-related malignancies.
Abstract
1. Introduction
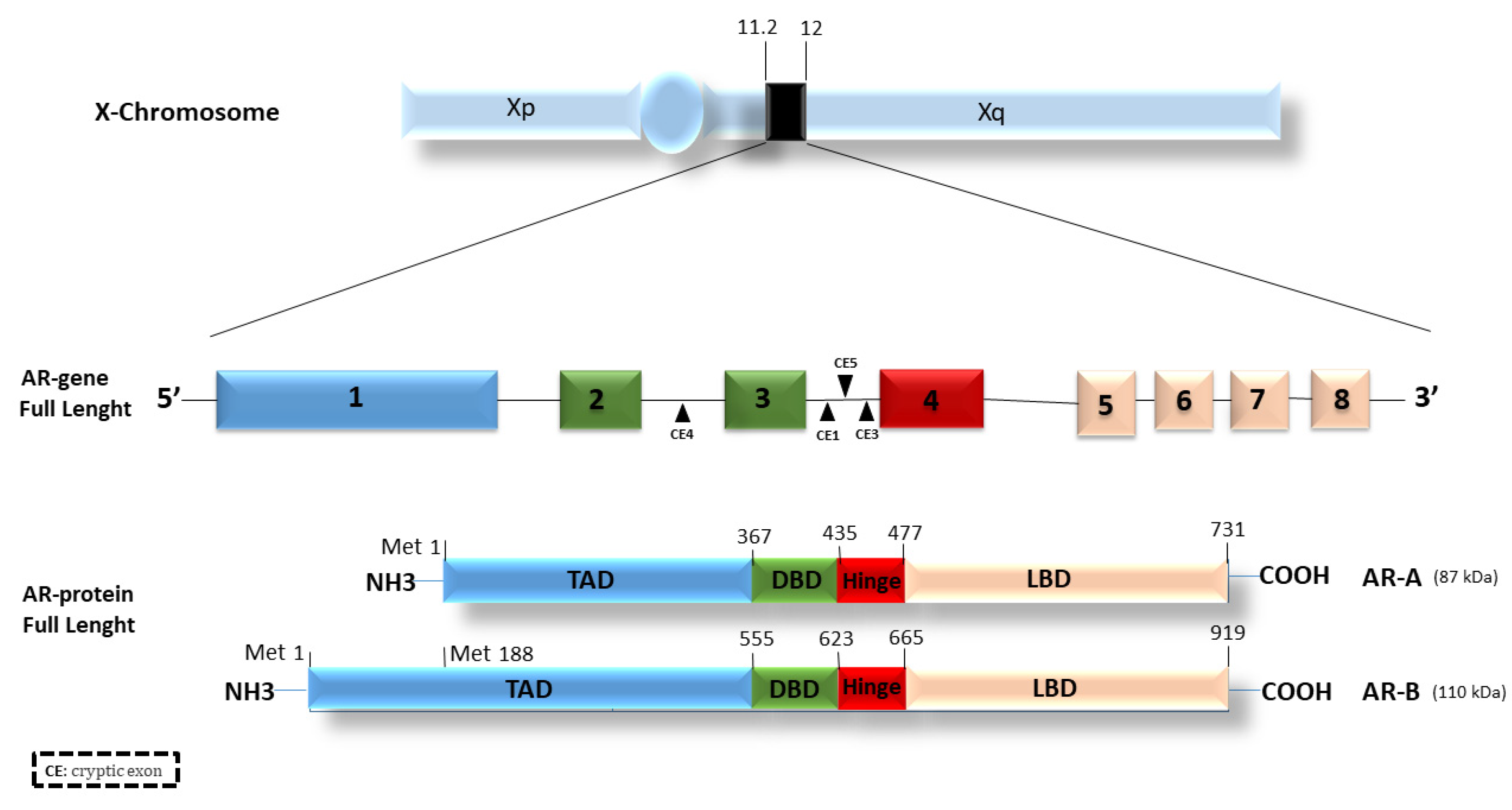
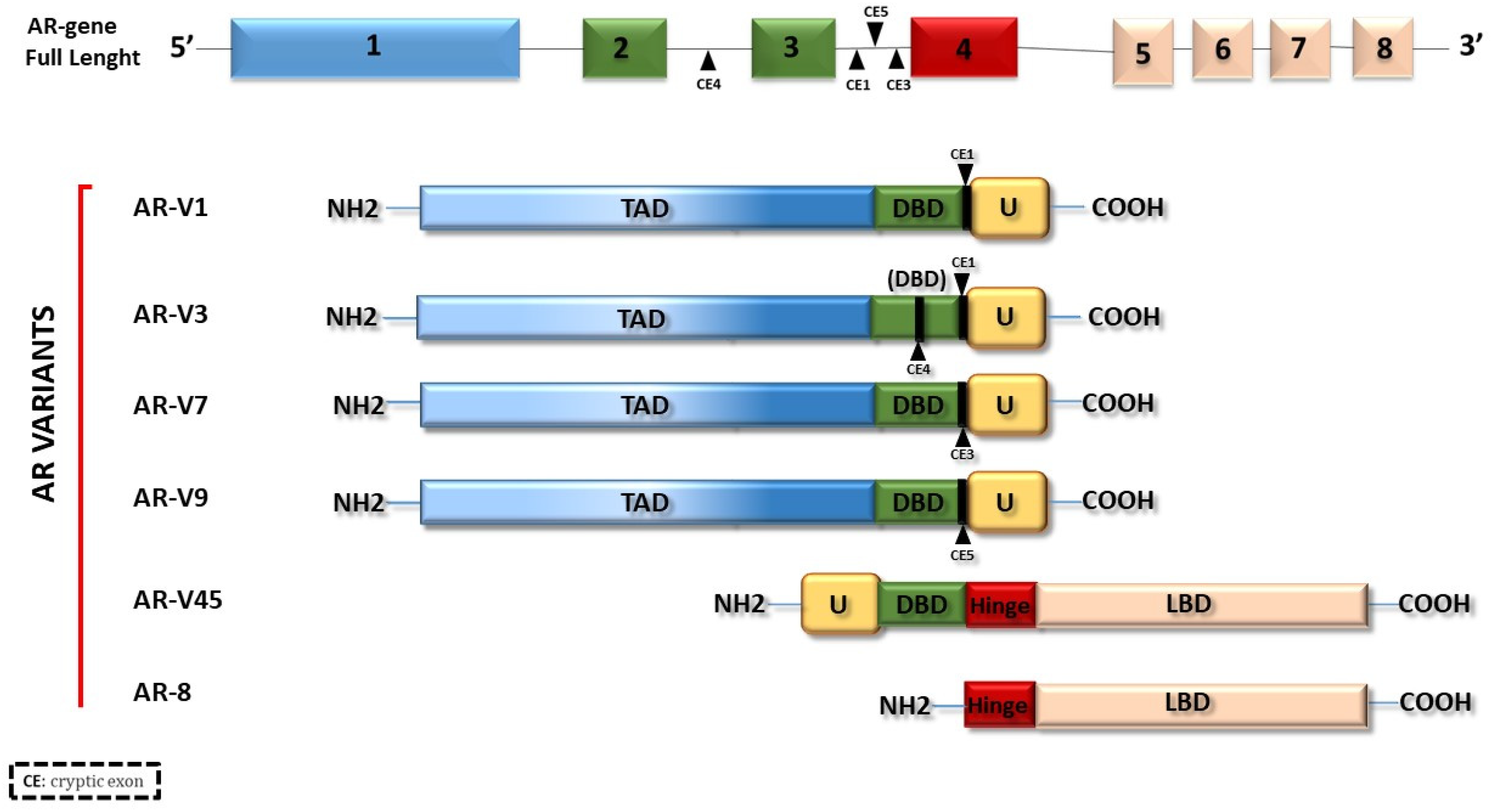
2. Androgen Receptor (AR)
3. AR in Lung Cancer
4. AR in Kidney Cancer
5. AR in Bladder Cancer
6. AR in Hepatocarcinoma
7. AR in Pancreatic Ductal Adenocarcinoma
8. AR in Gastric Cancer
9. Discussion and Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Giovannelli, P.; Ramaraj, P.; Williams, C. Editorial: Role of Sex Steroids and Their Receptor in Cancers. Front. Endocrinol. 2022, 13, 883229. [Google Scholar] [CrossRef]
- Lopes-Ramos, C.M.; Quackenbush, J.; DeMeo, D.L. Genome-Wide Sex and Gender Differences in Cancer. Front. Oncol. 2020, 10, 597788. [Google Scholar] [CrossRef] [PubMed]
- Brannon, A.R.; Haake, S.M.; Hacker, K.E.; Pruthi, R.S.; Wallen, E.M.; Nielsen, M.E.; Rathmell, W.K. Meta-Analysis of Clear Cell Renal Cell Carcinoma Gene Expression Defines a Variant Subgroup and Identifies Gender Influences on Tumor Biology. Eur. Urol. 2012, 61, 258–268. [Google Scholar] [CrossRef] [PubMed]
- Markle, J.G.; Fish, E.N. SeXX Matters in Immunity. Trends Immunol. 2014, 35, 97–104. [Google Scholar] [CrossRef]
- Giovannelli, P.; Di Donato, M.; Giraldi, T.; Migliaccio, A.; Castoria, G.; Auricchio, F. Targeting Rapid Action of Sex Steroid Receptors in Breast and Prostate Cancers. Front. Biosci. 2011, 16, 2224–2232. [Google Scholar] [CrossRef]
- Heinlein, C.A.; Chang, C. The Roles of Androgen Receptors and Androgen-Binding Proteins in Nongenomic Androgen Actions. Mol. Endocrinol. 2002, 16, 2181–2187. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.D.; Welsbie, D.S.; Tran, C.; Baek, S.H.; Chen, R.; Vessella, R.; Rosenfeld, M.G.; Sawyers, C.L. Molecular Determinants of Resistance to Antiandrogen Therapy. Nat. Med. 2004, 10, 33–39. [Google Scholar] [CrossRef]
- Helsen, C.; Van den Broeck, T.; Voet, A.; Prekovic, S.; Van Poppel, H.; Joniau, S.; Claessens, F. Androgen Receptor Antagonists for Prostate Cancer Therapy. Endocr. Relat. Cancer 2014, 21, T105–T118. [Google Scholar] [CrossRef]
- Niu, Y.; Chang, T.M.; Yeh, S.; Ma, W.L.; Wang, Y.Z.; Chang, C. Differential Androgen Receptor Signals in Different Cells Explain Why Androgen-Deprivation Therapy of Prostate Cancer Fails. Oncogene 2010, 29, 3593–3604. [Google Scholar] [CrossRef]
- Tan, M.H.; Li, J.; Xu, H.E.; Melcher, K.; Yong, E. Androgen Receptor: Structure, Role in Prostate Cancer and Drug Discovery. Acta Pharmacol. Sin. 2015, 36, 3–23. [Google Scholar] [CrossRef]
- Giovannelli, P.; Di Donato, M.; Auricchio, F.; Castoria, G.; Migliaccio, A. Androgens Induce Invasiveness of Triple Negative Breast Cancer Cells through AR/Src/PI3-K Complex Assembly. Sci. Rep. 2019, 9, 4490. [Google Scholar] [CrossRef]
- Giovannelli, P.; Di Donato, M.; Galasso, G.; Di Zazzo, E.; Bilancio, A.; Migliaccio, A. The Androgen Receptor in Breast Cancer. Front. Endocrinol. 2018, 9, 492. [Google Scholar] [CrossRef]
- Claessens, F.; Denayer, S.; Van Tilborgh, N.; Kerkhofs, S.; Helsen, C.; Haelens, A. Diverse Roles of Androgen Receptor (AR) Domains in AR-Mediated Signaling. Nucl. Recept. Signal. 2008, 6, e008. [Google Scholar] [CrossRef]
- Contrò, V.; Basile, J.R.; Proia, P. Sex Steroid Hormone Receptors, Their Ligands, and Nuclear and Non-Nuclear Pathways. AIMS Mol. Sci. 2015, 2, 294–310. [Google Scholar] [CrossRef]
- Zeng, R.; Liu, Z.; Sun, Y.; Xu, C. Differential Expression and Function of AR Isoforms in Prostate Cancer. Oncol. Rep. 2012, 27, 492–498. [Google Scholar]
- Wilson, C.M.; McPhaul, M.J. A and B Forms of the Androgen Receptor Are Expressed in a Variety of Human Tissues. Mol. Cell. Endocrinol. 1996, 120, 51–57. [Google Scholar] [CrossRef]
- Gao, T.; McPhaul, M.J. Functional Activities of the A and B Forms of the Human Androgen Receptor in Response to Androgen Receptor Agonists and Antagonists. Mol. Endocrinol. 1998, 12, 654–663. [Google Scholar] [CrossRef] [PubMed]
- Liegibel, U.M.; Sommer, U.; Boercsoek, I.; Hilscher, U.; Bierhaus, A.; Schweikert, H.U.; Nawroth, P.; Kasperk, C. Androgen Receptor Isoforms AR-A and AR-B Display Functional Differences in Cultured Human Bone Cells and Genital Skin Fibroblasts. Steroids 2003, 68, 1179–1187. [Google Scholar] [CrossRef] [PubMed]
- Catalano, M.G.; Pfeffer, U.; Raineri, M.; Ferro, P.; Curto, A.; Capuzzi, P.; Corno, F.; Berta, L.; Fortunati, N. Altered Expression of Androgen-Receptor Isoforms in Human Colon-Cancer Tissues. Int. J. Cancer 2000, 86, 325–330. [Google Scholar] [CrossRef]
- Nagandla, H.; Robertson, M.J.; Putluri, V.; Putluri, N.; Coarfa, C.; Weigel, N.L. Isoform-Specific Activities of Androgen Receptor and Its Splice Variants in Prostate Cancer Cells. Endocrinology 2021, 162, bqaa227. [Google Scholar] [CrossRef]
- Dehm, S.M.; Tindall, D.J. Alternatively Spliced Androgen Receptor Variants. Endocr. Relat. Cancer 2011, 18, R183–R196. [Google Scholar] [CrossRef]
- Yang, X.; Guo, Z.; Sun, F.; Li, W.; Alfano, A.; Shimelis, H.; Chen, M.; Brodie, A.M.H.; Chen, H.; Xiao, Z.; et al. Novel Membrane-Associated Androgen Receptor Splice Variant Potentiates Proliferative and Survival Responses in Prostate Cancer Cells. J. Biol. Chem. 2011, 286, 36152–36160. [Google Scholar] [CrossRef]
- Hara, T.; Miyazaki, J.; Araki, H.; Yamaoka, M.; Kanzaki, N.; Kusaka, M.; Miyamoto, M. Novel Mutations of Androgen Receptor: A Possible Mechanism of Bicalutamide Withdrawal Syndrome. Cancer Res. 2003, 63, 149–153. [Google Scholar] [PubMed]
- Yoshida, T.; Kinoshita, H.; Segawa, T.; Nakamura, E.; Inoue, T.; Shimizu, Y.; Kamoto, T.; Ogawa, O. Antiandrogen Bicalutamide Promotes Tumor Growth in a Novel Androgen-Dependent Prostate Cancer Xenograft Model Derived from a Bicalutamide-Treated Patient. Cancer Res. 2005, 65, 9611–9616. [Google Scholar] [CrossRef]
- Culig, Z.; Santer, F.R. Androgen Receptor Signaling in Prostate Cancer. Cancer Metastasis Rev. 2014, 33, 413–427. [Google Scholar] [CrossRef]
- Li, A.J.; McAllister, P.; Karlan, B.Y. Impact of Androgen Receptor Cytosine-Adenine-Guanine Polymorphisms on Clinical Outcome in BRCA Mutation-Associated Epithelial Ovarian Cancers. Gynecol. Oncol. 2010, 116, 105–108. [Google Scholar] [CrossRef]
- Mizushima, T.; Miyamoto, H. The Role of Androgen Receptor Signaling in Ovarian Cancer. Cells 2019, 8, 176. [Google Scholar] [CrossRef]
- Zhu, H.; Zhu, X.; Zheng, L.; Hu, X.; Sun, L.; Zhu, X. The Role of the Androgen Receptor in Ovarian Cancer Carcinogenesis and Its Clinical Implications. Oncotarget 2017, 8, 29395–29405. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.-M.; Chen, L.; Chang, W.-C.; Su, S.-Y.; Hung, Y.-C.; Ma, W.-L. Androgen/Androgen Receptor Signaling in Ovarian Cancer: Molecular Regulation and Therapeutic Potentials. Int. J. Mol. Sci. 2021, 22, 7748. [Google Scholar] [CrossRef] [PubMed]
- Calvillo-Robledo, A.; Pedernera, E.; Morales-Vásquez, F.; Pérez-Montiel, D.; Gómora, M.J.; Almaraz, M.Á.; De Alba Graue, P.G.; Rendón, E.; López-Basave, H.N.; Quintanar-Stephano, A.; et al. Simultaneous Expression of Steroid Sulfatase and Androgen Receptor Reduced Overall Survival of Patients with Epithelial Ovarian Tumors. J. Ovarian Res. 2021, 14, 98. [Google Scholar] [CrossRef]
- Manning-Geist, B.L.; Gordhandas, S.B.; Giri, D.D.; Iasonos, A.; Zhou, Q.; Girshman, J.; O’Cearbhaill, R.E.; Zamarin, D.; Lichtman, S.M.; Sabbatini, P.J.; et al. Phase II Study of Enzalutamide in Androgen Receptor Positive, Recurrent, High- and Low-Grade Serous Ovarian Cancer. Gynecol. Oncol. 2022, 164, 12–17. [Google Scholar] [CrossRef]
- Shiota, M.; Akamatsu, S.; Tsukahara, S.; Nagakawa, S.; Matsumoto, T.; Eto, M. Androgen Receptor Mutations for Precision Medicine in Prostate Cancer. Endocr. Relat. Cancer 2022, 29, R143–R155. [Google Scholar] [CrossRef]
- Konda, P.; Viswanathan, S.R. How Splicing Confers Treatment Resistance in Prostate Cancer. eLife 2022, 11, e82070. [Google Scholar] [CrossRef]
- Watson, P.A.; Chen, Y.F.; Balbas, M.D.; Wongvipat, J.; Socci, N.D.; Viale, A.; Kim, K.; Sawyers, C.L. Constitutively Active Androgen Receptor Splice Variants Expressed in Castration-Resistant Prostate Cancer Require Full-Length Androgen Receptor. Proc. Natl. Acad. Sci. USA 2010, 107, 16759–16765. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Zhan, Y.; Dong, Y. Emerging Data on Androgen Receptor Splice Variants in Prostate Cancer. Endocr. Relat. Cancer 2016, 23, T199–T210. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Luo, J. Decoding the Androgen Receptor Splice Variants. Transl. Androl. Urol. 2013, 2, 178–186. [Google Scholar] [CrossRef]
- Zhang, H.; Zhan, Y.; Liu, X.; Qi, Y.; Zhang, G.; Sartor, O.; Dong, Y. Splicing Variants of Androgen Receptor in Prostate Cancer. Am. J. Clin. Exp. Urol. 2013, 1, 18–24. [Google Scholar] [PubMed]
- Di Croce, L.; Okret, S.; Kersten, S.; Gustafsson, J.-Å.; Parker, M.; Wahli, W.; Beato, M. Steroid and Nuclear Receptors Villefranche-Sur-Mer, France, May 25–27, 1999. EMBO J. 1999, 18, 6201–6210. [Google Scholar] [CrossRef]
- Vicent, G.P.; Nacht, A.S.; Zaurín, R.; Ballaré, C.; Clausell, J.; Beato, M. Minireview: Role of Kinases and Chromatin Remodeling in Progesterone Signaling to Chromatin. Mol. Endocrinol. 2010, 24, 2088–2098. [Google Scholar] [CrossRef]
- Castoria, G.; Giovannelli, P.; Lombardi, M.; De Rosa, C.; Giraldi, T.; de Falco, A.; Barone, M.V.; Abbondanza, C.; Migliaccio, A.; Auricchio, F. Tyrosine Phosphorylation of Estradiol Receptor by Src Regulates Its Hormone-Dependent Nuclear Export and Cell Cycle Progression in Breast Cancer Cells. Oncogene 2012, 31, 4868–4877. [Google Scholar] [CrossRef] [PubMed]
- Peters, A.A.; Buchanan, G.; Ricciardelli, C.; Bianco-Miotto, T.; Centenera, M.M.; Harris, J.M.; Jindal, S.; Segara, D.; Jia, L.; Moore, N.L. Androgen Receptor Inhibits Estrogen Receptor-α Activity and Is Prognostic in Breast Cancer. Cancer Res. 2009, 69, 6131–6140. [Google Scholar] [CrossRef]
- Alberg, A.J.; Brock, M.V.; Ford, J.G.; Samet, J.M.; Spivack, S.D. Epidemiology of Lung Cancer: Diagnosis and Management of Lung Cancer: American College of Chest Physicians Evidence-Based Clinical Practice Guidelines. Chest 2013, 143, e1S–e29S. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Center, M.M.; DeSantis, C.; Ward, E.M. Global Patterns of Cancer Incidence and Mortality Rates and TrendsGlobal Patterns of Cancer. Cancer Epidemiol. Biomarkers Prev. 2010, 19, 1893–1907. [Google Scholar] [CrossRef]
- Stabellini, N.; Bruno, D.S.; Dmukauskas, M.; Barda, A.J.; Cao, L.; Shanahan, J.; Waite, K.; Montero, A.J.; Barnholtz-Sloan, J.S. Sex Differences in Lung Cancer Treatment and Outcomes at a Large Hybrid Academic-Community Practice. JTO Clin. Res. Rep. 2022, 3, 100307. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Wagle, N.S.; Jemal, A. Cancer Statistics, 2023. CA Cancer J. Clin. 2023, 73, 17–48. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.B.; Kau, T.Y.; Severson, R.K.; Kalemkerian, G.P. Lung Cancer in Women: Analysis of the National Surveillance, Epidemiology, and End Results Database. Chest 2005, 127, 768–777. [Google Scholar] [CrossRef]
- Investigators, I.E.L.C.A.P. Survival of Patients with Stage I Lung Cancer Detected on CT Screening. N. Engl. J. Med. 2006, 355, 1763–1771. [Google Scholar]
- Mikkonen, L.; Pihlajamaa, P.; Sahu, B.; Zhang, F.-P.; Jänne, O.A. Androgen Receptor and Androgen-Dependent Gene Expression in Lung. Mol. Cell. Endocrinol. 2010, 317, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Migliaccio, A.; Castoria, G.; DOMENICO, M.D.; Ciociola, A.; Lombardi, M.; De Falco, A.; Nanayakkara, M.; Bottero, D.; De Stasio, R.; Varricchio, L. Crosstalk between EGFR and Extranuclear Steroid Receptors. Ann. N. Y. Acad. Sci. 2006, 1089, 194–200. [Google Scholar] [CrossRef]
- Recchia, A.G.; Musti, A.M.; Lanzino, M.; Panno, M.L.; Turano, E.; Zumpano, R.; Belfiore, A.; Andò, S.; Maggiolini, M. A Cross-Talk between the Androgen Receptor and the Epidermal Growth Factor Receptor Leads to P38MAPK-Dependent Activation of MTOR and CyclinD1 Expression in Prostate and Lung Cancer Cells. Int. J. Biochem. Cell Biol. 2009, 41, 603–614. [Google Scholar] [CrossRef]
- Lu, H.-H.; Yeh, S.-D.; Chou, Y.-T.; Tsai, Y.-T.; Chang, C.; Wu, C.-W. Androgen Receptor Regulates Lung Cancer Progress through Modulation of OCT-4 Expression. Cancer Res. 2011, 71, 2126. [Google Scholar] [CrossRef]
- Padua, D.; Zhang, X.H.-F.; Wang, Q.; Nadal, C.; Gerald, W.L.; Gomis, R.R.; Massagué, J. TGFβ Primes Breast Tumors for Lung Metastasis Seeding through Angiopoietin-like 4. Cell 2008, 133, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, W.; Liu, X.S.; Carroll, J.S.; Jänne, O.A.; Keeton, E.K.; Chinnaiyan, A.M.; Pienta, K.J.; Brown, M. A Hierarchical Network of Transcription Factors Governs Androgen Receptor-Dependent Prostate Cancer Growth. Mol. Cell 2007, 27, 380–392. [Google Scholar] [CrossRef]
- Kumar-Sinha, C.; Tomlins, S.A.; Chinnaiyan, A.M. Recurrent Gene Fusions in Prostate Cancer. Nat. Rev. Cancer 2008, 8, 497–511. [Google Scholar] [CrossRef]
- Perner, S.; Demichelis, F.; Beroukhim, R.; Schmidt, F.H.; Mosquera, J.-M.; Setlur, S.; Tchinda, J.; Tomlins, S.A.; Hofer, M.D.; Pienta, K.G.; et al. TMPRSS2:ERG Fusion-Associated Deletions Provide Insight into the Heterogeneity of Prostate Cancer. Cancer Res. 2006, 66, 8337–8341. [Google Scholar] [CrossRef]
- Lara, P.N.; Heilmann, A.M.; Elvin, J.A.; Parikh, M.; De Vere White, R.; Gandour-Edwards, R.; Evans, C.P.; Pan, C.-X.; Schrock, A.B.; Erlich, R.; et al. TMPRSS2-ERG Fusions Unexpectedly Identified in Men Initially Diagnosed with Nonprostatic Malignancies. JCO Precis. Oncol. 2017, 1, 1–6. [Google Scholar] [CrossRef]
- Gockel, L.M.; Pfeifer, V.; Baltes, F.; Bachmaier, R.D.; Wagner, K.G.; Bendas, G.; Gütschow, M.; Sosič, I.; Steinebach, C. Design, Synthesis, and Characterization of PROTACs Targeting the Androgen Receptor in Prostate and Lung Cancer Models. Arch. Pharm. 2022, 355, 2100467. [Google Scholar] [CrossRef] [PubMed]
- Yeh, S.-D.; Yang, P.-C.; Lu, H.-H.; Chang, C.; Wu, C.-W. Targeting Androgen Receptor as a New Potential Therapeutic Approach to Battle Tobacco Carcinogens-Induced Non-Small Cell Lung Cancer. J. Transl. Med. 2012, 10, A8. [Google Scholar] [CrossRef]
- Jia, X.; Han, X. Targeting Androgen Receptor Degradation with PROTACs from Bench to Bedside. Biomed. Pharmacother. 2023, 158, 114112. [Google Scholar] [CrossRef]
- PROTAC Shrinks Mutated Prostate Tumors. Cancer Discov. 2022, 12, OF2. [CrossRef]
- Békés, M.; Langley, D.R.; Crews, C.M. PROTAC Targeted Protein Degraders: The Past Is Prologue. Nat. Rev. Drug Discov. 2022, 21, 181–200. [Google Scholar] [CrossRef]
- Compérat, E.; Camparo, P. Histological Classification of Malignant Renal Tumours at a Time of Major Diagnostic and Therapeutic Changes. Diagn. Interv. Imaging 2012, 93, 221–231. [Google Scholar] [CrossRef]
- Gupta, K.; Miller, J.D.; Li, J.Z.; Russell, M.W.; Charbonneau, C. Epidemiologic and Socioeconomic Burden of Metastatic Renal Cell Carcinoma (MRCC): A Literature Review. Cancer Treat. Rev. 2008, 34, 193–205. [Google Scholar] [CrossRef]
- Giannarini, G.; Nguyen, D.P.; Thalmann, G.N.; Thoeny, H.C. Diffusion-Weighted Magnetic Resonance Imaging Detects Local Recurrence after Radical Prostatectomy: Initial Experience. Eur. Urol. 2012, 61, 616–620. [Google Scholar] [CrossRef] [PubMed]
- Orth, S.R. Cigarette Smoking: An Important Renal Risk Factor–Far beyond Carcinogenesis. Tob. Induc. Dis. 2003, 1, 137. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Li, L.; Zhu, G.; Liang, L.; Guan, Z.; Chang, L.; Chen, Y.; Yeh, S.; Chang, C. ASC-J9 Suppresses Renal Cell Carcinoma Progression by Targeting an Androgen Receptor–Dependent HIF2α/VEGF Signaling PathwayTargeting AR to Suppress Kidney Cancer Progression. Cancer Res. 2014, 74, 4420–4430. [Google Scholar] [CrossRef]
- Noh, S.J.; Kang, M.J.; Kim, K.M.; Bae, J.S.; Park, H.S.; Moon, W.S.; Chung, M.J.; Lee, H.; Lee, D.G.; Jang, K.Y. Acetylation Status of P53 and the Expression of DBC1, SIRT1, and Androgen Receptor Are Associated with Survival in Clear Cell Renal Cell Carcinoma Patients. Pathology 2013, 45, 574–580. [Google Scholar] [CrossRef]
- Bialek, J.; Piwonka, M.; Kawan, F.; Fornara, P.; Theil, G. Differential Expression of the Androgen Receptor, Splice Variants and Relaxin 2 in Renal Cancer. Life 2021, 11, 731. [Google Scholar] [CrossRef] [PubMed]
- Langner, C.; Ratschek, M.; Rehak, P.; Schips, L.; Zigeuner, R. Steroid Hormone Receptor Expression in Renal Cell Carcinoma: An Immunohistochemical Analysis of 182 Tumors. J. Urol. 2004, 171, 611–614. [Google Scholar] [CrossRef]
- Yuan, P.; Ge, Y.; Liu, X.; Wang, S.; Ye, Z.; Xu, H.; Chen, Z. The Association of Androgen Receptor Expression with Renal Cell Carcinoma Risk: A Systematic Review and Meta-Analysis. Pathol. Oncol. Res. 2020, 26, 605–614. [Google Scholar] [CrossRef]
- Gul, A.; Rini, B.I. Adjuvant Therapy in Renal Cell Carcinoma. Cancer 2019, 125, 2935–2944. [Google Scholar] [CrossRef]
- Motzer, R.J.; Michaelson, M.D.; Redman, B.G.; Hudes, G.R.; Wilding, G.; Figlin, R.A.; Ginsberg, M.S.; Kim, S.T.; Baum, C.M.; DePrimo, S.E. Activity of SU11248, a Multitargeted Inhibitor of Vascular Endothelial Growth Factor Receptor and Platelet-Derived Growth Factor Receptor, in Patients with Metastatic Renal Cell Carcinoma. J. Clin. Oncol. 2006, 24, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Motzer, R.J.; Russo, P. Systemic Therapy for Renal Cell Carcinoma. J. Urol. 2000, 163, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Boorman, G.A. Animal Model of Human Disease: Carcinoma of the Ureter and Urinary Bladder. Am. J. Pathol. 1977, 88, 251. [Google Scholar]
- Wang, C.-S.; Li, C.-C.; Juan, Y.-S.; Wu, W.-J.; Lee, H.-Y. 5α-Reductase Inhibitors Impact Prognosis of Urothelial Carcinoma. BMC Cancer 2020, 20, 872. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Murray, T.; Samuels, A. I Wsp. Cancer Statistics. CA Cancer J. Clin. 2003, 53, 5–26. [Google Scholar] [CrossRef] [PubMed]
- Imada, S.; Akaza, H.; Ami, Y.; Koiso, K.; Ideyama, Y.; Takenaka, T. Promoting Effects and Mechanisms of Action of Androgen in Bladder Carcinogenesis in Male Rats. Eur. Urol. 1997, 31, 360–364. [Google Scholar] [CrossRef]
- Okajima, E.; Hiramatsu, T.; Iriya, K.; Ijuin, M.; Matsushima, S.; Yamada, K. Effect of Sex Hormones on Development of Urinary Bladder Tumours in Rats Induced by N-Butyl-N-(4-Hydroxybutyl) Nitrosamine. Urol. Res. 1975, 3, 73–79. [Google Scholar] [CrossRef]
- Filippi, S.; Morelli, A.; Sandner, P.; Fibbi, B.; Mancina, R.; Marini, M.; Gacci, M.; Vignozzi, L.; Vannelli, G.B.; Carini, M. Characterization and Functional Role of Androgen-Dependent PDE5 Activity in the Bladder. Endocrinology 2007, 148, 1019–1029. [Google Scholar] [CrossRef]
- Juan, Y.-S.; Onal, B.; Broadaway, S.; Cosgrove, J.; Leggett, R.E.; Whitbeck, C.; De, E.; Sokol, R.; Levin, R.M. Effect of Castration on Male Rabbit Lower Urinary Tract Tissue Enzymes. Mol. Cell. Biochem. 2007, 301, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Laor, E.; Schiffman, Z.J.; Braunstein, J.D.; Reid, R.E.; Tolia, B.M.; Koss, L.G.; Freed, S.Z. Androgen Receptors in Bladder Tumors. Urology 1985, 25, 161–163. [Google Scholar] [CrossRef]
- Boorjian, S.; Ugras, S.; Mongan, N.P.; Gudas, L.J.; You, X.; Tickoo, S.K.; Scherr, D.S. Androgen Receptor Expression Is Inversely Correlated with Pathologic Tumor Stage in Bladder Cancer. Urology 2004, 64, 383–388. [Google Scholar] [CrossRef]
- Dai, S.-L.; Wei, S.-S.; Zhang, C.; Li, X.-Y.; Liu, Y.-P.; Ma, M.; Lv, H.-L.; Zhang, Z.; Zhao, L.-M.; Shan, B.-E. MTA2 Promotes the Metastasis of Esophageal Squamous Cell Carcinoma via EIF4E-Twist Feedback Loop. Cancer Sci. 2021, 112, 1060–1074. [Google Scholar] [CrossRef]
- Leung, J.K.; Sadar, M.D. Non-Genomic Actions of the Androgen Receptor in Prostate Cancer. Front. Endocrinol. 2017, 8, 2. [Google Scholar] [CrossRef]
- Zhang, Y.; Ou, D.; Zhuang, D.; Zheng, Z.; Lin, M. In Silico Analysis of the Immune Microenvironment in Bladder Cancer. BMC Cancer 2020, 20, 265. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, Y.-H.; Bläuer, M.; Tammela, T.; Tuohimaa, P. Immunodetection of Androgen Receptor in Human Urinary Bladder Cancer. Histopathology 1997, 30, 556–562. [Google Scholar] [CrossRef]
- Ou, Z.; Wang, Y.; Liu, L.; Li, L.; Yeh, S.; Qi, L.; Chang, C. Tumor Microenvironment B Cells Increase Bladder Cancer Metastasis via Modulation of the IL-8/Androgen Receptor (AR)/MMPs Signals. Oncotarget 2015, 6, 26065. [Google Scholar] [CrossRef]
- Shiota, M.; Kiyoshima, K.; Yokomizo, A.; Takeuchi, A.; Kashiwagi, E.; Dejima, T.; Takahashi, R.; Inokuchi, J.; Tatsugami, K.; Eto, M. Suppressed Recurrent Bladder Cancer after Androgen Suppression with Androgen Deprivation Therapy or 5α-Reductase Inhibitor. J. Urol. 2017, 197, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Kim, M.S.; Ahn, J.-H.; Choi, W.S.; Park, H.K.; Kim, H.G.; Paick, S.H. Clinical Significance of 5-α Reductase Inhibitor and Androgen Deprivation Therapy in Bladder Cancer Incidence, Recurrence, and Survival: A Meta-Analysis and Systemic Review. Aging Male 2020, 23, 971–978. [Google Scholar] [CrossRef]
- Izumi, K.; Ito, Y.; Miyamoto, H.; Miyoshi, Y.; Ota, J.; Moriyama, M.; Murai, T.; Hayashi, H.; Inayama, Y.; Ohashi, K.; et al. Expression of Androgen Receptor in Non-Muscle-Invasive Bladder Cancer Predicts the Preventive Effect of Androgen Deprivation Therapy on Tumor Recurrence. Oncotarget 2016, 7, 14153–14160. [Google Scholar] [CrossRef]
- Wu, S.-C.; Kwon, D.; Jue, J.S.; Chen, F.V.; Velasquez Escobar, M.C.; Punnen, S.; Parekh, D.J.; Ritch, C.R.; Gonzalgo, M.L. Androgen Suppression Therapy Is Associated with Lower Recurrence of Non–Muscle-Invasive Bladder Cancer. Eur. Urol. Focus 2021, 7, 142–147. [Google Scholar] [CrossRef]
- Moschini, M.; Zaffuto, E.; Karakiewicz, P.; Mattei, A.; Gandaglia, G.; Fossati, N.; Montorsi, F.; Briganti, A.; Shariat, S.F. The Effect of Androgen Deprivation Treatment on Subsequent Risk of Bladder Cancer Diagnosis in Male Patients Treated for Prostate Cancer. World J. Urol. 2019, 37, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Santella, C.; Rouette, J.; Brundage, M.D.; Filion, K.B.; Azoulay, L. Androgen Deprivation Therapy for Prostate Cancer and the Risk of Bladder Cancer: A Systematic Review of Observational Studies. Urol. Oncol. Semin. Orig. Investig. 2020, 38, 816–825. [Google Scholar] [CrossRef] [PubMed]
- Kashiwagi, E.; Ide, H.; Inoue, S.; Kawahara, T.; Zheng, Y.; Reis, L.O.; Baras, A.S.; Miyamoto, H. Androgen Receptor Activity Modulates Responses to Cisplatin Treatment in Bladder Cancer. Oncotarget 2016, 7, 49169. [Google Scholar] [CrossRef] [PubMed]
- Kameyama, K.; Horie, K.; Mizutani, K.; Kato, T.; Fujita, Y.; Kawakami, K.; Kojima, T.; Miyazaki, T.; Deguchi, T.; Ito, M. Enzalutamide Inhibits Proliferation of Gemcitabine-Resistant Bladder Cancer Cells with Increased Androgen Receptor Expression. Int. J. Oncol. 2017, 50, 75–84. [Google Scholar] [CrossRef]
- Inoue, S.; Mizushima, T.; Miyamoto, H. Role of the Androgen Receptor in Urothelial Cancer. Mol. Cell. Endocrinol. 2018, 465, 73–81. [Google Scholar] [CrossRef]
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular Carcinoma. Nat. Rev. Dis. Primer 2021, 7, 6. [Google Scholar] [CrossRef]
- Acosta-Lopez, S.; Diaz-Bethencourt, D.; Concepcion-Massip, T.; Martin-Fernandez de Basoa, M.C.; Plata-Bello, A.; Gonzalez-Rodriguez, A.; Perez-Hernandez, F.; Plata-Bello, J. The Androgen Receptor Expression and Its Activity Have Different Relationships with Prognosis in Hepatocellular Carcinoma. Sci. Rep. 2020, 10, 22046. [Google Scholar] [CrossRef]
- Wu, M.-H.; Ma, W.-L.; Hsu, C.-L.; Chen, Y.-L.; Ou, J.-H.J.; Ryan, C.K.; Hung, Y.-C.; Yeh, S.; Chang, C. Androgen Receptor Promotes Hepatitis B Virus–Induced Hepatocarcinogenesis through Modulation of Hepatitis B Virus RNA Transcription. Sci. Transl. Med. 2010, 2, 32ra35. [Google Scholar] [CrossRef]
- Ren, Q.-N.; Zhang, H.; Sun, C.-Y.; Zhou, Y.-F.; Yang, X.-F.; Long, J.-W.; Li, X.-X.; Mai, S.-J.; Zhang, M.-Y.; Zhang, H.-Z. Phosphorylation of Androgen Receptor by MTORC1 Promotes Liver Steatosis and Tumorigenesis. Hepatology 2022, 75, 1123–1138. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, X.-X.; Yang, Y.; Zhang, Y.; Wang, H.-Y.; Zheng, X.S. Significance and Mechanism of Androgen Receptor Overexpression and Androgen Receptor/Mechanistic Target of Rapamycin Cross-talk in Hepatocellular Carcinoma. Hepatology 2018, 67, 2271–2286. [Google Scholar] [CrossRef] [PubMed]
- Dimri, M.; Satyanarayana, A. Molecular Signaling Pathways and Therapeutic Targets in Hepatocellular Carcinoma. Cancers 2020, 12, 491. [Google Scholar] [CrossRef]
- Cheng, Y.-W.; Chen, K.-W.; Kuo, H.-C.; Kuo, C.-H.; Lin, W.-H.; Chen, P.-J.; Yeh, S.-H. Specific Diacylglycerols Generated by Hepatic Lipogenesis Stimulate the Oncogenic Androgen Receptor Activity in Male Hepatocytes. Int. J. Obes. 2019, 43, 2469–2479. [Google Scholar] [CrossRef]
- Feng, H.; Cheng, A.S.; Tsang, D.P.; Li, M.S.; Go, M.Y.; Cheung, Y.S.; Zhao, G.; Ng, S.S.; Lin, M.C.; Yu, J. Cell Cycle–Related Kinase Is a Direct Androgen Receptor–Regulated Gene That Drives β-Catenin/T Cell Factor–Dependent Hepatocarcinogenesis. J. Clin. Investig. 2011, 121, 3159–3175. [Google Scholar] [CrossRef]
- Sun, H.; Yang, W.; Tian, Y.; Zeng, X.; Zhou, J.; Mok, M.T.; Tang, W.; Feng, Y.; Xu, L.; Chan, A.W. An Inflammatory-CCRK Circuitry Drives MTORC1-Dependent Metabolic and Immunosuppressive Reprogramming in Obesity-Associated Hepatocellular Carcinoma. Nat. Commun. 2018, 9, 5214. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Yang, D.; Yin, C.; Zhang, J. Androgen Receptor (AR)-TLR4 Crosstalk Mediates Gender Disparities in Hepatocellular Carcinoma Incidence and Progression. J. Cancer 2020, 11, 1094. [Google Scholar] [CrossRef]
- Groupe d’Etude et de Traitement du Carcinome Hépatocellulaire (GRETCH). Randomized Trial of Leuprorelin and Flutamide in Male Patients with Hepatocellular Carcinoma Treated with Tamoxifen. Hepatology 2004, 40, 1361–1369. [Google Scholar] [CrossRef] [PubMed]
- Manesis, E.K.; Giannoulis, G.; Zoumboulis, P.; Vafiadou, I.; Hadziyannis, S.J. Treatment of Hepatocellular Carcinoma with Combined Suppression and Inhibition of Sex Hormones: A Randomized, Controlled Trial. Hepatology 1995, 21, 1535–1542. [Google Scholar]
- Jiang, G.; Shi, L.; Zheng, X.; Zhang, X.; Wu, K.; Liu, B.; Yan, P.; Liang, X.; Yu, T.; Wang, Y. Androgen Receptor Affects the Response to Immune Checkpoint Therapy by Suppressing PD-L1 in Hepatocellular Carcinoma. Aging 2020, 12, 11466. [Google Scholar] [CrossRef]
- Andrén-Sandberg, Å.; Hoem, D.; Bäckman, P.L. Other Risk Factors for Pancreatic Cancer: Hormonal Aspects. Ann. Oncol. 1999, 10, S131–S135. [Google Scholar] [CrossRef]
- Kreiger, N.; Lacroix, J.; Sloan, M. Hormonal Factors and Pancreatic Cancer in Women. Ann. Epidemiol. 2001, 11, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Konduri, S.; Schwarz, M.A.; Cafasso, D.; Schwarz, R.E. Androgen Receptor Blockade in Experimental Combination Therapy of Pancreatic Cancer. J. Surg. Res. 2007, 142, 378–386. [Google Scholar] [CrossRef] [PubMed]
- Nacusi, L.P.; Debes, J.D. Primers on Molecular Pathways: Nuclear Receptors in Pancreatic Cancer: The Ligand-Independent Way. Pancreatology 2008, 8, 422–424. [Google Scholar] [CrossRef]
- Selvan, R.S.; Metzgar, R.S.; Petrow, V. Growth Modulatory Effects of Some 6-Methylenic Steroids on Human and Hamster Pancreatic Adenocarcinoma Cells in Vitro. Drug Des. Discov. 1992, 9, 119–133. [Google Scholar]
- Mahipal, A.; Tella, S.H.; Kommalapati, A.; Goyal, G.; Soares, H.; Neuger, A.; Copolla, D.; Kim, J.; Kim, R. Phase 1 Trial of Enzalutamide in Combination with Gemcitabine and Nab-Paclitaxel for the Treatment of Advanced Pancreatic Cancer. Investig. New Drugs 2019, 37, 473–481. [Google Scholar] [CrossRef]
- Noh, K.W.; Pungpapong, S.; Wallace, M.B.; Woodward, T.A.; Raimondo, M. Do Cytokine Concentrations in Pancreatic Juice Predict the Presence of Pancreatic Diseases? Clin. Gastroenterol. Hepatol. 2006, 4, 782–789. [Google Scholar] [CrossRef]
- Okitsu, K.; Kanda, T.; Imazeki, F.; Yonemitsu, Y.; Ray, R.B.; Chang, C.; Yokosuka, O. Involvement of Interleukin-6 and Androgen Receptor Signaling in Pancreatic Cancer. Genes Cancer 2010, 1, 859–867. [Google Scholar] [CrossRef]
- Ueda, T.; Bruchovsky, N.; Sadar, M.D. Activation of the Androgen Receptor N-Terminal Domain by Interleukin-6 via MAPK and STAT3 Signal Transduction Pathways. J. Biol. Chem. 2002, 277, 7076–7085. [Google Scholar] [CrossRef] [PubMed]
- Correa, P. Gastric Cancer: Overview. Gastroenterol. Clin. 2013, 42, 211–217. [Google Scholar] [CrossRef]
- Karimi, P.; Islami, F.; Anandasabapathy, S.; Freedman, N.D.; Kamangar, F. Gastric Cancer: Descriptive Epidemiology, Risk Factors, Screening, and PreventionGastric Cancer. Cancer Epidemiol. Biomark. Prev. 2014, 23, 700–713. [Google Scholar] [CrossRef]
- Orditura, M.; Galizia, G.; Sforza, V.; Gambardella, V.; Fabozzi, A.; Laterza, M.M.; Andreozzi, F.; Ventriglia, J.; Savastano, B.; Mabilia, A. Treatment of Gastric Cancer. World J. Gastroenterol. 2014, 20, 1635. [Google Scholar] [CrossRef]
- Fard, S.S.; Yazdanbod, M.; Sotoudeh, M.; Bashash, D.; Mahmoodzadeh, H.; Saliminejad, K.; Mousavi, S.A.; Ghaffari, S.H.; Alimoghaddam, K. Prognostic and Therapeutic Significance of Androgen Receptor in Patients with Gastric Cancer. OncoTargets Ther. 2020, 13, 9821. [Google Scholar] [CrossRef]
- Wu, C.-W.; Chi, C.-W.; Chang, T.-J.; Lui, W.-Y.; P’eng, F.-K. Sex Hormone Receptors in Gastric Cancer. Cancer 1990, 65, 1396–1400. [Google Scholar] [CrossRef]
- Zhang, B.; Du, T.; Chang, Q.; Fan, Z.; Li, J.; Yu, B.; Su, L.; Li, C.; Yan, C.; Gu, Q. Androgen Receptor Promotes Gastric Cancer Cell Migration and Invasion via AKT-Phosphorylation Dependent Upregulation of Matrix Metalloproteinase 9. Oncotarget 2014, 5, 10584. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Liu, R.; Yan, Y.; Pan, X.; Wang, M.; Han, X.; Ren, H.; Zhang, Z. Expression of Estrogen Receptors and Androgen Receptor and Their Clinical Significance in Gastric Cancer. Oncotarget 2017, 8, 40765–40777. [Google Scholar] [CrossRef] [PubMed]
- Fard, S.S.; Sotoudeh, M.; Yazdanbod, M.; Ghavamzadeh, A.; Malekzadeh, R.; Yaghmaie, M.; Mousavi, S.A.; Ghaffari, S.H.; Alimoghaddam, K. Evaluation of the Association between Androgen Receptor and AURKA and Its Prognostic Value in Gastric Cancer. Int. J. Hematol. Oncol. Stem Cell Res. 2019, 13, 174. [Google Scholar]
- Peng, L.; Li, Y.; Wei, S.; Li, X.; Dang, Y.; Zhang, W.; Zhang, G. LAMA4 Activated by Androgen Receptor Induces the Cisplatin Resistance in Gastric Cancer. Biomed. Pharmacother. 2020, 124, 109667. [Google Scholar] [CrossRef]
- Zhao, L.; Zhou, S.; Gustafsson, J.-Å. Nuclear Receptors: Recent Drug Discovery for Cancer Therapies. Endocr. Rev. 2019, 5, 1207–1249. [Google Scholar] [CrossRef]
- Deng, G.; Wang, R.; Sun, Y.; Huang, C.-P.; Yeh, S.; You, B.; Feng, C.; Li, G.; Ma, S.; Chang, C. Targeting Androgen Receptor (AR) with Antiandrogen Enzalutamide Increases Prostate Cancer Cell Invasion yet Decreases Bladder Cancer Cell Invasion via Differentially Altering the AR/CircRNA-ARC1/MiR-125b-2-3p or MiR-4736/PPARγ/MMP-9 Signals. Cell Death Differ. 2021, 28, 2145–2159. [Google Scholar] [CrossRef]
- Besançon, M.; Gris, T.; Joncas, F.-H.; Picard, V.; Bergeron, A.; Fradet, Y.; Toren, P. Combining Antiandrogens with Immunotherapy for Bladder Cancer Treatment. Eur. Urol. Open Sci. 2022, 43, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Longo, J.; Van Leeuwen, J.E.; Elbaz, M.; Branchard, E.; Penn, L.Z. Statins as Anticancer Agents in the Era of Precision Medicine. Clin. Cancer Res. 2020, 26, 5791–5800. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Cheng, C.; Wang, Y.; Chen, T.; Tu, J.; Niu, C.; Xing, R.; Wang, Y.; Xu, Y. Synergistic Effect of Statins and Abiraterone Acetate on the Growth Inhibition of Neuroblastoma via Targeting Androgen Receptor. Front. Oncol. 2021, 11, 595285. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.Z.; Chang, J.I.; Li, E.; Xiang, A.H.; Wu, B.U. Influence of Statins and Cholesterol on Mortality Among Patients with Pancreatic Cancer. J. Natl. Cancer Inst. 2017, 109, djw275. [Google Scholar] [CrossRef] [PubMed]
- Reese, J.M.; Bruinsma, E.S.; Nelson, A.W.; Chernukhin, I.; Carroll, J.S.; Li, Y.; Subramaniam, M.; Suman, V.J.; Negron, V.; Monroe, D.G.; et al. ERβ-Mediated Induction of Cystatins Results in Suppression of TGFβ Signaling and Inhibition of Triple-Negative Breast Cancer Metastasis. Proc. Natl. Acad. Sci. USA 2018, 115, E9580–E9589. [Google Scholar] [CrossRef]
- Neuwirt, H.; Bouchal, J.; Kharaishvili, G.; Ploner, C.; Jöhrer, K.; Pitterl, F.; Weber, A.; Klocker, H.; Eder, I.E. Cancer-Associated Fibroblasts Promote Prostate Tumor Growth and Progression through Upregulation of Cholesterol and Steroid Biosynthesis. Cell Commun. Signal. 2020, 18, 11. [Google Scholar] [CrossRef] [PubMed]
- Amin, F.; Fathi, F.; Reiner, Ž.; Banach, M.; Sahebkar, A. The Role of Statins in Lung Cancer. Arch. Med. Sci. 2021, 18, 141. [Google Scholar] [CrossRef]
- Feng, J.-L.; Dixon-Suen, S.C.; Jordan, S.J.; Webb, P.M. Statin Use and Survival among Women with Ovarian Cancer: An Australian National Data-Linkage Study. Br. J. Cancer 2021, 125, 766–771. [Google Scholar] [CrossRef]
- Ibrahim, A.B.; Zaki, H.F.; Ibrahim, W.W.; Omran, M.M.; Shouman, S.A. Evaluation of Tamoxifen and Simvastatin as the Combination Therapy for the Treatment of Hormonal Dependent Breast Cancer Cells. Toxicol. Rep. 2019, 6, 1114–1126. [Google Scholar] [CrossRef]
- Mayengbam, S.S.; Singh, A.; Pillai, A.D.; Bhat, M.K. Influence of Cholesterol on Cancer Progression and Therapy. Transl. Oncol. 2021, 14, 101043. [Google Scholar] [CrossRef]
- Braun, H.; Nash, R.; Tangpricha, V.; Brockman, J.; Ward, K.; Goodman, M. Cancer in Transgender People: Evidence and Methodological Considerations. Epidemiol. Rev. 2017, 39, 93–107. [Google Scholar] [CrossRef]
- De Blok, C.J.M.; Dreijerink, K.M.A.; Den Heijer, M. Cancer Risk in Transgender People. Endocrinol. Metab. Clin. N. Am. 2019, 48, 441–452. [Google Scholar] [CrossRef] [PubMed]
- De Blok, C.J.M.; Wiepjes, C.M.; Nota, N.M.; Van Engelen, K.; Adank, M.A.; Dreijerink, K.M.A.; Barbé, E.; Konings, I.R.H.M.; Den Heijer, M. Breast Cancer Risk in Transgender People Receiving Hormone Treatment: Nationwide Cohort Study in the Netherlands. BMJ 2019, 365, l1652. [Google Scholar] [CrossRef]
- Bertoncelli Tanaka, M.; Sahota, K.; Burn, J.; Falconer, A.; Winkler, M.; Ahmed, H.U.; Rashid, T.G.; The Gender Research Collaborative. Prostate Cancer in Transgender Women: What Does a Urologist Need to Know? BJU Int. 2022, 129, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.B.; McGlynn, K.A.; Devesa, S.S.; Freedman, N.D.; Anderson, W.F. Sex Disparities in Cancer Mortality and Survival. Cancer Epidemiol. Biomark. Prev. 2011, 20, 1629–1637. [Google Scholar] [CrossRef]
- Bertram, J.S.; Craig, A.W. Specific Induction of Bladder Cancer in Mice by Butyl-(4-Hydroxybutyl)-Nitrosamine and the Effects of Hormonal Modifications on the Sex Difference in Response. Eur. J. Cancer (1965) 1972, 8, 587–594. [Google Scholar] [CrossRef]
- Zhang, Y. Understanding the Gender Disparity in Bladder Cancer Risk: The Impact of Sex Hormones and Liver on Bladder Susceptibility to Carcinogens. J. Environ. Sci. Health Part C 2013, 31, 287–304. [Google Scholar] [CrossRef] [PubMed]
- Lucca, I.; Fajkovic, H.; Klatte, T. Sex Steroids and Gender Differences in Nonmuscle Invasive Bladder Cancer. Curr. Opin. Urol. 2014, 24, 500–505. [Google Scholar] [CrossRef]
- Levin, G.; Meyer, R. Association of Polycystic Ovary Syndrome with Pancreatic Cancer. JAMA Oncol. 2023. [Google Scholar] [CrossRef]
- Mravec, B.; Tibensky, M. Increased Cancer Risk in Polycystic Ovary Syndrome: An (Un)Sympathetic Connection? Med. Hypotheses 2020, 134, 109437. [Google Scholar] [CrossRef]
- Peeri, N.C.; Du, M. Association of Polycystic Ovary Syndrome with Pancreatic Cancer—Reply. JAMA Oncol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Peeri, N.C.; Landicino, M.V.; Saldia, C.A.; Kurtz, R.C.; Rolston, V.S.; Du, M. Association between Polycystic Ovary Syndrome and Risk of Pancreatic Cancer. JAMA Oncol. 2022, 8, 1845. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabbatino, E.; Tutino, V.; Licitra, F.; Di Donato, M.; Castoria, G.; Migliaccio, A.; Giovannelli, P. Role of the Androgen Receptor in Gender-Related Cancers. Endocrines 2023, 4, 407-426. https://doi.org/10.3390/endocrines4020031
Sabbatino E, Tutino V, Licitra F, Di Donato M, Castoria G, Migliaccio A, Giovannelli P. Role of the Androgen Receptor in Gender-Related Cancers. Endocrines. 2023; 4(2):407-426. https://doi.org/10.3390/endocrines4020031
Chicago/Turabian StyleSabbatino, Emilia, Viviana Tutino, Fabrizio Licitra, Marzia Di Donato, Gabriella Castoria, Antimo Migliaccio, and Pia Giovannelli. 2023. "Role of the Androgen Receptor in Gender-Related Cancers" Endocrines 4, no. 2: 407-426. https://doi.org/10.3390/endocrines4020031
APA StyleSabbatino, E., Tutino, V., Licitra, F., Di Donato, M., Castoria, G., Migliaccio, A., & Giovannelli, P. (2023). Role of the Androgen Receptor in Gender-Related Cancers. Endocrines, 4(2), 407-426. https://doi.org/10.3390/endocrines4020031