Abstract
The organic sulfur-containing compounds glucosinolates (GSLs) and the novel gasotransmitter H2S are known to have cardioprotective effects. This study investigated the antioxidant effects and H2S-releasing potential of three GSLs ((3E)-4-(methylsulfanyl)but-3-enyl GSL or glucoraphasatin, 4-hydroxybenzyl GSL or glucosinalbin, and (RS)-6-(methylsulfinyl)hexyl GSL or glucohesperin) in rat cardiac cells. It was found that all three GSLs had no effect on cardiac cell viability but were able to protect against H2O2-induced oxidative stress and cell death. NaHS, a H2S donor, also protected the cells from H2O2-stimulated oxidative stress and cell death. The GSLs alone or mixed with cysteine, N-acetylcysteine, glutathione, H2O2, iron and pyridoxal-5′-phosphate, or mouse liver lysates did not induce H2S release. The addition of GSLs also did not alter endogenous H2S levels in cardiac cells. H2O2 significantly induced cysteine oxidation in the cystathionine gamma-lyase (CSE) protein and inhibited the H2S production rate. In conclusion, this study found that the three tested GSLs protect cardiomyocytes from oxidative stress and cell death but independently of H2S signaling.
1. Introduction
Plants of the Brassicaceae family contain many natural products, including abundant glucosinolates (GSLs), which are a class of sulfur-containing molecules with known health benefits. GSLs are characterized by their core structural S-β-D-glucopyrano unit anomerically connected to an O-sulfated (Z)-thiohydroximate function and differ by the structure of their R chain [1,2]. Among the different GSLs, (RS)-6-(methylsulfinyl)hexyl GSL or glucohesperin (Gh), (3E)-4-(methylsulfanyl)but-3-enyl GSL or glucoraphasatin (Gr), and 4-hydroxybenzyl GSL or glucosinalbin (Gs) are very common [3,4]. GSLs serve as defense mechanisms when hydrolyzed into isothiocyanates (ITCs), thiocyanates, epithionitriles, and/or nitriles by the myrosinase enzyme (Figure 1). These compounds and their corresponding degradation products have been widely investigated for their antioxidant, antimicrobial, and anti-cancer capacities [5,6,7].
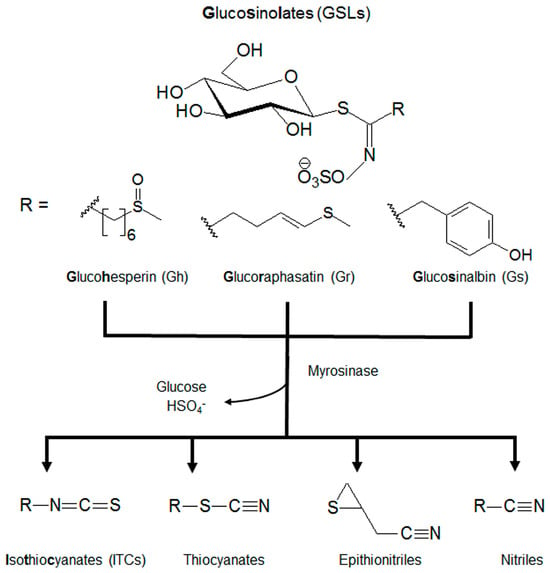
Figure 1.
The core structure and metabolism of glucosinolates (GSLs). GSLs are an S-β-D-glucopyrano unit anomerically connected to an O-sulfated (Z)-thiohydroximate function with an alkyl, arylalkyl, or indolyl side chain (R). GSLs can be further hydrolyzed by myrosinases to isothiocyanates (ITCs), thiocyanates, epithionitriles, and nitriles.
The gaseous molecule H2S is emerging as a novel regulator with a wide range of physical properties and physiological functions [8,9,10]. H2S can be generated via both enzymatic and non-enzymatic pathways. Enzymatically, cystathionine beta-synthase, cystathionine gamma-lyase (CSE), and 3-mercaptopyruvate sulfurtransferase, along with cysteine aminotransferase, use the cofactor pyridoxal 5′-phosphate (P5P) to catalyze cysteines and produce H2S [11]. Non-enzymatically, H2S can be released from sulfur-containing amino acids and natural sulfur products via chemical interaction with P5P, glutathione (GSH), and/or iron [12,13]. Other sulfur-containing molecules, i.e., diallyldisulfide from garlic, can release H2S in the bloodstream through interaction with GSH [12,14]. ITCs (the byproducts of GSLs) and glucoraphanin (a GSL) have been shown to release H2S in mammalian cells in a cysteine-dependent non-enzymatic reaction [15,16].
Cardiovascular diseases are the leading cause of death globally for both men and women, resulting in 17.9 million deaths each year. Commonly, cardiovascular diseases can be caused by disrupted heart structures and functions due to medical conditions (hypertension, diabetes, high lipid levels, etc.), lifestyle risk factors (unhealthy diet, physical inactivity, smoking, stress, etc.), and genetic factors [17,18]. Cardiomyocytes are extremely vulnerable to oxidative stress and have limited regenerative capacity [19,20]. A large amount of evidence has shown that both GSLs and H2S have cardio-protective effects. The consumption of GSL-enriched foods regularly, such as cruciferous vegetables, can prevent or reduce the onset of cardiovascular diseases [21,22]. H2S is widely shown to protect heart dysfunctions in various animal models, including myocardial ischemia/reperfusion injury, myocardial infarction, cardiac arrhythmia, cardiac hypertrophy, myocardial fibrosis, heart failure, etc. [23,24,25]. The cardioprotective roles of GSLs and H2S can be mostly attributed to their anti-inflammatory and antioxidant features [21,26]. So far, no study has explored the interplay between GSLs and H2S as it affects cardiac cell survival and oxidative stress regulation. This study hypothesized that GSLs act as H2S-releasing donors and protect from cardiomyocyte death induced by oxidative stress in a H2S-dependent manner.
This present study explored the protective roles of three GSLs (Gh, Gr, and Gs) and H2S in H2O2-induced oxidative stress and cell death in cultured rat cardiac cells. The effects of GSLs on CSE protein expression and endogenous H2S generation were also investigated. The non-enzymatic release of H2S from GSLs was further tested by mixing GSLs with or without cysteine, glutathione (GSH), N-acetyl-L-cysteine (NAC), H2O2, iron and pyridoxal 5′-phosphate (P5P). The potential of GSLs to act as a substrate for enzyme-catalyzed H2S generation was investigated with mouse liver lysates and cardiac cells. It was found that GSLs were able to protect cardiac cells from oxidative stress and cell death but did not release H2S under all the tested conditions.
2. Results
2.1. GSLs and H2S Protected H2O2-Induced Cell Death and Oxidative Stress
The effects of GSLs on the cell growth of rat cardiac cells (H9C2) were analyzed with an MTT assay. As shown in Figure 2, the three GSLs (Gh, Gr, and Gs) applied at the tested concentrations (1–100 μg/mL) did not alter cell viability in comparison to the control. The addition of H2O2 (800 μM) reduced cell viability by 52%; however, co-incubation with GSL (Gh, Gr, or Gs) at a dose higher than 5 μg/mL started to restore H2O2-induced cell death. GSLs at doses of 50 μg/mL and 100 μg/mL showed no significant difference compared with the dose of 10 μg/mL (Figure 2). Thus, a single dose of 10 μg/mL was chosen to further investigate the regulatory effects of GSLs on oxidative stress. It was found that the three GSLs (10 μg/mL) inhibited H2O2-induced cell death and oxidative stress (Figure 3A,B). The content of MDA, a lipid peroxidation marker, was dramatically higher in H2O2-treated cells but significantly reduced by coincubation with the three GSLs (Figure 3C). In addition, the supplement of H2S (30 μM) also markedly abolished the stimulatory effect of H2O2 (800 μM) on cell death, oxidative stress, and MDA content (Figure 3). The three tested GSLs seem to have similar activities.

Figure 2.
GSLs protected from H2O2-inhibited cell viability. After rat cardiac cells (H9C2) were incubated with or without 800 µM H2O2 in the presence of 1–100 µg/mL of Gh (A), Gr (B), or Gs (C) for 24 h, cell viability was detected by MTT. * p < 0.05 versus control; # p < 0.05 versus the cells with H2O2 only. The experiments were repeated 4 times.
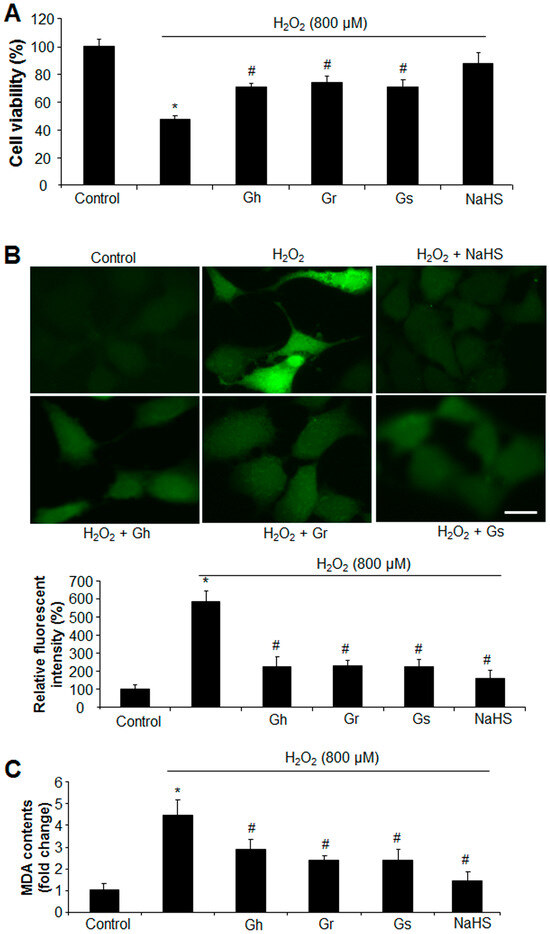
Figure 3.
GSLs and H2S protected cell death and oxidative stress induced by H2O2. Rat cardiac cells (H9C2) were exposed to 800 μM H2O2 in the presence of 10 μg/mL GSL or 30 μM H2S for 24 h. The cell viability was detected by MTT (A). ROS levels were examined by staining the cells with a fluorescent probe (H2DCFDA) (B). The fluorescent intensity was quantified with ImageJ software (Version 1.43) and normalized to the total cell number. The fluorescent intensity in control cells was considered 100%. Scale bar: 20 μm. Lipid peroxidation was also detected by measuring the content of MDA (C). * p < 0.05 versus control; # p < 0.05 versus H2O2 group. The experiments were repeated 4 times.
2.2. GSLs Had No Effect on CSE Protein Expression and H2S Generation in H9C2 Cells
As shown in Figure 4A, the incubation of the cells with H2O2 (800 μM) for 24 h did not affect CSE protein level, even in the presence of GSLs (10 μg/mL). However, H2O2 significantly reduced the H2S production rate by 60% in comparison to the control cells (Figure 4B). Coincubation with GSLs did not restore H2O2-inhibited H2S generation (Figure 4B). It was further found that H2O2 stimulated a higher level of cysteine sulfonate derived from total proteins and also CSE, pointing to possible protein post-translational modifications caused by H2O2 through cysteine oxidation (Figure 4C).
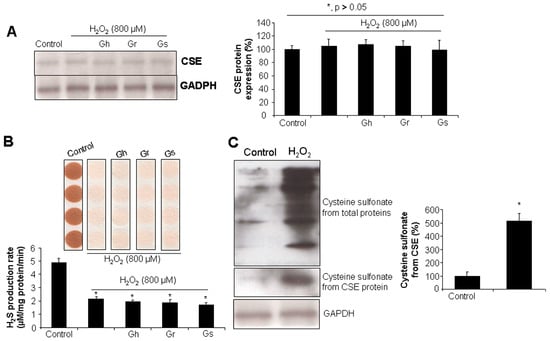
Figure 4.
The effects of H2O2 and/or GSLs on CSE protein expression and endogenous H2S generation. Rat cardiac cells (H9C2) were exposed to 800 μM H2O2 in the presence or absence of 10 μg/mL GSL for 24 h. The cells were then collected for CSE protein expression analysis by Western blot (A), H2S production rate analysis using lead acetate paper (B), and cysteine sulfonate production from CSE assessment using a Co-IP assay (C). In (A) * p > 0.05. In (B,C) * p < 0.05 versus control. The experiments were repeated 4 times.
2.3. No H2S Was Released from GSLs under Various Tested Conditions
By using a lead acetate paper assay, it was first found that the three GSLs alone (10 μg/mL) were not capable of releasing H2S at 37 °C for 2 h (Figure 5). Mixing GSLs with three sulfur-containing amino acids (cysteine, GSH, or NAC) also did not produce H2S. In addition, under oxidative conditions with H2O2, no H2S release was observed from GSLs. The additions of 100 μM FeCl2 and 1 mM P5P did not liberate H2S from GSLs. H2S release was clearly detected from NaHS, which was not altered by the presence of GSLs (Figure 5).
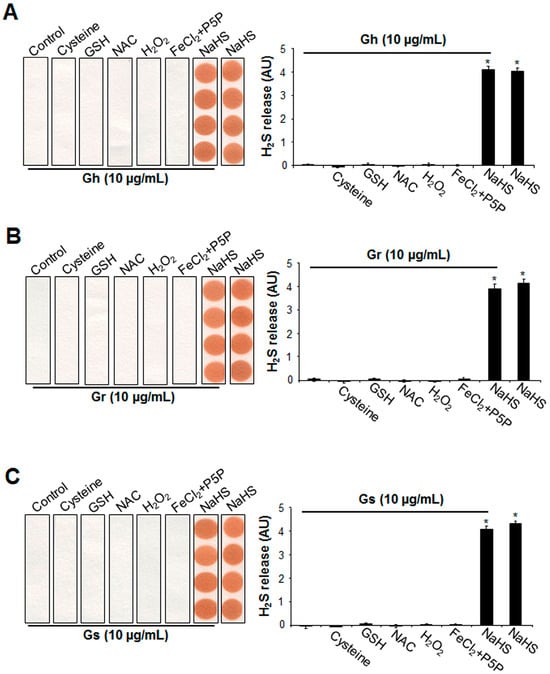
Figure 5.
The supplement of cysteine, GSH, NAC, H2O2, FeCl2 P5P, or NaHS did not induce H2S release from the 3 tested GSLs. Gh (A), Gr (B), or Gs (C) at 10 μg/mL was mixed with 10 mM cysteine, 10 mM GSH, 10 mM NAC, 100 μM H2O2, 100 μΜ FeCl2 and 1 mM P5P, or 100 μM NaHS, then incubated at 37 °C for 2 h. H2S released was trapped with lead acetate paper and quantified with ImageJ software (Version 1.43). * p < 0.05 versus all other groups. The experiments were repeated 4 times.
We then tested the potential of H2S release from GSLs when mixed with mouse liver lysates in the presence or absence of cysteine and P5P. H2S can be endogenously produced from the substrate cysteine via enzymatic pathways with P5P as a cofactor [11]. Significant H2S release was observed from mouse liver lysates when cysteine and P5P were present, which was not altered by the further addition of GSLs (Figure 6). No H2S release from the mixture of mouse liver lysates and GSLs only was detected, even in the presence of P5P (Figure 6). By using a H2S fluorescent probe WSP1, we further detected the real H2S level inside H9C2 cells. As shown in Figure 7A, the addition of GSLs had no effect on endogenous H2S levels compared with the control cells. Consistently, using a methylene blue method, the H2S level in the cell culture medium showed no difference among all groups (Figure 7B).
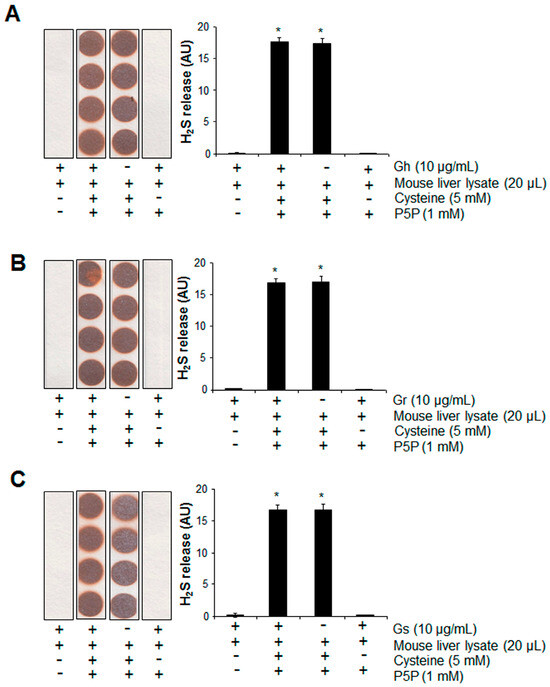
Figure 6.
The presence of mouse liver lysates did not induce H2S release from the 3 tested GSLs. Gh (A), Gr (B), or Gs (C) at 10 μg/mL was mixed with 20 μL mouse liver lysates in the presence of 5 mM cysteine and 1 mM P5P, then incubated at 37 °C for 2 h. H2S released was trapped with lead acetate paper and quantified with ImageJ software (Version 1.43). * p < 0.05 versus all other groups. “-/+” indicates the absence/presence of each component. The experiments were repeated 4 times.
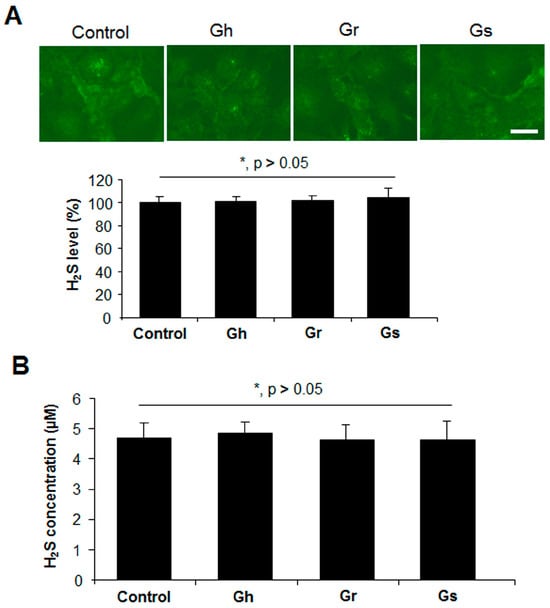
Figure 7.
GSLs did not alter endogenous H2S levels in H9C2 cells. Rat cardiac cells (H9C2) were incubated with 10 μg/mL of each GSL for 24 h. The H2S level in the cells and cell culture medium was determined with a H2S fluorescent probe WSP1 (A) and a methylene blue method (B), respectively. Scale bar: 20 μm. * p > 0.05. The experiments were repeated 4 times.
3. Discussion
GSLs occur naturally in high concentrations in plants of the Brassicaceae family and are known to have beneficial effects on humans at relatively low doses [1,2]. H2S is an important gasotransmitter with known cardioprotective effects [8,9,10]. H2S has been reported to be released from the enzymatic degradation products of some GSLs, i.e., ITCs, in physiological conditions [15,27,28]. This study explored the antioxidant effects of GSLs in cardiac cells and examined the possible utility of H2S signaling in mediating the functions of GSLs. It was found that GSLs could not release H2S under all the tested conditions, and GSLs protected cardiac cells from H2O2-induced oxidative stress and cell death in an H2S-independent manner.
GSL intake is reported to be associated with a lower risk of cardiovascular disease, and the administration of GSLs would induce the activities of certain Phase I enzymes and alter the generation of reactive oxygen species in vitro [2,5]. This present study showed that the three GSLs (Gh, Gs, and Gr) applied at a dose range of 1–100 µg/mL did not affect cell growth. We then investigated the protective effects of GSLs on H2O2-induced cell death in cardiac cells. The supplement of 10 µg/mL of each GSL showed a significant protective role against H2O2-induced cell death, oxidative stress, and MDA content. Similarly, we also observed that H2S was able to attenuate H2O2-stimulated cell death and oxidative stress. Given the findings that GSLs are a group of organosulfur compounds and are biologically inactive molecules, we then tested whether H2S would be liberated from GSLs and mediate the beneficial effects of GSLs in cardiac cells.
Differently from our hypothesis, no H2S release was detected from GSLs under various conditions. It has been reported that sulfur-containing amino acids could react with sulfur-containing natural products to release H2S non-enzymatically [12,14]. However, our studies show that the co-incubation of three sulfur-containing amino acids (cysteine, NAC, and GSH) with GSLs did not induce H2S release. In contrast to our findings, Lucarini et al. reported that the GSL glucoraphanin would release H2S in the presence of cysteine, thus protecting against chemotherapy-induced neuropathic pain by regulating Kv7 potassium channels [16]. Gambari et al. also showed that glucoraphanin increased intracellular H2S levels and stimulated osteogenic differentiation in human mesenchymal stromal cells [29]. The discrepancy may be due to the tested dose of GSLs, cell types, and H2S detection methods. In their studies, the concentration of glucoraphanin used for testing H2S release was more than 40 times higher than in this present study. They utilized an electrochemical approach with an Apollo-4000 amperometric detector and also a H2S-selective mini-electrode for the detection of the total sulfide. In contrast, we used lead acetate paper, which detects the free gaseous form of H2S aside from sulfide ion, hydrosulfide, polysulfide, and/or elemental sulfur, etc. [30]. H2S gas has higher lipid solubility and can be freely permeable through a cellular membrane. Compared with other forms of sulfide, the intracellular and intercellular transportations of H2S gas do not need any receptor or transporter, thus facilitating the gasotransmitter’s roles of free H2S gas inside the cells.
With the aid of P5P, iron could react with the sulfur-containing amino acid cysteine for H2S release [13]. However, our data with lead acetate paper do not support this notion since the addition of iron did not liberate H2S released from GSLs even in the presence of P5P. We also tested the possibility that the SH group from NaHS would react with the sulfur atom in GSLs to boost H2S release, but the result demonstrated that NaHS failed to generate H2S from the three tested GSLs. It was then hypothesized that GSLs may react with strong oxidant or reducing agents to liberate H2S. In contrast to this hypothesis, GSLs could not generate H2S in the presence of additional oxidants (H2O2) or reducing agents (NAC). In mammalian cells, the H2S-producing enzyme CSE uses cysteine as a substrate for producing H2S [11]. The CSE gene is highly expressed in liver tissues and contributes to a majority of H2S generation in our body [8,31]. By replacing cysteine with GSLs, we tried to observe whether the CSE enzyme can use GSLs as a substrate for generating H2S in mouse liver tissues. In opposition to the hypothesis, we did not detect any H2S release in mouse liver tissues when cysteine was replaced with GSLs. To determine the possibility of H2S release from GSLs when incubated with H9C2 cells for a time period of 24 h, we utilized two additional methods for detecting the real level of H2S inside the cells and also in the cell culture medium. WSP1 is a reactive disulfide-containing fluorescent probe used for the selective detection of H2S inside the cells [32]. No significant difference in fluorescent intensity was observed between control and GSL-incubated cells. The methylene blue method is an efficient and convenient assay for the determination of absolute H2S in biological samples, including blood and cell culture medium [33]. Similar to the results with WSP1, we did not see any change in the H2S level in the cell culture medium with or without GSL incubation. Different from the methods with lead acetate paper and WSP-1 probe, it should be noted that the presence of a high level of thiol or other reductants in the sample may interfere with the results of the methylene blue method [33]. Given all this evidence, it can be concluded that all of the conditions tested did not elicit any H2S release from GSLs. Future studies might explore the H2S-releasing potential of GSLs under other conditions, i.e., in the presence of GSL-metabolizing enzymes. In plants, GSLs can be further degraded by myrosinase via the hydrolytic cleavage of the GSLs for generating ITCs, which have been widely shown to release H2S under physiological conditions [15].
In this present study, we observed that H2O2 did not alter CSE protein expression in cardiac cells but inhibited endogenous H2S generation, suggesting that H2O2 may alter CSE activity via the post-translational modification of the CSE protein. This was supported by the data showing that H2O2 induced more cysteine oxidation in total proteins and CSE. Cysteine residues are essential for maintaining protein structure, stability, and functions. The free thiol group in cysteine can be easily oxidized to sulfenic, sulfinic, and sulfonic acids, thus changing the activities of enzymes [34]. In cultured smooth muscle cells, H2O2 incubation did not change the mRNA and protein levels of CSE but was able to promote the activity of CSE and increase H2S production [35]. However, in another study with human umbilical vein endothelial cells, H2O2 suppressed H2S generation by directly reducing CSE protein expression [36]. In all these studies, H2S protected the cells from H2O2-induced cell death [35,36]. This evidence suggests that, in response to oxidative stress, the H2S signal may be differently regulated to provide cardioprotection in a cell-dependent-type manner.
In summary, this study found that both GSLs and H2S protected cardiomyocytes from oxidative stress and cell death, while GSLs could not release H2S under any of the tested conditions. This study suggests that the antioxidant properties of GSLs in cardiomyocytes are independent of H2S signaling. Future studies should look at the pathway/mechanism by which GSLs generate their antioxidant effects and validate the cardioprotective functions of GSLs with human trials.
4. Materials and Methods
4.1. Isolation of the 3 GSLs (Gh, Gr, and Gs)
The 3 authenticated GSLs (Gh, Gr, and Gs) tested in this study were prepared as previously reported [37]. In brief, (RS)-6-(methylsulfinyl)hexyl GSL (Gh) was isolated from Dithyrea wislizenii Engelm. fruits. Fresh nine-day-old Cherry Belle radish seedlings were used to isolate (3E)-4-(methylsulfanyl)but-3-enyl (Gr). 4-hydroxybenzyl GSL (Gs) was isolated from Lepidium densiflorum Schrad. seeds. All these GSLs were characterized using mass spectrometry (MS) and nuclear magnetic resonance (NMR) spectroscopy. The electrospray mass spectra were recorded on a LC-MS instrument (Agilent Technologies HP 1100, New Castle, DE, USA) with a Chemstation data system LC–MSD B.03.01 at Laurentian University (Sudbury, ON, Canada). NMR spectra were recorded on a Bruker DRX 500 spectrometer (Milton, ON, Canada) at 500 MHz (1H) and 125 MHz (13C) at Laurentian University (Sudbury, ON, Canada).
4.2. Cell Culture
Rat cardiomyocytes (H9C2, ATCC, Manassas, VA, USA, CRL-1446TM) were cultured with Dulbecco’s modified eagle’s medium (Sigma-Aldrich, St. Louis, MO, USA) with 10% heat-inactivated fetal bovine serum, 100 U/mL penicillin and 100 mg/mL streptomycin at 37 °C in a humidified atmosphere of 5% CO2. Most of the experiments were performed when the cells reached 70–80% confluence unless otherwise indicated.
4.3. Animal Care and Tissue Collection
Male mice with a genetic background C57BL/6 × 129/Sv were fed a standard rodent feed with free access to water [8]. The liver tissues were collected from 8- to 10-week-old male mice and stored at −80 °C until they were used for experimentation. The animal experiments were conducted in compliance with the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH Publication No. 85-23, revised 1996) and approved by the Animal Care Committees of Laurentian University, Canada.
4.4. Cell Viability
Cell viabilities were assessed based on the conversion of tetrazolium salt 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) to formazan. Briefly, cells were plated at equal numbers onto each well of 96-well plates to form 70–80% confluence. The cells were then treated with H2O2, GSLs, and/or NaHS in 96-well plates for 24 h. Next, the cells were incubated with MTT reagent (5 mg/mL) for an additional 4 h of incubation at 37 °C. Subsequently, the formazan salts were solubilized by adding dimethylsulfoxide (Thermo Fisher Scientific, Toronto, ON, Canada) for 5 min. The FLUOstar OPTIMA microplate reader (BMG Labtech Inc., Ortenberg, Germany) was used to read the absorbance at 570 nm. The control cells without any treatment were considered 100% viable.
4.5. Measurement of H2S Production
A lead acetate paper was first used to analyze the H2S generated by the 3 GSLs [30]. Briefly, 100 µL of PBS solution containing the tested GSLs, cell/tissue lysates, L-cysteine, NAC, GSH, P5P, FeCl2, H2O2, and/or NaHS was added to 96-well plates, and then lead acetate paper (Sigma-Aldrich) was immediately placed over the wells. The reaction was incubated at 37 °C for 2 h in the dark. The intensity of the darkening on the lead acetate paper was quantified with ImageJ software (Version 1.43), and the amount of H2S release was expressed as an arbitrary unit (AU) or normalized to protein concentration and expressed as μM/mg protein/min.
The endogenous H2S level in cardiac cells was also detected using a H2S fluorescent probe WSP1 from Cayman Chemical (Ann Arbor, MI, USA) [32]. Briefly, after various treatments, the cells were washed once with PBS and then incubated with WSP1 working solution (10 μg/mL) in PBS for 30 min at 37 °C. Afterward, the cells were washed with PBS three times, and the fluorescence signal was immediately observed with an EVOS M5000 fluorescent microscope (Thermo Fisher Scientific). The fluorescent intensity was analyzed using ImageJ 1.43 software.
A methylene blue method was further used to measure H2S generation from cardiac cells [33]. Briefly, 100 μL of culture media from the control cells or GSLs-treated cells were added to a 1.5 mL Eppendorf tube containing zinc acetate (1% w/v, 300 μL) to trap H2S. After 5 min, the reaction was stopped by adding 200 μL of N,N-dimethyl-p-phenylenediamine sulfate (20 μM in 7.2 M HCl), and 200 μL of FeCl3 (30 mM in 1.2 M HCl). After the mixture was kept in the dark for 20 min, 100 μL of trichloroacetic acid (10% w/v) was added for 30 min to precipitate any protein that might be present in the culture media. Subsequently, the mixture was centrifuged at 15,000× g for 15 min. The absorbance at 670 nm for the supernatants was then read with a FLUOstar OPTIMA microplate reader (BMG Labtech Inc.). The H2S concentration in the culture media was calculated against the calibration curve of standard NaHS.
4.6. Western Blotting
After various treatments, the cells were collected and lysed with a radioimmunoprecipitation assay buffer supplemented with a protease inhibitor cocktail (Sigma-Aldrich) for protein extraction. A Pierce™ BCA protein assay reagent (Thermo Fisher Scientific) was used to determine the concentration of all protein samples. Equal amounts of proteins (90 μg) were separated in a 10% SDS-PAGE gel with the Mini-Protean II system for 90 min at 120 V. The protein was then transferred to a PVDF membrane (Pall Corporation, Pensacola, FL, USA), which was then blocked in PBS with 0.1% Tween 20 containing 3% skim milk at 4 °C overnight. The expression of target proteins was determined using appropriate primary antibodies and peroxidase-conjugated secondary antibodies (Sigma-Aldrich, 1:5000) followed by visualization using an ECL solution (Bio-Rad Laboratories, Mississauga, ON, Canada). After incubation with the antibodies, the membranes were washed in blocking solution 3 times for 5 min each. The antibodies were diluted as follows: CSE (1:1000, Abnova, Taipei, Taiwan), cysteine sulfonate (1:1000, Abcam, Toronto, ON, Canada), and GAPDH (1:500, Sigma-Aldrich). The densitometry of the CSE protein was analyzed with ImageJ software (Version 1.43) by normalizing to the intensity of GAPDH.
4.7. Detection of Oxidative Stress
Oxidative stress was monitored in H9C2 cells by staining the cells with 2,7-dichlorodihydrofluorescein diacetate (H2DCFDA) dye (Thermo Fisher Scientific). Briefly, after various treatments, the cells were washed once with PBS and then the H2DCFDA dye (1 μM) was added to the cells and incubated in the dark for 30 min, followed by visualization with an EVOS M5000 fluorescent microscope (Thermo Fisher Scientific). The fluorescent intensity was analyzed using ImageJ 1.43 software and normalized to cell number.
4.8. Co-IP Assay
Oxidized cysteines in the CSE protein were detected with a Co-IP assay, as previously described [34]. Briefly, the proteins (120 µg) were first incubated with 1 μg of anti-cysteine sulfonate antibody (Abcam) overnight at 4 °C, followed by incubation with protein A/G-agarose beads (Sigma-Aldrich) for 2 h at 4 °C. The beads were washed three times with lysis buffer, and the bound proteins were then analyzed by Western blotting with an anti-CSE antibody. The total proteins that had not undergone the Co-IP procedure were assessed for the detection of GAPDH expression with anti-GAPDH antibody. The cysteine sulfonate from the CSE protein was normalized to the GAPDH protein level.
4.9. Malondialdehyde (MDA) Content
Lipid peroxidation was determined by measuring the content of MDA with an assay kit from Abcam (Cambridge, MA, USA) [38]. Briefly, after various treatments, the cells were lysed in sodium phosphate buffer (20 mM, pH 3.0) containing 0.5% Triton X-100. A volume of 25 μL lysate was mixed with 10 μL MDA color reagent solution in a 96-well plate and incubated at room temperature for 30 min. Afterward, 40 μL of reaction solution was added to the mixture for an additional 30 min. The absorbance was recorded at 600 nm using a FLUOstar OPTIMA microplate spectrophotometer (BMG Labtech Inc.). MDA levels were calculated using a standard curve, normalized to the total protein amounts, and then expressed as fold change from the control.
4.10. Statistical Analysis
The data are presented as means ± SEM, representing at least 4 independent experiments. A statistical analysis was performed using a two-tailed Student’s t-test or a one-tailed ANOVA. A p-value < 0.05 was considered statistically significant.
Author Contributions
F.H., B.A., S.M. and G.Y. conceived this study. F.H., B.A. and G.Y. performed the experiments and analyzed the data. S.M. provided the three tested GSLs. F.H., S.M. and G.Y. wrote and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by a discovery grant from the Natural Sciences and Engineering Research Council of Canada (RGPIN/5369-2022) and a grant-in-aid from the Heart and Stroke Foundation of Canada (G-18-0022098).
Institutional Review Board Statement
The animal study protocol was approved by the Animal Care Committees of Laurentian University (protocol code: 6013832; date of approval: 9 April 2020).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Miękus, N.; Marszałek, K.; Podlacha, M.; Iqbal, A.; Puchalski, C.; Świergiel, A.H. Health benefits of plant-derived sulfur compounds, glucosinolates, and organosulfur compounds. Molecules 2020, 25, 3804. [Google Scholar] [CrossRef] [PubMed]
- Kamal, R.M.; Abdull Razis, A.F.; Mohd Sukri, N.S.; Perimal, E.K.; Ahmad, H.; Rollin, P.; Djedaini-Pilard, F.; Mazzon, E.; Rigaud, S. Beneficial health effects of glucosinolates-derived isothiocyanates on cardiovascular and neurodegenerative diseases. Molecules 2022, 27, 624. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Subramanian, P.; Hahn, B.S. Glucosinolate diversity analysis in Choy Sum (Brassica rapa subsp. chinensis var. parachinensis) Germplasms for functional food breeding. Foods 2023, 12, 2400. [Google Scholar] [PubMed]
- Dinkova-Kostova, A.T.; Kostov, R.V. Glucosinolates and isothiocyanates in health and disease. Trends Mol. Med. 2012, 18, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Armah, C.N.; Derdemezis, C.; Traka, M.H.; Dainty, J.R.; Doleman, J.F.; Saha, S.; Leung, W.; Potter, J.F.; Lovegrove, J.A.; Mithen, R.F. Diet rich in high glucoraphanin broccoli reduces plasma LDL cholesterol: Evidence from randomised controlled trials. Mol. Nutr. Food Res. 2015, 59, 918–926. [Google Scholar] [CrossRef] [PubMed]
- García-Ibañez, P.; Yepes-Molina, L.; Ruiz-Alcaraz, A.J.; Martínez-Esparza, M.; Moreno, D.A.; Carvajal, M.; García-Peñarrubia, P. Brassica bioactives could ameliorate the chronic inflammatory condition of endometriosis. Int. J. Mol. Sci. 2020, 21, 9397. [Google Scholar] [CrossRef] [PubMed]
- Galuppo, M.; Iori, R.; De Nicola, G.R.; Bramanti, P.; Mazzon, E. Anti-inflammatory and anti-apoptotic effects of (RS)-glucoraphanin bioactivated with myrosinase in murine sub-acute and acute MPTP-induced Parkinson’s disease. Bioorg. Med. Chem. 2013, 21, 5532–5547. [Google Scholar] [CrossRef]
- Yang, G.; Wu, L.; Jiang, B.; Yang, W.; Qi, J.; Cao, K.; Meng, Q.; Mustafa, A.K.; Mu, W.; Zhang, S.; et al. H2S as a physiologic vasorelaxant: Hypertension in mice with deletion of cystathionine gamma-lyase. Science 2008, 322, 587–590. [Google Scholar] [CrossRef]
- Powell, C.R.; Dillon, K.M.; Matson, J.B. A review of hydrogen sulfide (H2S) donors: Chemistry and potential therapeutic applications. Biochem. Pharmacol. 2018, 149, 110–123. [Google Scholar] [CrossRef]
- Nishida, M.; Sawa, T.; Kitajima, N.; Ono, K.; Inoue, H.; Ihara, H.; Motohashi, H.; Yamamoto, M.; Suematsu, M.; Kurose, H.; et al. Hydrogen sulfide anion regulates redox signaling via electrophile sulfhydration. Nat. Chem. Biol. 2012, 8, 714–724. [Google Scholar]
- Sen, U.; Sathnur, P.B.; Kundu, S.; Givvimani, S.; Coley, D.M.; Mishra, P.K.; Qipshidze, N.; Tyagi, N.; Metreveli, N.; Tyagi, S.C. Increased endogenous H2S generation by CBS, CSE, and 3MST gene therapy improves ex vivo renovascular relaxation in hyperhomocysteinemia. Am. J. Physiol. Cell Physiol. 2012, 303, C41–C51. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Kurboniyon, M.S.; Zou, Y.; Luo, K.; Fang, S.; Xia, P.; Ning, S.; Zhang, L.; Wang, C. GSH-triggered/photothermal-enhanced H2S signaling molecule release for gas therapy. Pharmaceutics 2023, 15, 2443. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Minkler, P.; Grove, D.; Wang, R.; Willard, B.; Dweik, R.; Hine, C. Non-enzymatic hydrogen sulfide production from cysteine in blood is catalyzed by iron and vitamin B6. Commun. Biol. 2019, 2, 194. [Google Scholar] [CrossRef] [PubMed]
- Predmore, B.L.; Kondo, K.; Bhushan, S.; Zlatopolsky, M.A.; King, A.L.; Aragon, J.P.; Grinsfelder, D.B.; Condit, M.E.; Lefer, D.J. The polysulfide diallyl trisulfide protects the ischemic myocardium by preservation of endogenous hydrogen sulfide and increasing nitric oxide bioavailability. Am. J. Physiol. Heart Circ. Physiol. 2012, 302, H2410–H2418. [Google Scholar] [CrossRef] [PubMed]
- Martelli, A.; Testai, L.; Citi, V.; Marino, A.; Bellagambi, F.; Ghimenti, G.; Breschi, M.; Calderone, V. Pharmacological characterization of the vascular effects of aryl isothiocyanates: Is hydrogen sulfide the real player? Vasc. Pharmacol. 2014, 60, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Lucarini, E.; Micheli, L.; Trallori, E.; Citi, V.; Martelli, A.; Testai, L.; De Nicola, G.R.; Iori, R.; Calderone, V.; Ghelardini, C.; et al. Effect of glucoraphanin and sulforaphane against chemotherapy-induced neuropathic pain: Kv7 potassium channels modulation by H2S release in vivo. Phytother. Res. 2018, 32, 2226–2234. [Google Scholar] [CrossRef] [PubMed]
- Mensah, G.A.; Roth, G.A.; Fuster, V. The global burden of cardiovascular diseases and risk factors: 2020 and beyond. J. Am. Coll. Cardiol. 2019, 74, 2529–2532. [Google Scholar] [CrossRef]
- Wojtasińska, A.; Kućmierz, J.; Tokarek, J.; Dybiec, J.; Rodzeń, A.; Młynarska, E.; Rysz, J.; Franczyk, B. New insights into cardiovascular diseases treatment based on molecular targets. Int. J. Mol. Sci. 2023, 24, 16735. [Google Scholar] [CrossRef]
- Stojanovic, D.; Stojanovic, M.; Milenkovic, J.; Velickov, A.; Ignjatovic, A.; Milojkovic, M. The multi-faceted nature of renalase for mitochondrial dysfunction improvement in cardiac disease. Cells 2023, 12, 1607. [Google Scholar] [CrossRef]
- Su, X.; Zhou, M.; Li, Y.; Zhang, J.; An, N.; Yang, F.; Zhang, G.; Yuan, C.; Chen, H.; Wu, H.; et al. Protective effects of natural products against myocardial ischemia/reperfusion: Mitochondria-targeted therapeutics. Biomed. Pharmacother. 2022, 149, 112893. [Google Scholar] [CrossRef]
- Connolly, E.L.; Sim, M.; Travica, N.; Marx, W.; Beasy, G.; Lynch, G.S.; Bondonno, C.P.; Lewis, J.R.; Hodgson, J.M.; Blekkenhorst, L.C. Glucosinolates from cruciferous vegetables and their potential role in chronic disease: Investigating the preclinical and clinical evidence. Front. Pharmacol. 2021, 12, 767975. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Liu, G.; Zong, G.; Sampson, L.; Hu, F.B.; Willett, W.C.; Rimm, E.B.; Manson, J.E.; Rexrode, K.M.; Sun, Q. Intake of glucosinolates and risk of coronary heart disease in three large prospective cohorts of US men and women. Clin. Epidemiol. 2018, 10, 749–762. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Tan, B.; Sun, Y.; Hu, Q. Cystathionine γ lyase S-sulfhydrates Drp1 to ameliorate heart dysfunction. Redox. Biol. 2022, 58, 102519. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zhang, L.; Lu, B.; Wen, J.; Wang, M.; Zhang, S.; Li, Q.; Shu, F.; Lu, F.; Liu, N.; et al. Hydrogen sulphide reduced the accumulation of lipid droplets in cardiac tissues of db/db mice via Hrd1 S-sulfhydration. J. Cell. Mol. Med. 2021, 25, 9154–9167. [Google Scholar] [CrossRef] [PubMed]
- Citi, V.; Piragine, E.; Testai, L.; Breschi, M.C.; Calderone, V.; Martelli, A. The role of hydrogen sulfide and H2S-donors in myocardial protection against ischemia/reperfusion injury. Curr. Med. Chem. 2018, 25, 4380–4401. [Google Scholar] [CrossRef] [PubMed]
- Bechelli, C.; Macabrey, D.; Deglise, S.; Allagnat, F. Clinical potential of hydrogen sulfide in peripheral arterial disease. Int. J. Mol. Sci. 2023, 24, 9955. [Google Scholar] [CrossRef] [PubMed]
- Micheli, L.; Mitidieri, E.; Turnaturi, C.; Vanacore, D.; Ciampi, C.; Lucarini, E.; Cirino, G.; Ghelardini, C.; Sorrentino, R.; Di Cesare Mannelli, L.; et al. Beneficial effect of H2S-releasing molecules in an in vitro model of sarcopenia: Relevance of glucoraphanin. Int. J. Mol. Sci. 2022, 23, 5955. [Google Scholar] [CrossRef]
- Lin, Y.; Yang, X.; Lu, Y.; Liang, D.; Huang, D. Isothiocyanates as H2S donors triggered by cysteine: Reaction mechanism and structure and activity relationship. Org. Lett. 2019, 21, 5977–5980. [Google Scholar] [CrossRef]
- Gambari, L.; Barone, M.; Amore, E.; Grigolo, B.; Filardo, G.; Iori, R.; Citi, V.; Calderone, V.; Grassi, F. Glucoraphanin increases intracellular hydrogen sulfide (H2S) levels and stimulates osteogenic differentiation in human mesenchymal stromal cell. Nutrients 2022, 14, 435. [Google Scholar] [CrossRef]
- Ahn, B.K.; Ahn, Y.J.; Lee, Y.J.; Lee, Y.H.; Lee, G.J. Simple and sensitive detection of bacterial hydrogen sulfide production using a paper-based colorimetric assay. Sensors 2022, 22, 5928. [Google Scholar] [CrossRef]
- Jeddi, S.; Gholami, H.; Gheibi, S.; Kashfi, K.; Ghasemi, A. Altered gene expression of hydrogen sulfide-producing enzymes in the liver and muscles tissues of hyperthyroid rats. J. Cell. Physiol. 2019, 234, 17937–17945. [Google Scholar] [CrossRef] [PubMed]
- Martelli, A.; Citi, V.; Calderone, V. Vascular Effects of H2S-donors: Fluorimetric detection of H2S generation and ion channel activation in human aortic smooth muscle cells. Methods Mol. Biol. 2019, 2007, 79–87. [Google Scholar] [PubMed]
- Bibli, S.I.; Luck, B.; Zukunft, S.; Wittig, J.; Chen, W.; Xian, M.; Papapetropoulos, A.; Hu, J.; Fleming, I. A selective and sensitive method for quantification of endogenous polysulfide production in biological samples. Redox Biol. 2018, 18, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Davies, M.J.; Krämer, A.C.; Miotto, G.; Zaccarin, M.; Zhang, H.; Ursini, F. Protein cysteine oxidation in redox signaling: Caveats on sulfenic acid detection and quantification. Arch. Biochem. Biophys. 2017, 617, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jia, G.; Li, H.; Yan, S.; Qian, J.; Guo, X.; Li, G.; Qi, H.; Zhu, Z.; Wu, Y.; et al. H2O2-mediated oxidative stress enhances cystathionine γ-lyase-derived H2S synthesis via a sulfenic acid intermediate. Antioxidants 2021, 10, 1488. [Google Scholar] [CrossRef]
- Wen, Y.D.; Wang, H.; Kho, S.H.; Rinkiko, S.; Sheng, X.; Shen, H.M.; Zhu, Y.Z. Hydrogen sulfide protects HUVECs against hydrogen peroxide induced mitochondrial dysfunction and oxidative stress. PLoS ONE 2013, 8, e53147. [Google Scholar] [CrossRef]
- Montaut, S.; Benson, H.J.; Kay, M.; Guido, B.S.; Mahboob, S.S.; Chénier, J.; Gasparetto, J.L.; Joly, H.A. Probing the free-radical scavenging activity of the extract, the major glucosinolate and isothiocyanate of Eruca sativa Mill and Lepidium densiflorum Schrad. seeds. J. Food Compos. Anal. 2017, 61, 52–58. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).